
The Potential Neuroprotective Effect of Cyperus esculentus L. Extract in Scopolamine-Induced Cognitive Impairment in Rats: Extensive Biological and Metabolomics Approaches

 ,
,  , , ,
, , ,  , ,
, ,  ,
,  and
and
Abstract
:1. Introduction
2. Results
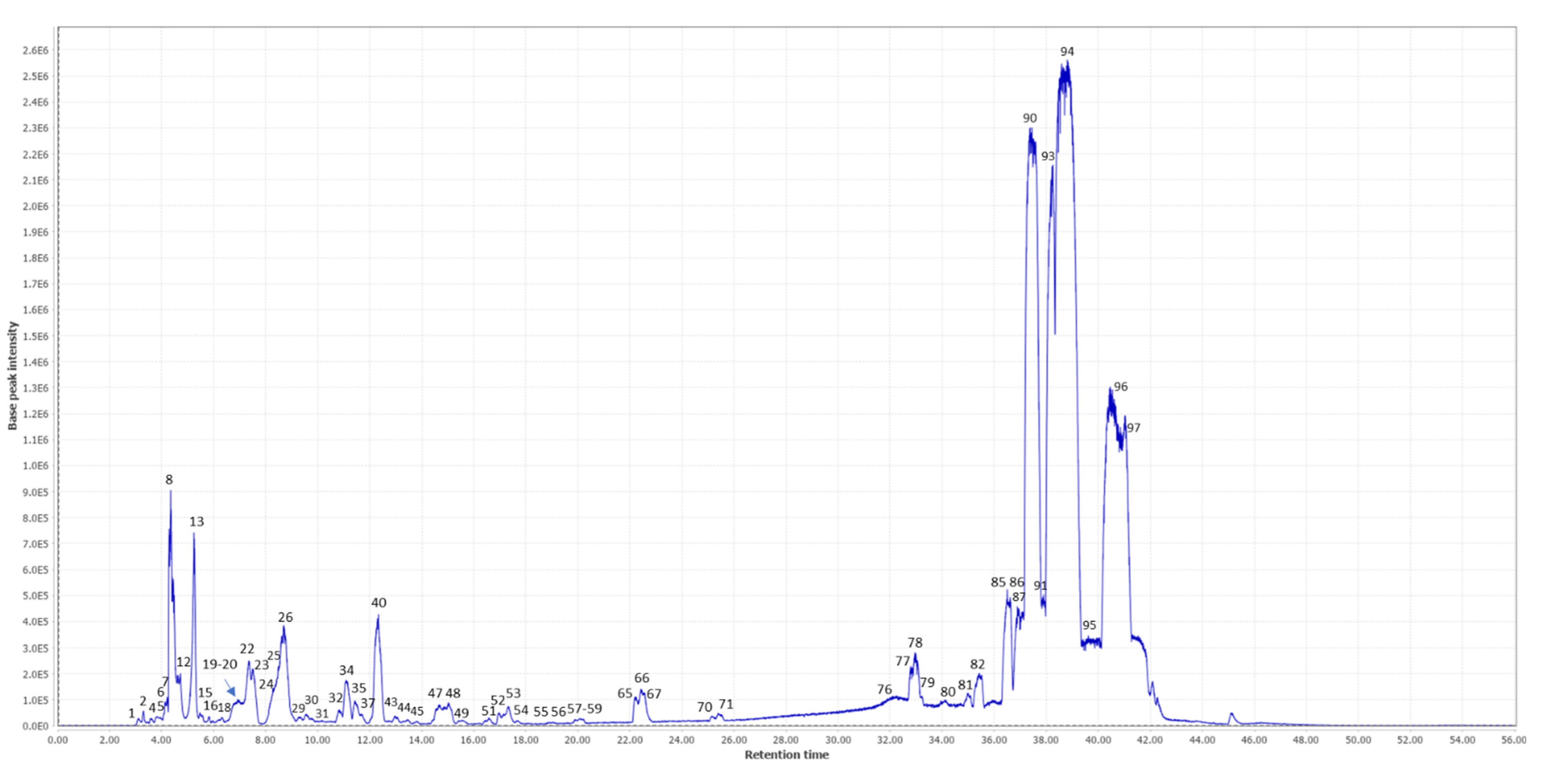
2.1. Metabolite Profiling Using LC/MS

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N ° | RT | m/z Experimental | m/z Theorical | Error (ppm) | Formula | Level of Annotation | Compounds | MS/MS Fragments | Rel. Ab. (%) | REF |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 3.11 | 201.0249 | 201.0260 | 5.47 | C5H6N4O5 | 2 | 2-Oxo-4-hydroxy-4 carboxy-5-ureidoimidazoline | 59/157 | 0.07 | FDB001617 |
| 2 | 3.31 | 141.0164 | 141.0188 | 17.0 | C6H6O4 | 2 | Kojic acid | 59/141 | 0.09 | HMDB32923 |
| 3 | 3.52 | 131.0821 | 131.0821 | 0 | C5H12N2O2 | 2 | L-ornithine | 70 | 0.03 | [32] |
| 4 | 3.80 | 173.1040 | 173.1039 | 0.58 | C6H14N4O2 | 2 | Arginine | 131 | 0.1 | [33,34] |
| 5 | 3.82 | 195.0502 | 195.0505 | 1.53 | C6H12O7 | 2 | Gluconic acid | 75/129/195 | 0.14 | FDB001980 |
| 6 | 4.22 | 267.0717 | 267.0716 | 0.37 | C9H16O9 | 3 | xylo-manno-nononic acid ɣ-lactone | 267 | 0.21 | - |
| 7 | 4.25 | 191.0553 | 191.0556 | 1.57 | C7H12O6 | 2 | Quinic acid | 85/92/191 | 0.06 | [35] |
| 8 | 4.36 | 341.1082 | 341.1084 | 0.59 | C12H22O11 | 2 | Galactinol dihydrate | 59/71/89/101/113/143/161 | 2.24 | [32] |
| 9 | 4.42 | 503.1607 | 503.1612 | 0.99 | C18H32O16 | 3 | Trisaccharide (raffinose) | 503 | 0.17 | [32], HMDB03213 |
| 10 | 4.50 | 337.0769 | 337.0771 | 0.59 | C12H18O11 | 2 | Ascorbyl glucoside isomer I | 59/161/277/289 | 0.03 | HMDB0253873 |
| 11 | 4.61 | 115.0031 | 115.0031 | 0 | C4H4O4 | 2 | Fumaric acid | 71/87/99 | 0.02 | [35] |
| 12 | 4.72 | 341.1082 | 341.1084 | 0.59 | C12H22O11 | 2 | Sucrose | 59/71/89/101/113/143/161 | 0.35 | [32] |
| 13 | 5.22 | 341.1082 | 341.1084 | 0.59 | C12H22O11 | 2 | Trehalose/maltose | 129/143/161/179 | 1.37 | [32] |
| 14 | 5.30 | 133.0135 | 133.0137 | 1.50 | C4H6O5 | 2 | Malic acid | 71/89/115/133 | 0.06 | [35] |
| 15 | 5.47 | 503.1607 | 503.1612 | 0.99 | C18H32O16 | 3 | Trisaccharide | 503 | 0.12 | [32] |
| 16 | 5.82 | 503.1597 | 503.1612 | 2.98 | C18H32O16 | 3 | Trisaccharide | 503 | 0.05 | [32] |
| 17 | 5.97 | 337.0763 | 337.0771 | 2.37 | C12H18O11 | 2 | Ascorbyl glucoside isomer II | 59/161/277/289 | 0.03 | HMDB0253873 |
| 18 | 6.32 | 251.0765 | 251.0780 | 5.97 | C10H12N4O4 | 4 | Unknown | 251 | 0.11 | - |
| 19 | 6.61 | 191.0189 | 192.0270 | 0.60 | C6H8O7 | 2 | Citric acid | 87 /111/129/173 | 0.03 | [36] |
| 20 | 6.76 | 295.1029 | 295.1029 | 0 | C11H20O9 | 3 | Aliphatic glucoside derivative | 295 | 0.11 | - |
| 21 | 7.31 | 243.0621 | 243.0617 | 1.64 | C9H12N2O6 | 2 | Uridine | 140/152/200 | 0.05 | [37] |
| 22 | 7.35 | 369.1404 | 369.1397 | 1.89 | C14H26O11 | 3 | Amylose | 369 | 1 | HMDB03403 |
| 23 | 7.56 | 295.1036 | 295.1029 | 2.37 | C11H20O9 | 3 | Aliphatic glucoside derivative | 295 | 0.07 | - |
| 24 | 8.15 | 130.0868 | 130.0868 | 0 | C6H13NO2 | 3 | Leucine | 130 | 1.10 | [32,33] |
| 25 | 8.34 | 329.0871 | 329.0873 | 0.60 | C14H18O9 | 2 | Dihydroxy benzoic acid methyl ester hexoside | 125/153/167/270 | 0.06 | [35] |
| 26 | 8.69 | 413.1654 | 413.1659 | 1.21 | C16H30O12 | 3 | Glucopyranoside derivate | 413 | 1.42 | - |
| 27 | 8.68 | 282.0843 | 282.0838 | 1.77 | C10H13N5O5 | 2 | Guanosine | 133/150 | 0.03 | FDB003632 |
| 28 | 8.95 | 493.1546 | 493.1557 | 2.23 | C20H30O14 | 2 | O-hexosyl-O-methyl-myoinositol-dihydroxy benzoic acid | 137/167/209/243/293/331 | 0.11 | [35] |
| 29 | 9.28 | 295.1027 | 295.1029 | 0.67 | C11H20O9 | 3 | Aliphatic glucoside derivative | 295 | 0.11 | - |
| 30 | 9.54 | 383.1550 | 383.1553 | 0.78 | C15H28O11 | 2 | Butanediol apiosylglucoside | 71/89/161 | 0.15 | HMDB0033063 |
| 31 | 10.53 | 164.0710 | 164.0712 | 1.22 | C9H11NO2 | 2 | Phenylalanine | 103/147/164 | 0.03 | [32,33] |
| 32 | 10.84 | 380.1545 | 380.1557 | 3.15 | C15H27NO10 | 4 | Unknown | 380 | 0.2 | - |
| 33 | 10.89 | 218.1026 | 218.1028 | 0.91 | C9H17NO5 | 2 | Pantothenic acid | 71/88/146 | 0.05 | FDB008322 |
| 34 | 11.09 | 559.2228 | 559.2238 | 1.78 | C22H40O16 | 3 | Trisaccharide derivative | 218 | 0.62 | - |
| 35 | 11.44 | 461.1652 | 461.1659 | 1.51 | C20H30O12 | 2 | Verbascoside | 119/137/299 | 0.32 | FDB018766 |
| 36 | 11.56 | 279.1078 | 279.1080 | 0.71 | C11H20O8 | 3 | Methyl glucopyranosyloxy butanoate | 279 | 0.16 | - |
| 37 | 11.65 | 309.1186 | 309.1186 | 0 | C12H22O9 | 3 | Dideoxy-glucopyranosyl-ribohexose | 309 | 0.15 | - |
| 38 | 11.84 | 359.0973 | 359.0978 | 1.39 | C15H20O10 | 2 | Glucosyringic acid | 153/197/315/341 | 0.03 | HMDB0303364 |
| 39 | 12.10 | 397.1657 | 397.1651 | 1.51 | C23H26O6 | 2 | Kanzonol M | 176/161/181 | 0.02 | HMDB0041101 |
| 40 | 12.34 | 397.1705 | 397.1710 | 1.26 | C16H30O11 | 3 | Glucopyranoside derivate | 397 | 1.37 | - |
| 41 | 12.35 | 203.0823 | 203.0821 | 0.98 | C11H12N2O2 | 2 | Tryptophan | 116/142 | 0.06 | [32] |
