
Anti-Phototoxicity Effect of Phenolic Compounds from Acetone Extract of Entada phaseoloides Leaves via Activation of COX-2 and iNOS in Human Epidermal Keratinocytes


 ,
,
Abstract
:1. Introduction
2. Material and Methods
2.1. Chemicals and Reagents
2.2. Identification of the Isolated Compounds
2.3. Extraction and Isolation
2.4. High-Performance Liquid Chromatography (HPLC) Analysis
2.5. Analysis of the Acetone Extract by LC-DAD-MS/qTOF
2.6. Cell Culture
2.7. Cell Viability Measurement by MTT Assay
2.8. UVB-Irradiation
2.9. Wound Healing Assay
2.10. Real-Time Quantitative PCR
2.11. Statistical Analysis
3. Results and Discussion
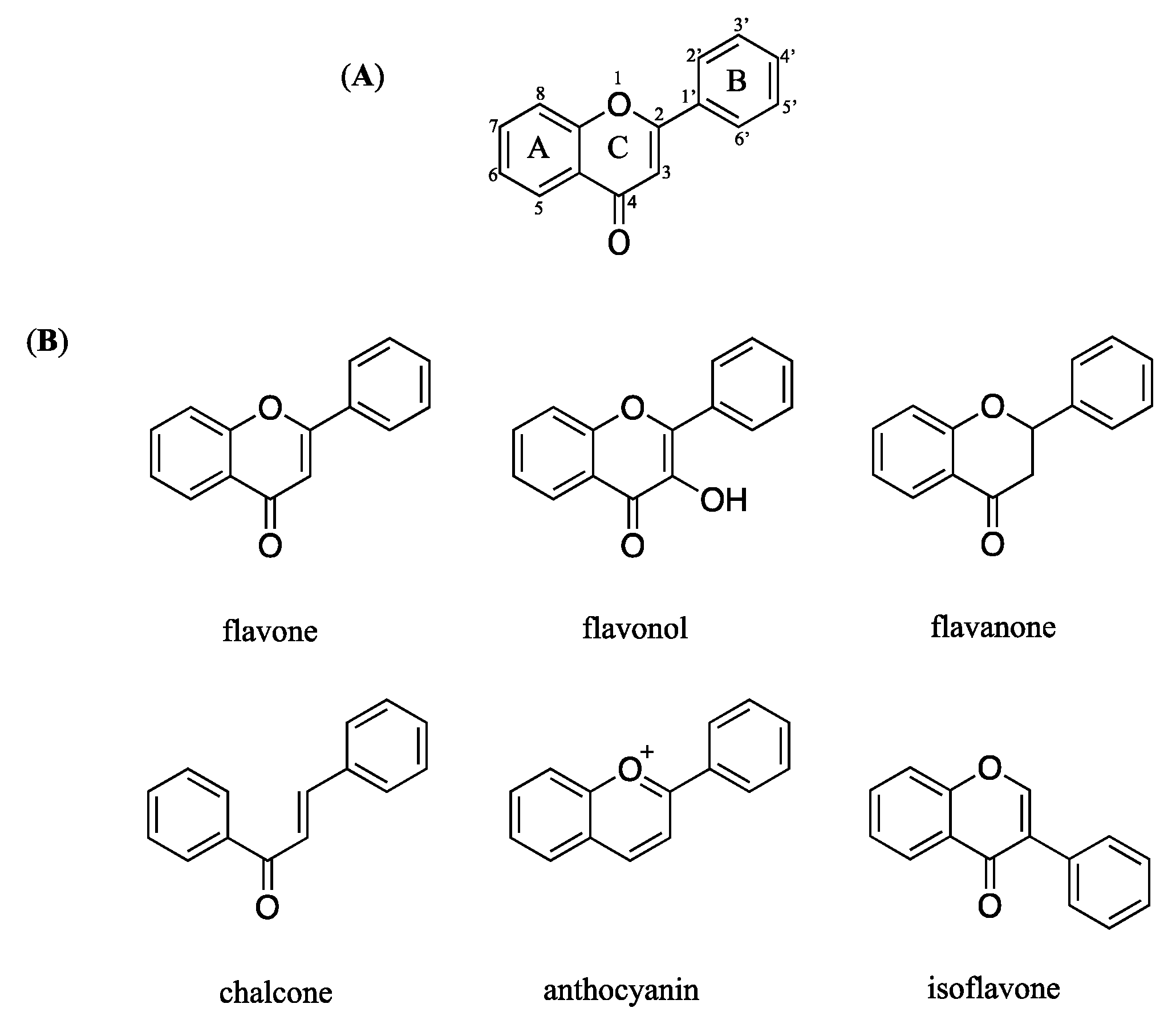
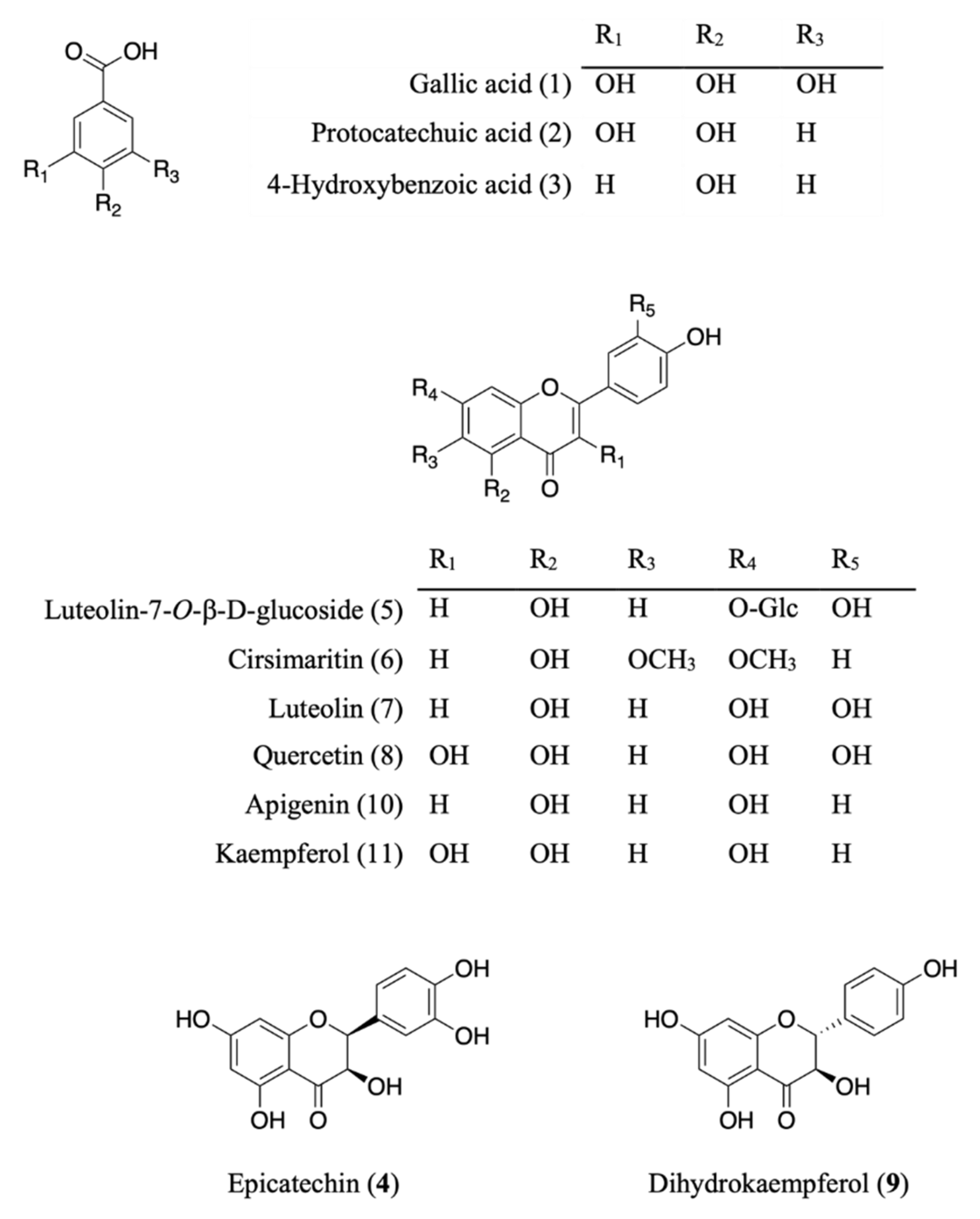
3.1. Identification of the Isolated Compounds
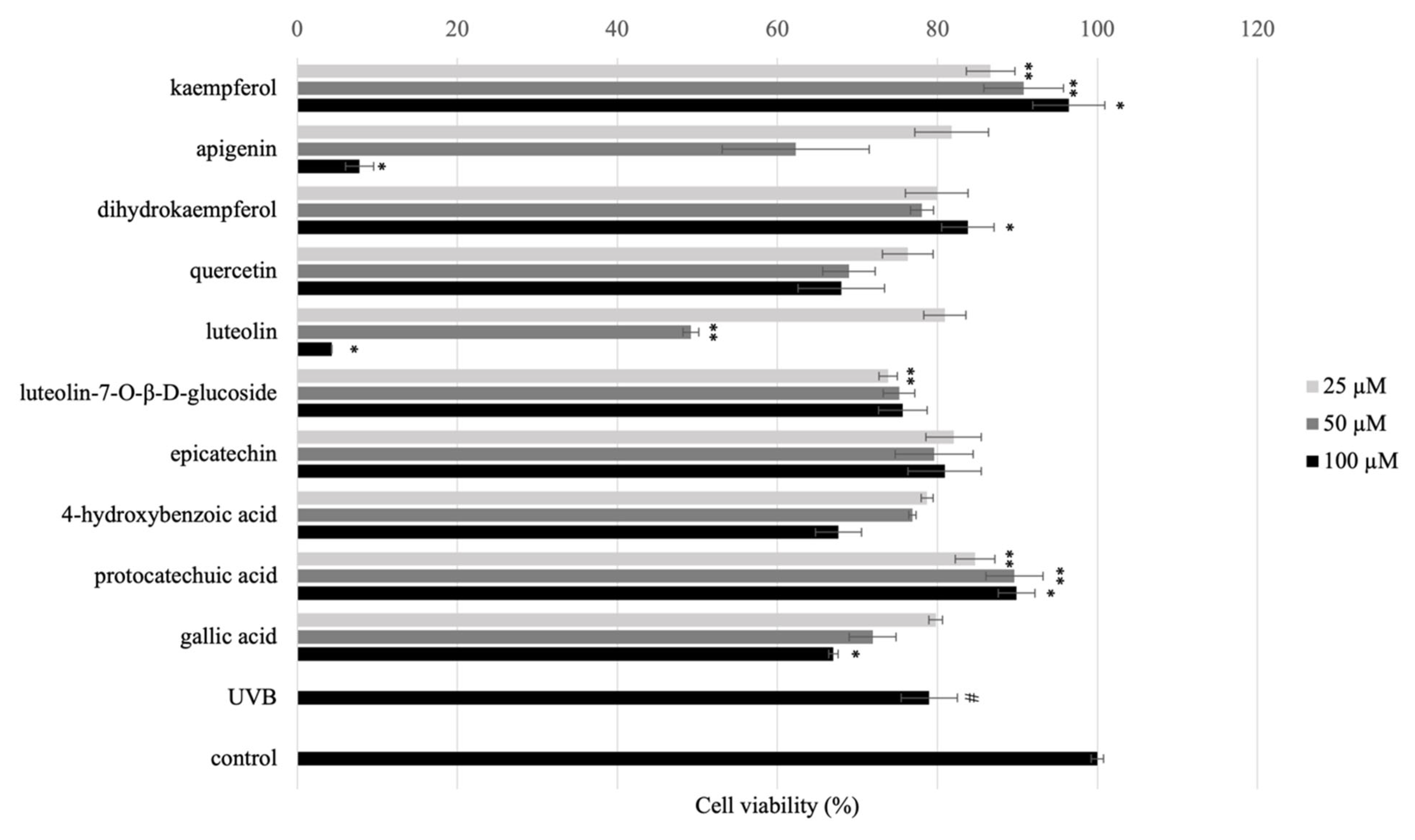
3.2. Cytotoxicity of Isolated/Standard Compounds
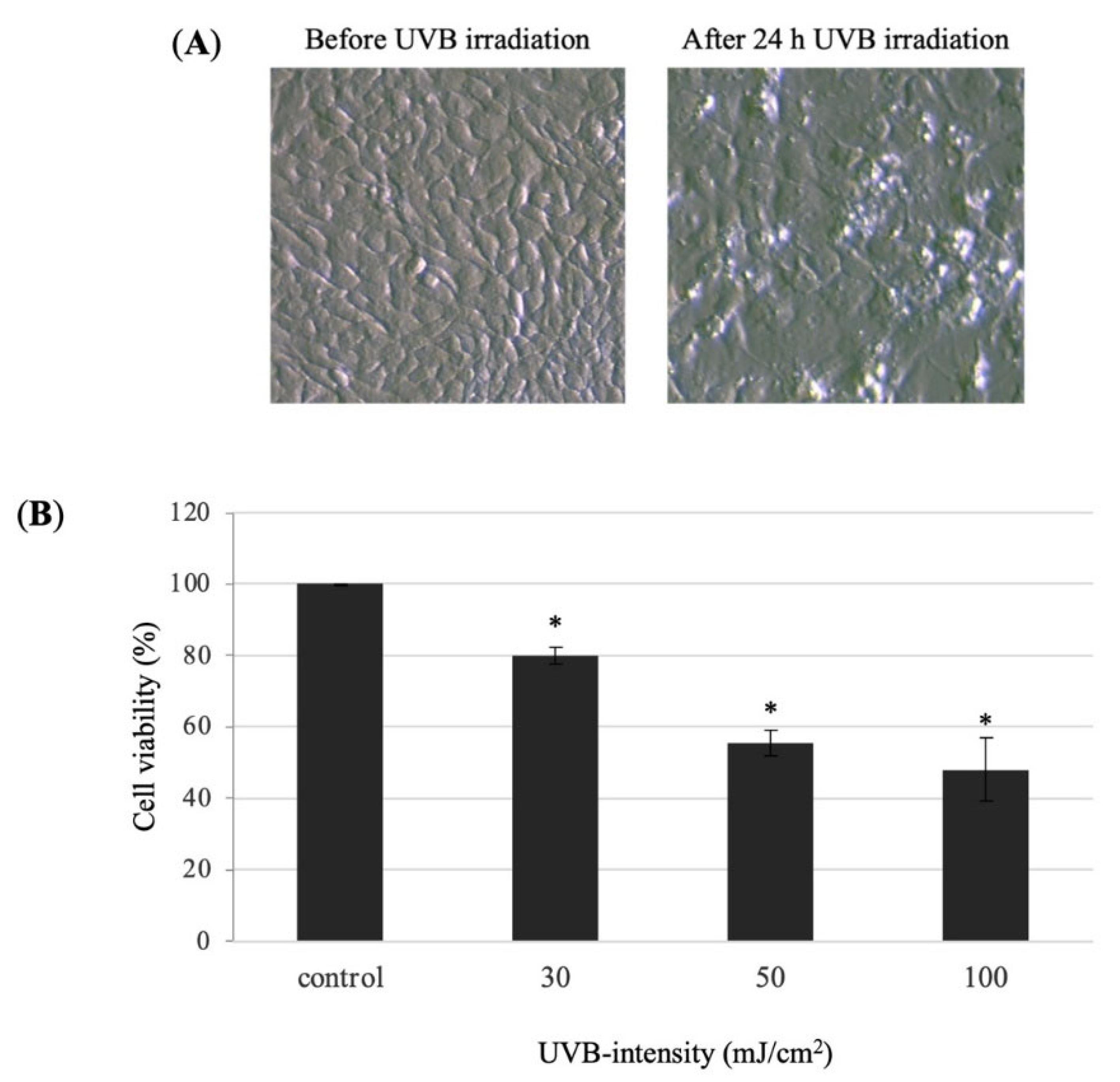
3.3. UVB-Induced HaCaT Cell Damage
3.4. Protective Effects of Isolated and/or Standard Compounds against UVB-Irradiation
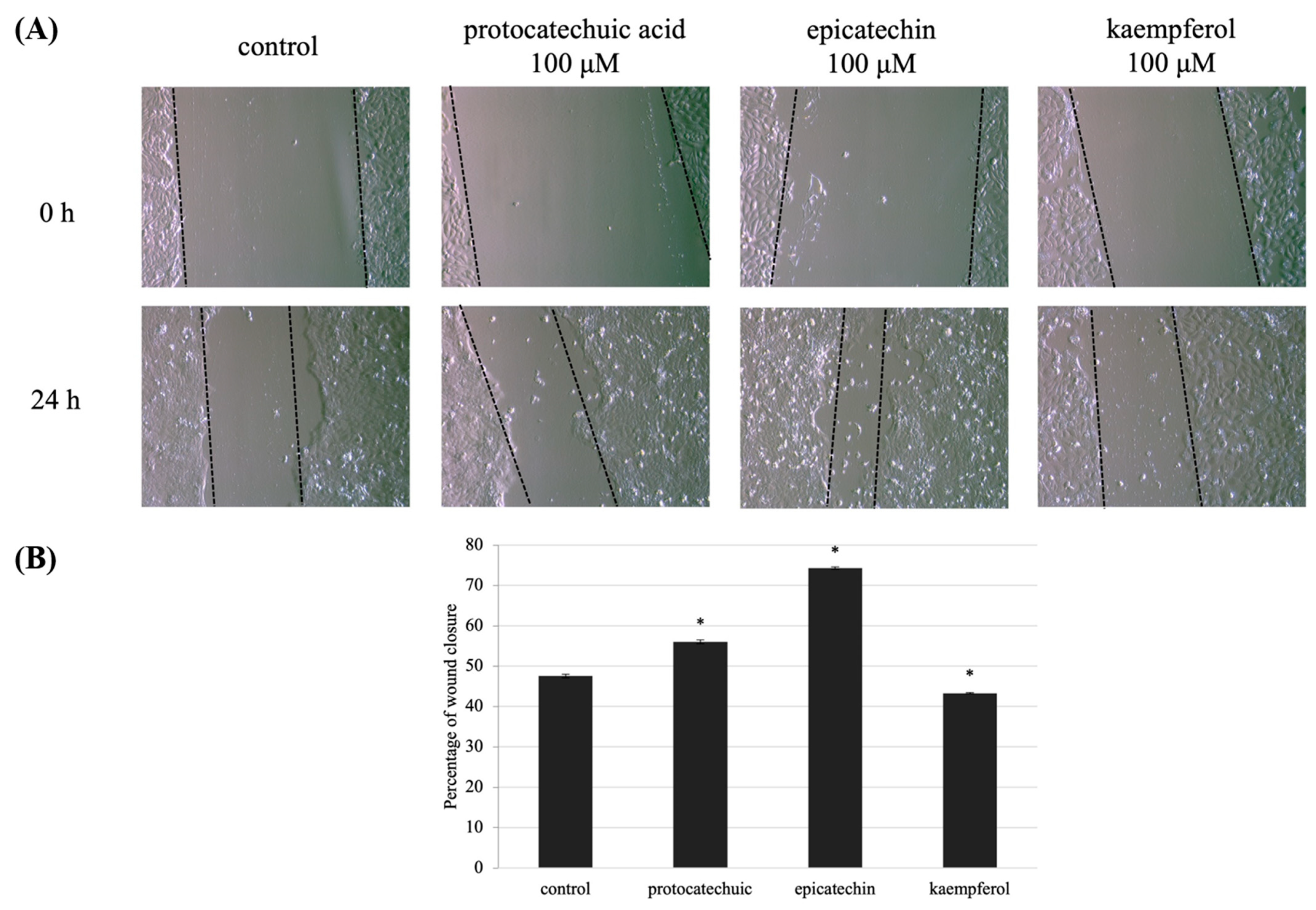
3.5. Protocatechuic Acid, Epicatechin, and Kaempferol Rescue UVB-Induced Migration Defect in HaCaT Cells
