
The Content of Phenolic Compounds and Organic Acids in Two Tagetes patula Cultivars Flowers and Its Dependence on Light Colour and Substrate

 , , , and
, , , and
Abstract
:1. Introduction
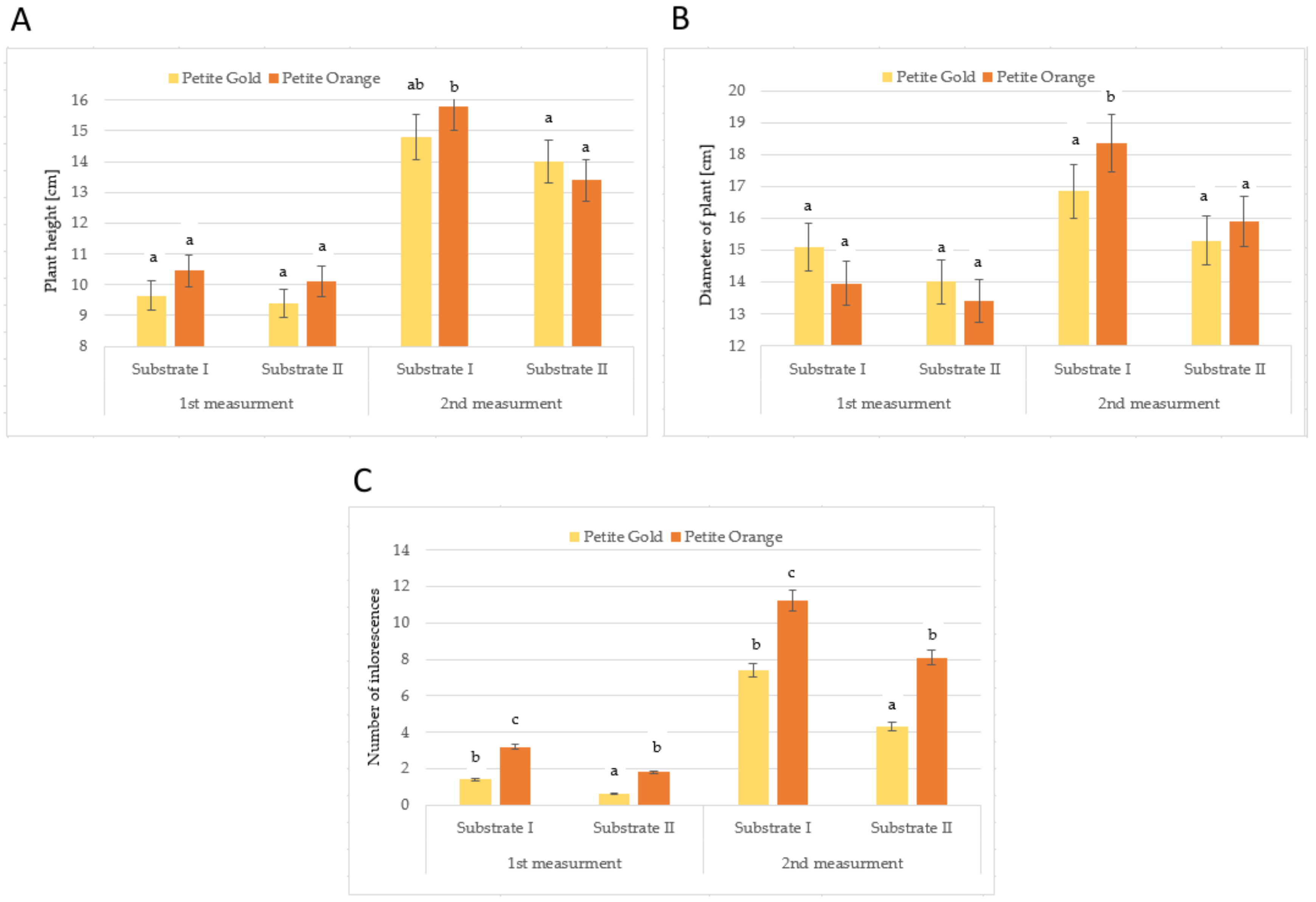
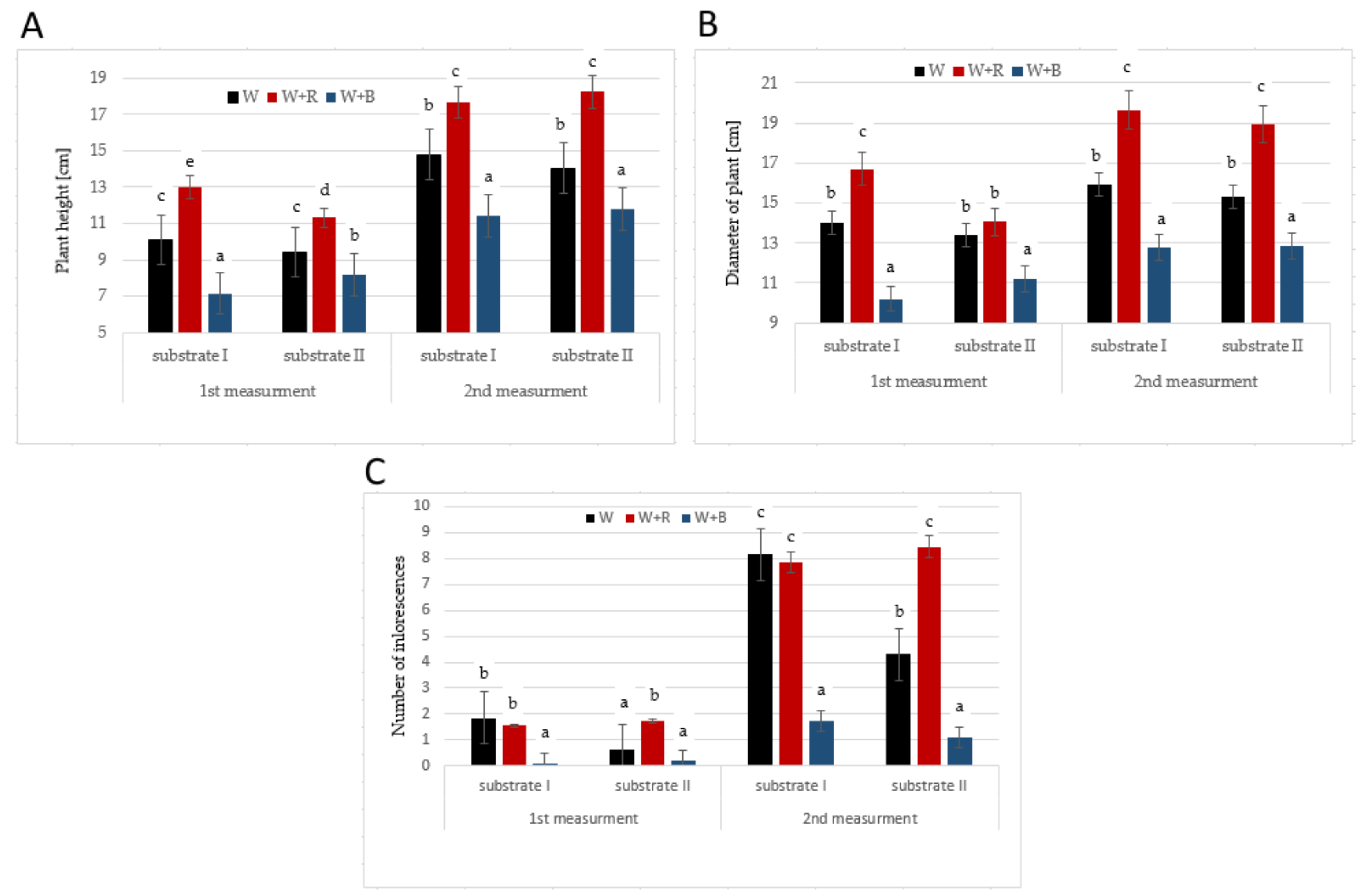
2. Results and Discussion
3. Materials and Methods
3.1. Experimental Materials and Design
3.2. Morphological Measurements
3.3. Sample Preparation
3.4. Determination of Phenolic Compounds and Organic Acids
3.5. Determination of Total Phenolic Content (TPC)
3.6. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Meléndez-Martínez, A.J.; Benítez, A.; Corell, M.; Hernanz, D.; Mapelli-Brahm, P.; Stinco, C.; Coyago-Cruz, E. Screening for innovative sources of carotenoids and phenolic antioxidants among flowers. Foods 2021, 10, 2625. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Meenu, M.; Xu, B. A systematic investigation on free phenolic acids and flavonoids profiles of commonly consumed edible flowers in China. J. Pharm. Biomed. Anal. 2019, 172, 268–277. [Google Scholar] [CrossRef]
- Vuolo, M.M.; Lima, V.S.; Maróstica, M.R., Jr. Phenolic compounds: Structure, classification, and antioxidant power. In Bioactive Compounds, Health Benefits and Potential Applications; Campos, M.R.S., Ed.; Elsevier: Sawston, UK; Cambridge, MA, USA, 2019; pp. 33–50. [Google Scholar]
- Lim, T.K. Tagetes patula. In Edible Medicinal and Non Medicinal Plants: Volume 7. Flowers; Lim, T.K., Ed.; Springer: Dordrecht, The Netherlands, 2014; pp. 456–468. [Google Scholar]
- Zambrano, V.M.L.; Rivadeneira, A.A.D.; Nevarez, J.G.C.; Rodriguez-Diaz, J.M. Phytochemical characterization, antioxidant and antibacterial activity of essential oil and extracts of Tagetes patula on Staphylococcus aureus. Rev. Fac. Agron. (LUZ) 2020, 37, 347–367. [Google Scholar] [CrossRef]
- Chitrakar, B.; Zhang, M.; Bhandari, B. Edible flowers with the common name “marigold’: Their therapeutic values and processing. Trends Food Sci. Technol. 2019, 89, 76–87. [Google Scholar] [CrossRef]
- Egebjerg, M.M.; Olesen, P.T.; Eriksen, F.D.; Ravn-Haren, G.; Bredsdorff, L.; Pilegaard, K. Are wild and cultivated flowers served in restaurants or sold by local producers in Denmark safe for the consumer? Food Chem. Toxicol. 2018, 120, 129–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopsell, D.A.; Sams, C.E. Increases in shoot tissue pigments, glucosinolates, and mineral elements in sprouting broccoli after exposure to short-duration blue light from light emitting diodes. J. Am. Soc. Hortic. Sci. 2013, 138, 31–37. