

Dual Topoisomerase I/II Inhibition-Induced Apoptosis and Necro-Apoptosis in Cancer Cells by a Novel Ciprofloxacin Derivative via RIPK1/RIPK3/MLKL Activation
 ,
,  , ,
, , 
Abstract
:1. Introduction
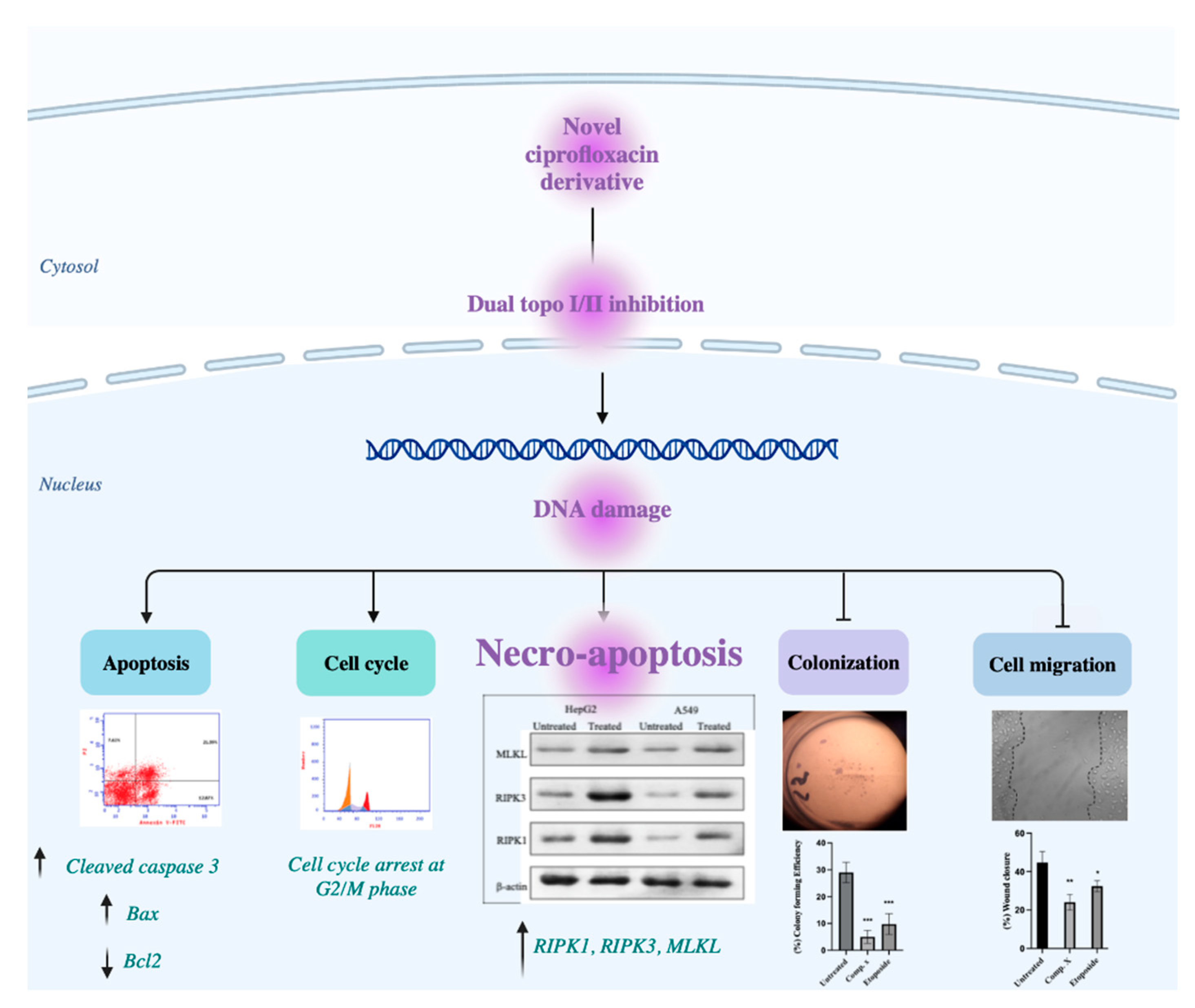
2. Results
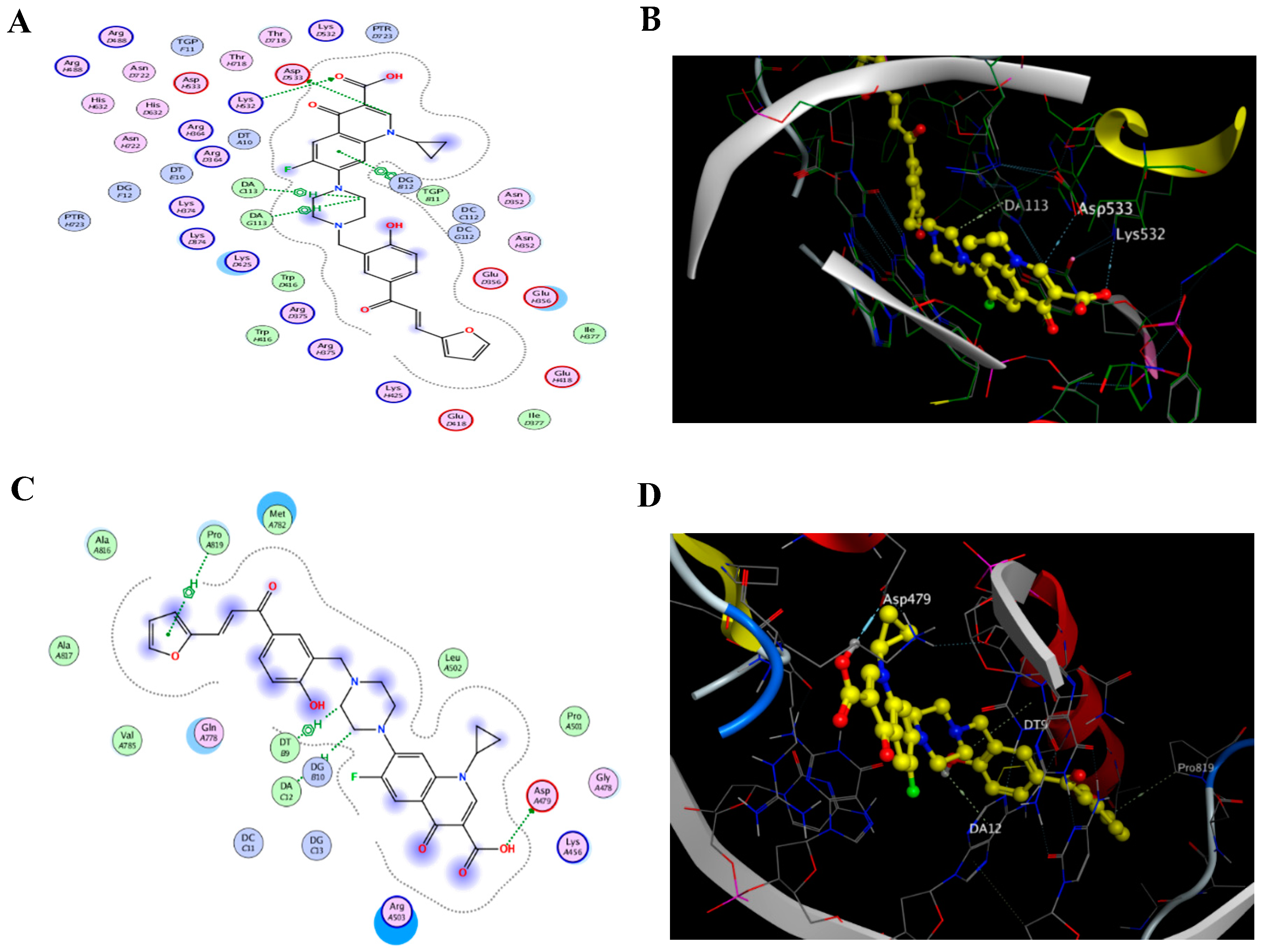
2.1. Docking of the CM Derivative to Topo I and II Enzymes
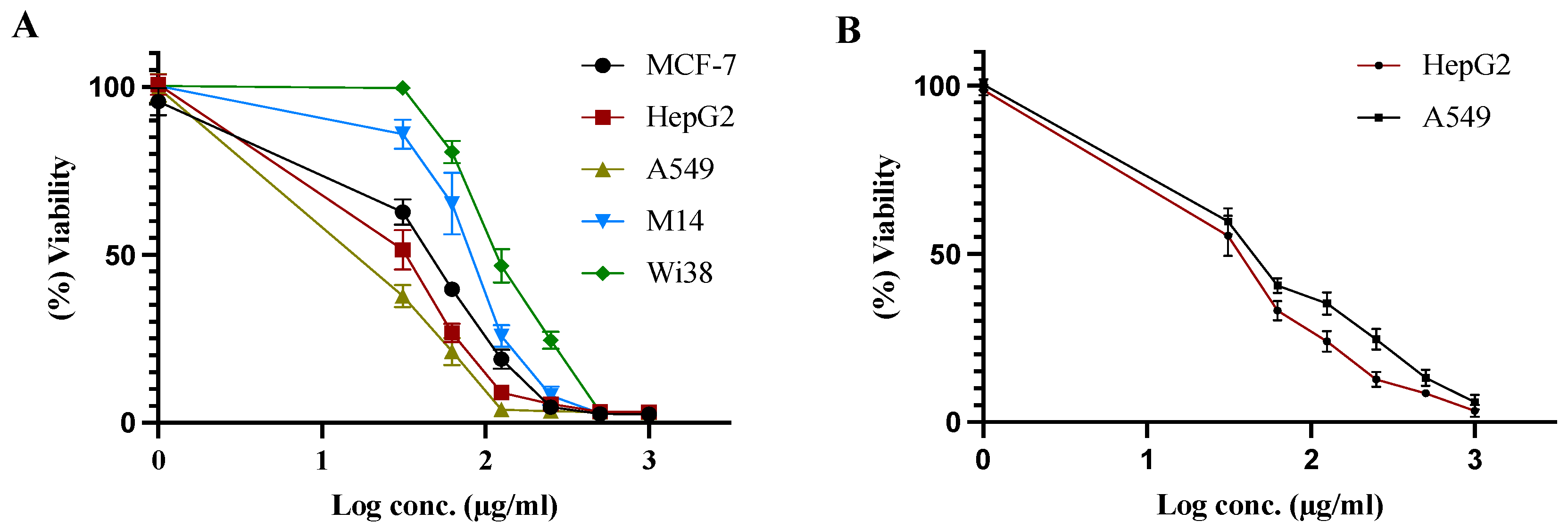
2.2. Cell Viability
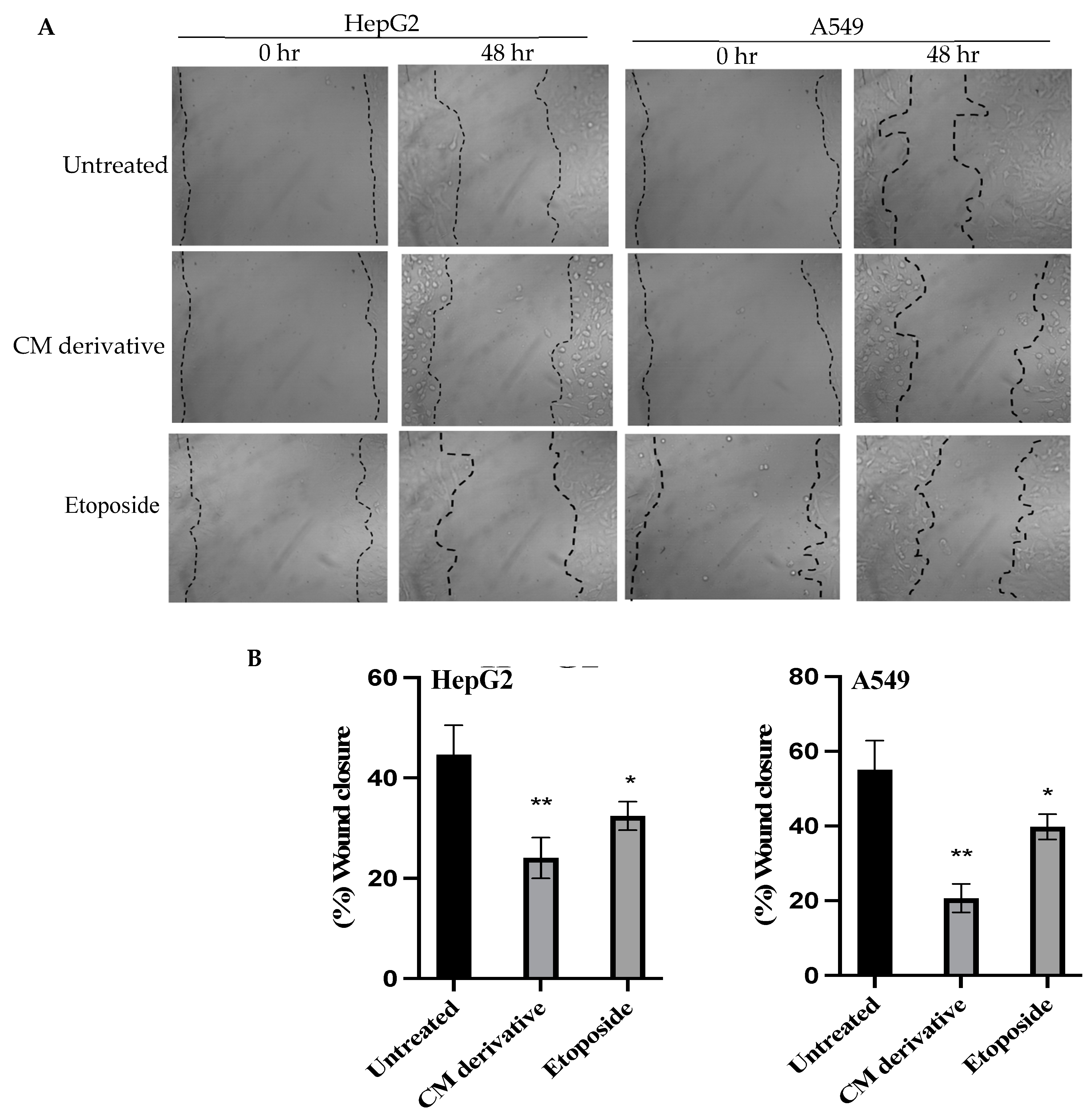
2.3. Cell Migration Assay
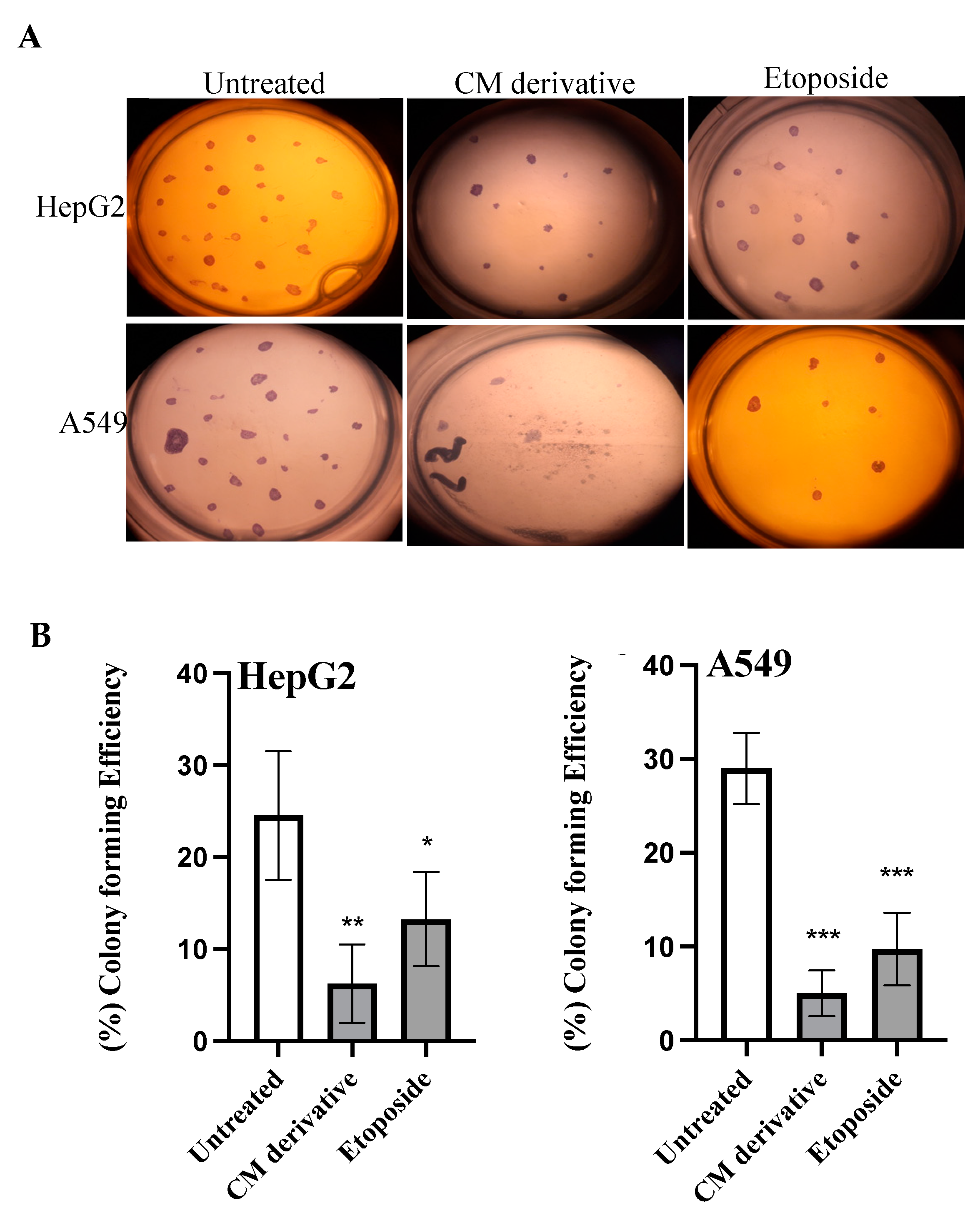
2.4. Colony Formation Assay
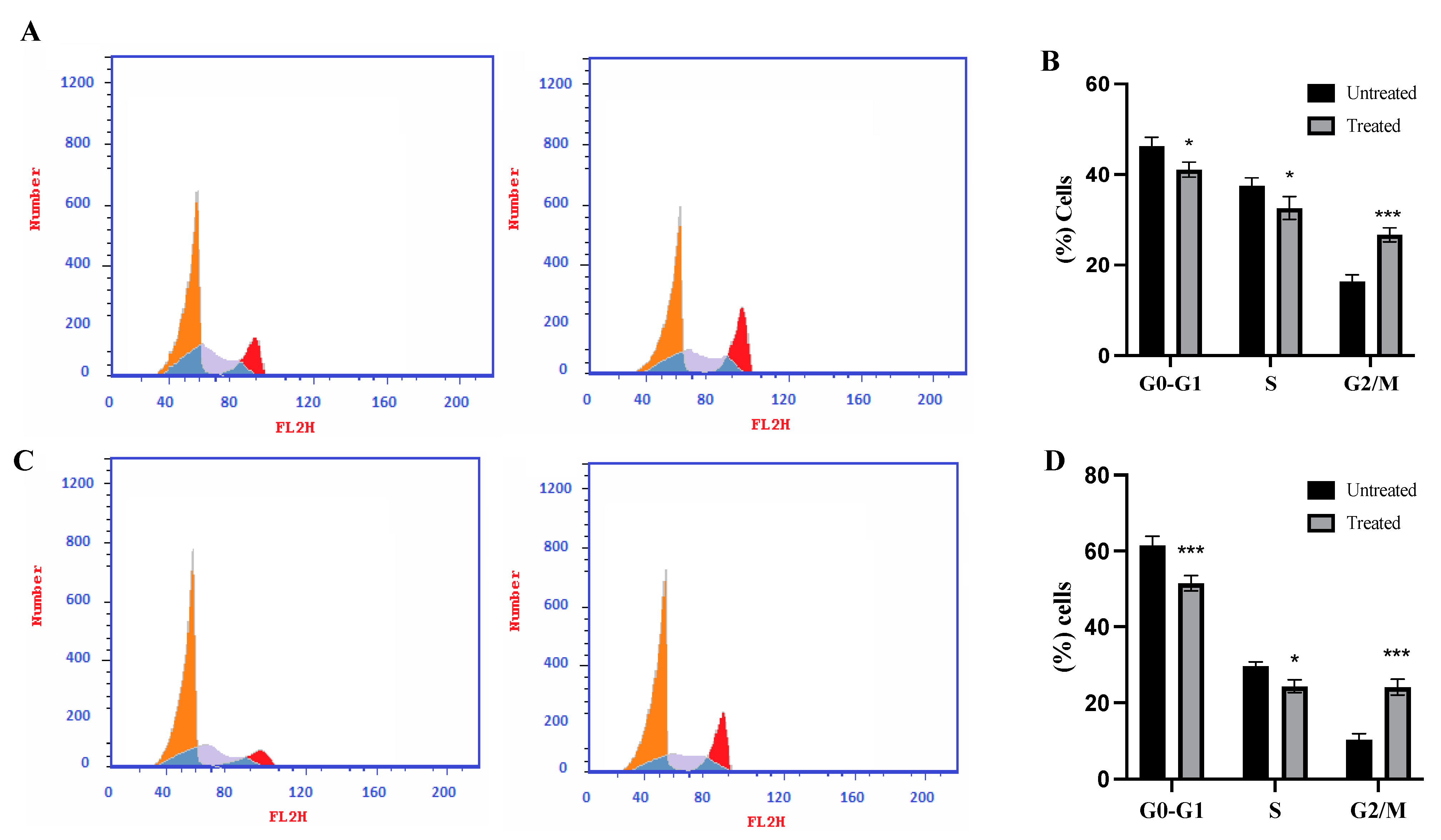
2.5. Cell Cycle Analysis
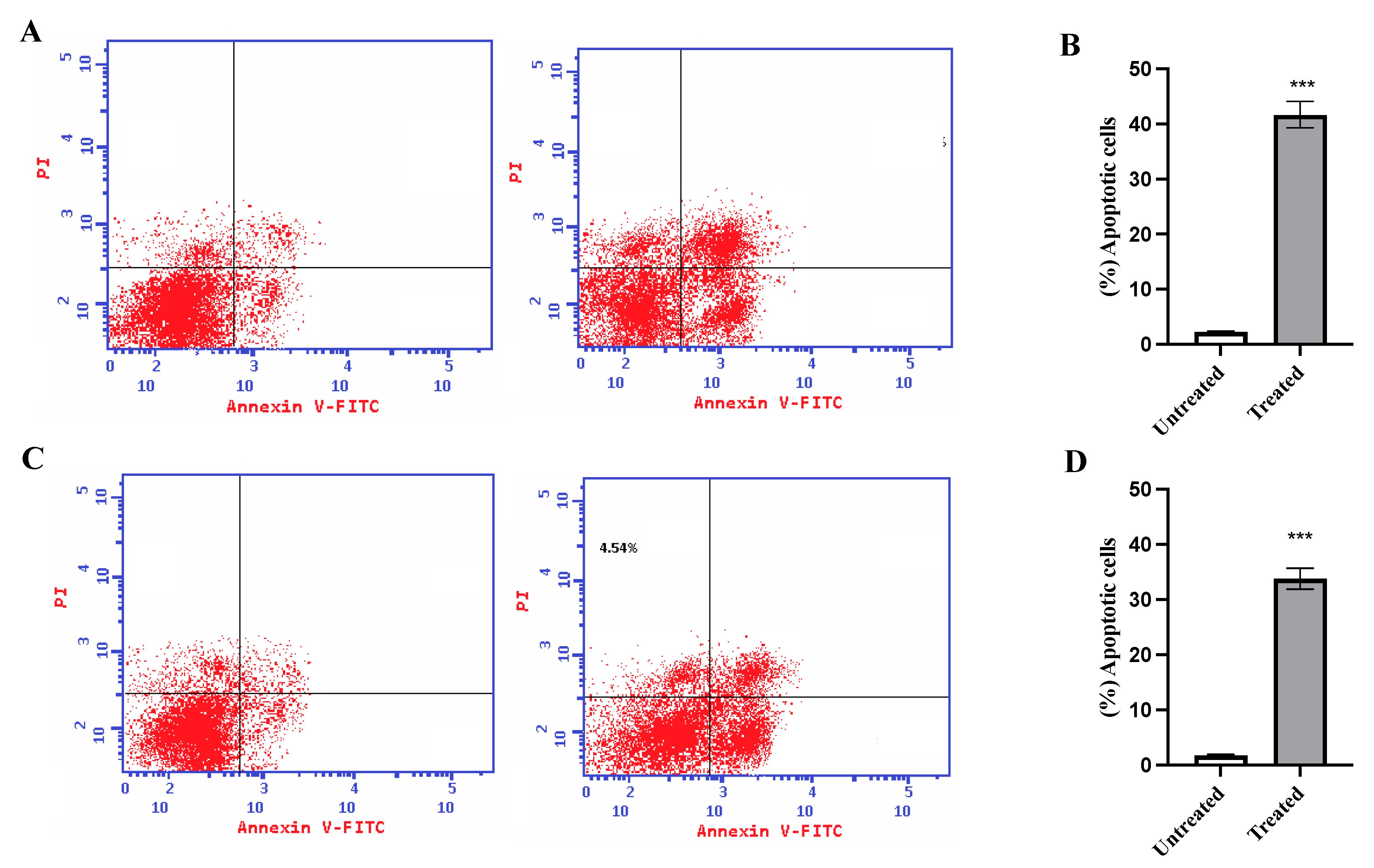
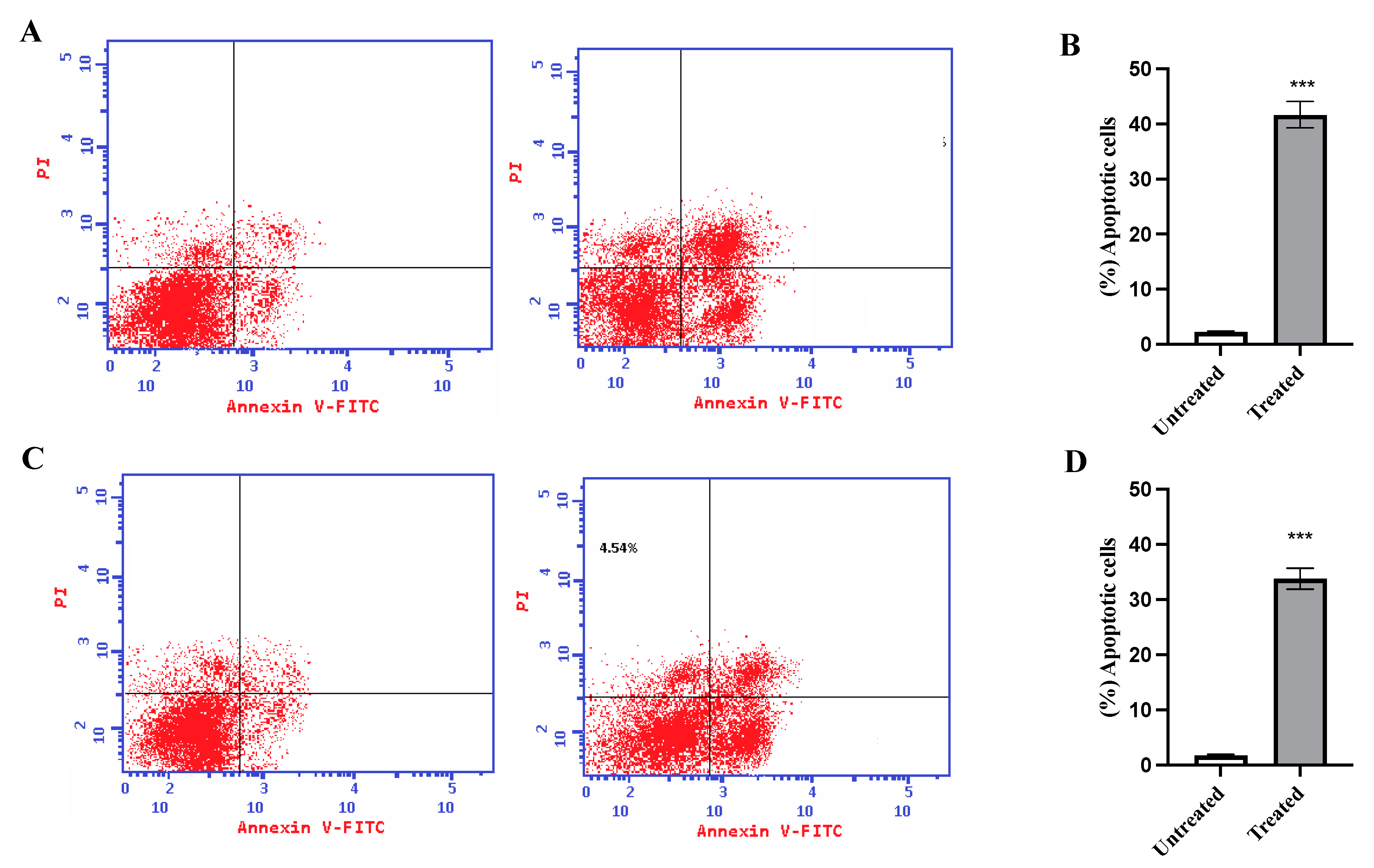
2.6. Annexin V Assay
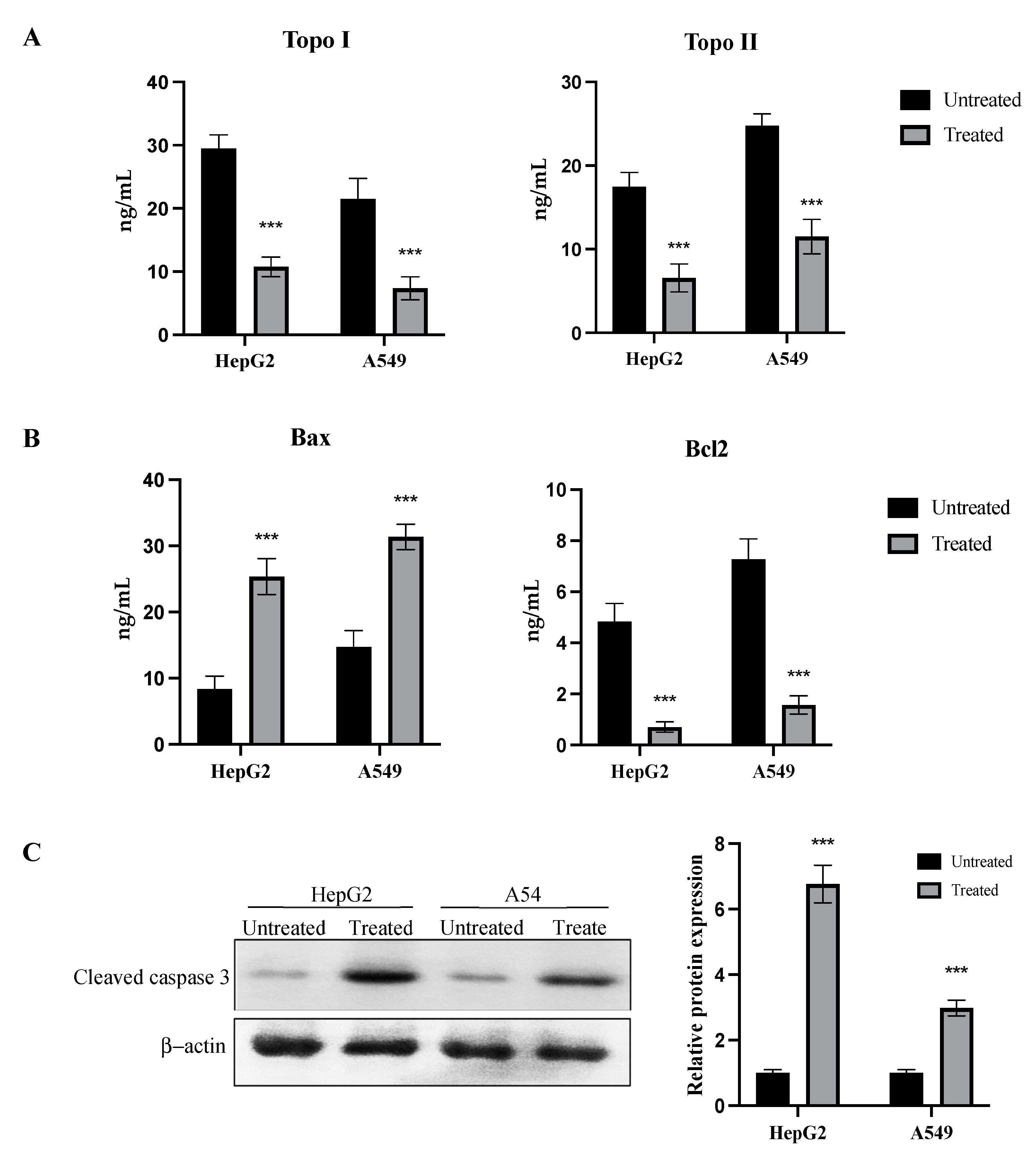
2.7. Inhibition of Topo I/II Enzymes and Activation of Apoptotic Pathway
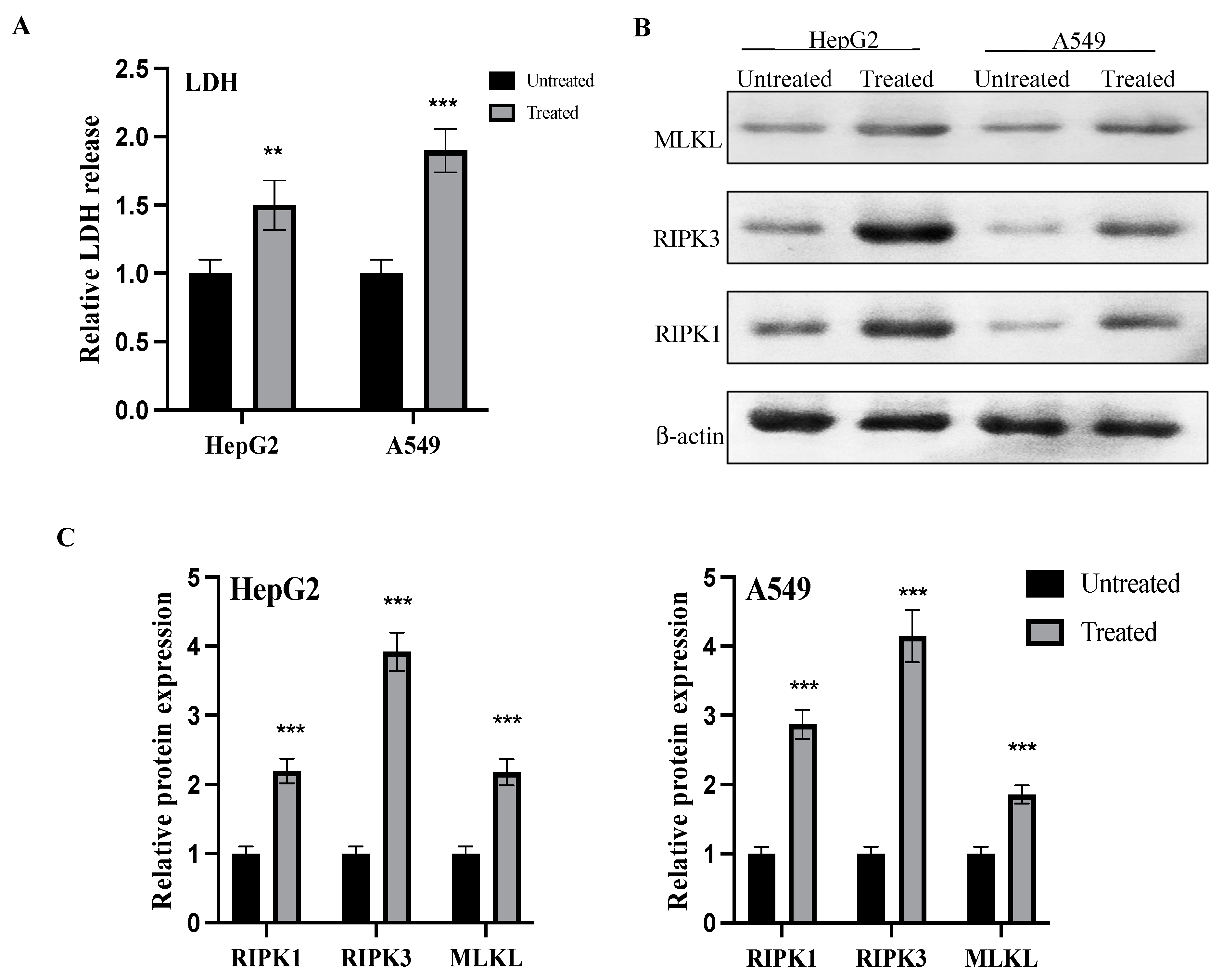
2.8. LDH Release and Activation of Necro-Apoptotic Pathway
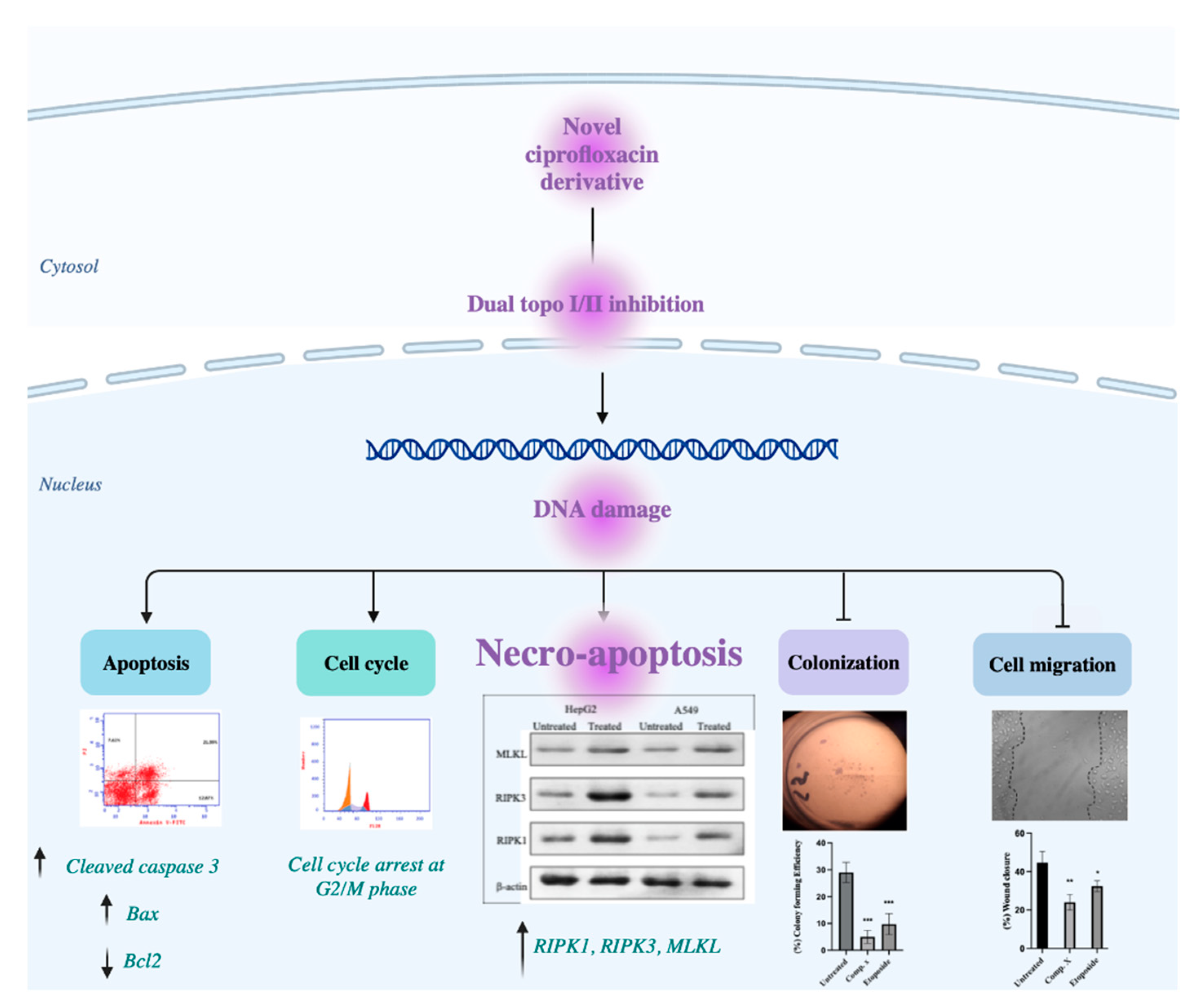
3. Discussion
4. Materials and Methods
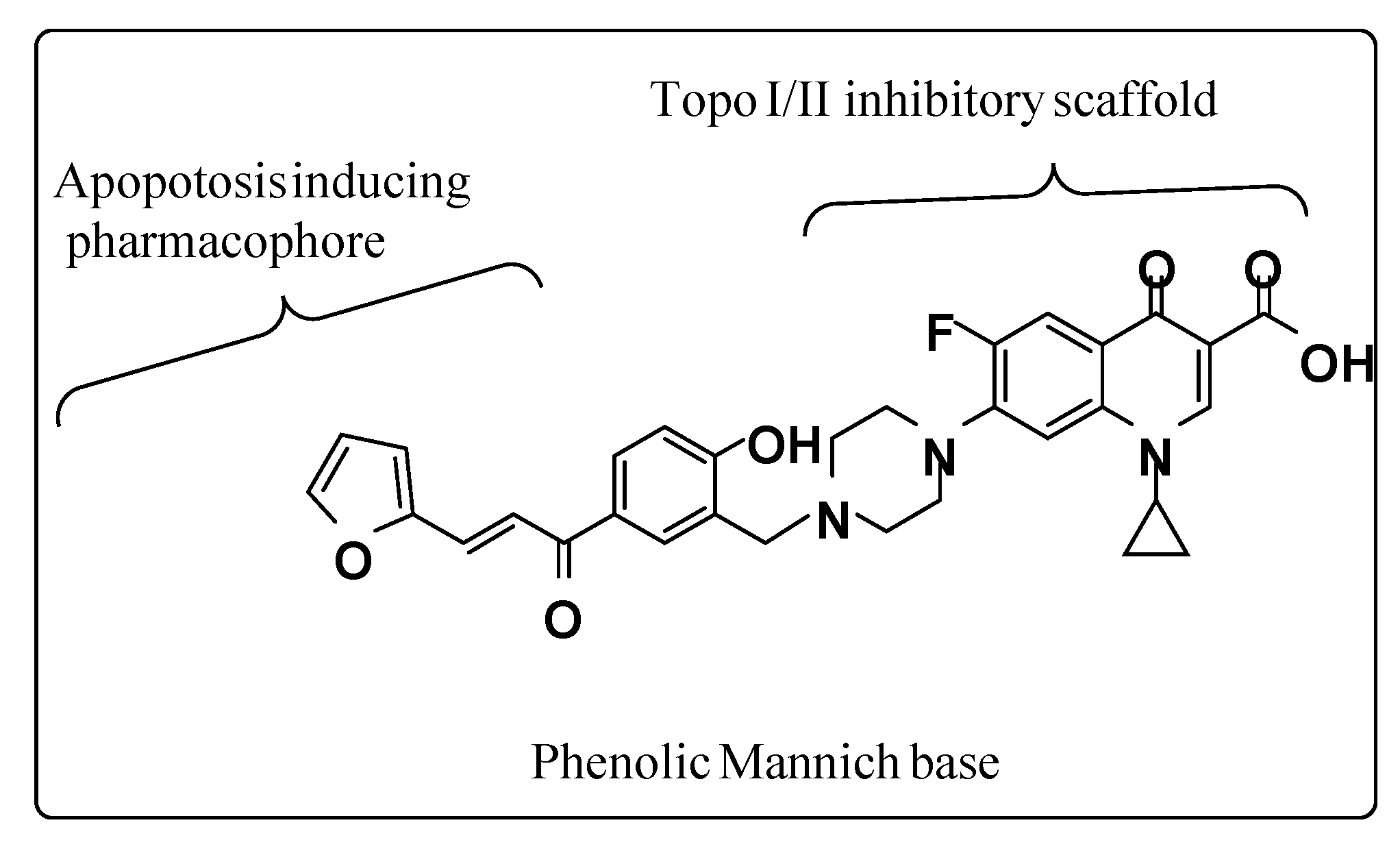
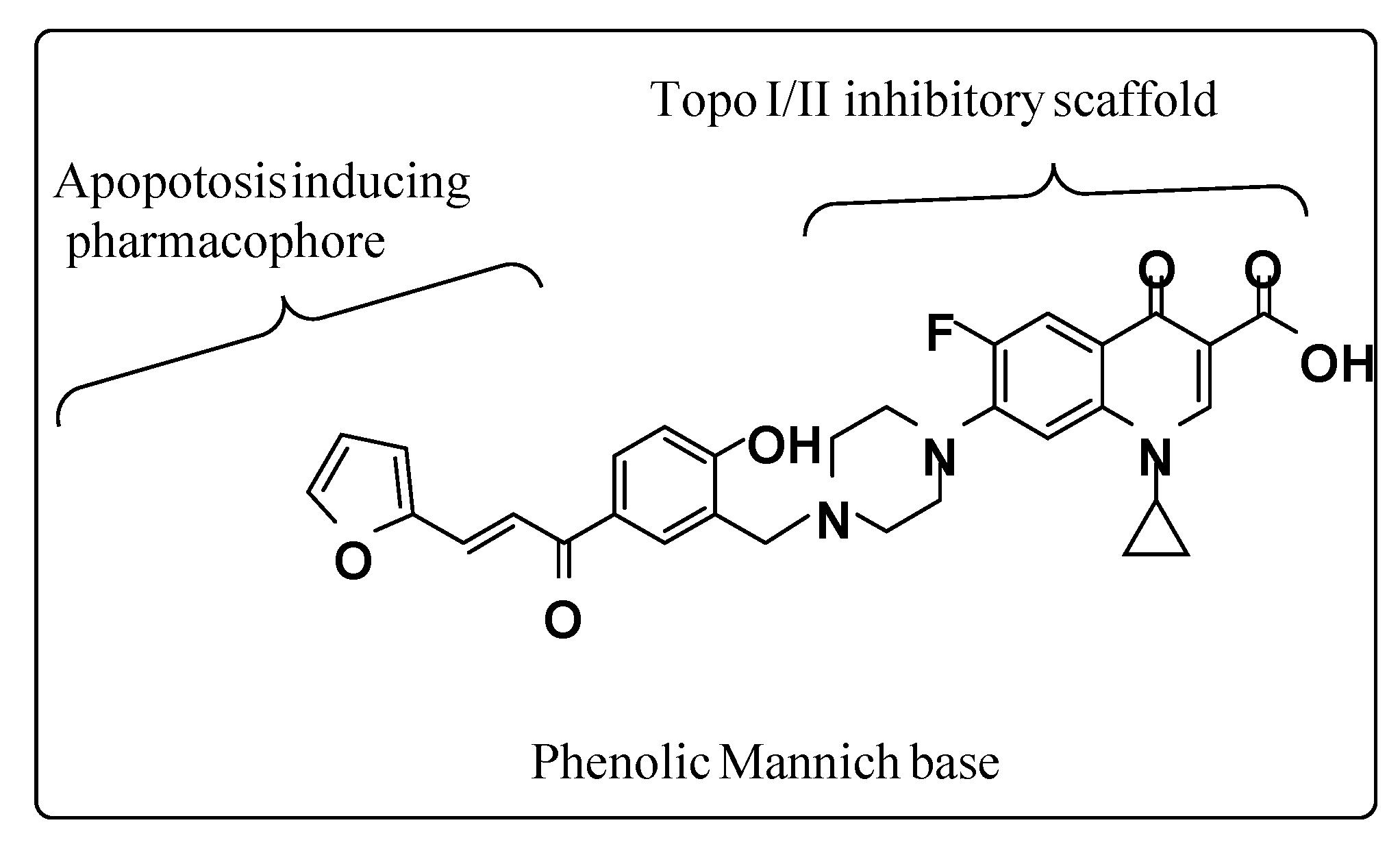
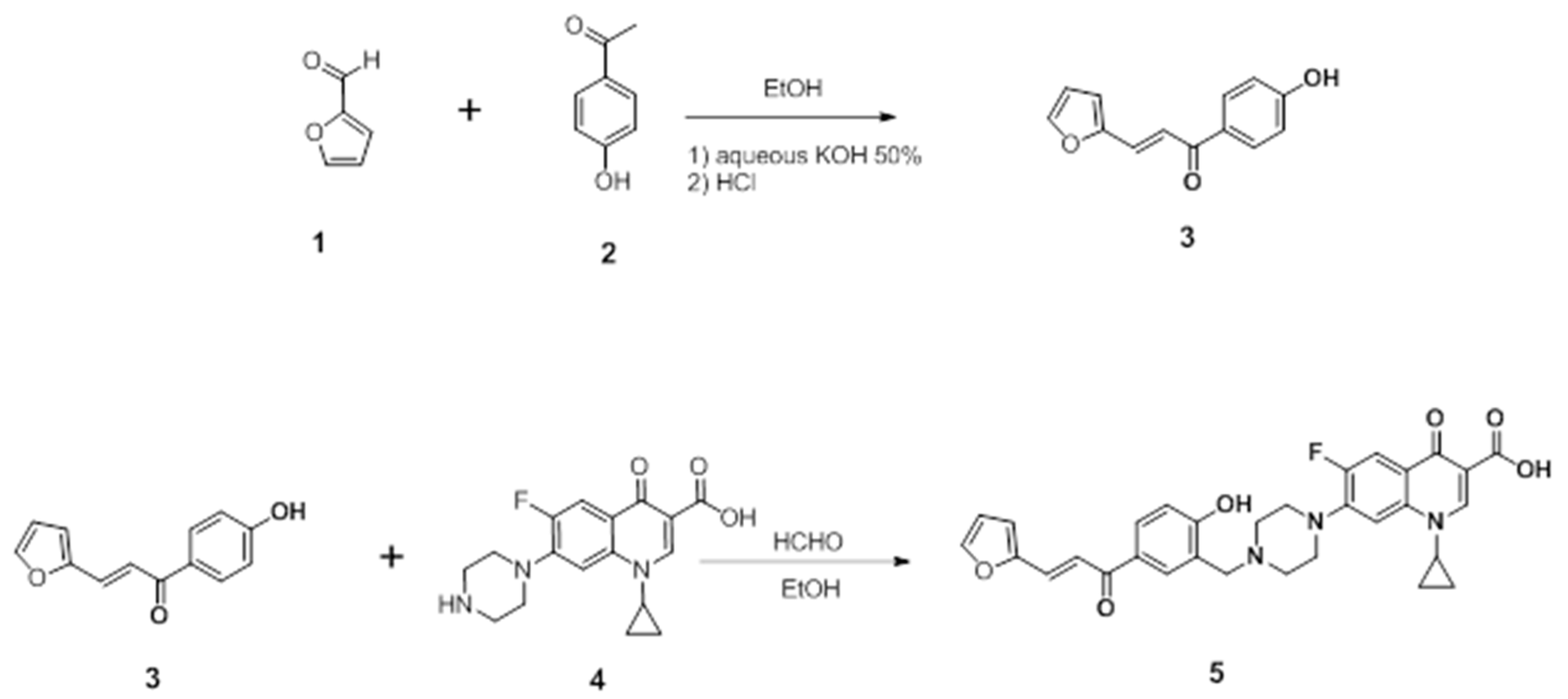
4.1. Synthesis of (E)-1-cyclopropyl-6-fluoro-7-(4-(5-(3-(furan-2-yl) acryloyl)-2 hydroxybenzyl) piperazin-1-yl)-4-oxo-1,4-dihydroquinoline-3-carboxylic acid
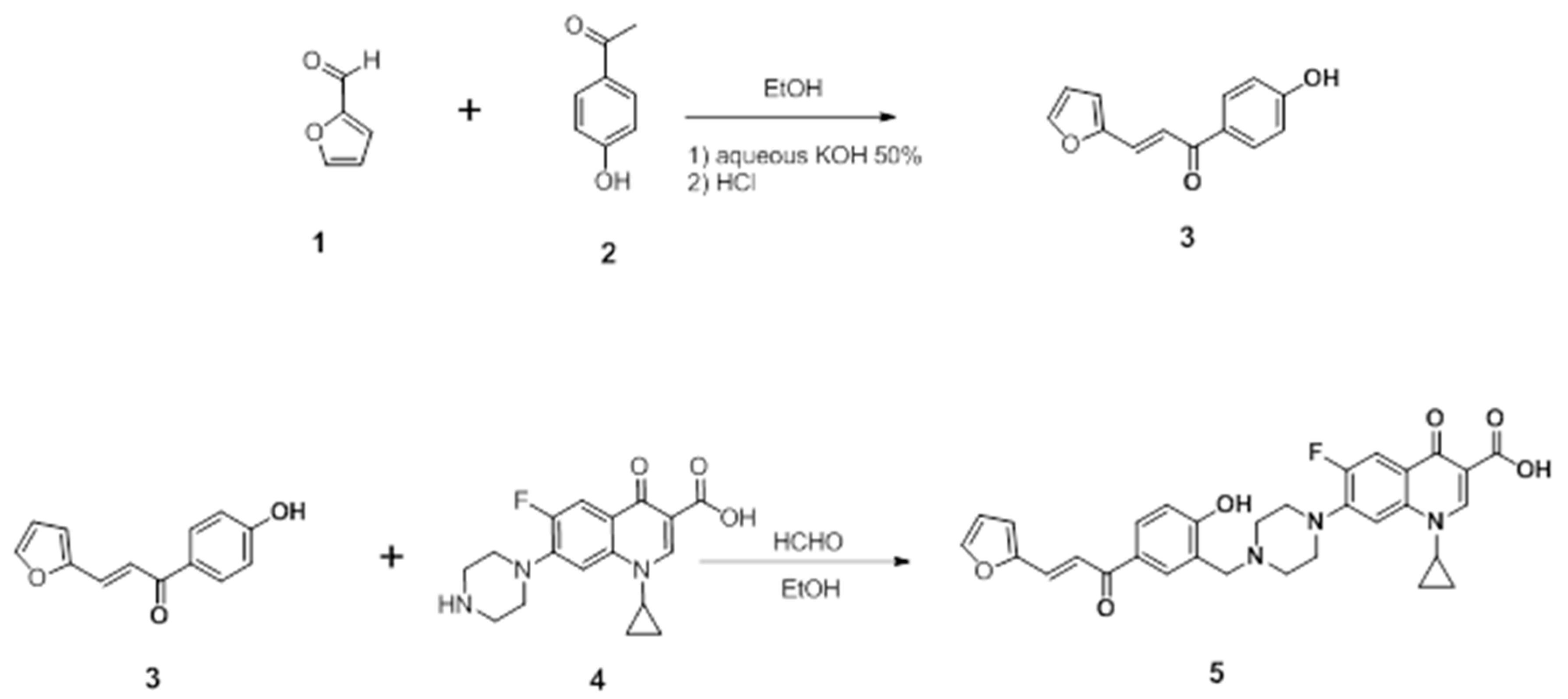
4.1.1. Preparation of the Chalcone
4.1.2. Preparation of the Mannich Base
4.2. Docking Studies of the CM Derivative on Human Topo I and Topo II Enzymes
4.3. Cell Culture
4.4. Cell Viability
4.5. Cell Migration Assay
4.6. Colony Formation Assay
4.7. Cell cycle Analysis
4.8. Annexin V assay
4.9. Evaluation the Activity on Topo I, Topo II, Bax, and bcl2
4.10. Lactate Dehydrogenase Release Assay
4.11. Analysis of Protein Expression via Western Blotting
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Gouzerh, F.; Bessière, J.-M.; Ujvari, B.; Thomas, F.; Dujon, A.M.; Dormont, L. Odors and cancer: Current status and future directions. Biochim. Et Biophys. Acta (BBA)—Rev. Cancer 2022, 1877, 188644. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Hamid, N.M.; Fathy, M.; Koike, C.; Yoshida, T.; Okabe, M.; Zho, K.; Abouzied, M.; Nikaido, T. Identification of Chemo and Radio-Resistant Sub-Population of Stem Cells in Human Cervical Cancer HeLa Cells. Cancer Investig. 2021, 39, 661–674. [Google Scholar] [CrossRef] [PubMed]
