
Rosmarinic Acid Prevents Cisplatin-Induced Liver and Kidney Injury by Inhibiting Inflammatory Responses and Enhancing Total Antioxidant Capacity, Thereby Activating the Nrf2 Signaling Pathway

 ,
,
Abstract
:1. Introduction
2. Results
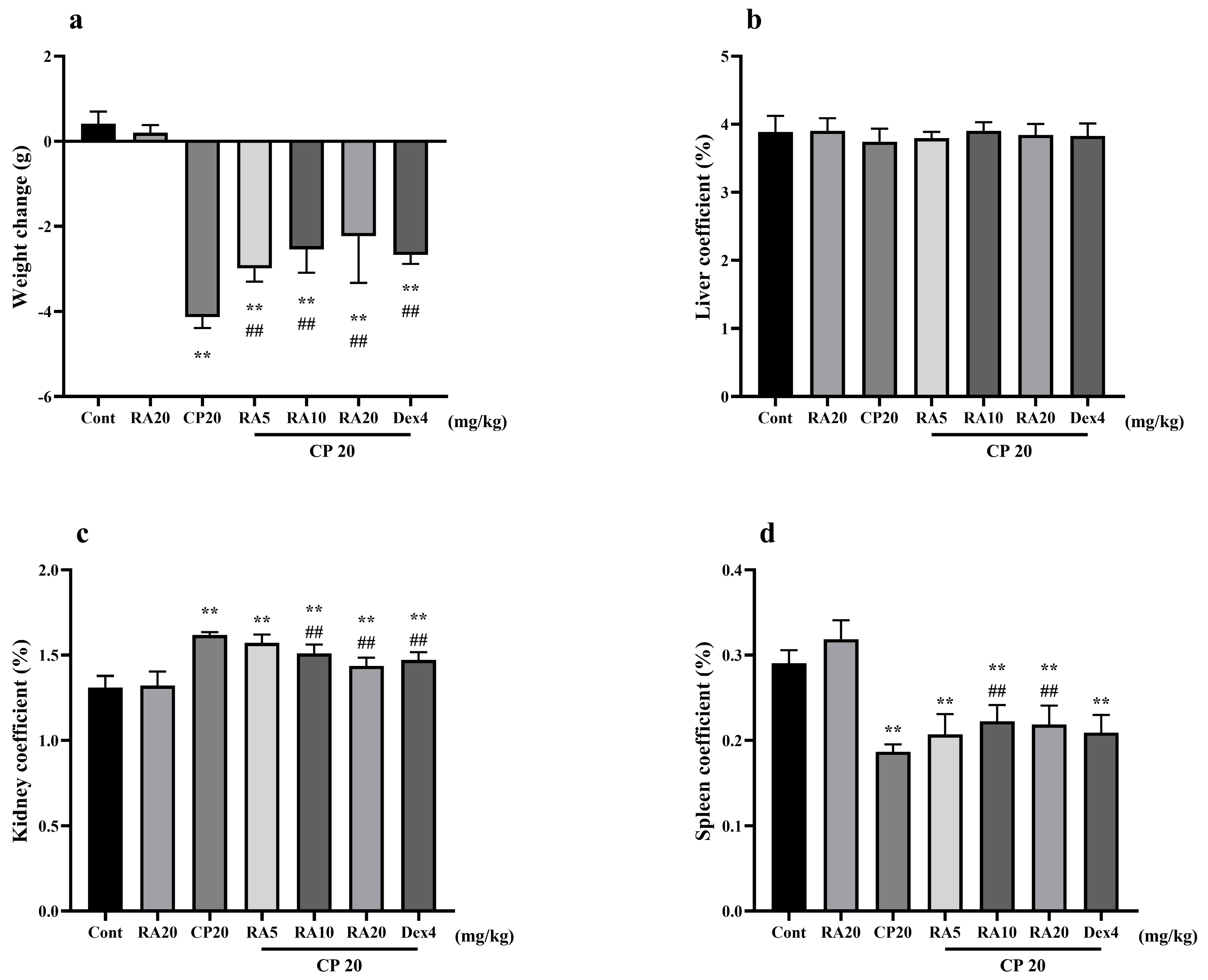
2.1. Effects of RA on Body Weight Changes and Organ Indices in Mice with CP-ALI and AKI
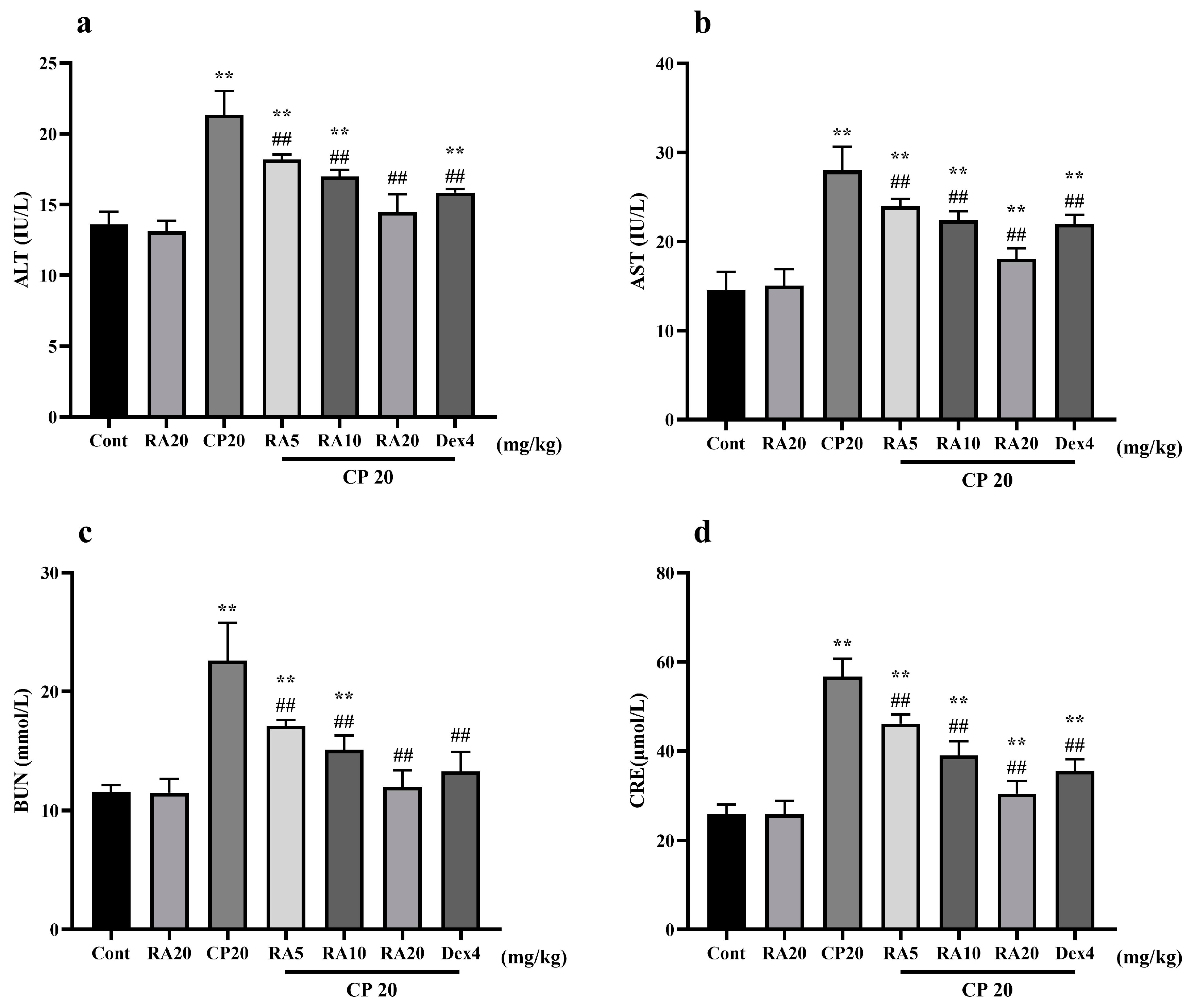
2.2. Effects of RA on Liver and Kidney Function in Mice with CP-ALI and AKI
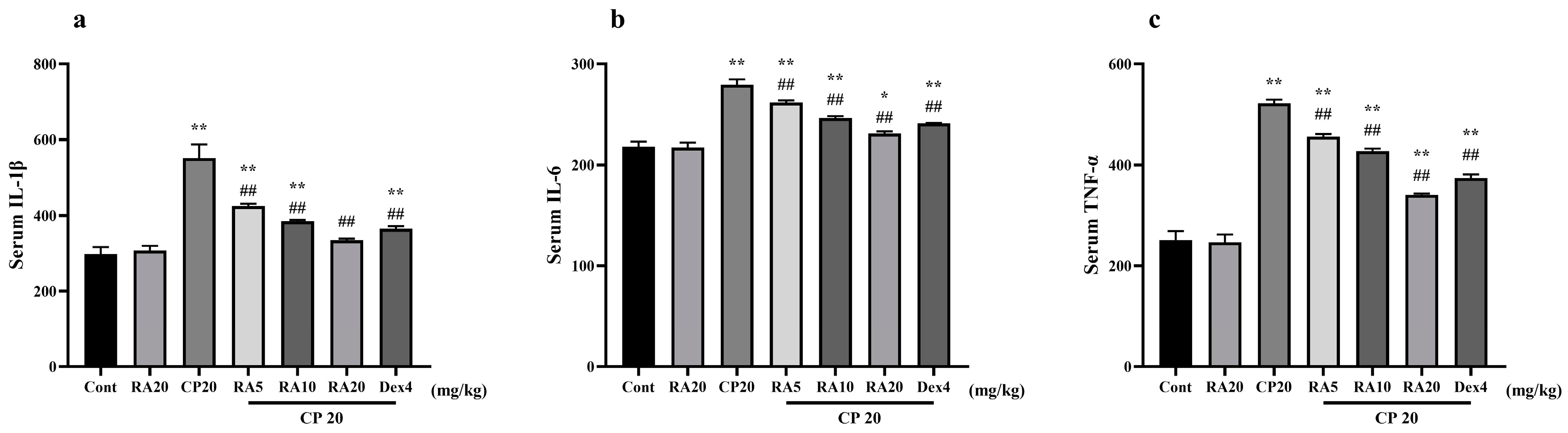
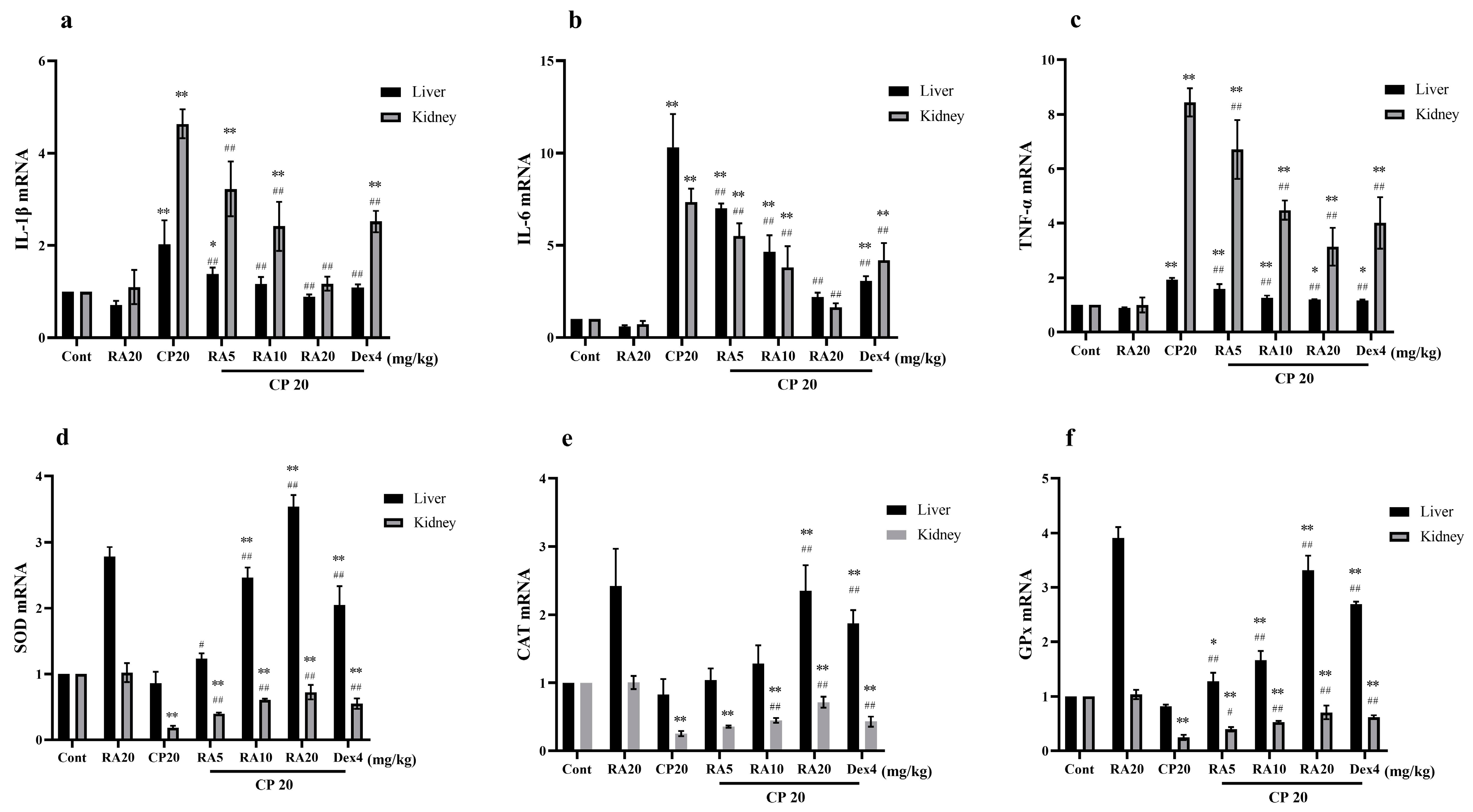
2.3. Effect of RA on Serum Levels of IL-1β, IL-6 and TNF-α Induced by CP
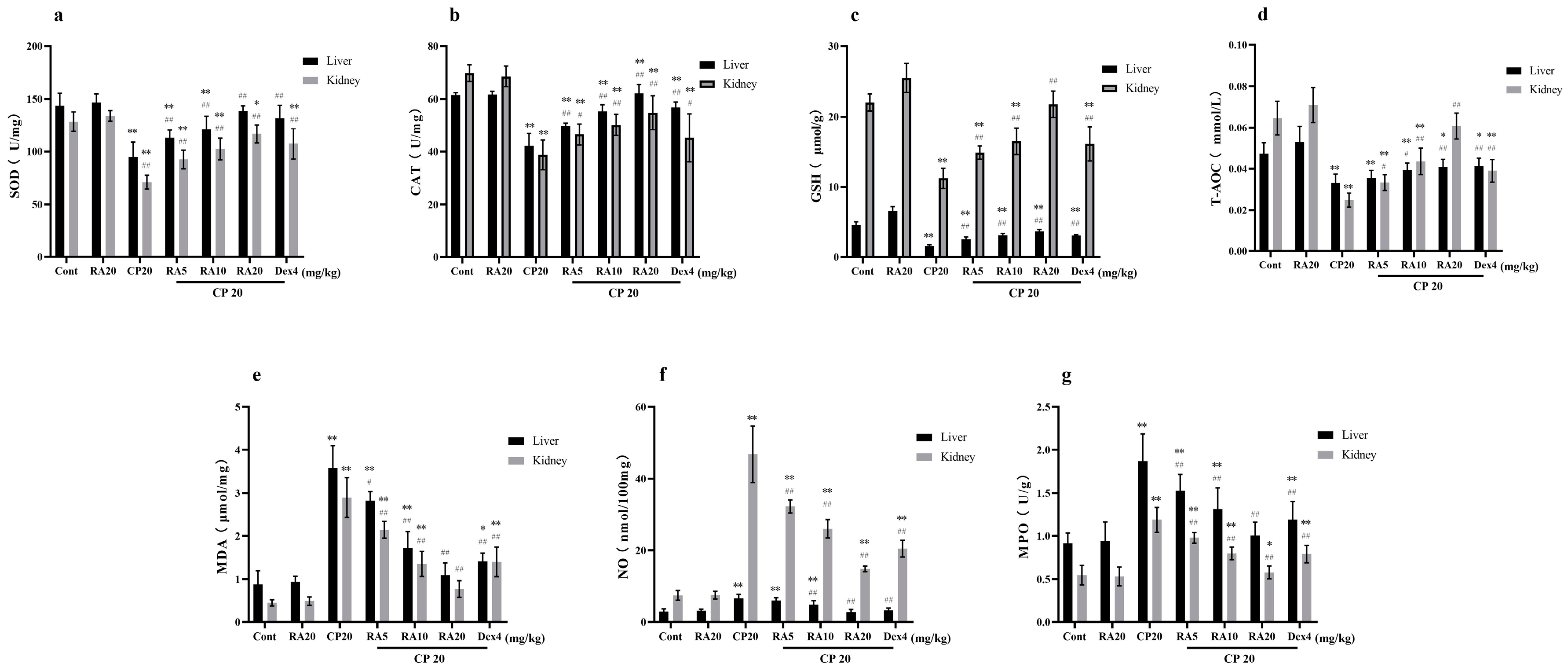
2.4. Effects of RA on the Antioxidant Capacity of the Liver and Kidney in the Presence of CP
2.5. Effects of RA on mRNA Expression in CP-Induced ALI and AKI
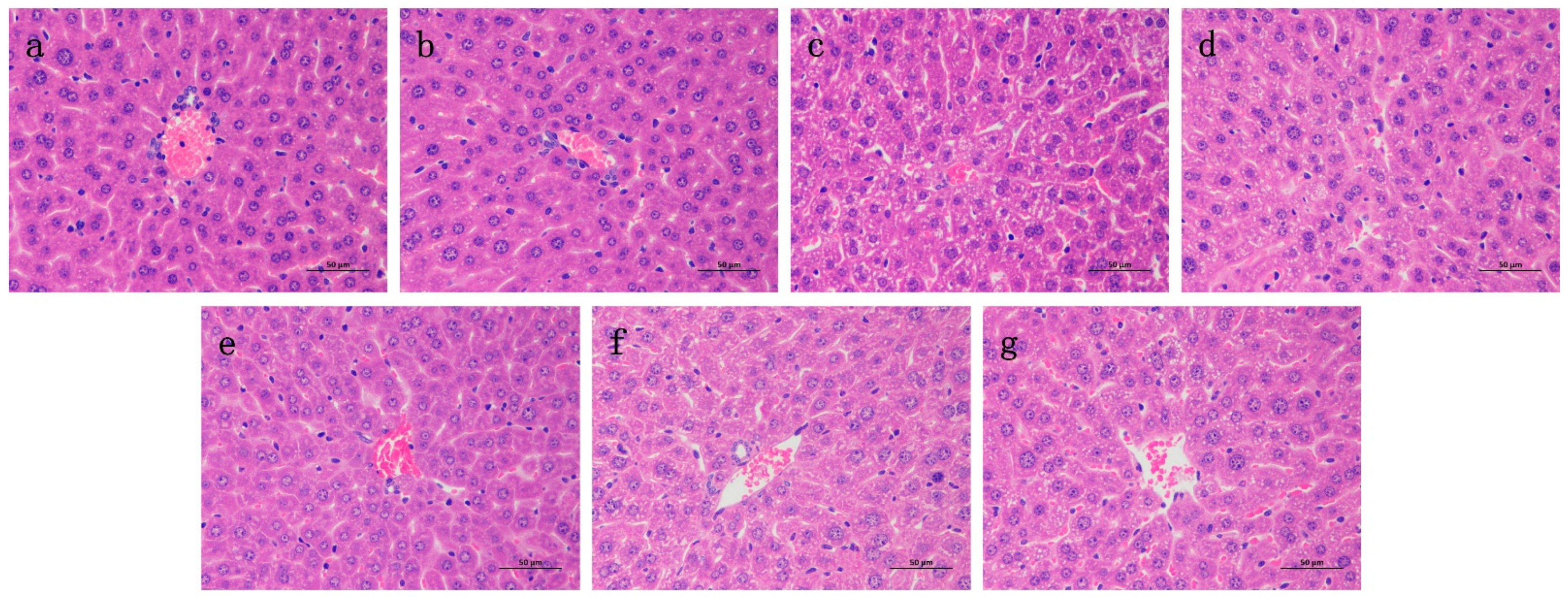
2.6. Effects of RA on Histological Changes of CP-Induced ALI and AKI in Mice
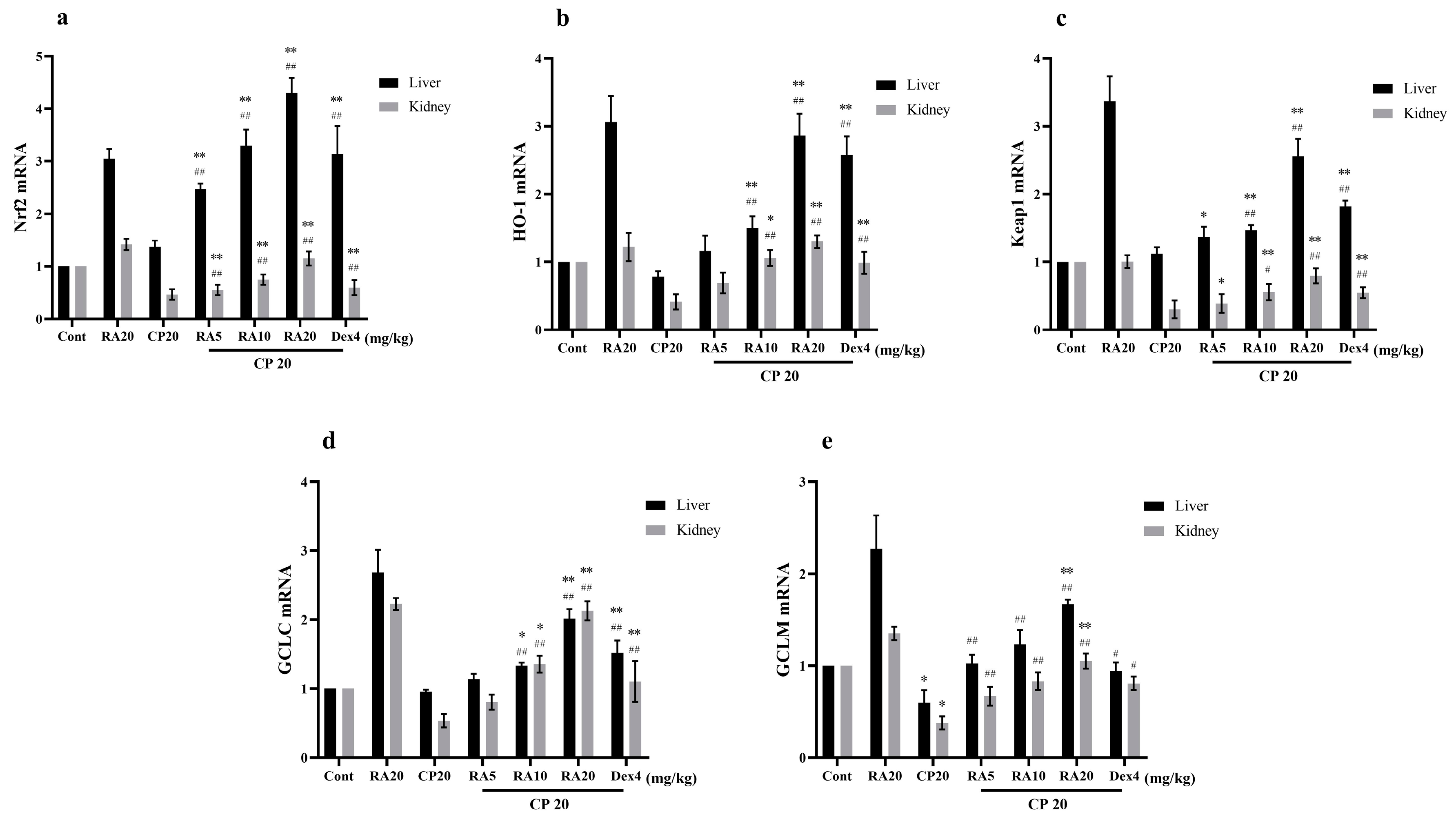
2.7. Effects of RA on CP-Induced mRNA Expression Levels of the Nrf2 Signaling Pathway
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Reagents
4.3. Challenge and Treatment of Mice
4.4. Collection and Testing of Blood and Organs
4.5. Real-Time Polymerase Chain Reaction (qPCR)
4.6. Liver and Kidney Histology
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Fan, X. Protective Effect and Mechanism of Daphnetin on Acute Kidney Injury; Jilin University: Changchun, China, 2019. [Google Scholar]