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, S.D.; Folta, K.M. Sequential light programs shape kale (Brassica napus) sprout appearance and alter metabolic and nutrient content. Hortic. Res. 2014, 1, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frąszczak, B. The effect of different doses of blue light on the biometric traits and photosynthesis of dill plants. Not. Bot. Horti Agrobot. Cluj Napoca 2016, 44, 34–40. [Google Scholar] [CrossRef] [Green Version]
- Bian, Z.H.; Yang, Q.C.; Liu, W.K. Effects of light quality on the accumulation of phytochemicals in vegetables produced in controlled environments: A review. J. Sci. Food Agric. 2015, 95, 869–877. [Google Scholar] [CrossRef]
- Son, K.-H.; Oh, M.-M. Leaf shape, growth, and antioxidant phenolic compounds of two lettuce cultivars grown under various combinations of blue and red light-emitting diodes. Hortic. Sci. 2013, 48, 988–995. [Google Scholar] [CrossRef]
- Kopsell, D.A.; Sams, C.E.; Morrow, R.C. Blue wavelengths from LED lighting increase nutritionally important metabolites in specialty crops. Hortic. Sci. 2015, 50, 1285–1288. [Google Scholar] [CrossRef] [Green Version]
- Hasan, M.M.; Bashir, T.; Ghosh, R.; Lee, S.K.; Bae, H. An Overview of LEDs’ effects on the production of bioactive compounds and crop quality. Molecules 2017, 22, 1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gam, D.T.; Khoi, P.H.; Ngoc, P.B.; Linh, L.K.; Hung, N.K.; Anh, P.T.L.; Thu, N.T.; Hien, N.T.T.; Khanh, T.D.; Ha, C.H. LED Lights promote growth and flavonoid accumulation of Anoectochilus roxburghii and are linked to the enhanced expression of several related genes. Plants 2020, 9, 1344. [Google Scholar] [CrossRef] [PubMed]
- Japundžić-Palenkić, B.; Jagodar, K.; Romanjek Fajdetić, N.; Božić-Ostojić, L. Effect of Growing Media on Tagetes patula Nana Plants Quality, Proceedings of the 9th International Scientific and Expert Conference, TEAM 2018, Novi Sad, Serbia, 10–12 October 2018; Lalić, B., Ed.; University of Novi Sad: Novi Sad, Serbia, 2018; pp. 314–320. [Google Scholar]
- Moccaldi, L.E.; Runkle, E.S. Modeling the effects of temperature and photosynthetic daily light integral on growth and flowering of Salvia splendens and Tagetes patula. J. Am. Soc. Hort. Sci. 2007, 132, 283–288. [Google Scholar] [CrossRef] [Green Version]