- Noto, Z.; Yoshida, T.; Okabe, M.; Koike, C.; Fathy, M.; Tsuno, H.; Tomihara, K.; Arai, N.; Noguchi, M.; Nikaido, T. CD44 and SSEA-4 positive cells in an oral cancer cell line HSC-4 possess cancer stem-like cell characteristics. Oral Oncol. 2013, 49, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Yoshida, T.; Okabe, M.; Fathy, M.; Sun, Y.; Koike, C.; Salto, S.; Nikaido, T. CD24+SSEA4+cells in Ovarian Carcinoma Cells Demonstrated the Characteristics as Cancer Stem Cells. J. Cancer Sci. Ther. 2017, 9, 343–352. [Google Scholar] [CrossRef]
- Abdellatef, A.A.; Fathy, M.; Mohammed, A.E.I.; Bakr, M.S.A.; Ahmed, A.H.; Abbass, H.S.; El-Desoky, A.H.; Morita, H.; Nikaido, T.; Hayakawa, Y. Inhibition of cell-intrinsic NF-kappaB activity and metastatic abilities of breast cancer by aloe-emodin and emodic-acid isolated from Asphodelus microcarpus. J. Nat. Med. 2021, 75, 840–853. [Google Scholar] [CrossRef]
- Fathy, M.; Nikaido, T. In vivo modulation of iNOS pathway in hepatocellular carcinoma by Nigella sativa. Env. Health Prev. Med. 2013, 18, 377–385. [Google Scholar] [CrossRef]
- Fathy, M.; Nikaido, T. In vivo attenuation of angiogenesis in hepatocellular carcinoma by Nigella sativa. Turk. J. Med. Sci. 2018, 48, 178–186. [Google Scholar] [CrossRef]
- Liang, X.; Wu, Q.; Luan, S.; Yin, Z.; He, C.; Yin, L.; Zou, Y.; Yuan, Z.; Li, L.; Song, X.; et al. A comprehensive review of topoisomerase inhibitors as anticancer agents in the past decade. Eur. J. Med. Chem. 2019, 171, 129–168. [Google Scholar] [CrossRef]
- Khadka, D.B.; Cho, W.-J. Topoisomerase inhibitors as anticancer agents: A patent update. Expert Opin. Ther. Pat. 2013, 23, 1033–1056. [Google Scholar] [CrossRef]
- Lin, C.; Chang, T.; Hsieh, W.; Hung, M.; Lin, I.; Lai, S.; Tzeng, Y. Simultaneous induction of apoptosis and necroptosis by Tanshinone IIA in human hepatocellular carcinoma HepG2 cells. Cell Death Discov. 2016, 2, 1–11. [Google Scholar] [CrossRef]