- Lv, H. The Protective Effect of Licorice Chalcone A on Acute Liver Injury and Its Mechanism; Jilin University: Changchun, China, 2019. [Google Scholar]
- Ren, C.; Yu, Z. Research progress of drug-induced acute kidney injury. China J. New Drugs Clin. Med. 2019, 38, 257–262. [Google Scholar]
- Zhao, H. The Role of Adenosine Receptor-Mediated cAMP Signaling Pathway in Acetaminophen-Induced Drug-Induced Liver Injury; Anhui Medical University: Hefei, China, 2015. [Google Scholar]
- Fang, K.; Xu, J.; Xu, K.; Zhan, Y. Mechanism of liver injury induced by chemotherapy drugs. J. Clin. Hepatobiliary Dis. 2020, 36, 677–679. [Google Scholar]
- Zhang, Q. Evaluation of Hepatotoxicity of Nanomaterials and Monomers of Traditional Chinese Medicine Based on Hepatocytes and Microtissues; Soochow University: Suzhou, China, 2019. [Google Scholar]
- Fan, X.; Wei, W.; Huang, J.; Peng, L.; Ci, X. Daphnetin Attenuated Cisplatin-Induced Acute Nephrotoxicity With Enhancing Antitumor Activity of Cisplatin by Upregulating SIRT1/SIRT6-Nrf2 Pathway. Front. Pharmacol. 2020, 11, 579178. [Google Scholar] [CrossRef] [PubMed]
- dos Santos, N.A.G.; Carvalho Rodrigues, M.A.; Martins, N.M.; Dos Santos, A.C. Cisplatin-induced nephrotoxicity and targets of nephroprotection: An update. Arch. Toxicol. 2012, 86, 1233–1250. [Google Scholar] [CrossRef]
- Peres, L. ADD Cunha Júnior. Acute nephrotoxicity of cisplatin: Molecular mechanisms. Braz. J. Nephrol. 2013, 35, 332–340. [Google Scholar] [CrossRef]