- Król, B. Yield and the chemical composition of flower heads of pot marigold (Calendula officinalis L. cv. Orange King) depending on nitrogen fertilization. Acta Sci. Pol. Hortorum Cultus 2011, 10, 235–243. [Google Scholar]
- Zhang, M.; Park, Y.; Runkle, E.S. Regulation of extension growth and flowering of seedlings by blue radiation and the red to far-red ratio of sole-source lighting. Sci. Hortic. 2020, 272, 109478. [Google Scholar] [CrossRef]
- Wollaeger, H.M.; Runkle, E.S. Growth and acclimation of impatiens, salvia, petunia, and tomato seedlings to blue and red light. Hort. Sci. 2015, 50, 522–529. [Google Scholar] [CrossRef] [Green Version]
- Snowden, M.C.; Cope, K.R.; Bugbee, B. Sensitivity of seven diverse species to blue and green light: Interactions with photon flux. PLoS ONE 2016, 11, e0163121. [Google Scholar] [CrossRef]
- Frąszczak, B.; Kula-Maximenko, M. The preferences of different cultivars of lettuce seedlings (Lactuca sativa L.) for the spectral composition of light. Agronomy 2021, 11, 1211. [Google Scholar] [CrossRef]
- Banerjee, R.; Batschauer, A. Plant blue-light receptors. Planta 2005, 20, 498–502. [Google Scholar] [CrossRef]
- Schwend, T.; Prucker, D.; Mempel, H. Red light promotes compact growth of sunflowers. Eur. J. Hortic. Sci. 2015, 80, 56–61. [Google Scholar] [CrossRef]
- Nissim-Levi, A.; Kitron, M.; Nishri, Y.; Ovadia, R.; Forer, I.; Oren-Shamir, M. Effects of blue and red LED lights on growth and flowering of Chrysanthemum morifolium. Sci. Hortic. 2019, 254, 77–83. [Google Scholar] [CrossRef]
- Bergstrand, K.-J.; Asp, H.; Schüssler, H.K. Development and acclimatisation of horticultural plants subjected to narrow-band lighting. Europ. J. Hort. Sci. 2014, 72, 45–51. [Google Scholar]
- Aliniaeifard, S.; Seif, M.; Arab, M.; Zare Mehrjerdi, M.; Li, T.; Lastochkina, O. Growth and photosynthetic performance of Calendula officinalis under monochromatic red light. Int. J. Hortic. Sci. Technol. 2018, 5, 123–132. [Google Scholar] [CrossRef]
- Sabzalian, M.R.; Heydarizadeh, P.; Zahedi, M.; Boroomand, A.; Agharokh, M.; Sahba, M.; Schoefs, B. High performance of vegetables, flowers, and medicinal plants in a red-blue LED incubator for indoor plant production. Agron. Sustain. Dev. 2014, 34, 879–886. [Google Scholar] [CrossRef] [Green Version]
