
Ameliorative Effect of Imperatorin on Dermatophagoides pteronyssinus-Induced Allergic Asthma by Suppressing the Th2 Response in Mice
 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Effects of Imperatorin on Der p-Specific IgE, IgG1, and IgG2a in Serum
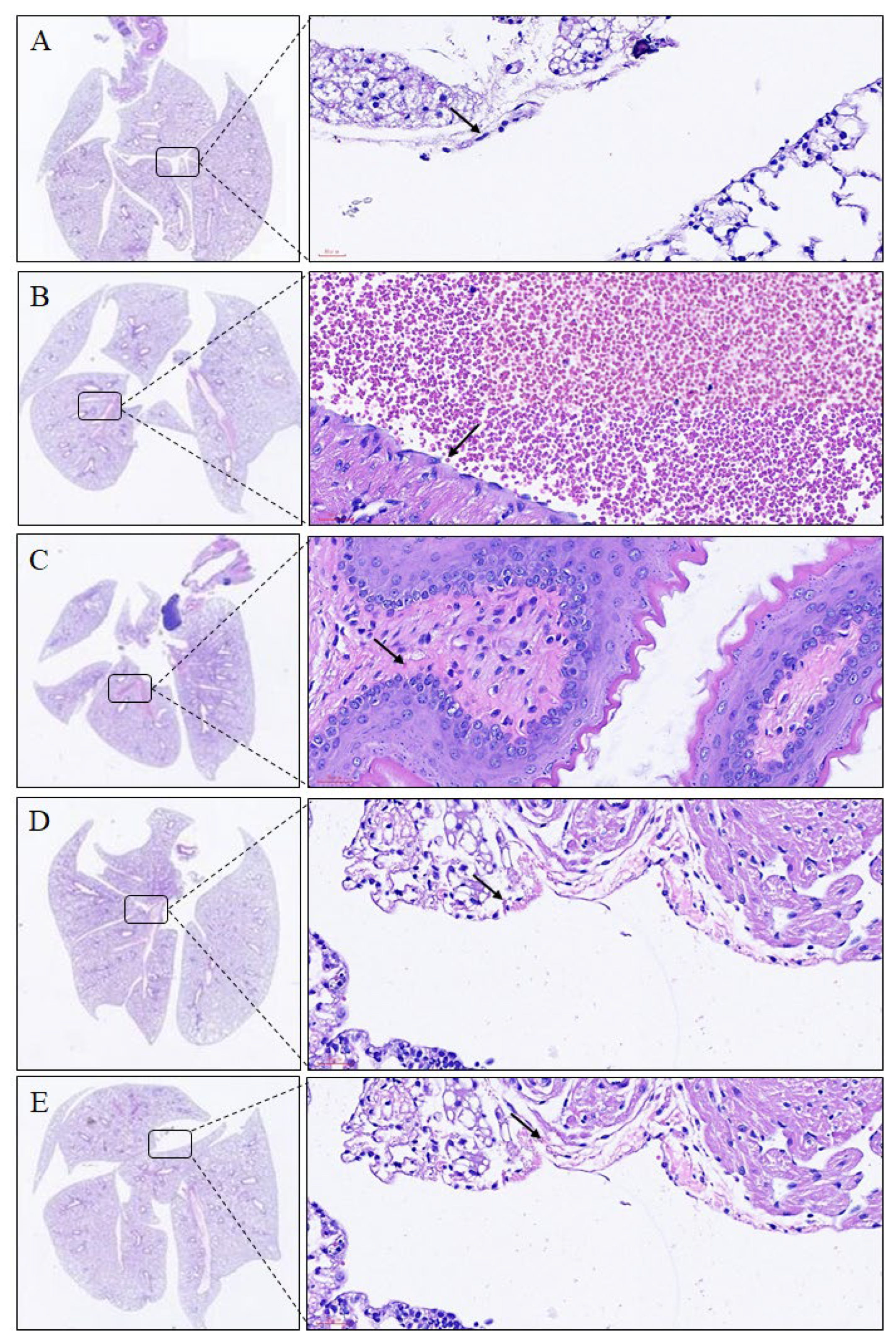
2.2. Histopathological Analyses of Lungs and Trachea
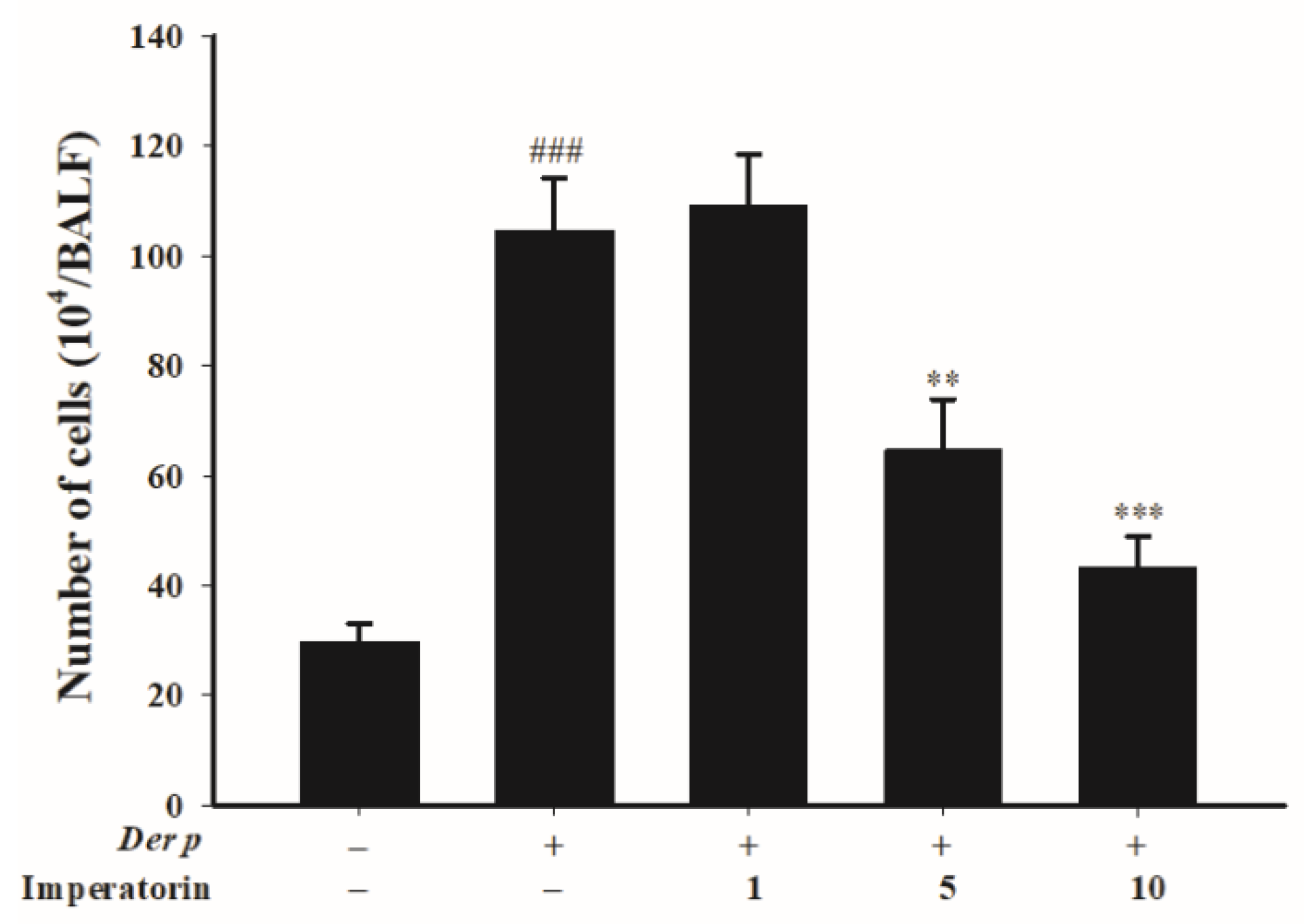
2.3. Effects of Imperatorin on Inflammatory Cell Count in Bronchoalveolar Lavage Fluid (BALF)
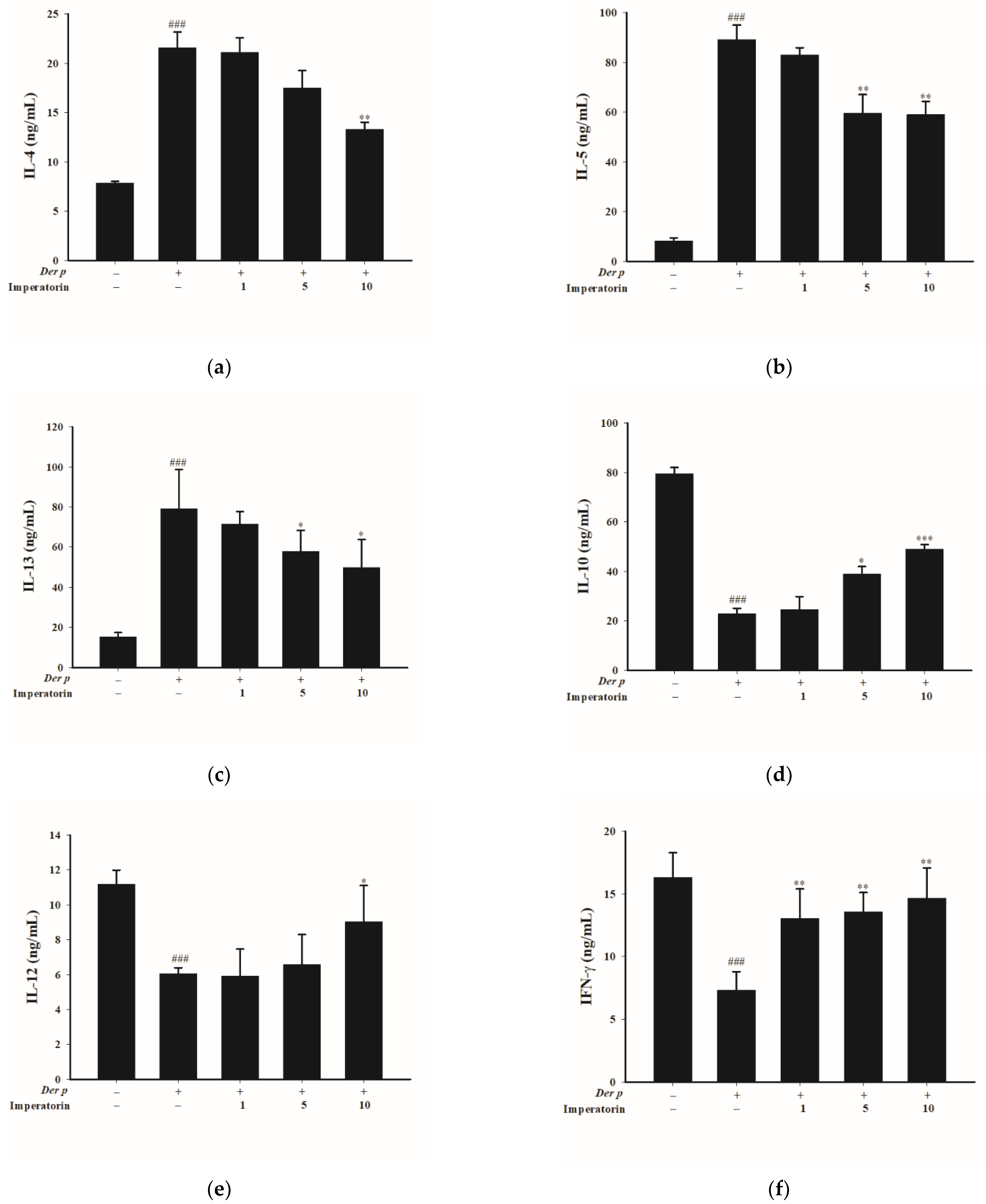
2.4. Effects of Imperatorin on Cytokines in BALF of Der p-Induced Allergic Asthma Mice
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals
4.3. Dust Mite Allergen Mouse Model
4.4. Collection of Mouse Serum and BALF
4.5. Lung Histology
4.6. Determination of Der p-Specific IgE, IgG1, and IgG2a in Serum
4.7. Th1 and Th2 Cytokine Expression Detection in BALF
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Froidure, A.; Shen, C.; Pilette, C. Dendritic cells revisited in human allergic rhinitis and asthma. Allergy 2016, 71, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Lambrecht, B.N.; Hammad, H. Allergens and the airway epithelium response: Gateway to allergic sensitization. J. Allergy Clin. Immunol. 2014, 134, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Lai, C.L.; Shyur, S.D.; Wu, C.Y.; Chang, C.L.; Chu, S.H. Specific IgE to 5 different major house dust mites among asthmatic children. Acta Paediatr. Taiwanica 2002, 43, 265–270. [Google Scholar]