- Zang, Y. Research on the Biological Activity of Perilla Rosmarinic Acid and Its Application; North Central University: Minneapolis, CA, USA, 2018. [Google Scholar]
- Baranauskaite, J.; Sadauskiene, I.; Liekis, A.; Kasauskas, A.; Lazauskas, R.; Zlabiene, U.; Masteikova, R.; Kopustinskiene, D.M.; Bernatoniene, J. Natural Compounds Rosmarinic Acid and Carvacrol Counteract Aluminium-Induced Oxidative Stress. Molecules 2020, 25, 1807. [Google Scholar] [CrossRef] [Green Version]
- Bacanlı, M.; Aydın, S.; Taner, G.; Göktaş, H.G.; Şahin, T.; Başaran, A.A.; Başaran, N. Does rosmarinic acid treatment have protective role against sepsis-induced oxidative damage in Wistar Albino rats? Hum. Exp. Toxicol. 2016, 35, 877–886. [Google Scholar] [CrossRef]
- Ding, Y.; Zhang, Z.; Yue, Z.; Ding, L.; Zhou, Y.; Huang, Z.; Huang, H. Rosmarinic acid Ameliorates H2O2-Induced Oxidative Stress on L02 Cells through MAPK and Nrf2 Pathways. Rejuvenation Res. 2019, 22, 289–298. [Google Scholar] [CrossRef]
- Chu, X.; Ci, X.; He, J.; Jiang, L.; Wei, M.; Cao, Q.; Guan, M.; Xie, X.; Deng, X.; He, J. Effects of a Natural Prolyl Oligopeptidase Inhibitor, Rosmarinic Acid, on Lipopolysaccharide-Induced Acute Lung Injury in Mice. Molecules 2012, 17, 3586–3598. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.; Xu, Y.; Wen, X.; Nie, H.; Hu, T.; Yang, X.; Chu, X.; Yang, J.; Deng, X.; He, J. Rosmarinic Acid Attenuates Airway Inflammation and Hyperresponsiveness in a Murine Model of Asthma. Molecules 2016, 21, 769. [Google Scholar] [CrossRef] [Green Version]