- Frąszczak, B.; Janowska, B.; Świerk, D. The morphological’ responses of Calendula officinalis L. ‘Radio’ to the foliar application of benzyladenine and different light spectra. Agronomy 2021, 11, 460. [Google Scholar] [CrossRef]
- Park, Y.; Runkle, E.S. Blue radiation attenuates the effects of the red to far-red ratio on extension growth but not on flowering. Environ. Exp. Bot. 2019, 168, 103871. [Google Scholar] [CrossRef]
- Youssef, H.; Ali, S.; Sanad, M.; Dawood, D. Chemical investigation of flavonoid, phenolic acids composition and antioxidant activity of Tagetes erecta flowers. Egypt. J. Chem. 2020, 63, 2605–2615. [Google Scholar] [CrossRef]
- Szopa, A.; Starzec, A.; Ekiert, H. The importance of monochromatic lights in the production of phenolic acids and flavonoids in shoot cultures of Aronia melanocarpa, Aronia arbutifolia and Aronia × prunifolia. J. Photochem. Photobiol. B Biol. 2018, 179, 91–97. [Google Scholar] [CrossRef]
- Jiang, H.; Li, X.; Tian, J.; Liu, H. Pre-harvest supplemental blue light enhanced antioxidant activity of flower stalk in chinese kale during storage. Plants 2021, 10, 1177. [Google Scholar] [CrossRef]
- Fu, B.; Ji, X.; Zhao, M.; He, F.; Wang, X.; Wang, Y.; Liu, P.; Niu, L. The influence of light quality on the accumulation of flavonoids in tobacco (Nicotiana tabacum L.) leaves. J. Photochem. Photobiol. B Biol. 2016, 162, 544–549. [Google Scholar] [CrossRef] [PubMed]
- Ravaglia, J.D.; Espley, R.V.; Henry-Kirk, R.A.; Andreotti, C.; Ziosi, V.; Hellens, R.P.; Costa, G.; Allan, A.C. Transcriptional regulation of flavonoid biosynthesis in nectarine (Prunus persica) by a set of R2R3 MYB transcription factors. BMC Plant Biol. 2013, 13, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sergejeva, D.; Alsina, I.; Duma, M.; Dubova, L.; Augspole, I.; Erdberga, I.; Berzina, K. Evaluation of different lighting sources on the growth and chemical composition of lettuce. Agron. Res. 2018, 16, 892–899. [Google Scholar] [CrossRef]
- Ayub, M.A.; Hussain, A.I.; Hanif, M.A.; Chatha, A.A.S.; Kamal, G.M.; Shahid, M.; Janneh, O. Variation in phenolic profile, β-carotene and flavonoid contents, biological activities of two Tagetes species from Pakistani flora. Chem. Biodivers. 2017, 14, e1600463. [Google Scholar] [CrossRef] [PubMed]
- López-Bucio, J.; Nieto-Jacobo, M.F.; Ramírez-Rodríguez, V.; Herrera-Estrella, L. Organic acid metabolism in plants: From adaptive physiology to transgenic varieties for cultivation in extreme soils. Plant Sci. 2000, 160, 1–13. [Google Scholar] [CrossRef]