- Hooper, D.C. Mode of Action of Fluoroquinolones. Drugs 1999, 58, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Alaaeldin, R.; Mustafa, M.; Abuo-Rahma, G.E.-D.A.; Fathy, M. In vitro inhibition and molecular docking of a new ciprofloxacin-chalcone against SARS-CoV-2 main protease. Fundam. Clin. Pharmacol. 2022, 36, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Alaaeldin, R.; Abuo-Rahma, G.E.-D.A.; Zhao, Q.-L.; Fathy, M. Modulation of apoptosis and epithelial-Mesenchymal transition E-cadherin/TGF-β/Snail/TWIST pathways by a new ciprofloxacin chalcone in breast cancer cells. Anticancer Res. 2021, 41, 2383–2395. [Google Scholar] [CrossRef]
- Esfandiari Mazandaran, K.; Mirshokraee, S.A.; Didehban, K.; Houshdar Tehrani, M.H. Design, Synthesis and Biological Evaluation of Ciprofloxacin- Peptide Conjugates as Anticancer Agents. Iran. J. Pharm. Res. 2019, 18, 1823–1830. [Google Scholar] [CrossRef] [PubMed]
- Zandi, A.; Moini Zanjani, T.; Ziai, S.A.; Khazaei Poul, Y.; Haji Molla Hoseini, M. Evaluation of the cytotoxic effects of ciprofloxacin on human glioblastoma A-172 cell line. Middle East. J. Cancer 2017, 8, 119–126. [Google Scholar]
- Chrzanowska, A.; Roszkowski, P.; Bielenica, A.; Olejarz, W.; Stępień, K.; Struga, M. Anticancer and antimicrobial effects of novel ciprofloxacin fatty acids conjugates. Eur. J. Med. Chem. 2020, 185, 111810. [Google Scholar] [CrossRef] [PubMed]
- Kloskowski, T.; Gurtowska, N.; Nowak, M.; Joachimiak, R.; Bajek, A.; Olkowska, J.; Drewa, T. The influence of ciprofloxacin on viability of A549, HepG2, A375. S2, B16 and C6 cell lines in vitro. Acta Pol. Pharm. 2011, 68, 859–865. [Google Scholar] [CrossRef]
- Eisa, M.A.; Fathy, M.; Abuo-Rahma, G.; Abdel-Aziz, M.; Nazmy, M.H. Anti-Proliferative and Pro-Apoptotic Activities of Synthesized 3,4,5 Tri-Methoxy Ciprofloxacin Chalcone Hybrid, through p53 Up-Regulation in HepG2 and MCF7 Cell Lines. Asian Pac. J. Cancer Prev. 2021, 22, 3393–3404. [Google Scholar] [CrossRef]
- Eisa, M.A.; Fathy, M.; Nazmy, M.H. Potential COX2 mediated therapeutic effect of ciprofloxacin. Minia J. Med. Res. 2021, 32, 47–57. [Google Scholar] [CrossRef]