- Liang, Z.; Nie, H.; Xu, Y.; Peng, J.; Zeng, Y.; Wei, Y.; Wen, X.; Qiu, J.; Zhong, W.; Deng, X.; et al. Therapeutic effects of rosmarinic acid on airway responses in a murine model of asthma. Int. Immunopharmacol. 2016, 41, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Liang, Z.; Wu, L.; Deng, X.; Liang, Q.; Xu, Y.; Deng, R.; Lv, L.; Ji, M.; Hao, Z.; He, J. The Antioxidant Rosmarinic Acid Ameliorates Oxidative Lung Damage in Experimental Allergic Asthma via Modulation of NADPH Oxidases and Antioxidant Enzymes. Inflammation 2020, 43, 1902–1912. [Google Scholar] [CrossRef] [PubMed]
- Duan, Z.; Cai, G.; Li, J.; Chen, X. Cisplatin-induced renal toxicity in elderly people. Ther. Adv. Med. Oncol. 2020, 12, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Barton, C.D.; Pizer, B.; Jones, C.; Oni, L.; Pirmohamed, M.; Hawcutt, D.B. Identifying cisplatin-induced kidney damage in paediatric oncology patients. Pediatric Nephrol. J. Int. Pediatr. Nephrol. Assoc. 2018, 33, 1467–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sumit, G. Cisplatin: The first metal based anticancer drug. Bioorganic Chem. 2019, 88, 102925. [Google Scholar]
- Pezeshki, Z.; Khosravi, A.; Nekuei, M.; Khoshnood, S.; Zandi, E.; Eslamian, M.; Talebi, A.; Nematbakhsh, M. Time course of cisplatin-induced nephrotoxicity and hepatotoxicity. J. Nephropathol. 2017, 6, 163–167. [Google Scholar] [CrossRef] [Green Version]