- Pires, T.C.; Dias, M.I.; Barros, L.; Ferreira, I.C. Nutritional and chemical characterization of edible petals and corresponding infusions: Valorization as new food ingredients. Food Chem. 2017, 220, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Nowosielski, O. Methods for Determining the Need for Fertilization, 2nd ed.; Państwowe Wydawnictwo Rolnicze i Leśne: Warszawa, Poland, 1974. [Google Scholar]
- Czerniawska, W.; Strahl, A. Metody Badań Laboratoryjnych w Stacjach Chemiczno-Rolniczych: Badania Gleb, Ziem i Podłoży Spod Warzyw i Kwiatów Oraz Części Wskaźnikowych Roślin w Celach Diagnostycznych (Methods of Laboratory Tests in Chemical-Agricultural Stations: Research of Soils, Soil and Substrates under Vegetables and Flowers as well as Indicator Parts of Plants for Diagnostic Purposes); Institute of Soil Science and Plant Cultivation: Puławy, Poland, 1983. [Google Scholar]

{kind=link}
{kind=link}
| Light Colour (A) | Substrate (B) | N-NO3 | N-NH4 | P | K | Ca | Mg | Na | S-SO4 |
|---|---|---|---|---|---|---|---|---|---|
| White | I | 7.0 a | 26.5 a | 63.5 a | 63.5 a | 1246.0 ab | 212.0 a | 138.0 a | 316.5 a |
| II | 14.0 a | 42.0 a | 103.5 ab | 118.0 ab | 1100.0 a | 225.5 ab | 125.5 a | 364.0 ab | |
| White + blue | I | 28.0 a | 17.5 a | 37.5 a | 188.0 bc | 1401.0 b | 244.0 ab | 150.0 a | 415.0 b |
| II | 19.5 a | 24.5 a | 58.5 a | 217.0 c | 1294.5 ab | 262.5 b | 136.0 a | 388.5 ab | |
| White + red | I | 2.0 a | 23.0 a | 183.0 b | 183.0 bc | 1246.0 ab | 252.5 ab | 139.0 a | 369.5 ab |
| II | 7.0 a | 26.5 a | 63.5 a | 63.5 a | 1246.0 ab | 212.0 a | 138.0 a | 316.5 a | |
| Mean for A | White | 10.5 ab | 34.3 a | 83.5 a | 90.8 a | 1173.0 a | 218.8 a | 131.8 a | 340.3 a |
| White + blue | 23.8 b | 21.0 a | 48.0 a | 202.5 b | 1347.8 b | 253.3 b | 143.0 a | 401.8 b | |
| White + red | 4.5 a | 24.8 a | 123.3 b | 123.8 a | 1246.0 ab | 232.3 ab | 138.5 a | 343.0 a | |
| Mean for B | I | 12.3 a | 22.4 a | 94.7 b | 144.8 b | 1297.7 a | 236.2 b | 142.3 a | 367.0 a |
| II | 13.5 a | 31.0 a | 75.2 a | 132.8 a | 1213.5 a | 233.3 a | 133.2 a | 356.3 a |
| Light Colour (A) | Substrate (B) | Fe | Mn | Zn | Cu | Cl | pH | EC |
|---|---|---|---|---|---|---|---|---|
| White | I | 13.05 ab | 1.45 ab | 16.70 a | 0.30 a | 52.0 a | 6.82 ab | 0.65 a |
| II | 12.55 a | 1.70 ab | 16.60 a | 0.30 a | 82.0 ab | 6.58 a | 1.05 ab | |