| 42 | 12.94 | 193.0504 | 193.0501 | 1.55 | C10H10O4 | 2 | Ferulic acid | 107/134/149 | 0.05 | [35,38] |
| 43 | 12.98 | 503.1398 | 503.1401 | 0.59 | C21H28O14 | 2 | 6-Caffeoylsucrose | 149/161/179/323/341/443 | 0.12 | FDB014172 |
| 44 | 13.45 | 353.0865 | 353.0873 | 2.26 | C16H18O9 | 2 | Chlorogenic acid | 127/135/191 | 0.04 | FDB002582 [37,39] |
| 45 | 13.81 | 597.2177 | 597.2183 | 1.00 | C28H38O14 | 4 | Unknown | 597 | 0.05 | - |
| 46 | 14.55 | 323.1337 | 323.1342 | 1.54 | C13H24O9 | 4 | Unknown disaccharide | 323 | 0.12 | - |
| 47 | 14.67 | 293.1232 | 293.1236 | 1.36 | C12H22O8 | 2 | Ethyl-glucopyranosyl-butanoate isomer I | 59/85/101/131 | 0.3 | HMDB0031693 |
| 48 | 15.02 | 293.1232 | 293.1236 | 1.36 | C12H22O8 | 2 | Ethyl-glucopyranosyl-butanoate isomer II | 59/85/101/131 | 0.32 | HMDB0031693 |
| 49 | 15.54 | 351.1286 | 351.1291 | 1.42 | C14H24O10 | 4 | Unknown disaccharide | 351 | 0.08 | - |
| 50 | 15.68 | 323.0976 | 323.0978 | 0.61 | C12H20O10 | 4 | Unknown disaccharide | 323 | 0.04 | - |
| 51 | 16.58 | 609.1462 | 609.1456 | 0.98 | C27H30O16 | 2 | Luteolin-3′,7-di-O-glucoside | 285/447 | 0.09 | [40] |
| 52 | 16.98 | 245.0923 | 245.0926 | 1.22 | C13H14N2O3 | 2 | cyclic 6-hydroxymelatonin | 74/116/142/159/203/245 | 0.1 | HMDB60810 |
| 53 | 17.32 | 245.0924 | 245.0926 | 0.81 | C13H14N2O3 | 2 | N-acetyl tryptophan | 74/116/142/159/203 | 0.23 | HMDB13713 |
| 54 | 17.67 | 683.1805 | 683.1823 | 2.63 | C30H36O18 | 2 | Rosmarinic acid di-O-hexoside | 359/521 | 0.07 | [41] |
| 55 | 19.05 | 461.1080 | 461.1084 | 0.86 | C22H22O11 | 2 | Kaempferide 7-glucoside | 283/269/299 | 0.05 | HMDB38455 |
| 56 | 19.35 | 285.0395 | 285.0399 | 1.40 | C15H10O6 | 3 | Aureusidin | 285 | 0.03 | [40] |
| 57 | 19.88 | 209.0792 | 209.0774 | 8.60 | C6H14N2O6 | 4 | Unknown | 209 | 0.04 | - |
| 58 | 20.06 | 209.0790 | 209.0787 | 1.43 | C7H10N6O2 | 4 | Unknown nitrogenous compound | 209 | 0.11 | - |
| 59 | 20.08 | 287.0560 | 287.0556 | 1.39 | C15H12O6 | 2 | Dihydrokaempferol | 107/135/151/175/229/243 | 0.06 | FDB012431 |
| 60 | 20.21 | 447.0923 | 447.0927 | 0.89 | C21H20O11 | 2 | Luteolin-O-glucoside | 285 | 0.04 | [40] |
| 61 | 20.94 | 299.0488 | 299.0556 | 22.7 | C16H12O6 | 3 | Luteolin methyl ether | 285 | 0.04 | HMDB37339 |
| 62 | 21.27 | 269.0396 | 269.0450 | 20.1 | C15H10O5 | 2 | Apigenin | 117/269 | 0.04 | [37,40] |
| 63 | 21.41 | 299.0556 | 299.0556 | 0 | C16H12O6 | 3 | Chrysoeriol | 284/299 | 0.02 | [34] |
| 64 | 21.81 | 271.0555 | 271.0606 | 18.8 | C15H12O5 | 2 | Naringenin | 107/119/151/177/217 | 0.04 | [34] |
| 65 | 22.22 | 285.0396 | 285.0399 | 1.05 | C15H10O6 | 3 | Kaempferol | 285 | 0.23 | [38] |
| 66 | 22.42 | 285.0397 | 285.0399 | 0.70 | C15H10O6 | 2 | Luteolin | 107/133/151/175 | 0.52 | [36,38,40] |
| 67 | 23.3 | 301.0714 | 301.0712 | 0.66 | C16H14O6 | 2 | Hesperitin | 135/151/285 | 0.03 | [42] |
| 68 | 24.21 | 299.0556 | 299.0556 | 0 | C16H12O6 | 2 | Kaempferide | 256/284 | 0.08 | [34,43] |
| 69 | 25.04 | 209.0815 | 209.0814 | 0.48 | C11H14O4 | 3 | Methylxanthoxylin | 209 | 0.05 | HMDB34047 |
| 70 | 25.39 | 299.0558 | 299.0556 | 0.69 | C16H12O6 | 2 | Isokaempferide | 183/227/255 | 0.17 | HMDB0302564 |
| 71 | 25.51 | 313.0713 | 313.0712 | 0.31 | C17H14O6 | 2 | Cirsimaritin | 283/297/313 | 0.11 | HMDB0250276 |
| 72 | 26.08 | 329.2324 | 329.2328 | 1.21 | C18H34O5 | 2 | Trihydroxy-octadecenoic acid | 171/211/229/285/311 | 0.2 | FDB002905 |
| 73 | 26.14 | 351.2144 | 351.2171 | 7.68 | C20H32O5 | 4 | Unknown | 351 | 0.03 | - |
| 74 | 28.63 | 373.1291 | 373.1287 | 1.07 | C20H22O7 | 3 | Isohydroxymatairesinol | 373 | 0.11 | HMDB0301737 |
| 75 | 28.66 | 339.1235 | 339.1232 | 0.88 | C20H20O5 | 3 | Prenylnaringenin | 339 | 0.09 | HMDB0247465 |
| 76 | 31.09 | 313.2380 | 313.2379 | 0.32 | C18H34O4 | 3 | Octadecanedioic acid | 313 | 0.05 | HMDB00782 |
| 77 | 32.44 | 315.2525 | 315.2535 | 3.17 | C18H36O4 | 3 | Dihydroxyoctadecanoic acid | 315 | 0.2 | HMDB31008 |
| 78 | 32.95 | 205.1593 | 205.1592 | 0.48 | C14H22O | 3 | 2,4-di-tert-butylphenol | 205 | 1.07 | HMDB13816 |
| 79 | 33.15 | 295.2272 | 295.2273 | 0.33 | C18H32O3 | 3 | Hydroxylinoleic acid | 277 | 0.25 | HMDB0247599 |
| 80 | 34.60 | 199.1698 | 199.1698 | 0 | C12H24O2 | 2 | Dodecanoic acid | 59/155 | 0.06 | FDB030978 |
| 81 | 34.98 | 299.2592 | 299.2586 | 2 | C18H36O3 | 2 | Hydroxyoctadecanoic acid | 255/269/281/299 | 0.52 | FDB006898 |
| 82 | 35.40 | 297.2432 | 297.2430 | 0.67 | C18H34O3 | 2 | Ricinoleic acid | 127/183/279 | 0.8 | FDB012640 |
| 83 | 36.03 | 255.2329 | 255.2324 | 1.96 | C16H32O2 | 3 | Isopalmitic acid | 255 | 0.18 | [32] |
| 84 | 36.27 | 281.2480 | 281.2481 | 0.35 | C18H34O2 | 3 | Elaidic acid | 281 | 0.17 | HMDB00573 |
| 85 | 36.50 | 277.2162 | 277.2168 | 2.16 | C18H30O2 | 2 | Linolenic Acid | 119 | 2.27 | [32] |
| 86 | 36.70 | 227.2007 | 227.2011 | 1.76 | C14H28O2 | 2 | Myristic acid | 209 | 0.77 | [32] |
| 87 | 36.89 | 253.2163 | 253.2168 | 1.97 | C16H30O2 | 2 | Palmitoleic acid | 71/253 | 1.89 | [32] |