- Qi, L.; Luo, Q.; Zhang, Y.; Jia, F.; Zhao, Y.; Wang, F. Advances in Toxicological Research of the Anticancer Drug Cisplatin. Chem. Res. Toxicol. 2019, 32, 1469–1486. [Google Scholar] [CrossRef]
- Xiang, Y.; Ji, M.; Wu, L.; He, J. Establishment of a pathological model of acute liver and kidney injury in mice. Adv. Anim. Med. 2022, 43, 56–59. [Google Scholar]
- Bian, X.; Liu, X.; Liu, J.; Zhao, Y.; Li, H.; Zhang, L.; Li, P.; Gao, Y. Hepatoprotective effect of chiisanoside from acanthopanax sessiliflorus against lps/d-galn-induced acute liver injury by inhibiting nf-κb and activating nrf2/ho-1 signaling pathways. J. Sci. Food Agric. 2018, 99, 3283–3290. [Google Scholar] [CrossRef]
- Sherif, I.O. The effect of natural antioxidants in cyclophosphamide-induced hepatotoxicity: Role of Nrf2/HO-1 pathway. Int. Immunopharmacol. 2018, 61, 29–36. [Google Scholar] [CrossRef]
- Ansari, M.A. Sinapic acid modulates Nrf2/HO-1 signaling pathway in cisplatin-induced nephrotoxicity in rats. Biomed. Pharmacother. 2017, 93, 646–653. [Google Scholar] [CrossRef] [PubMed]
- Holditch, S.J.; Brown, C.N.; Lombardi, A.M.; Nguyen, K.N.; Edelstein, C.L. Recent Advances in Models, Mechanisms, Biomarkers, and Interventions in Cisplatin-Induced Acute Kidney Injury. Int. J. Mol. Sci. 2019, 20, 3011. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, W.; Fu, Z.; Zou, Y.; Wen, D.; Ma, H.; Zhou, F.; Chen, Y.; Zhang, M.; Zhang, W. MicroRNA-140-5p attenuated oxidative stress in Cisplatin induced acute kidney injury by activating Nrf2/ARE pathway through a Keap1-independent mechanism. Exp. Cell Res. 2017, 360, 292–302. [Google Scholar] [CrossRef] [PubMed]