- Alaaeldin, R.; Nazmy, M.H.; Abdel-Aziz, M.; Abuo-Rahma, G.E.-D.A.; Fathy, M. Cell Cycle Arrest and Apoptotic Effect of 7-(4-(N-substituted carbamoylmethyl) piperazin-1-yl) Ciprofloxacin-derivative on HCT 116 and A549 Cancer Cells. Anticancer Res. 2020, 40, 2739–2749. [Google Scholar] [CrossRef]
- Abd-Elhamid, R.A.; Nazmy, M.H.; Fathy, M. Targeting Apoptosis as a Therapeutic Approach in Cancer. Minia J. Med. Res. 2020, 31, 321–334. [Google Scholar] [CrossRef]
- Abdel-Aal, M.A.; Abdel-Aziz, S.A.; Shaykoon, M.S.A.; Abuo-Rahma, G.E.D.A. Towards anticancer fluoroquinolones: A review article. Arch. Der Pharm. 2019, 352, 1800376. [Google Scholar] [CrossRef] [PubMed]
- Fathy, M.; Sun, S.; Zhao, Q.-L.; Abdel-Aziz, M.; Abuo-Rahma, G.E.-D.A.; Awale, S.; Nikaido, T. A New Ciprofloxacin-derivative Inhibits Proliferation and Suppresses the Migration Ability of HeLa Cells. Anticancer Res. 2020, 40, 5025–5033. [Google Scholar] [CrossRef] [PubMed]
- Fathy, M.; Khalifa, E.; Fawzy, M.A. Modulation of inducible nitric oxide synthase pathway by eugenol and telmisartan in carbon tetrachloride-induced liver injury in rats. Life Sci. 2019, 216, 207–214. [Google Scholar] [CrossRef]
- Fawzy, M.A.; Maher, S.A.; Bakkar, S.M.; El-Rehany, M.A.; Fathy, M. Pantoprazole Attenuates MAPK (ERK1/2, JNK, p38)-NF-kappaB and Apoptosis Signaling Pathways after Renal Ischemia/Reperfusion Injury in Rats. Int. J. Mol. Sci 2021, 22, 10669. [Google Scholar] [CrossRef]
- Fawzy, M.A.; Maher, S.A.; El-Rehany, M.A.; Welson, N.N.; Albezrah, N.K.A.; Batiha, G.E.-S.; Fathy, M. Vincamine Modulates the Effect of Pantoprazole in Renal Ischemia/Reperfusion Injury by Attenuating MAPK and Apoptosis Signaling Pathways. Molecules 2022, 27, 1383. [Google Scholar] [CrossRef]
- Oba, J.; Okabe, M.; Yoshida, T.; Soko, C.; Fathy, M.; Amano, K.; Kobashi, D.; Wakasugi, M.; Okudera, H. Hyperdry human amniotic membrane application as a wound dressing for a full-thickness skin excision after a third-degree burn injury. Burn. Trauma 2020, 8, tkaa014. [Google Scholar] [CrossRef]
- Okabe, M.; Yoshida, T.; Suzuki, M.; Goto, M.; Omori, M.; Taguchi, M.; Toda, A.; Suzuki, T.; Nakagawa, K.; Hiramoto, F.; et al. Hyperdry Human Amniotic Membrane (HD-AM) is Supporting Aciclovir Included Device of Poly-N-p-Vinylbenzyl-D-Lactonamide (PVLA) Sphere for Treatment of HSV-1 Infected Rabbit Keratitis Model. J. Biotechnol. Biomater. 2017, 7, 251. [Google Scholar] [CrossRef] [Green Version]