- Wan, H.; Winton, H.L.; Soeller, C.; Tovey, E.R.; Gruenert, D.C.; Thompson, P.J.; Stewart, G.A.; Taylor, G.W.; Garrod, D.R.; Cannell, M.B.; et al. Der p 1 facilitates transepithelial allergen delivery by disruption of tight junctions. J. Clin. Investig. 1999, 104, 123–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambrecht, B.N.; Salomon, B.; Klatzmann, D.; Pauwels, R.A. Dendritic cells are required for the development of chronic eosinophilic airway inflammation in response to inhaled antigen in sensitized mice. J. Immunol. 1998, 160, 4090–4097. [Google Scholar] [PubMed]
- Novak, N.; Kraft, S.; Bieber, T. IgE receptors. Curr. Opin. Immunol. 2001, 13, 721–726. [Google Scholar] [CrossRef]
- Rothenberg, M.E. Eosinophilia. N. Engl. J. Med. 1998, 338, 1592–1600. [Google Scholar] [CrossRef]
- Sumi, Y.; Hamid, Q. Airway remodeling in asthma. Allergol. Int. 2007, 56, 341–348. [Google Scholar] [CrossRef] [Green Version]
- Taher, Y.A.; Henricks, P.A.; van Oosterhout, A.J. Allergen-specific subcutaneous immunotherapy in allergic asthma: Immunologic mechanisms and improvement. Libyan J. Med. 2010, 5, 5303. [Google Scholar] [CrossRef]
- Deng, M.; Xie, L.; Zhong, L.; Liao, Y.; Liu, L.; Li, X. Imperatorin: A review of its pharmacology, toxicity and pharmacokinetics. Eur. J. Pharmacol. 2020, 879, 173124. [Google Scholar] [CrossRef]
- Mukherjee, A.B.; Zhang, Z. Allergic asthma: Influence of genetic and environmental factors. J. Biol. Chem. 2011, 286, 32883–32889. [Google Scholar] [CrossRef] [PubMed]
- Afshar, R.; Medoff, B.D.; Luster, A.D. Allergic asthma: A tale of many T cells. Clin. Exp. Allergy 2008, 38, 1847–1857. [Google Scholar] [CrossRef] [PubMed]
- Shum, B.O.; Rolph, M.S.; Sewell, W.A. Mechanisms in allergic airway inflammation-lessons from studies in the mouse. Expert. Rev. Mol. Med. 2008, 10, e15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barnes, P.J. Corticosteroids, IgE, and atopy. J. Clin. Investig. 2001, 107, 265–266. [Google Scholar] [CrossRef]