- ALHaithloul, H.A.; Alotaibi, M.F.; Bin-Jumah, M.; Elgebaly, H.; Mahmoud, A.M. Olea europaea leaf extract up-regulates Nrf2/ARE/HO-1 signaling and attenuates cyclophosphamide-induced oxidative stress, inflammation and apoptosis in rat kidney. Biomed. Pharmacother. 2019, 111, 676–685. [Google Scholar] [CrossRef]
- Hitl, M.; Kladar, N.; Gavarić, N.; Božin, B. Rosmarinic Acid–Human Pharmacokinetics and Health Benefits. Planta Med. 2021, 87, 273–282. [Google Scholar] [CrossRef]
- Fan, X.; Wei, W.; Huang, J.; Liu, X.; Ci, X. Isoorientin Attenuates Cisplatin-Induced Nephrotoxicity Through the Inhibition of Oxidative Stress and Apoptosis via Activating the SIRT1/SIRT6/Nrf-2 Pathway. Front. Pharmacol. 2020, 11, 264. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Gu, W.; Ma, N.; Fan, X.; Ci, X. Leonurine hydrochloride alleviates ferroptosis in cisplatin-induced acute kidney injury by activating the Nrf2 signaling pathway. Br. J. Pharmacol. 2022, 179, 3991–4009. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, M.; Mou, J.; Zhao, Z.; Yang, J.; Zhu, F.; Pei, G.; Zhu, H.; Wang, Y.; Xu, G.; et al. Pretreatment of Huaiqihuang extractum protects against cisplatin-induced nephrotoxicity. Sci. Rep. 2018, 8, 7333. [Google Scholar] [CrossRef]
- Gabbia, D.; Pozzo, L.; Zigiotto, G.; Roverso, M.; Sacchi, D.; Dalla Pozza, A.; Carrara, M.; Bogialli, S.; Floreani, A.; Guido, M.; et al. Dexamethasone counteracts hepatic inflammation and oxidative stress in cholestatic rats via CAR activation. PLoS ONE 2018, 13, e0204336. [Google Scholar] [CrossRef]
