
Neuroprotective Effects of the Psychoactive Compound Biatractylolide (BD) in Alzheimer’s Disease

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. MTT Assay
2.2. Detection of MMP by Rhodamine 123 Staining
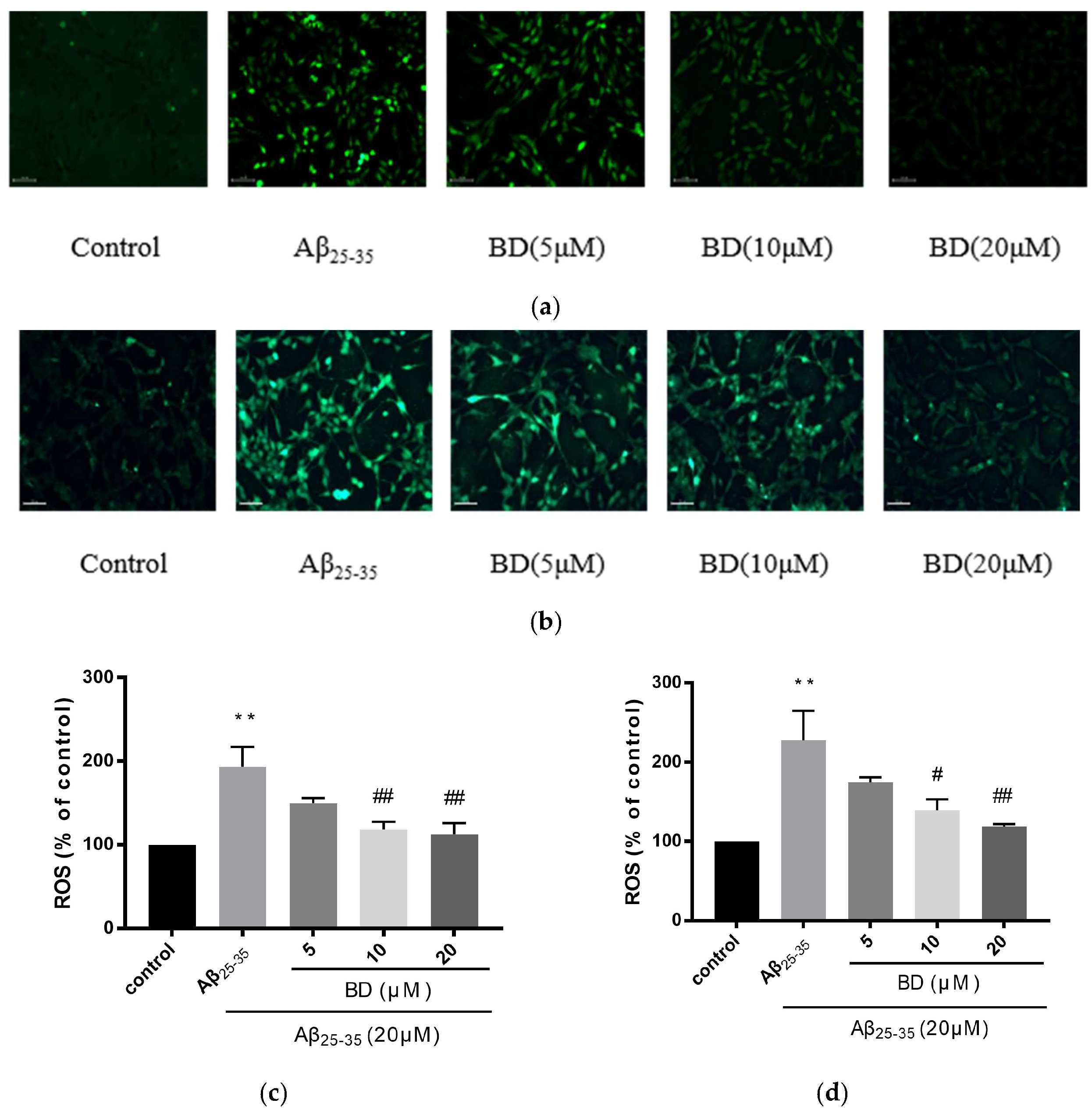