| 88 | 36.99 | 327.2315 | 327.2324 | 2.75 | C22H32O2 | 2 | DHA | 229/283/309 | 0.92 | HMDB0244316 |
| 89 | 37.38 | 581.4541 | 581.4570 | 4.99 | C38H62O4 | 2 | Oxygenated fatty acid derivatives | 253/271 | 0.87 | - |
| 90 | 37.45 | 279.2337 | 279.2324 | 4.65 | C18H32O2 | 2 | Linoleic acid | 71/261/279 | 17 | [32] |
| 91 | 37.95 | 267.2325 | 267.2324 | 0.37 | C17H32O2 | 3 | Heptadecenoic acid | 267 | 0.35 | HMDB31046 |
| 92 | 38.22 | 533.4538 | 533.4570 | 5.99 | C34H62O4 | 2 | Oxygenated fatty acid derivatives | 293/533 | 0.77 | - |
| 93 | 38.26 | 255.2372 | 255.2324 | 18.8 | C16H32O2 | 2 | Palmitic acid | 237 | 12 | [32] |
| 94 | 38.32 | 281.2480 | 281.2481 | 0.35 | C18H34O2 | 2 | Oleic acid | 253/255/267 | 29.9 | [32] |
| 95 | 39.40 | 269.2483 | 269.2481 | 0.74 | C17H34O2 | 3 | Margaric acid | 269 | 0.25 | [32] |
| 96 | 40.46 | 283.2640 | 283.2637 | 1.05 | C18H36O2 | 2 | Stearic acid | 265 | 14.5 | [32] |
| 97 | 41.47 | 309.2792 | 309.2794 | 0.65 | C20H38O2 | 3 | Gondoic acid | 309 | 0.32 | [32] |
2.2. Effect of Different Doses of TN on Scop-Induced Behavioral Changes in Rats
2.3. Effect of Different Doses of TN on Scop-Induced Alterations in AChE Activity
2.4. Effect of Different Doses of TN on Scop-Induced Oxidative Stress
2.5. Effect of Different Doses of TN on Scop-Induced Neuroinflammation
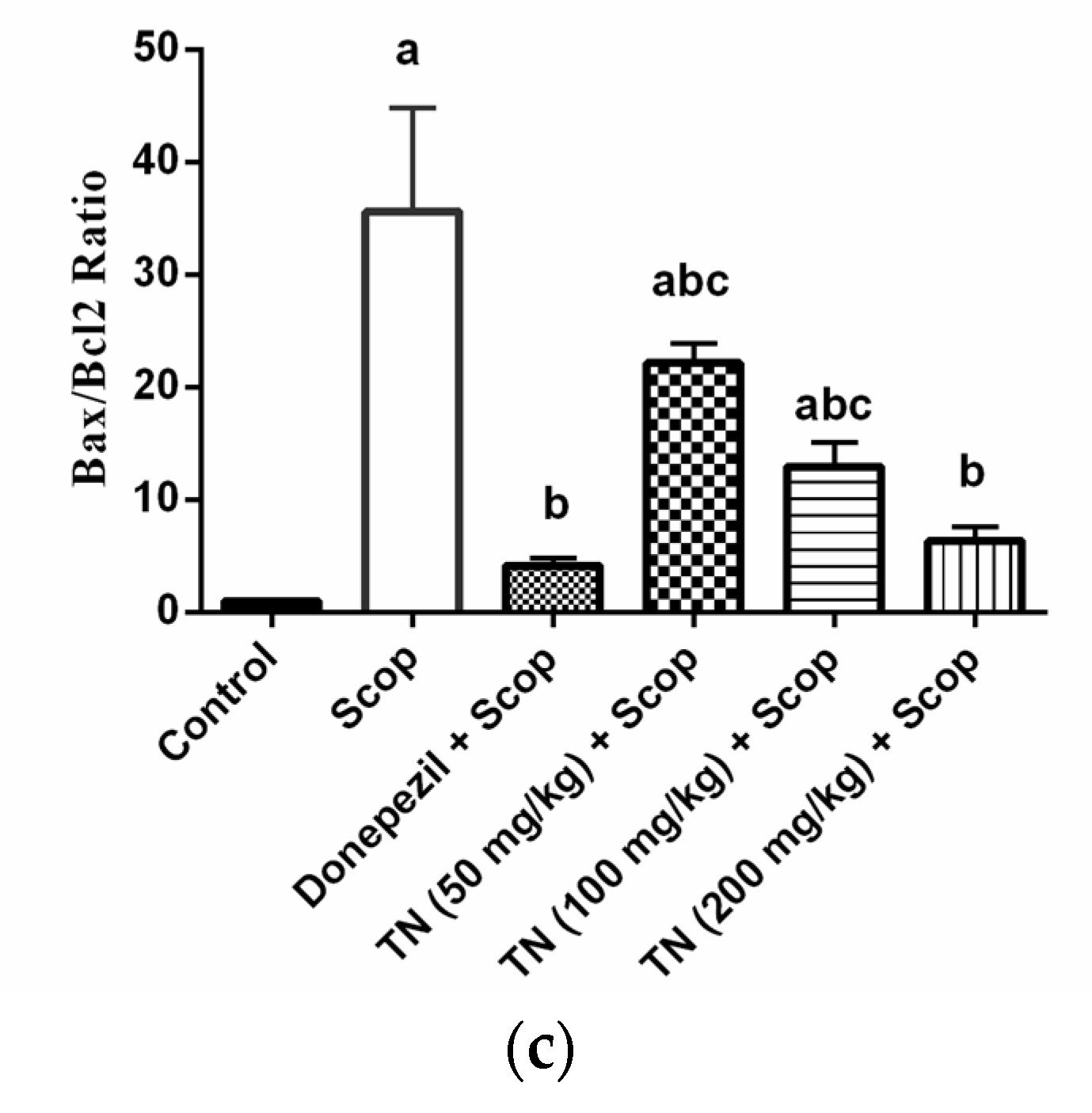
2.6. Effect of Different Doses of TN on Scop-Induced Apoptosis
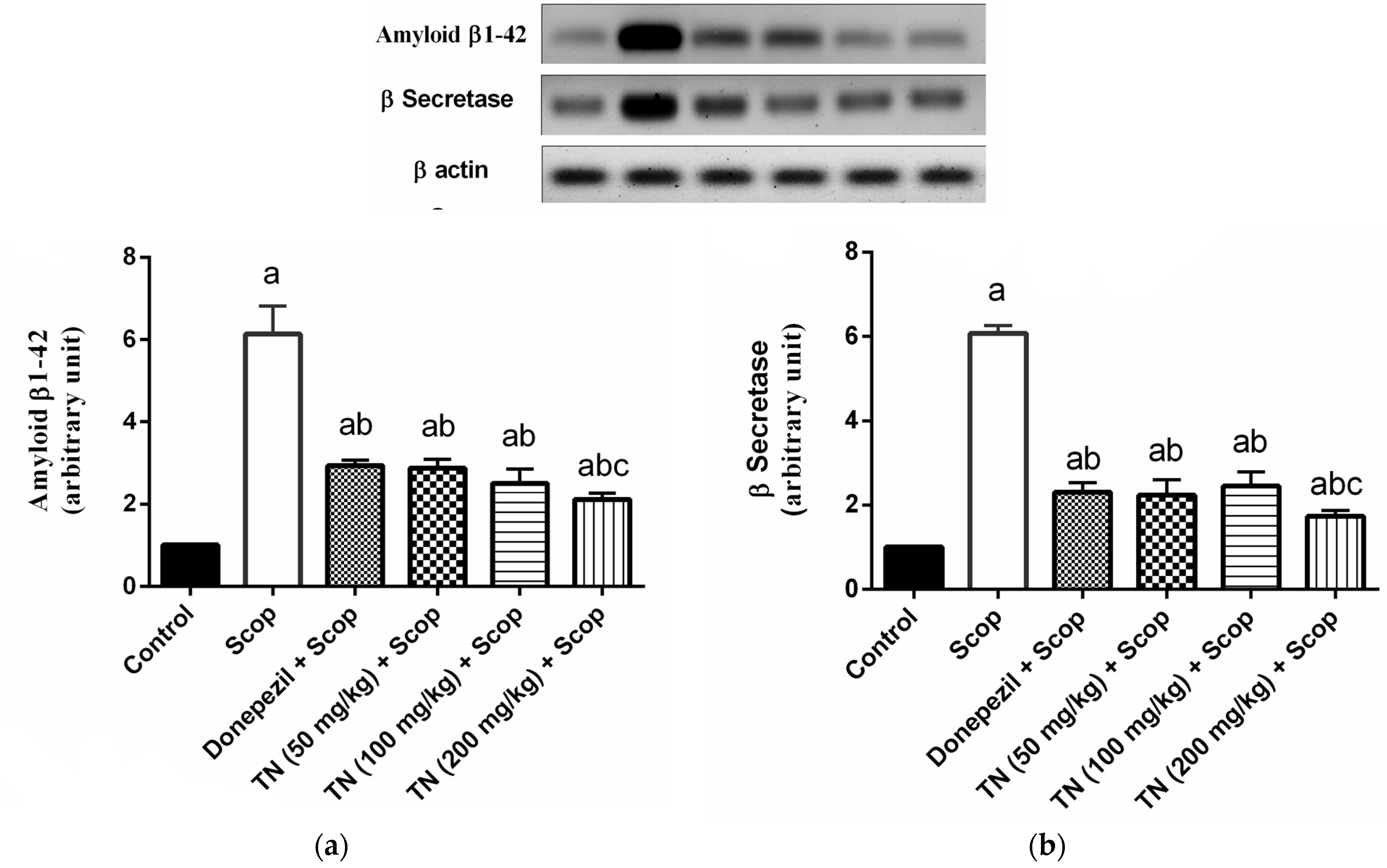
2.7. Effect of Different Doses of TN on Scop-Induced Alterations in Aβ and β-Secretase Protein Expression
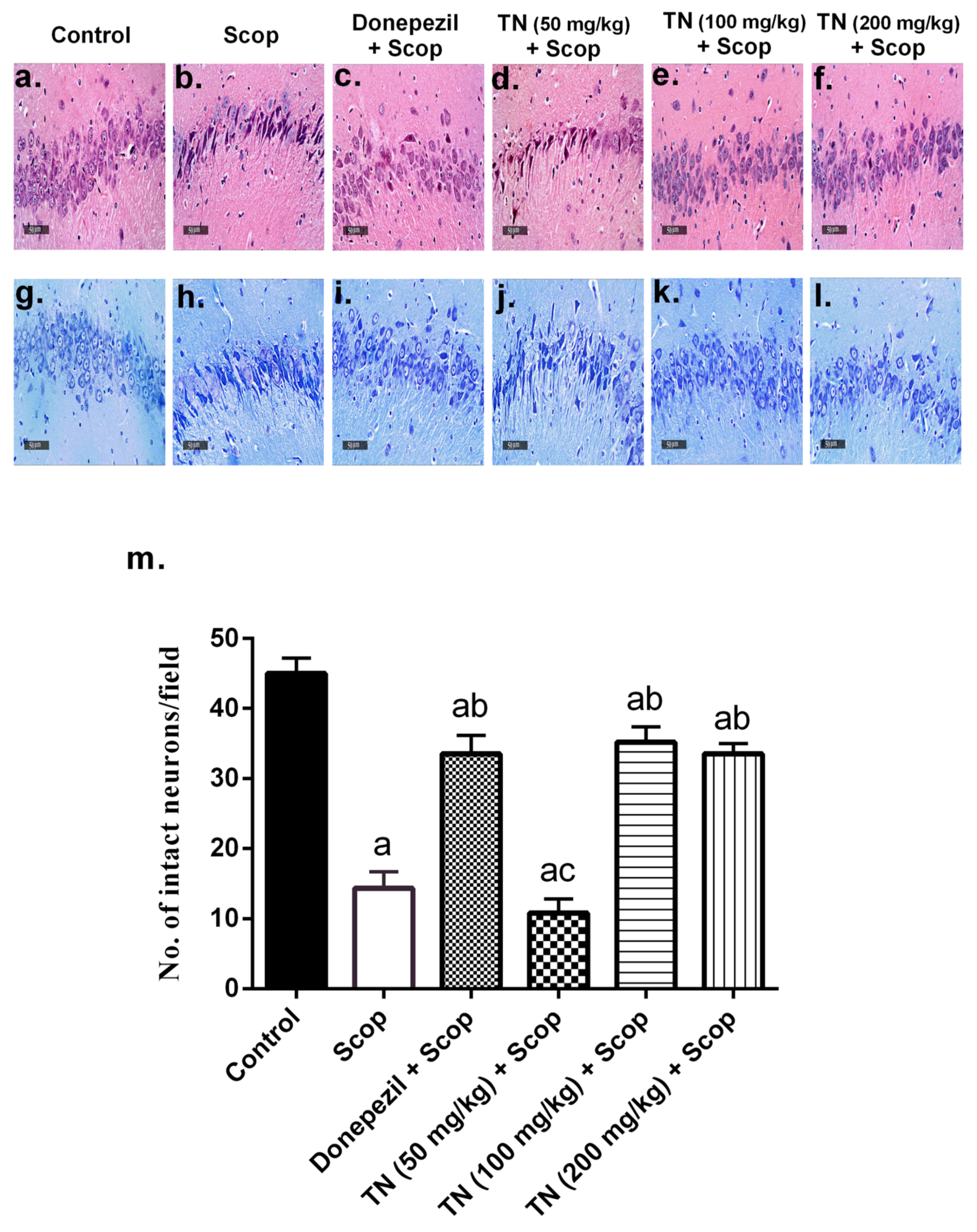
2.8. Effect of Different Doses of TN on Scop-Induced Histopathological Alterations and Neuronal Loss
3. Discussion
4. Materials and Methods
4.1. Plant Material and Extraction
4.2. UHPLC-ESI-QTOF-MS Profiling
4.3. UHPLC-ESI-QTOF-MS Data Processing
4.4. Biological Study
4.4.1. Animals
4.4.2. Drugs and Chemicals
4.4.3. Experimental Design
4.4.4. Behavioral Assessments
Morris Water Maze Test
Y-Maze Test
4.4.5. Brain Processing
4.4.6. Biochemical Measurements
Acetylcholinesterase Activity
Determination of Oxidative Stress Biomarkers
Enzyme-Linked Immunosorbent Assay
Quantitative Real-Time Polymerase Chain Reaction
Western Blot Analysis
4.4.7. Histopathological Examination
4.4.8. Statistical Analysis
5. Conclusions
Abbreviations
| Aβ. | amyloid-beta |
| AD | Alzheimer’s disease |
| AChE | acetylcholinestrase |
| ALA | alpha-linolenic acid |
| Bax | Bcl2-associated X protein |
| Bcl-2 | B-cell lymphoma 2 |
| CAT | catalase |
| DHA | docosahexaenoic acid |
| DTNB | 5,5-dithiobis 2-nitrobenzoic acid |
| EPA | eicosapentaenoic acid |
| GSH | glutathione |
| GSK-3β | glycogen synthase kinase-3β |
| IL-1β | interleukin 1 beta |
| LC-PUFA | long chain polyunsaturated fatty acids |
| MAPK | mitogen-activated protein kinase |
| MDA | malodialdehyde |
| MWM | Morris water maze |
| PI3K/AKT | phosphatidylinositol-3-kinase/protein kinase B |
| Scop | scopolamine |
| SOD | superoxide dismutase |
| TN | tiger nut |
| TNF-α | tumor necrosis factor alpha |
| UHPLC-ESI-QTOF-MS | ultra performance liquid chromatography with electrospray ionization and quadrupole time-of-flight mass spectrometry |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Fawzi, S.F.; Menze, E.T.; Tadros, M.G. Deferiprone ameliorates memory impairment in Scopolamine-treated rats: The impact of its iron-chelating effect on beta-amyloid disposition. Behav. Brain. Res. 2020, 378, 112314. [Google Scholar] [CrossRef] [PubMed]
- Thakur, A.K.; Kamboj, P.; Goswami, K.; Ahuja, K.J.J.A.P.R. Pathophysiology and management of Alzheimer’s disease: An overview. J. Anal. Pharm. Res 2018, 9, 226–235. [Google Scholar]
- Tang, K.S. The cellular and molecular processes associated with scopolamine-induced memory deficit: A model of Alzheimer’s biomarkers. Life Sci. 2019, 233, 116695. [Google Scholar] [CrossRef]
- Aisen, P.S.; Cummings, J.; Jack, C.R.; Morris, J.C.; Sperling, R.; Frölich, L.; Jones, R.W.; Dowsett, S.A.; Matthews, B.R.; Raskin, J.J.A.s.r.; et al. On the path to 2025: Understanding the Alzheimer’s disease continuum. Alzheimer’s Res. Ther. 2017, 9, 60. [Google Scholar]
- Tiwari, S.; Soni, R.J.J.A.D.P. Alzheimer’s disease pathology and oxidative stress: Possible therapeutic options. J. Alzheimers. Dis Park. 2014, 4, 162. [Google Scholar] [CrossRef] [Green Version]
- Shabani, S.; Mirshekar, M.A. Diosmin is neuroprotective in a rat model of scopolamine-induced cognitive impairment. Biomed. Pharmacother. 2018, 108, 1376–1383. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Hu, J.; Li, J.; Yang, Z.; Xin, X.; Wang, J.; Ding, J.; Geng, M. Effect of acidic oligosaccharide sugar chain on scopolamine-induced memory impairment in rats and its related mechanisms. Neurosci. Lett. 2005, 374, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Singh, A.; Ekavali. A review on Alzheimer’s disease pathophysiology and its management: An update. Pharmacol. Rep. 2015, 67, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Gambo, A.; Da’u, A. Tiger nut (Cyperus esculentus): Composition, products, uses and health benefits-a review. Bayero. J. Pure. Appl. Sci. 2014, 7, 56–61. [Google Scholar] [CrossRef] [Green Version]