- Zhai, Z.; Ouyang, W.; Yao, Y.; Zhang, Y.; Zhang, H.; Xu, F.; Gao, C. Dexamethasone-loaded ROS-responsive poly(thioketal)nanoparticles suppress inflammation and oxidative stress of acute lung injur. Bioact. Mater. 2022, 14, 430–442. [Google Scholar] [CrossRef]
- Yi, L.; Cui, J.; Wang, W.; Tang, W.; Teng, F.; Zhu, X.; Qin, J.; Wuniqiemu, T.; Sun, J.; Wei, Y.; et al. Formononetin Attenuates Airway Inammation and Oxidative Stress in Murine Allergic Asthma. Front. Pharmacol. 2020, 11, 533841. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Wang, W.; Li, Z.; Cheng, J.; Wang, W.; Zhang, J. Establishment and evaluation of ischemia-reperfusion and cisplatin-induced acute kidney injury model in mice. Exp. Anim. Comp. Med. 2014, 34, 187–192. [Google Scholar]
- Guo, Y. Protective Effect and Mechanism of Sophora Japonica Yellow Infusion on Cisplatin-Induced Acute Kidney Injury in Mice; Huazhong University of Science and Technology: Wuhan, China, 2016. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targets | Forward | Reverse |
|---|---|---|
| IL-1β | AAAAAAGCCTCGTGCTGTCG | GTCGTTGCTTGGTTCTCCTTG |
| IL-6 | CTAGTGCGTTATGCCTAAGC | ATAGTGTCCCAACATTCATATTGTC |
| TNF-α | AGCACAGAAAGCATGATCCG | CTGATGAGAGGGAGGCCATT |
| SOD | CGGCTTCTCGTCTTGCT | GTTCACCGCTTGCCTTC |
| CAT | GCCAACTACCAGCGTGA | CCGCACCTGAGTGACAT |
| GPx | GGCATTGGCTTGGTGATT | AGGTGGAAAGGCATCGG |