3.6. Effect of Protocatechuic Acid, Epicatechin, and Kaempferol on UVB-Irradiated Inflammatory Marker in HaCaT Cells
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Barua, C.C.; Buragohain, L.; Rizavi, H.; Gogoi, S.B.; Rahman, R.; Siva, B.; Mounika, K.; Babu, K.S.; Pathak, D.C.; Phukan, A. Effect of seeds of Entada phaseoloides on chronic restrain stress in mice. J. Ayurveda Integr. Med. 2020, 11, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Ikegami, F.; Sekine, T.; Duangteraprecha, S.; Matsushita, N.; Matsuda, N.; Ruangrungsi, N.; Murakoshi, I.; Entadamide, C. A sulphur-containing amide from Entada phaseoloides. Phytochemistry 1988, 28, 881–882. [Google Scholar] [CrossRef]
- Sugimoto, S.; Matsunami, K.; Otsuka, H. Biological activity of Entada phaseoloides and Entada rheedei. J. Nat. Med. 2018, 72, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Mauriz, J.L.; Collado, P.S.; Veneroso, C.; Reiter, J.R.; González-Gallego, J. A review of the molecular aspects of melatonin’s anti-inflammatory actions: Recent insights and new perspectives. J. Pineal Res. 2013, 54, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Marabini, L.; Melzi, G.; Lolli, F.; Dell’Agli, M.; Piazza, S.; Sangiovanni, E.; Marinovich, M. Effects of Vitis vinifera L. leaves extract on UV radiation damage in human keratinocytes (HaCaT). J. Photochem. Photobiol. B Biol. 2020, 204, 111810–111817. [Google Scholar] [CrossRef] [PubMed]
- Sitailo, L.A.; Tibudan, S.S.; Denning, M.F. Activation of caspase-9 is required for UV-induced apoptosis of human keratinocytes. J. Biol. Chem. 2002, 277, 19346–19352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Li, N.; Wang, Y.; Ding, L.; Chen, H.; Yu, Y.; Shi, X. Protective effects of quercetin on UVB irradiation-induced cytotoxicity through ROS clearance in keratinocyte cells. Oncol. Rep. 2017, 37, 209–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Oh, J.Y.; Kim, Y.S.; Lee, H.G.; Lee, J.S.; Jeon, Y.J. Anti-photoaging and Anti-Melanogenesis Effects of Fucoidan Isolated from Hizikia fusiforme and Its Underlying Mechanisms. Mar. Drugs 2020, 18, 427. [Google Scholar] [CrossRef] [PubMed]