- Oderinde, R.; Tairu, O. Evaluation of the properties of yellow nutsedge (Cyperus esculentus) tuber oil. Food Chem. 1988, 28, 233–237. [Google Scholar] [CrossRef]
- Ejoh, R.A.; Djomdi; Ndjouenkeu, R. Characteristics of tigernut (Cyperus esculentus) tubers and their performance in the production of a milky drink. J. Food Process. Preserv. 2006, 30, 145–163. [Google Scholar] [CrossRef]
- Tackholm, V. Students’ flora of Egypt, 2nd ed.; Cairo University Press: Cairo, Egypt, 1974. [Google Scholar]
- Obadina, A.; Oyawole, O.; Ayoola, A. Quality assessment of gari produced using rotary drier. In Food Processing: Methods, Techniques and Trends; VC, B., Ed.; Nova Science Publishers: New York, NY, USA, 2008. [Google Scholar]
- Arafat, S.M.; Gaafar, A.M.; Basuny, A.M.; Nassef, S.L. Chufa tubers (Cyperus esculentus L.): As a new source of food. World Appl. Sci. J. 2009, 7, 151–156. [Google Scholar]
- Yu, Y.; Lu, X.; Zhang, T.; Zhao, C.; Guan, S.; Pu, Y.; Gao, F. Tiger Nut (Cyperus esculentus L.): Nutrition, Processing, Function and Applications. Foods 2022, 11, 601. [Google Scholar] [CrossRef] [PubMed]
- Yeboah, S.O.; Mitei, Y.C.; Ngila, J.C.; Wessjohann, L.; Schmidt, J. Compositional and structural studies of the oils from two edible seeds: Tiger nut, Cyperus esculentum, and asiato, Pachira insignis, from Ghana. Food Res. Int. 2012, 47, 259–266. [Google Scholar] [CrossRef]
- Nofouzi, K.; Mahmudi, R.; Tahapour, K.; Amini, E.; Yousefi, K. Verbascum speciosum methanolic extract: Phytochemical components and antibacterial properties. J. Essent. Oil Bear Plants 2016, 19, 499–505. [Google Scholar] [CrossRef]
- Allahyari, S.; Pakbin, B.; Amani, Z.; Mahmoudi, R.; Hamidiyan, G.; Peymani, A.; Qajarbeygi, P.; Mousavi, S. Antiviral activity of Phoenix dactylifera extracts against herpes simplex virus type 1: An animal study. Comp. Clin. Pathol. 2021, 30, 945–951. [Google Scholar] [CrossRef]
- Nwosu, L.C.; Edo, G.I.; Ozgor, E. The phytochemical, proximate, pharmacological, GC-MS analysis of Cyperus esculentus (Tiger nut): A fully validated approach in health, food and nutrition. Food Biosci. 2022, 46, 101551. [Google Scholar] [CrossRef]
- Sánchez-Zapata, E.; Fernández-López, J.; Angel Pérez-Alvarez, J. Tiger nut (Cyperus esculentus) commercialization: Health aspects, composition, properties, and food applications. Compr. Rev. Food Sci. Food Saf. 2012, 11, 366–377. [Google Scholar] [CrossRef]
- Abimbade, S.F.; Oloyede, G.K.; Nwabueze, C.C. Antioxidant and toxicity screenings of extracts obtained from Cyperus esculentus. Acad. Arena 2014, 6, 77–83. [Google Scholar]
- Saber, F.R.; Mahrous, E.A. Novel Functional Foods From Plants of the Mediterranean Area: Biological, Chemical, Metabolomic Approaches. In Reference Module in Food Science; Ferranti, P., Ed.; Elsevier: Amsterdam, The Netherlands, In Press. [CrossRef]
- Sumner, L.W.; Amberg, A.; Barrett, D.; Beale, M.H.; Beger, R.; Daykin, C.A.; Fan, T.W.-M.; Fiehn, O.; Goodacre, R.; Griffin, J.L. Proposed minimum reporting standards for chemical analysis. Metabolomics 2007, 3, 211–221. [Google Scholar] [CrossRef] [Green Version]
- Ijarotimi, O.S.; Yinusa, M.A.; Adegbembo, P.A.; Adeniyi, M.D. Chemical compositions, functional properties, antioxidative activities, and glycaemic indices of raw and fermented tigernut tubers (Cyperus esculentus Lativum) flour. J. Food Biochem. 2018, 42, e12591. [Google Scholar] [CrossRef]
- Bazinet, R.P.; Layé, S. Polyunsaturated fatty acids and their metabolites in brain function and disease. Nat. Rev. Neurosci. 2014, 15, 771–785. [Google Scholar] [CrossRef]
- Song, J.; Kim, Y.-S.; Lee, D.H.; Lee, S.H.; Park, H.J.; Lee, D.; Kim, H. Neuroprotective effects of oleic acid in rodent models of cerebral ischaemia. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, M.; Sadiq, A.; Junaid, M.; Ullah, F.; Ovais, M.; Ullah, I.; Ahmed, J.; Shahid, M. Flavonoids as prospective neuroprotectants and their therapeutic propensity in aging associated neurological disorders. Front. Aging. Neurosci. 2019, 11, 155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kempuraj, D.; Thangavel, R.; Kempuraj, D.D.; Ahmed, M.E.; Selvakumar, G.P.; Raikwar, S.P.; Zaheer, S.A.; Iyer, S.S.; Govindarajan, R.; Chandrasekaran, P.N. Neuroprotective effects of flavone luteolin in neuroinflammation and neurotrauma. Biofactors 2021, 47, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Szwajgier, D.; Borowiec, K.; Pustelniak, K. The neuroprotective effects of phenolic acids: Molecular mechanism of action. Nutrients 2017, 9, 477. [Google Scholar] [CrossRef] [Green Version]