| White + blue | I | 19.10 b | 3.20 b | 23.65 b | 0.40 a | 116.5 b | 6.61 ab | 1.05 ab |
| II | 13.15 ab | 2.40 ab | 18.85 a | 0.30 a | 111.0 b | 6.62 ab | 1.15 b | |
| Red + white | I | 15.10 ab | 1.40 b | 16.35 a | 0.30 a | 55.5 a | 6.88 b | 0.70 ab |
| II | 11.55 a | 1.30 a | 15.10 a | 0.30 a | 92.0 ab | 6.73 ab | 0.90 ab | |
| Mean for A | White | 12.80 a | 1.58 ab | 16.65 a | 0.30 a | 67.0 a | 6.70 a | 0.85 a |
| White + blue | 16.13 a | 2.80 b | 21.25 b | 0.35 a | 113.8 b | 6.62 a | 1.10 a | |
| White + red | 13.33 a | 1.35 a | 15.73 a | 0.30 a | 73.8 a | 6.81 a | 0.80 a | |
| Mean for B | I | 15.75 a | 2.02 a | 18.90 a | 0.33 a | 74.7 a | 6.77 a | 0.80 a |
| II | 12.42 a | 1.80 a | 16.85 a | 0.30 a | 95.0 a | 6.64 a | 1.03 a |
| Cultivar (A) | Light Colour (B) | Substrate (C, c) | TPC [mg GAE·g−1FW] | Catechin | Quercetin | Rutin | Sum |
|---|---|---|---|---|---|---|---|
| Petite Orange | White | I | 1.40 A | 1.99 A | 869.3 C | 3.53 A | 874.8 C |
| II | 1.55 A | 2.85 A | 180.1 A | 3.80 A | 186.7 A | ||
| Petite Gold | White | I | 1.32 A ab | 2.33 A a | 788.9 C a | 11.83 B b | 803.1 C a |
| II | 1.19 A ab | 6.43 B b | 482.3 B a | 32.78 C c | 521.5 B a | ||
| White + blue | I | 1.19 ab | 5.18 b | 420.8 a | 1.73 a | 421.7 a | |
| II | 1.35 b | 9.25 c | 425.8 a | nd | 435.0 a | ||
| White + red | I | 1.22 ab | 2.87 a | 1574.8 c | 7.73 b | 1585.4 c | |
| II | 1.12 a | 5.36 b | 1201.5 b | 1.25 a | 1208.2 b | ||
| Mean for A | Petite Orange | 1.48 A | 2.41 A | 524.7 A | 3.66 A | 530.8 A | |
| Petite Gold | 1.26 A | 4.38 B | 635.6 B | 22.31 B | 662.3 A | ||
| Mean for B | White | 1.25 a | 4.38 a | 635.6 a | 22.31 b | 662.3 a | |
| White + blue | 1.27 a | 7.22 b | 423.3 a | 0.87 a | 431.4 a | ||
| White + red | 1.17 a | 4.12 a | 1388.2 b | 4.49 a | 1396.8 b | ||
| Mean for C | I | 1.36 A | 2.16 A | 829.1 B | 7.68 A | 839.0 B | |
| II | 1.37 A | 4.64 B | 331.2 A | 18.29 B | 354.1 A | ||
| Mean for c | I | 1.24 a | 3.46 a | 928.2 b | 7.10 a | 938.7 b | |
| II | 1.22 a | 7.02 b | 703.2 a | 11.34 b | 721.6 a |
| Cultivar (A) | Light Colour (b) | Substrate (C, c) | 2.5-DHBA | 4-HBA | Caffeic | Chlorogenic | Trans-Cinnamic | Gallic | Protocatechuic | Sinapic | Syringic | Vanillic | Sum |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Petite Orange | White | I II | nd nd | 1.32 A 1.62 A | nd 1.17 B | 1.78 B 3.05 C | nd nd | 2.70 A 4.06 B | nd nd | nd nd | nd nd | 1.32 A 5.07 B | 7.12 A 14.96 B |