- Shytaj, I.L.; Fares, M.; Gallucci, L.; Lucic, B.; Tolba, M.M.; Zimmermann, L.; Adler, J.M.; Xing, N.; Bushe, J.; Gruber, A.D.; et al. The FDA-Approved Drug Cobicistat Synergizes with Remdesivir To Inhibit SARS-CoV-2 Replication In Vitro and Decreases Viral Titers and Disease Progression in Syrian Hamsters. mBio 2022, 13, e0370521. [Google Scholar] [CrossRef]
- Zang, X.; Song, J.; Li, Y.; Han, Y. Targeting necroptosis as an alternative strategy in tumor treatment: From drugs to nanoparticles. J. Control. Release 2022, 349, 213–226. [Google Scholar] [CrossRef]
- Xiong, K.; Qian, C.; Yuan, Y.; Wei, L.; Liao, X.; He, L.; Rees, T.W.; Chen, Y.; Wan, J.; Ji, L. Necroptosis induced by ruthenium (II) complexes as dual catalytic inhibitors of topoisomerase I/II. Angew. Chem. Int. Ed. 2020, 59, 16631–16637. [Google Scholar] [CrossRef] [PubMed]
- Hevener, K.; Verstak, T.A.; Lutat, K.E.; Riggsbee, D.L.; Mooney, J.W. Recent developments in topoisomerase-targeted cancer chemotherapy. Acta Pharm. Sin. B 2018, 8, 844–861. [Google Scholar] [CrossRef] [PubMed]
- Thakur, D.S. Topoisomerase II inhibitors in cancer treatment. Int. J. Pharm. Sci. Nanotechnol. 2010, 3, 1173–1181. [Google Scholar] [CrossRef]
- Rasheed, Z.A.; Rubin, E.H. Mechanisms of resistance to topoisomerase I-targeting drugs. Oncogene 2003, 22, 7296–7304. [Google Scholar] [CrossRef] [Green Version]
- Tugrak, M.; Yamali, C.; Sakagami, H.; Gul, H.I. Synthesis of mono Mannich bases of 2-(4-hydroxybenzylidene)-2, 3-dihydroinden-1-one and evaluation of their cytotoxicities. J. Enzym. Inhib. Med. Chem. 2016, 31, 818–823. [Google Scholar] [CrossRef] [Green Version]
- Canturk, P.; Kucukoglu, K.; Topcu, Z.; Gul, M.; Gul, H.I. Effect of some bis Mannich bases and corresponding piperidinols on DNA topoisomerase I. Arzneim. -Forsch. 2008, 58, 686–691. [Google Scholar]
- Sabra, R.T.; Abdellatef, A.A.; Abdel-Sattar, E.; Fathy, M.; Meselhy, M.R.; Hayakawa, Y. Russelioside A, a Pregnane Glycoside from Caralluma tuberculate, Inhibits Cell-Intrinsic NF-κB Activity and Metastatic Ability of Breast Cancer Cells. Biol. Pharm. Bull. 2022, 45, 1564–1571. [Google Scholar] [CrossRef]