- Marim, F.M.; Teixeira, D.C.; Queiroz-Junior, C.M.; Valiate, B.V.S.; Alves-Filho, J.C.; Cunha, T.M.; Dantzer, R.; Teixeira, M.M.; Teixeira, A.L.; Costa, V.V. Inhibition of Tryptophan Catabolism Is Associated With Neuroprotection During Zika Virus Infection. Front. Immunol. 2021, 12, 702048. [Google Scholar] [CrossRef]
- Wang, J.; Song, Y.; Gao, M.; Bai, X.; Chen, Z. Neuroprotective effect of several phytochemicals and its potential application in the prevention of neurodegenerative diseases. Geriatrics 2016, 1, 29. [Google Scholar] [CrossRef] [Green Version]
- Aljuhaimi, F.; Şimşek, Ş.; Özcan, M.M. Comparison of chemical properties of taro (Colocasia esculenta L.) and tigernut (Cyperus esculentus) tuber and oils. J. Food Process Preserv. 2018, 42, e13534. [Google Scholar] [CrossRef]
- Bosch, L.; Alegria, A.; Farre, R. RP-HPLC determination of tiger nut and orgeat amino acid contents. Food Sci. Technol. Int. 2005, 11, 33–40. [Google Scholar] [CrossRef]
- Soto Mayer, L. Phytochemical Analysis of the methanolic extract of tigernut, tuber of Cyperus esculentus, by ultra-high performance liquid chromatography coupled with electrospray ionization-quadrupole-time of flight-mass spectrometry (UHPLC/ESI-Q.-TOF-MS). M.Sc. Thesis, Universidad CEU San pablo, Madrid, 2019. [Google Scholar]
- Abd-ElGawad, A.M.; Elshamy, A.I.; Al-Rowaily, S.L.; El-Amier, Y.A. Habitat Affects the Chemical Profile, Allelopathy, and Antioxidant Properties of Essential Oils and Phenolic Enriched Extracts of the Invasive Plant Heliotropium Curassavicum. Plants 2019, 8, 482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elshamy, A.I.; Farrag, A.R.H.; Ayoub, I.M.; Mahdy, K.A.; Taher, R.F.; Gendy, A.E.-N.G.; Mohamed, T.A.; Al-Rejaie, S.S.; Ei-Amier, Y.A.; Abd-EIGawad, A.M. UPLC-qTOF-MS phytochemical profile and antiulcer potential of Cyperus conglomeratus Rottb. alcoholic extract. Molecules 2020, 25, 4234. [Google Scholar] [CrossRef] [PubMed]
- Sayed, H.M.; Mohamed, M.H.; Farag, S.F.; Mohamed, G.A.; Omobuwajo, O.R.; Proksch, P. Fructose-amino acid conjugate and other constituents from Cyperus rotundus L. Nat. Prod. Res. 2008, 22, 1487–1497. [Google Scholar] [CrossRef] [PubMed]
- Sayed, H.M.; Mohamed, M.H.; Farag, S.F.; Mohamed, G.A. Phytochemical and biological investigations of Cyperus rotundus L. Bull. Facul.t Pharm. Cairo. Uni. 2001, 39, 195–203. [Google Scholar]
- Rocha, F.G.; de Mello Brandenburg, M.; Pawloski, P.L.; da Silva Soley, B.; Costa, S.C.A.; Meinerz, C.C.; Baretta, I.P.; Otuki, M.F.; Cabrini, D.A. Preclinical study of the topical anti-inflammatory activity of Cyperus rotundus L. extract (Cyperaceae) in models of skin inflammation. J. Ethnopharmacol. 2020, 254, 112709. [Google Scholar] [CrossRef] [PubMed]
- El-Habashy, I.; Mansour, R.; Zahran, M.; El-Hadidi, M.; Saleh, N. Leaf flavonoids of Cyperus species in Egypt. Biochem. Syst. Ecol. 1989, 17, 191–195. [Google Scholar] [CrossRef]
- Uysal, S.; Zengin, G.; Sinan, K.I.; Ak, G.; Ceylan, R.; Mahomoodally, M.F.; Uysal, A.; Sadeer, N.B.; Jekő, J.; Cziáky, Z. Chemical characterization, cytotoxic, antioxidant, antimicrobial, and enzyme inhibitory effects of different extracts from one sage (Salvia ceratophylla L.) from Turkey: Open a new window on industrial purposes. RSC Adv. 2021, 11, 5295–5310. [Google Scholar] [CrossRef]
- Allan, R.; Wells, R.; MacLeod, J. Flavanone quinones from Cyperus species. Tetrahedron Lett. 1973, 1, 7–8. [Google Scholar] [CrossRef]
- Farrag, A.R.H.; Abdallah, H.M.; Khattab, A.R.; Elshamy, A.I.; El Gendy, A.E.-N.G.; Mohamed, T.A.; Farag, M.A.; Efferth, T.; Hegazy, M.-E.F. Antiulcer activity of Cyperus alternifolius in relation to its UPLC-MS metabolite fingerprint: A mechanistic study. Phytomedicine 2019, 62, 152970. [Google Scholar] [CrossRef]
- Innih, S.O.; Eluehike, N.; Francis, B. Effects of aqueous extract of Cyperus esculentus (tiger nut) on antioxidant status and hematological indices in the heart of cadmium-induced wistar rats. Niger. J. Experiment. Clin. Biosci. 2021, 9, 17. [Google Scholar] [CrossRef]
- Sudha, T.S. Evaluation of anticonvulsant and antioxidant properties of Cyperus esculentus Linn. in various types of experimentally induced seizures in rats. Int. J. Green Pharm. 2021, 14(4), 381–387. [Google Scholar]
- Hussein, J.S.; Medhat, D.; Abdel-Latif, Y.; Morsy, S.; Gaafar, A.A.; Ibrahim, E.A.; Al-kashef, A.S.; Nooman, M.U. Amelioration of neurotoxicity induced by esfenvalerate: Impact of Cyperus rotundus L. tuber extract. Comparat Clin. Pathol. 2021, 30, 1–10. [Google Scholar] [CrossRef]
- Umukoro, S.; Okoh, L.; Igweze, S.C.; Ajayi, A.M.; Ben-Azu, B. Protective effect of Cyperus esculentus (tiger nut) extract against scopolamine-induced memory loss and oxidative stress in mouse brain. Drug Metab. Person. Ther. 2020, 35(3), 20200112. [Google Scholar] [CrossRef]
- El-Marasy, S.A.; Abd-Elsalam, R.M.; Ahmed-Farid, O.A. Ameliorative effect of silymarin on scopolamine-induced dementia in rats. Maced. Journal Med. Sci. 2018, 6, 1215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barai, P.; Raval, N.; Acharya, S.; Borisa, A.; Bhatt, H.; Acharya, N. Neuroprotective effects of bergenin in Alzheimer’s disease: Investigation through molecular docking, in vitro and in vivo studies. Behav. Brain. Res. 2019, 356, 18–40. [Google Scholar] [CrossRef]