- Oh, H.A.; Kim, H.M.; Jeong, H.J. Distinct effects of imperatorin on allergic rhinitis: Imperatorin inhibits caspase-1 activity in vivo and in vitro. J. Pharmacol. Exp. Ther. 2011, 339, 72–81. [Google Scholar] [CrossRef]
- Lin, C.L.; Hsiao, G.; Wang, C.C.; Lee, Y.L. Imperatorin exerts antiallergic effects in Th2-mediated allergic asthma via induction of IL-10-producing regulatory T cells by modulating the function of dendritic cells. Pharmacol. Res. 2016, 110, 111–121. [Google Scholar] [CrossRef]
- Li, D.; Wu, L. Coumarins from the roots of Angelica dahurica cause anti-allergic inflammation. Exp. Ther. Med. 2017, 14, 874–880. [Google Scholar] [CrossRef] [Green Version]
- Halwani, R.; Vazquez-Tello, A.; Sumi, Y.; Pureza, M.A.; Bahammam, A.; Al-Jahdali, H.; Soussi-Gounni, A.; Mahboub, B.; Al-Muhsen, S.; Hamid, Q. Eosinophils induce airway smooth muscle cell proliferation. J. Clin. Immunol. 2013, 33, 595–604. [Google Scholar] [CrossRef]
- Truyen, E.; Coteur, L.; Dilissen, E.; Overbergh, L.; Dupont, L.J.; Ceuppens, J.L.; Bullens, D.M. Evaluation of airway inflammation by quantitative Th1/Th2 cytokine mRNA measurement in sputum of asthma patients. Thorax 2006, 61, 202–208. [Google Scholar] [CrossRef] [Green Version]
- Matucci, A.; Vultaggio, A.; Maggi, E.; Kasujee, I. Is IgE or eosinophils the key player in allergic asthma pathogenesis? Are we asking the right question? Respir. Res. 2018, 19, 113. [Google Scholar] [CrossRef] [Green Version]