- Sordet, O.; Khan, Q.A.; Kohn, K.W.; Pommier, Y. Apoptosis induced by topoisomerase inhibitors. Curr. Med. Chem.-Anti-Cancer Agents 2003, 3, 271–290. [Google Scholar] [CrossRef]
- Zhao, W.; Feng, H.; Sun, W.; Liu, K.; Lu, J.-J.; Chen, X. Tert-butyl hydroperoxide (t-BHP) induced apoptosis and necroptosis in endothelial cells: Roles of NOX4 and mitochondrion. Redox Biol. 2017, 11, 524–534. [Google Scholar] [CrossRef]
- Zhu, Y.; Cui, H.; Xia, Y.; Gan, H. RIPK3-mediated necroptosis and apoptosis contributes to renal tubular cell progressive loss and chronic kidney disease progression in rats. PLoS ONE 2016, 11, e0156729. [Google Scholar] [CrossRef] [Green Version]
- Alaaeldin, R.; Hassan, H.A.; Abdel-Rahman, I.M.; Mohyeldin, R.H.; Youssef, N.; Allam, A.E.; Abdelwahab, S.F.; Zhao, Q.-L.; Fathy, M. A New EGFR Inhibitor from Ficus benghalensis Exerted Potential Anti-Inflammatory Activity via Akt/PI3K Pathway Inhibition. Curr. Issues Mol. Biol. 2022, 44, 2967–2981. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Xie, X.; Ren, Y.; Shao, Z.; Zhang, N.; Li, L.; Ding, X.; Zhang, L. The role of necroptosis in disease and treatment. MedComm 2021, 2, 730–755. [Google Scholar] [CrossRef]
- Alaaeldin, R.; Abdel-Rahman, I.A.M.; Hassan, H.A.; Youssef, N.; Allam, A.E.; Abdelwahab, S.F.; Zhao, Q.-L.; Fathy, M. Carpachromene Ameliorates Insulin Resistance in HepG2 Cells via Modulating IR/IRS1/PI3k/Akt/GSK3/FoxO1 Pathway. Molecules 2021, 26, 7629. [Google Scholar] [CrossRef]
- Saeed, W.K.; Jun, D.W. Necroptosis: An emerging type of cell death in liver diseases. World J. Gastroenterol. 2014, 20, 12526–12532. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Wang, H.; Zhu, M.; Song, W.; Wang, J.; Wu, C.; Kong, Y.; Guo, J.; Li, N.; Liu, J. Ophiopogonin D′, a natural product from radix ophiopogonis, induces in vitro and in vivo RIPK1-dependent and caspase-independent apoptotic death in androgen-independent human prostate cancer cells. Front. Pharmacol. 2018, 9, 432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najafov, A.; Chen, H.; Yuan, J. Necroptosis and Cancer. Trends Cancer 2017, 3, 294–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, Z.; Yang, Z.; Xie, L.; DeWitt, J.; Chen, Y. Cancer therapy in the necroptosis era. Cell Death Differ. 2016, 23, 748–756. [Google Scholar] [CrossRef] [Green Version]