- Kim, M.-S.; Lee, D.Y.; Lee, J.; Kim, H.W.; Sung, S.H.; Han, J.-S.; Jeon, W.K. Terminalia chebula extract prevents scopolamine-induced amnesia via cholinergic modulation and anti-oxidative effects in mice. BMC Complem Altern. Med. 2018, 18, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Tucker, L.B.; Velosky, A.G.; McCabe, J.T. Applications of the Morris water maze in translational traumatic brain injury research. Neurosci. Biobehav. Rev. 2018, 88, 187–200. [Google Scholar] [CrossRef] [PubMed]
- Birla, H.; Keswani, C.; Rai, S.N.; Singh, S.S.; Zahra, W.; Dilnashin, H.; Rathore, A.S.; Singh, S.P. Neuroprotective effects of Withania somnifera in BPA induced-cognitive dysfunction and oxidative stress in mice. Behav. Brain Funct. 2019, 15, 9. [Google Scholar] [CrossRef] [Green Version]
- Sarter, M.; Bodewitz, G.; Stephens, D.N. Attenuation of scopolamine-induced impairment of spontaneous alternation behaviour by antagonist but not inverse agonist and agonist β-carbolines. Psychopharmacology 1988, 94, 491–495. [Google Scholar] [CrossRef]
- Brinza, I.; Boiangiu, R.S.; Hancianu, M.; Cioanca, O.; Erdogan Orhan, I.; Hritcu, L. Bay Leaf (Laurus Nobilis, L.) Incense Improved Scopolamine-Induced Amnesic Rats by Restoring Cholinergic Dysfunction and Brain Antioxidant Status. Antioxidants 2021, 10, 259. [Google Scholar] [CrossRef]
- He, C.; Qu, X.; Cui, L.; Wang, J.; Kang, J.X. Improved spatial learning performance of fat-1 mice is associated with enhanced neurogenesis and neuritogenesis by docosahexaenoic acid. Proc. Natl. Acad. Sci. USA 2009, 106, 11370–11375. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Wang, H.; Cheng, H.; Che, Z. Ameliorating effect of luteolin on memory impairment in an Alzheimer’s disease model. Mol. Med. Rep. 2016, 13, 4215–4220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bakoyiannis, I.; Daskalopoulou, A.; Pergialiotis, V.; Perrea, D. Phytochemicals and cognitive health: Are flavonoids doing the trick? Biomed. Pharmacother. 2019, 109, 1488–1497. [Google Scholar] [CrossRef]
- Selkoe, D.J.; Hardy, J. The amyloid hypothesis of Alzheimer’s disease at 25 years. EMBO Mol. Med. 2016, 8, 595–608. [Google Scholar] [CrossRef]
- Wang, W.-Y.; Tan, M.-S.; Yu, J.-T.; Tan, L. Role of pro-inflammatory cytokines released from microglia in Alzheimer’s disease. Annal. Transl. Med. 2015, 3, 136. [Google Scholar]
- Das, H.; Sarkar, S.; Paidi, R.K.; Biswas, S.C. Subtle genomic DNA damage induces intraneuronal production of amyloid-β (1-42) by increasing β-secretase activity. FASEB J. 2021, 35, e21569. [Google Scholar] [CrossRef]
- Fourriere, L.; Gleeson, P.A. Amyloid β production along the neuronal secretory pathway: Dangerous liaisons in the Golgi? Traffic 2021, 22, 319–327. [Google Scholar] [CrossRef]
- Patel, P.; Shah, J.S. Effect of Vitamin D Supplementation on the Progression of Alzheimer’s Disease in Rats: A Mechanistic Approach. Res. Sq. Prepr. 2021. [Google Scholar] [CrossRef]
- Djeuzong, E.; Kandeda, A.K.; Djiogue, S.; Stéphanie, L.; Nguedia, D.; Ngueguim, F.; Djientcheu, J.P.; Kouamouo, J.; Dimo, T. Antiamnesic and Neuroprotective Effects of an Aqueous Extract of Ziziphus jujuba Mill.(Rhamnaceae) on Scopolamine-Induced Cognitive Impairments in Rats. Evid-Based Compl. Alt. Med. 2021, 2021. 5577163. [Google Scholar] [CrossRef] [PubMed]
- Kandeda, A.K.; Nguedia, D.; Ayissi, E.R.; Kouamouo, J.; Dimo, T. Ziziphus jujuba (Rhamnaceae) Alleviates Working Memory Impairment and Restores Neurochemical Alterations in the Prefrontal Cortex of D-Galactose-Treated Rats. Evid-Based Compl. Alt. Med. 2021, 2021. 6610864. [Google Scholar] [CrossRef] [PubMed]
- Ali, W.; Ikram, M.; Park, H.Y.; Jo, M.G.; Ullah, R.; Ahmad, S.; Abid, N.B.; Kim, M.O. Oral administration of alpha linoleic acid rescues Aβ-induced glia-mediated neuroinflammation and cognitive dysfunction in C57BL/6N mice. Cells 2020, 9, 667. [Google Scholar] [CrossRef] [Green Version]
- Baptista, F.I.; Henriques, A.G.; Silva, A.M.; Wiltfang, J.; da Cruz e Silva, O.A. Flavonoids as therapeutic compounds targeting key proteins involved in Alzheimer’s disease. ACS Chem. Neurosci. 2014, 5, 83–92. [Google Scholar] [CrossRef] [Green Version]
- Schroeter, H.; Boyd, C.; Spencer, J.P.; Williams, R.J.; Cadenas, E.; Rice-Evans, C. MAPK signaling in neurodegeneration: Influences of flavonoids and of nitric oxide. Neurobiol. Aging 2002, 23, 861–880. [Google Scholar] [CrossRef] [Green Version]
- Walker, E.H.; Pacold, M.E.; Perisic, O.; Stephens, L.; Hawkins, P.T.; Wymann, M.P.; Williams, R.L. Structural determinants of phosphoinositide 3-kinase inhibition by wortmannin, LY294002, quercetin, myricetin, and staurosporine. Mol. Cell 2000, 6, 909–919. [Google Scholar] [CrossRef]