2.3. Measurement of Intracellular ROS
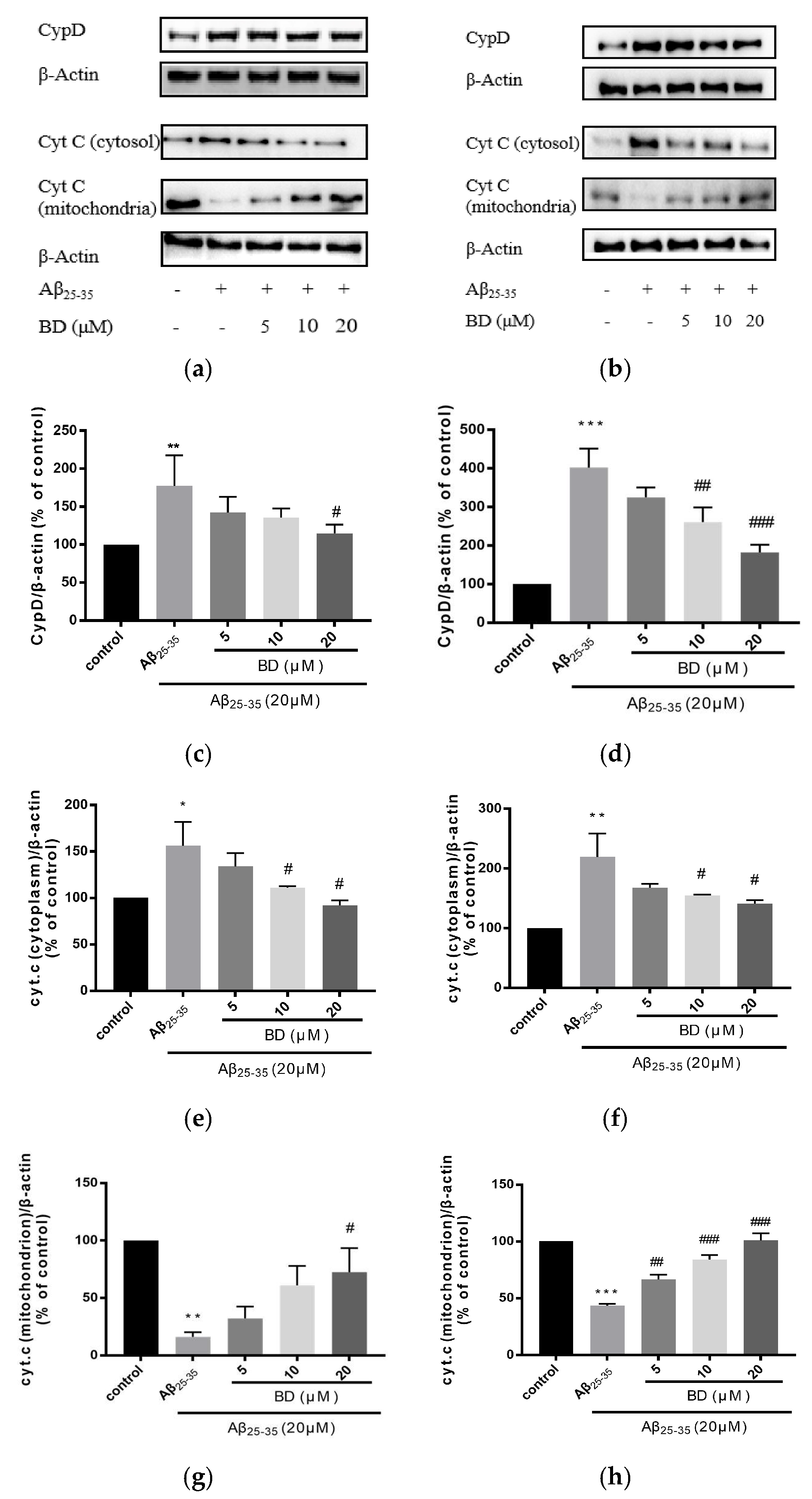
2.4. Protein Characterization
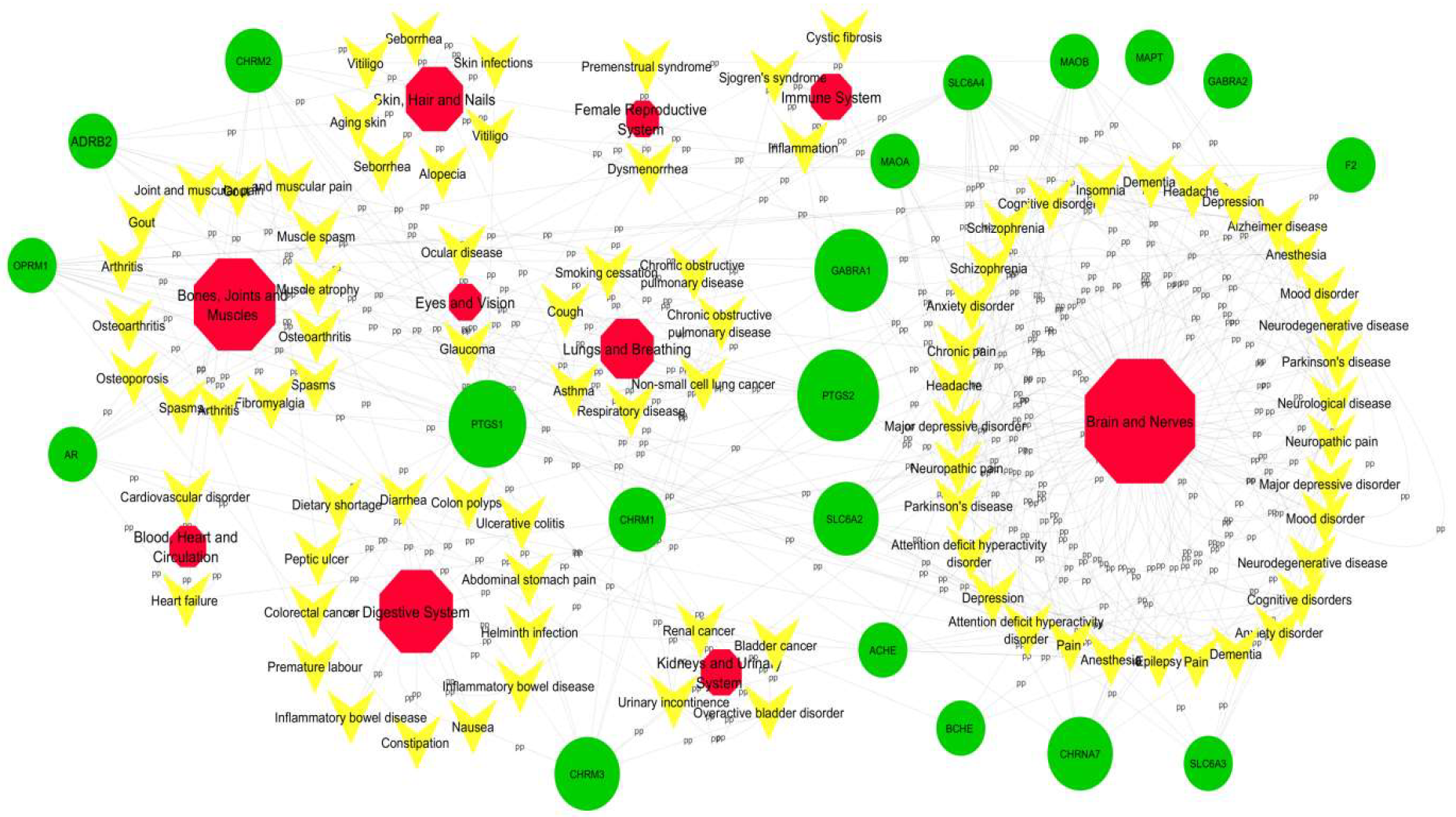