- Ohshima, Y.; Yamada, A.; Hiraoka, M.; Katamura, K.; Ito, S.; Hirao, T.; Akutagawa, H.; Kondo, N.; Morikawa, A.; Mayumi, M. Early sensitization to house dust mite is a major risk factor for subsequent development of bronchial asthma in Japanese infants with atopic dermatitis: Results of a 4-year followup study. Ann. Allergy Asthma Immunol. Off. Publ. Am. Coll. Allergy Asthma Immunol. 2002, 89, 265–270. [Google Scholar] [CrossRef]
- Xian, Z.; Jin, G.; Li, H.; Jiang, J.; Wang, C.; Zhu, L.; Jin, Z.; Li, L.; Piao, H.; Zheng, M.; et al. Imperatorin Suppresses Anaphylactic Reaction and IgE-Mediated Allergic Responses by Inhibiting Multiple Steps of FceRI Signaling in Mast Cells: IMP Alleviates Allergic Responses in PCA. Biomed. Res. Int. 2019, 2019, 7823761. [Google Scholar] [CrossRef] [PubMed]
- Sehra, S.; Pynaert, G.; Tournoy, K.; Haegeman, A.; Matthys, P.; Tagawa, Y.; Pauwels, R.; Grooten, J. Airway IgG counteracts specific and bystander allergen-triggered pulmonary inflammation by a mechanism dependent on Fc gamma R and IFN-gamma. J. Immunol. 2003, 171, 2080–2089. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jurado, A.; Carballido, J.; Griffel, H.; Hochkeppel, H.K.; Wetzel, G.D. The immunomodulatory effects of interferon-gamma on mature B-lymphocyte responses. Experientia 1989, 45, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Bossie, A.; Vitetta, E.S. IFN-gamma enhances secretion of IgG2a from IgG2a-committed LPS-stimulated murine B cells: Implications for the role of IFN-gamma in class switching. Cell. Immunol. 1991, 135, 95–104. [Google Scholar] [CrossRef]
- Platts-Mills, T.A.E. The Role of Immunoglobulin E in Allergy and Asthma. Am. J. Respir. Crit. Care Med. 2001, 164, S1–S5. [Google Scholar] [CrossRef]
- Epstein, M.M. Targeting memory Th2 cells for the treatment of allergic asthma. Pharmacol. Ther. 2006, 109, 107–136. [Google Scholar] [CrossRef]