- Fathy, M.; Fawzy, M.A.; Hintzsche, H.; Nikaido, T.; Dandekar, T.; Othman, E.M. Eugenol exerts apoptotic effect and modulates the sensitivity of HeLa cells to cisplatin and radiation. Molecules 2019, 24, 3979. [Google Scholar] [CrossRef] [Green Version]
- Chou, H.L.; Fong, Y.; Wei, C.K.; Tsai, E.M.; Chen, J.Y.; Chang, W.T.; Wu, C.Y.; Huang, H.W.; Chiu, C.C. A Quinone-Containing Compound Enhances Camptothecin-Induced Apoptosis of Lung Cancer Through Modulating Endogenous ROS and ERK Signaling. Arch. Immunol. Exp. 2017, 65, 241–252. [Google Scholar] [CrossRef]
- Karki, R.; Song, C.; Kadayat, T.M.; Magar, T.B.; Bist, G.; Shrestha, A.; Na, Y.; Kwon, Y.; Lee, E.S. Topoisomerase I and II inhibitory activity, cytotoxicity, and structure-activity relationship study of dihydroxylated 2,6-diphenyl-4-aryl pyridines. Bioorg. Med. Chem. 2015, 23, 3638–3654. [Google Scholar] [CrossRef]
- Goel, A.; Prasad, A.K.; Parmar, V.S.; Ghosh, B.; Saini, N. Apoptogenic effect of 7,8-diacetoxy-4-methylcoumarin and 7,8-diacetoxy-4-methylthiocoumarin in human lung adenocarcinoma cell line: Role of NF-kappaB, Akt, ROS and MAP kinase pathway. Chem. Biol. Interact. 2009, 179, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Greenfield, E.A. Antibodies: A Laboratory Manual; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2013. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO | Compound | S a kcal/mole | RMSD_Refine b | Amino Acid Bond | Distance Å | E (Kcal mol) |
|---|---|---|---|---|---|---|
| 1 | CM derivative | −11.605 | 1.901 | Asp 533/H-donor | 3.34 | −0.7 |
| Lys 532/H-acceptor | 3.67 | −0.9 | ||||
| TGP 11/pi-pi | 3.95 | 0.0 | ||||
| DA 113/H-pi | 3.52 | −0.6 | ||||
| 2 | Camptothecin | −9.9577 | 1.390 | Asp 533/H-donor | 3.06 | −2.9 |
| Lys 532/H-acceptor | 3.10 | −3.0 | ||||
| Arg 364/H-acceptor | 3.57 | −1.6 | ||||
| TGP 11/pi-pi | 3.88 | 0.0 | ||||
| DA 113/pi-pi | 3.56 | 0.0 | ||||