| Nrf2 | GCTCCTATGCGTGAATCCCA | TTCTGGGCGGCGACTTTATT |
| HO-1 | GTCAAGCACAGGGTGACAGA | AAGTGACGCCATCTGTGAGG |
| Keap1 | ACCTCGGACTCGCAGCGTAC | CCAAGCAGGAGGAGTTCTTCAACC |
| GCLC | CTGCACATCTACCACGCAGT | GTCTCAAGAACATCGCCTCC |
| GCLM | CGGGAACCTGCTCAACTG | CCAAAACATCTGGAAACTCCC |
| β-actin | ATCACTATTGGCAACGAGCG | TCAGCAATGCCTGGGTACAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiang, Y.; Ji, M.; Wu, L.; Lv, L.; Liang, Q.; Deng, R.; Deng, Z.; Liu, X.; Ren, L.; Feng, X.; et al. Rosmarinic Acid Prevents Cisplatin-Induced Liver and Kidney Injury by Inhibiting Inflammatory Responses and Enhancing Total Antioxidant Capacity, Thereby Activating the Nrf2 Signaling Pathway. Molecules 2022, 27, 7815. https://doi.org/10.3390/molecules27227815
Xiang Y, Ji M, Wu L, Lv L, Liang Q, Deng R, Deng Z, Liu X, Ren L, Feng X, et al. Rosmarinic Acid Prevents Cisplatin-Induced Liver and Kidney Injury by Inhibiting Inflammatory Responses and Enhancing Total Antioxidant Capacity, Thereby Activating the Nrf2 Signaling Pathway. Molecules. 2022; 27(22):7815. https://doi.org/10.3390/molecules27227815
Chicago/Turabian StyleXiang, Yifei, Min Ji, Liqin Wu, Li Lv, Qiuling Liang, Ruihan Deng, Zhaoyou Deng, Xia Liu, Lingyi Ren, Xin Feng, and et al. 2022. "Rosmarinic Acid Prevents Cisplatin-Induced Liver and Kidney Injury by Inhibiting Inflammatory Responses and Enhancing Total Antioxidant Capacity, Thereby Activating the Nrf2 Signaling Pathway" Molecules 27, no. 22: 7815. https://doi.org/10.3390/molecules27227815