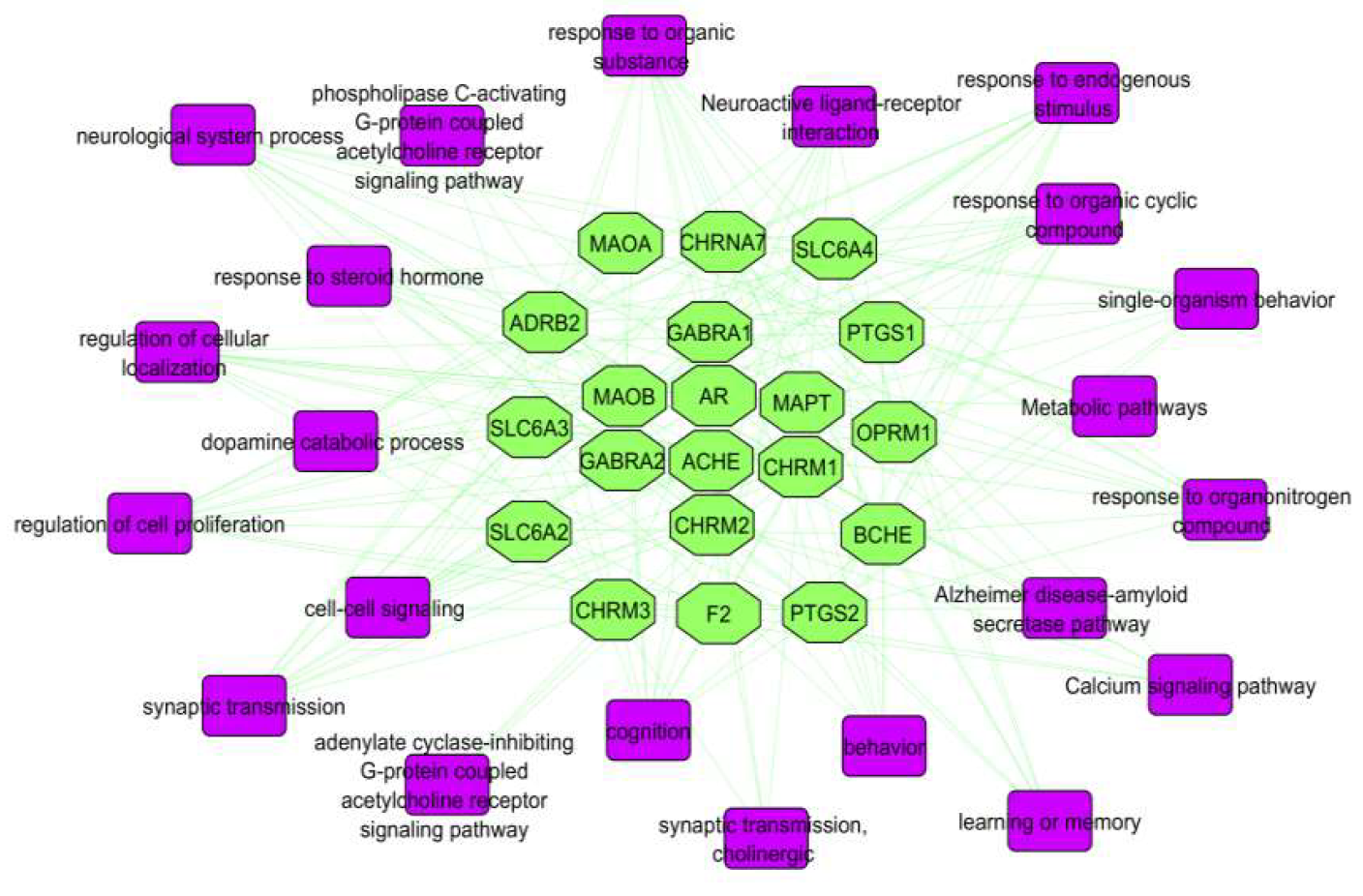
2.5. Networks
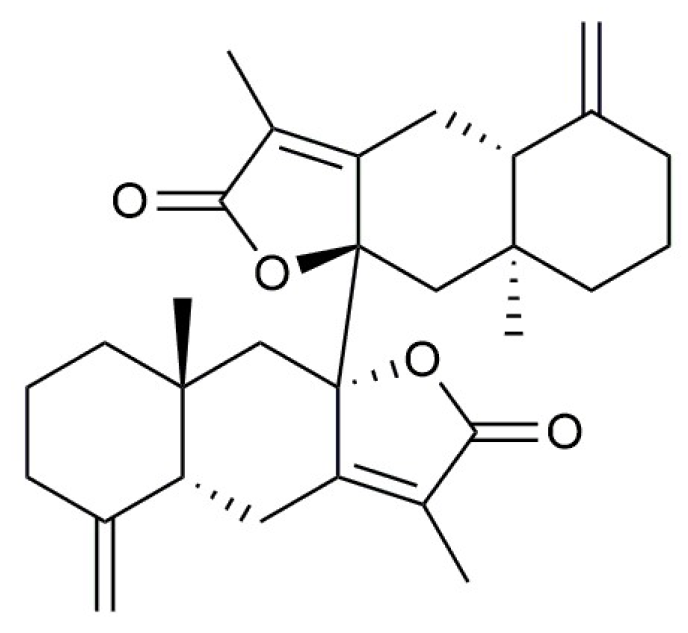
2.5.1. Active Compound Screening