| DC 112/pi-pi | 3.68 | 0.0 |
| NO | Compound | S a kcal/mole | RMSD_Refine b | Amino acid bond | Distance Å | E (Kcal mol) |
|---|---|---|---|---|---|---|
| 1 | CM derivative | −8.247 | 1.727 | Gln789/H-donor | 2.79 | −7.5 |
| Pro 819/pi-H | 4.12 | −0.5 | ||||
| DA 12/H-pi | 4.01 | −0.7 | ||||
| DT 9/H-pi | 4.33 | −0.5 | ||||
| 2 | Etoposide | −7.671 | 1.603 | Asp 479/H-donor | 3.08 | −2.3 |
| DG 13/H-donor | 3.06 | −2.0 | ||||
| DA 12/H-pi | 3.57 | −0.6 |
| Cell Lines | IC50 (Mean ± SD) (μg/mL) | |
|---|---|---|
| CM Derivative | Etoposide | |
| MCF-7 | 51.58 ± 4.82 | ND |
| HepG2 | 22.09 ± 1.7 | 34.32 ± 2.81 |
| A549 | 27.71 ± 2.4 | 48.32 ± 3.93 |
| M14 | 90.48 ± 7.4 | ND |
| Wi38 | 118.65 ± 10.3 | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alaaeldin, R.; Abdel-Rahman, I.M.; Ali, F.E.M.; Bekhit, A.A.; Elhamadany, E.Y.; Zhao, Q.-L.; Cui, Z.-G.; Fathy, M. Dual Topoisomerase I/II Inhibition-Induced Apoptosis and Necro-Apoptosis in Cancer Cells by a Novel Ciprofloxacin Derivative via RIPK1/RIPK3/MLKL Activation. Molecules 2022, 27, 7993. https://doi.org/10.3390/molecules27227993
Alaaeldin R, Abdel-Rahman IM, Ali FEM, Bekhit AA, Elhamadany EY, Zhao Q-L, Cui Z-G, Fathy M. Dual Topoisomerase I/II Inhibition-Induced Apoptosis and Necro-Apoptosis in Cancer Cells by a Novel Ciprofloxacin Derivative via RIPK1/RIPK3/MLKL Activation. Molecules. 2022; 27(22):7993. https://doi.org/10.3390/molecules27227993
Chicago/Turabian StyleAlaaeldin, Rania, Islam M. Abdel-Rahman, Fares E. M. Ali, Amany Abdlrehim Bekhit, Eyad Y. Elhamadany, Qing-Li Zhao, Zheng-Guo Cui, and Moustafa Fathy. 2022. "Dual Topoisomerase I/II Inhibition-Induced Apoptosis and Necro-Apoptosis in Cancer Cells by a Novel Ciprofloxacin Derivative via RIPK1/RIPK3/MLKL Activation" Molecules 27, no. 22: 7993. https://doi.org/10.3390/molecules27227993
APA StyleAlaaeldin, R., Abdel-Rahman, I. M., Ali, F. E. M., Bekhit, A. A., Elhamadany, E. Y., Zhao, Q.-L., Cui, Z.-G., & Fathy, M. (2022). Dual Topoisomerase I/II Inhibition-Induced Apoptosis and Necro-Apoptosis in Cancer Cells by a Novel Ciprofloxacin Derivative via RIPK1/RIPK3/MLKL Activation. Molecules, 27(22), 7993. https://doi.org/10.3390/molecules27227993