| Petite Gold | White | I | 2.21 A a | 1.24 A a | nd | nd | 4.38 B b | 6.93 C c | 1.88 B c | 1.71 B b | 1.40 B a | 4.33 B a | 24.10 C a |
| II | 29.35 B c | 6.90 B b | 4.54 C c | 3.21 C c | nd | 3.88 B ab | 1.59 B bc | nd | 1.29 B a | 4.32 B a | 55.08 D c | ||
| White + blue | I | 28.90 c | 4.26 ab | 2.57 bc | nd | 17.83 d | 8.06 c | nd | 2.57 b | 7.95 c | 8.41 a | 80.54 d | |
| II | 44.48 d | 5.97 ab | 1.96 ab | 1.53 b | 4.04 b | 5.84 bc | 1.25 b | 2.56 b | 8.20 c | 7.26 a | 83.09 d | ||
| White + red | I II | 2.71 a 12.82 b | 6.86 b 1.47 a | 1.43 ab nd | 3.98 c nd | 3.87 b 11.96 c | 7.40 c 2.50 a | 3.38 d 2.16 c | 1.66 b nd | 4.96 b 1.11 a | 5.41 a 4.45 a | 41.65 b 36.47 b | |
| Mean for A | Petite Orange Petite Gold | nd 15.80 B | 1.47 A 4.07 B | 0.58 A 2.27 B | 2.41 B 1.61 A | nd 2.19 B | 3.38 A 5.41 B | nd 1.74 B | nd 0.85 B | nd 1.35 B | 3.19 A 4.33 A | 11.04 A 39.59 B | |
| Mean for b | White White + blue White + red | 15.78 b 36.69 c 7.76 a | 4.07 a 5.11 a 4.17 a | 2.27 a 2.26 a 0.71 a | 1.61 b 0.77 a 1.99 b | 2.19 a 10.94 c 7.91 b | 5.41 ab 6.95 b 4.95 a | 1.74 b 0.62 a 2.77 c | 0.85 a 2.56 b 0.83 a | 1.34 a 8.07 b 3.04 b | 4.32 a 7.84 a 4.93 a | 39.59 a 81.82 b 39.06 a | |
| Mean for C | I | 1.11 A | 1.28 A | nd | 0.89 A | 2.19 B | 4.82 B | 0.94 A | 0.85 B | 0.70 A | 2.82 A | 15.61 A | |
| II | 14.67 B | 4.26 B | 2.85 B | 3.13 B | nd | 3.97 A | 0.79 A | nd | 0.65 A | 4.70 A | 35.02 B | ||
| Mean for c | I | 11.27 a | 4.12 a | 1.33 a | 1.32 a | 8.69 b | 7.47 b | 1.76 a | 1.98 b | 4.77 b | 6.05 a | 48.76 a | |
| II | 28.88 b | 4.78 a | 2.16 a | 1.58 a | 5.33 a | 4.07 a | 1.67 a | 0.85 a | 3.53 a | 5.35 a | 58.22 b |
| Cultivar (A) | Light Colour (b) | Substrate (C, c) | Acetic | Citric | Fumaric | Malic | Malonic | Succinic | Quinic | Sum |
|---|---|---|---|---|---|---|---|---|---|---|
| Petite Orange | White | I II | nd nd | nd nd | 0.84 A nd | 3.63 AB 3.16 AB | 4.21 A 4.55 A | nd 3.45 B | 0.70 A 2.70 B | 9.39 A 14.11 A |
| Petite Gold | White | I | nd | 5.97 B bc | nd | 4.85 B a | 9.68 B c | nd | 1.24 A a | 21.74 B a |
| II | 0.91 Bab | 9.26 C cd | 9.28 B b | 2.65 A a | 10.65 B c | 0.52 A ab | 1.23 A b | 34.51 C b | ||
| White + blue | I | 2.41 ab | 3.91 ab | nd | 12.42 b | 4.89 ab | 2.38 ab | 2.63 c | 28.64 ab | |
| II | 11.62 c | 3.20 ab | nd | 16.91 c | 8.78 bc | 4.74 b | 0.98 ab | 46.22 c | ||