- Sobeh, M.; El-Raey, M.; Rezq, S.; Abdelfattah, M.A.; Petruk, G.; Osman, S.; El-Shazly, A.M.; El-Beshbishy, H.A.; Mahmoud, M.F.; Wink, M. Chemical profiling of secondary metabolites of Eugenia uniflora and their antioxidant, anti-inflammatory, pain killing and anti-diabetic activities: A comprehensive approach. J. Ethnopharmacol. 2019, 240, 111939–111950. [Google Scholar] [CrossRef] [PubMed]
- Sobeh, M.; Petruk, G.; Osman, S.; El Raey, M.A.; Imbimbo, P.; Monti, D.M.; Wink, M. Isolation of myricitrin and 3, 5-di-O-methyl gossypetin from Syzygium samarangense and evaluation of their involvement in protecting keratinocytes against oxidative stress via activation of the Nrf-2 pathway. Molecules 2019, 24, 1839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okba, M.M.; El Awdan, S.A.E.; Yousif, M.F.; El Deeb, K.S.; Soliman, F.M. Entada rheedii seeds thioamides, phenolics, and saponins and its antiulcerogenic and antimicrobial activities. J. Appl. Pharm. Sci. 2018, 8, 101–108. [Google Scholar]
- Cho, J.Y.; Moon, J.H.; Seong, K.Y.; Park, K.H. Antimicrobial Activity of 4-Hydroxybenzoic Acid and trans 4-Hydroxycinnamic Acid Isolated and Identified from Rice Hull. Biosci. Biotechol. Biochem. 