- Renauld, J.C. New insights into the role of cytokines in asthma. J. Clin. Pathol. 2001, 54, 577–589. [Google Scholar] [CrossRef] [Green Version]
- Racke, M.K.; Bonomo, A.; Scott, D.E.; Cannella, B.; Levine, A.; Raine, C.S.; Shevach, E.M.; Röcken, M. Cytokine-induced immune deviation as a therapy for inflammatory autoimmune disease. J. Exp. Med. 1994, 180, 1961–1966. [Google Scholar] [CrossRef] [Green Version]
- Chung, F. Anti-inflammatory cytokines in asthma and allergy: Interleukin-10, interleukin-12, interferon-gamma. Mediat. Inflamm. 2001, 10, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, A.A.; Gawali, N.B.; Shinde, P.; Munshi, R.; Juvekar, A.R. Imperatorin ameliorates lipopolysaccharide induced memory deficit by mitigating proinflammatory cytokines, oxidative stress and modulating brain-derived neurotropic factor. Cytokine 2018, 110, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.C.; Liu, K.F.; Liu, P.C.; Ho, Y.T.; Li, W.S.; Peng, W.H.; Tsai, J.C. Comparing the Protection Imparted by Different Fraction Extracts of Garlic (Allium sativum L.) against Der p-Induced Allergic Airway Inflammation in Mice. Int. J. Mol. Sci. 2019, 20, 4879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.Y.; Lee, S.Y.; Park, S.Y.; Choi, H.J.; Kim, E.G.; Han, J.S. Therapeutic potential of a phospholipase D1 inhibitory peptide fused with a cell-penetrating peptide as a novel anti-asthmatic drug in a Der f 2-induced airway inflammation model. Exp. Mol. Med. 2018, 50, 1–11. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organ | Histopathological Findings | Groups 1 | ||||
|---|---|---|---|---|---|---|
| Control | Der p | Der p | ||||