- Baier, A.; Szyszka, R. Compounds from Natural Sources as Protein Kinase Inhibitors. Biomolecules 2020, 10, 1546. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, W.; Ijaz, B.; Shabbiri, K.; Ahmed, F.; Rehman, S. Oxidative toxicity in diabetes and Alzheimer’s disease: Mechanisms behind ROS/RNS generation. J. Biomed. Sci. Eng. 2017, 24, 1–10. [Google Scholar] [CrossRef]
- Adedayo, B.C.; Jesubowale, O.S.; Adebayo, A.A.; Oboh, G. Effect of Andrographis paniculata leaves extract on neurobehavioral and biochemical indices in scopolamine-induced amnesic rats. J. Food Biochem. 2021, 45, e13280. [Google Scholar] [CrossRef]
- Kouémou, N.E.; Taiwe, G.S.; Moto, F.C.; Pale, S.; Ngoupaye, G.T.; Njapdounke, J.S.; Nkantchoua, G.C.; Pahaye, D.B.; Bum, E.N. Nootropic and neuroprotective effects of Dichrocephala integrifolia on scopolamine mouse model of Alzheimer’s disease. Front. Pharmacol. 2017, 8, 847. [Google Scholar] [CrossRef] [Green Version]
- Sun, K.; Bai, Y.; Zhao, R.; Guo, Z.; Su, X.; Li, P.; Yang, P. Neuroprotective effects of matrine on scopolamine-induced amnesia via inhibition of AChE/BuChE and oxidative stress. Metab. Brain Dis. 2019, 34, 173–181. [Google Scholar] [CrossRef]
- Pattanashetti, L.A.; Patil, B.M.; Hegde, H.V.; Kangle, R.P. Potential ameliorative effect of Cynodon dactylon (L.) pers on scopolamine-induced amnesia in rats: Restoration of cholinergic and antioxidant pathways. Ind. J. Pharmacol. 2021, 53, 50. [Google Scholar] [CrossRef]
- Procházková, D.; Boušová, I.; Wilhelmová, N. Antioxidant and prooxidant properties of flavonoids. Fitoterapia 2011, 82, 513–523. [Google Scholar] [CrossRef]
- Moghaddam, A.H.; Zare, M. Neuroprotective effect of hesperetin and nano-hesperetin on recognition memory impairment and the elevated oxygen stress in rat model of Alzheimer’s disease. Biomed. Pharmacother. 2018, 97, 1096–1101. [Google Scholar]
- Ishola, I.O.; Tota, S.; Adeyemi, O.O.; Agbaje, E.O.; Narender, T.; Shukla, R. Protective effect of Cnestis ferruginea and its active constituent on scopolamine-induced memory impairment in mice: A behavioral and biochemical study. Pharm. Biol. 2013, 51, 825–835. [Google Scholar] [CrossRef]
- Zhao, J.; Ren, T.; Yang, M.; Zhang, Y.; Wang, Q.; Zuo, Z. Reduced systemic exposure and brain uptake of donepezil in rats with scopolamine-induced cognitive impairment. Xenobiotica 2020, 50, 389–400. [Google Scholar] [CrossRef]
- Bruce, K.D.; Zsombok, A.; Eckel, R.H. Lipid Processing in the Brain: A Key Regulator of Systemic Metabolism. Front. Endocrinol. 2017, 8, 60. [Google Scholar] [CrossRef] [Green Version]
- Lesa, G.M.; Palfreyman, M.; Hall, D.H.; Clandinin, M.T.; Rudolph, C.; Jorgensen, E.M.; Schiavo, G. Long chain polyunsaturated fatty acids are required for efficient neurotransmission in C. elegans. J. Cell Sci. 2003, 116, 4965–4975. [Google Scholar] [CrossRef] [Green Version]
- Willis, L.M.; Shukitt-Hale, B.; Joseph, J.A. Dietary polyunsaturated fatty acids improve cholinergic transmission in the aged brain. Genes Nutr. 2009, 4, 309–314. [Google Scholar] [CrossRef]
- Liu, Y.; Fu, X.; Lan, N.; Li, S.; Zhang, J.; Wang, S.; Li, C.; Shang, Y.; Huang, T.; Zhang, L. Luteolin protects against high fat diet-induced cognitive deficits in obesity mice. Behav. Brain Res. 2014, 267, 178–188. [Google Scholar] [CrossRef] [PubMed]
- Uriarte-Pueyo, I.; Calvo, M.I. Flavonoids as acetylcholinesterase inhibitors. Curr. Med. Chem. 2011, 18, 5289–5302. [Google Scholar] [CrossRef]
- Demirci, K.; Nazıroğlu, M.; Övey, İ.S.; Balaban, H. Selenium attenuates apoptosis, inflammation and oxidative stress in the blood and brain of aged rats with scopolamine-induced dementia. Metab. Brain Dis. 2017, 32, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Oyama, J.-i.; Maeda, T.; Sasaki, M.; Kozuma, K.; Ochiai, R.; Tokimitsu, I.; Taguchi, S.; Higuchi, Y.; Makino, N. Green tea catechins improve human forearm vascular function and have potent anti-inflammatory and anti-apoptotic effects in smokers. Internal. Med. 2010, 49, 2553–2559. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.-Z.; Deng, X.-H.; Bentivoglio, M. Differential response of apoptosis-regulatory Bcl-2 and Bax proteins to an inflammatory challenge in the cerebral cortex and hippocampus of aging mice. Brain Res. Bull. 2007, 74, 329–335. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Kim, J.-H.; He, M.-T.; Lee, A.-Y.; Cho, E.-J. Apigenin Ameliorates Scopolamine-Induced Cognitive Dysfunction and Neuronal Damage in Mice. Molecules 2021, 26, 5192. [Google Scholar] [CrossRef]
- Li, D.; Cai, C.; Liao, Y.; Wu, Q.; Ke, H.; Guo, P.; Wang, Q.; Ding, B.; Fang, J.; Fang, S. Systems pharmacology approach uncovers the therapeutic mechanism of medicarpin against scopolamine-induced memory loss. Phytomedicine 2021, 91, 153662. [Google Scholar] [CrossRef]
- Wang, R.; Shen, X.; Xing, E.; Guan, L.; Xin, L. Scutellaria baicalensis stem-leaf total flavonoid reduces neuronal apoptosis induced by amyloid beta-peptide (25–35). Neural Regen. Res. 2013, 8, 1081. [Google Scholar]