1998, 62, 2273–2276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jibril, S.; Sirat, H.M.; Basar, N. Bioassay-Guided Isolation of Antioxidants and α-Glucosidase Inhibitors from the Root of Cassia sieberiana D.C. (Fabaceae). Rec. Nat. Prod. 2017, 11, 406–410. [Google Scholar]
- Ragasa, C.Y.; Pendon, Z.; Sngalang, V.; Rideout, J.A. Antimicrobial Flavones from Coleus amboinicuc. Philipp. J. Sci. 1999, 128, 347–351. [Google Scholar]
- Qian, Z.M.; Li, H.J.; Ping, L.; Rena, M.T.; Tang, D. Simultaneous Qualitation and Quantification of Thirteen Bioactive Compounds in Flos Lonicerae by High-Performance Liquid Chromatography with Diode Array Detector and Mass Spectrometry. Biol. Pharm. Bull. 2007, 55, 1073–1076. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Kong, Y.; Zu, Y.; Fu, Y.; Luo, M.; Zhang, L. Determination and quantification of active phenolic compounds in pigeon pea leaves and its medicinal product using liquid chromatography–tandem mass spectrometry. J. Chromatogr. A 2010, 1217, 4723–4731. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cao, Y.; Chen, S.; Lin, J.; Bian, J.; Huang, D. Anti-inflammation Activity of Flavones and Their Structure-Activity Relationship. J. Agric. Food Chem. 2021, 69, 7285–7302. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Woo, Y.; Cho, H. Regulation of Anti-Oxidative, Anti-Inflammatory, and Anti-Apoptotic Activity of Advanced Cooling Composition (ACC) in UVB-Irradiated Human HaCaT Keratinocytes. Int. J. Mol. Sci. 2020, 21, 6527. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Bi, Z.; Xu, A. Green tea polyphenol epigallocatechin-3-gallate inhibits the expression of nitric oxide synthase and generation of nitric oxide induced by ultraviolet B in HaCaT cells. Chin. Med. J. 2006, 119, 282–287. [Google Scholar] [CrossRef] [PubMed]
- Chen, F.; Tang, Y.; Sun, Y.; Veeraraghavan, V.P.; Mohan, S.K.; Cui, C. 6-shogaol, a active constiuents of ginger prevent UVB radiation mediated inflammation and oxidative stress through modulating NrF2 signaling in human epidermal keratinocytes (HaCaT cells). J. Photochem. Photobiol. B Biol. 2019, 197, 11518–11524. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target | Sequence | |
|---|---|---|
| Forward | Reverse | |
| GADPH | 5′-GCACCGTCAAGGCTGAGAAC-3′ | 5′-ATGGTGGTGAAGACGCCAGT-3′ |
| COX-2 | 5′-AAGTTGGCAGCAAATTGAGCA-3′ | 5′-TCCTTTTCTCCTGTGAAGGCG-3′ |