| Imperatorin (1 mg/kg) | Imperatorin (5 mg/kg) | Imperatorin (10 mg/kg) | ||||
| Lung | Inflammation, eosinophilic and lymphocytic cells, perivascular and per bronchial, focal | 0.0 ± 0.00 | 3.8 ± 0.43 # | 3.5 ± 0.50 | 2.5 ± 0.50 | 1.8 ± 0.43 * |
| Aggregation, macrophage and giant cells, alveolar, focal | 0.0 ± 0.00 | 3.5 ± 0.50 # | 3.3 ± 0.43 | 2.3 ± 0.43 | 1.5 ± 0.50 * | |
| Epithelial hyperplasia, bronchial, focal | 0.0 ± 0.00 | 3.8 ± 0.43 # | 3.3± 0.83 | 2.5 ± 0.50 * | 2.3 ± 0.43 * | |
| Mucification, goblet, bronchial, focal | 0.0 ± 0.00 | 3.0 ± 0.00 # | 2.8 ± 0.43 | 2.3 ± 0.43 * | 1.5 ± 0.50 * | |
| Bronchial airways | Inflammation, submucosal focal | 0.0 ± 0.00 | 3.6 ± 0.41 # | 3.8 ± 0.49 | 3.2 ± 0.43 | 2.5 ± 0.32 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsieh, C.-C.; Ng, Y.-Y.; Li, W.-S.; Tzeng, C.-Y.; Hsu, T.-Y.; Huang, W.-H.; Tsai, J.-C. Ameliorative Effect of Imperatorin on Dermatophagoides pteronyssinus-Induced Allergic Asthma by Suppressing the Th2 Response in Mice. Molecules 2022, 27, 7028. https://doi.org/10.3390/molecules27207028
Hsieh C-C, Ng Y-Y, Li W-S, Tzeng C-Y, Hsu T-Y, Huang W-H, Tsai J-C. Ameliorative Effect of Imperatorin on Dermatophagoides pteronyssinus-Induced Allergic Asthma by Suppressing the Th2 Response in Mice. Molecules. 2022; 27(20):7028. https://doi.org/10.3390/molecules27207028
Chicago/Turabian StyleHsieh, Chia-Chen, Yan-Yan Ng, Wei-Sung Li, Chung-Yuh Tzeng, Tsai-Yi Hsu, Wan-Hsiang Huang, and Jen-Chieh Tsai. 2022. "Ameliorative Effect of Imperatorin on Dermatophagoides pteronyssinus-Induced Allergic Asthma by Suppressing the Th2 Response in Mice" Molecules 27, no. 20: 7028. https://doi.org/10.3390/molecules27207028