| White + red | I II | nd 3.15 b | nd 13.93 d | 0.37 a nd | 6.88 a 3.98 a | 11.55 c 2.16 a | 19.66 d 9.63 c | 0.55 a 2.20 c | 39.02 bc 35.06 b | |
| Mean for A | Petite Orange Petite Gold | nd 0.45 B | nd 7.61 B | 0.42 A 4.64 B | 3.40 A 3.75 A | 4.38 A 10.17 B | 1.73 B 0.26 A | 1.70 A 1.24 A | 11.75 A 28.12 B | |
| Mean for b | White | 15.78 b 36.69 c 7.76 a | 4.07 a 5.11 a 4.17 a | 4.64 b nd 0.18 a | 1.61 b 0.77 a 1.99 b | 2.19 a 10.94 c 7.91 b | 5.41 ab 6.95 b 4.95 a | 1.74 b 0.62 a 2.77 c | 0.85 a 2.56 b 0.83 a | |
| White + blue | ||||||||||
| White + red | ||||||||||
| Mean for C | I | nd | 2.99 A | 0.42 A | 4.24 B | 6.95 A | nd | 0.97 A | 15.56 A | |
| II | 0.45 B | 4.63 A | 4.64 B | 2.91 A | 7.60 A | 1.99 B | 1.97 B | 24.31 B | ||
| Mean for c | I | 11.27 a | 4.12 a | 0.12 a | 1.32 a | 8.69 b | 7.47 b | 1.76 a | 1.98 b | |
| II | 28.88 b | 4.78 a | 3.09 b | 1.58 a | 5.33 a | 4.07 a | 1.67 a | 0.85 a |
| Light Colour | Wavelength (nm) | PFD * (μmol m−2 s−1) | % for W Light | % for W + R | % for W + B |
|---|---|---|---|---|---|
| UV | 320–380 | 0.5 | 0.3 | 0.2 | 0.2 |
| Violet | 380–450 | 15.4 | 8.9 | 6.6 | 6.6 |
| Blue | 450–495 | 30.3 | 17.6 | 13.0 | 38.9 |
| Green | 495–570 | 53.5 | 31.1 | 23.0 | 23.0 |
| Gold | 570–590 | 18.7 | 10.9 | 8.1 | 8.1 |
| Orange | 590–620 | 21.8 | 12.7 | 9.4 | 9.4 |
| Red (R) | 620–700 | 26.4 | 15.3 | 37.2 | 11.4 |
| Far Red (FR) | 700–780 | 5.6 | 3.3 | 2.4 | 2.4 |
| sum | 320–780 | 172.2 | 100 | 100 | 100 |
| R:FR | 4.7 | - | 15.5 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krzymińska, A.; Frąszczak, B.; Gąsecka, M.; Magdziak, Z.; Kleiber, T. The Content of Phenolic Compounds and Organic Acids in Two Tagetes patula Cultivars Flowers and Its Dependence on Light Colour and Substrate. Molecules 2022, 27, 527. https://doi.org/10.3390/molecules27020527
Krzymińska A, Frąszczak B, Gąsecka M, Magdziak Z, Kleiber T. The Content of Phenolic Compounds and Organic Acids in Two Tagetes patula Cultivars Flowers and Its Dependence on Light Colour and Substrate. Molecules. 2022; 27(2):527. https://doi.org/10.3390/molecules27020527
Chicago/Turabian StyleKrzymińska, Agnieszka, Barbara Frąszczak, Monika Gąsecka, Zuzanna Magdziak, and Tomasz Kleiber. 2022. "The Content of Phenolic Compounds and Organic Acids in Two Tagetes patula Cultivars Flowers and Its Dependence on Light Colour and Substrate" Molecules 27, no. 2: 527. https://doi.org/10.3390/molecules27020527