- Cutuli, D.; Pagani, M.; Caporali, P.; Galbusera, A.; Laricchiuta, D.; Foti, F.; Neri, C.; Spalletta, G.; Caltagirone, C.; Petrosini, L.; et al. Effects of Omega-3 Fatty Acid Supplementation on Cognitive Functions and Neural Substrates: A Voxel-Based Morphometry Study in Aged Mice. Front. Aging Neurosci. 2016, 8, 38. [Google Scholar] [CrossRef] [Green Version]
- Ajami, M.; Eghtesadi, S.; Razaz, J.M.; Kalantari, N.; Habibey, R.; Nilforoushzadeh, M.A.; Zarrindast, M.; Pazoki-Toroudi, H. Expression of Bcl-2 and Bax after hippocampal ischemia in DHA+ EPA treated rats. Neurol. Sci. 2011, 32, 811–818. [Google Scholar] [CrossRef]
- Farooq, S.; Mir, S.A.; Shah, M.A.; Manickavasagan, A. Chapter 2—Extraction techniques. In Plant Extracts: Applications in the Food Industry; Mir, S.A., Manickavasagan, A., Shah, M.A., Eds.; Academic Press: Cambridge, MA, USA, 2022; pp. 23–37. [Google Scholar] [CrossRef]
- Adusumilli, R.; Mallick, P. Data conversion with ProteoWizard msConvert. In Proteomics, Humana Press: New York, 2017; 339–368. In Proteomics; Humana Press: New York, NY, USA, 2017; pp. 339–368. [Google Scholar]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pluskal, T.; Korf, A.; Smirnov, A.; Schmid, R.; Fallon, T.R.; Du, X.; Weng, J.-K. CHAPTER 7 Metabolomics Data Analysis Using MZmine. In Processing Metabolomics and Proteomics Data with Open Software: A Practical Guide; The Royal Society of Chemistry: London, UK, 2020; pp. 232–254. [Google Scholar]
- Dührkop, K.; Fleischauer, M.; Ludwig, M.; Aksenov, A.A.; Melnik, A.V.; Meusel, M.; Dorrestein, P.C.; Rousu, J.; Böcker, S. SIRIUS 4: Turning tandem mass spectra into metabolite structure information. Nat. Methods 2019, 16, 299–302. [Google Scholar] [CrossRef] [Green Version]
- Aksoz, E.; Gocmez, S.S.; Sahin, T.D.; Aksit, D.; Aksit, H.; Utkan, T. The protective effect of metformin in scopolamine-induced learning and memory impairment in rats. Pharmacol. Rep. 2019, 71, 818–825. [Google Scholar] [CrossRef] [PubMed]
- Ademosun, A.O.; Adebayo, A.A.; Popoola, T.V.; Oboh, G. Shaddock (Citrus maxima) peels extract restores cognitive function, cholinergic and purinergic enzyme systems in scopolamine-induced amnesic rats. Drug Chem. Toxicol. 2022, 45, 1073–1080. [Google Scholar] [CrossRef] [PubMed]
- Sayed, R.H.; Ghazy, A.H.; Yammany, M.F.E. Recombinant human erythropoietin and interferon-beta-1b protect against 3-nitropropionic acid-induced neurotoxicity in rats: Possible role of JAK/STAT signaling pathway. Inflammopharmacology 2022, 30, 667–681. [Google Scholar] [CrossRef] [PubMed]
- Nunez, J. Morris Water Maze Experiment. J. Vis. Exp. 2008, 19, 897. [Google Scholar] [CrossRef] [Green Version]
- Biggan, S.L.; Beninger, R.J.; Cockhill, J.; Jhamandas, K.; Boegman, R.J. Quisqualate lesions of rat NBM: Selective effects on working memory in a double Y-maze. Brain Res. Bull. 1991, 26, 613–616. [Google Scholar] [CrossRef]
- Mihara, M.; Uchiyama, M. Determination of malonaldehyde precursor in tissues by thiobarbituric acid test. Anal. Biochem. 1978, 86, 271–278. [Google Scholar] [CrossRef] [PubMed]
- Beutler, E.; Duron, O.; Kelly, B.M. Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 1963, 61, 882–888. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Saeed, M.M.; Fernández-Ochoa, Á.; Saber, F.R.; Sayed, R.H.; Cádiz-Gurrea, M.d.l.L.; Elmotayam, A.K.; Leyva-Jiménez, F.J.; Segura-Carretero, A.; Nadeem, R.I. The Potential Neuroprotective Effect of Cyperus esculentus L. Extract in Scopolamine-Induced Cognitive Impairment in Rats: Extensive Biological and Metabolomics Approaches. Molecules 2022, 27, 7118. https://doi.org/10.3390/molecules27207118
Saeed MM, Fernández-Ochoa Á, Saber FR, Sayed RH, Cádiz-Gurrea MdlL, Elmotayam AK, Leyva-Jiménez FJ, Segura-Carretero A, Nadeem RI. The Potential Neuroprotective Effect of Cyperus esculentus L. Extract in Scopolamine-Induced Cognitive Impairment in Rats: Extensive Biological and Metabolomics Approaches. Molecules. 2022; 27(20):7118. https://doi.org/10.3390/molecules27207118
Chicago/Turabian StyleSaeed, Marwa M., Álvaro Fernández-Ochoa, Fatema R. Saber, Rabab H. Sayed, María de la Luz Cádiz-Gurrea, Amira K. Elmotayam, Francisco Javier Leyva-Jiménez, Antonio Segura-Carretero, and Rania I. Nadeem. 2022. "The Potential Neuroprotective Effect of Cyperus esculentus L. Extract in Scopolamine-Induced Cognitive Impairment in Rats: Extensive Biological and Metabolomics Approaches" Molecules 27, no. 20: 7118. https://doi.org/10.3390/molecules27207118
APA StyleSaeed, M. M., Fernández-Ochoa, Á., Saber, F. R., Sayed, R. H., Cádiz-Gurrea, M. d. l. L., Elmotayam, A. K., Leyva-Jiménez, F. J., Segura-Carretero, A., & Nadeem, R. I. (2022). The Potential Neuroprotective Effect of Cyperus esculentus L. Extract in Scopolamine-Induced Cognitive Impairment in Rats: Extensive Biological and Metabolomics Approaches. Molecules, 27(20), 7118. https://doi.org/10.3390/molecules27207118