| iNOS | 5′-TACTCCACCAACAATGGCAA-3′ | 5′-ATAGCGGATGAGCTGAGCAT-3′ |
| Peak No. | Retention Time, tR (min) | Formula | [M-H]− (m/z exp) | Extract Mass | Error (ppm) | Identification |
|---|---|---|---|---|---|---|
| 1 | 1.80 | C7H6O5 | 169.0142 | 170.0215 | −0.19 | Gallic acid |
| 2 | 2.67 | C7H6O4 | 153.0199 | 154.0271 | 4.04 | Protocatechuic acid |
| 3 | 3.57 | C7H6O3 | 137.0243 | 138.0322 | −0.9 | 4-Hydroxybenzoic acid |
| 4 | 3.90 | C15H14O6 | 289.0734 | 290.0795 | 5.29 | Epicatechin |
| 5 | 5.69 | C21H20O11 | 447.0941 | 448.1011 | 1.71 | Luteolin-7-O-β-d-glucoside |
| 6 | 7.27 | C17H14O6 | 313.0716 | 314.0790 | −0.26 | Cirsimaritin |
| 7 | 7.57 | C15H10O6 | 285.0408 | 286.0477 | 1.24 | Luteolin |
| 8 | 7.65 | C15H10O7 | 301.0358 | 302.0432 | 0.68 | Quercetin |
| 9 | 7.92 | C15H12O6 | 287.0567 | 288.2500 | −1.55 | Dihydrokaempferol |
| 10 | 8.65 | C15H10O5 | 269.0459 | 270.0533 | 0.78 | Apigenin |
| 11 | 8.90 | C15H10O6 | 285.0408 | 286.0477 | 1.24 | Kaempferol |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mittraphab, Y.; Amen, Y.; Nagata, M.; Matsumoto, M.; Wang, D.; Shimizu, K. Anti-Phototoxicity Effect of Phenolic Compounds from Acetone Extract of Entada phaseoloides Leaves via Activation of COX-2 and iNOS in Human Epidermal Keratinocytes. Molecules 2022, 27, 440. https://doi.org/10.3390/molecules27020440
Mittraphab Y, Amen Y, Nagata M, Matsumoto M, Wang D, Shimizu K. Anti-Phototoxicity Effect of Phenolic Compounds from Acetone Extract of Entada phaseoloides Leaves via Activation of COX-2 and iNOS in Human Epidermal Keratinocytes. Molecules. 2022; 27(2):440. https://doi.org/10.3390/molecules27020440
Chicago/Turabian StyleMittraphab, Yanisa, Yhiya Amen, Maki Nagata, Masako Matsumoto, Dongmei Wang, and Kuniyoshi Shimizu. 2022. "Anti-Phototoxicity Effect of Phenolic Compounds from Acetone Extract of Entada phaseoloides Leaves via Activation of COX-2 and iNOS in Human Epidermal Keratinocytes" Molecules 27, no. 2: 440. https://doi.org/10.3390/molecules27020440
APA StyleMittraphab, Y., Amen, Y., Nagata, M., Matsumoto, M., Wang, D., & Shimizu, K. (2022). Anti-Phototoxicity Effect of Phenolic Compounds from Acetone Extract of Entada phaseoloides Leaves via Activation of COX-2 and iNOS in Human Epidermal Keratinocytes. Molecules, 27(2), 440. https://doi.org/10.3390/molecules27020440