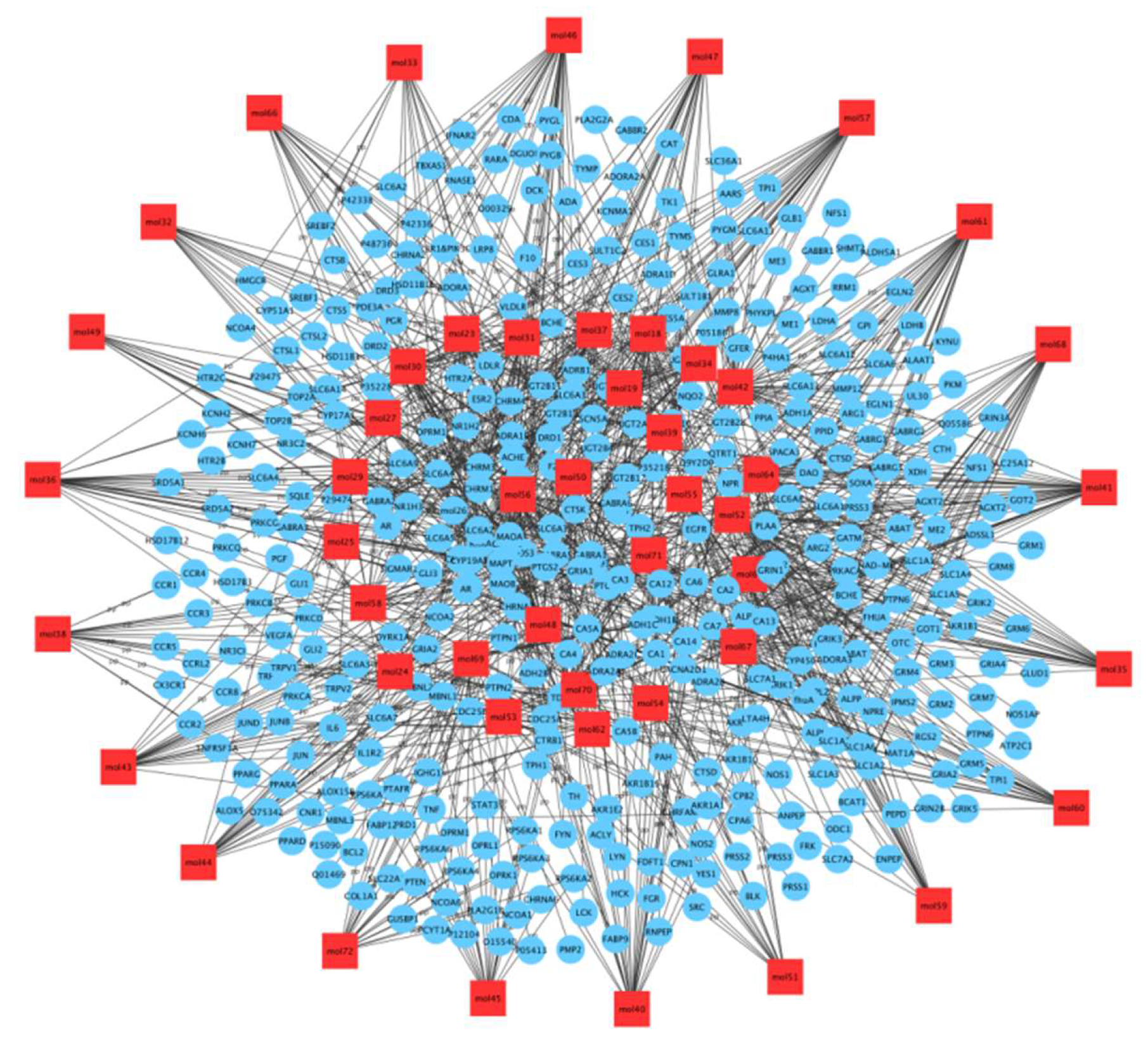
2.5.2. Network Construction and Analysis: C-T Network
2.5.3. T-D Network
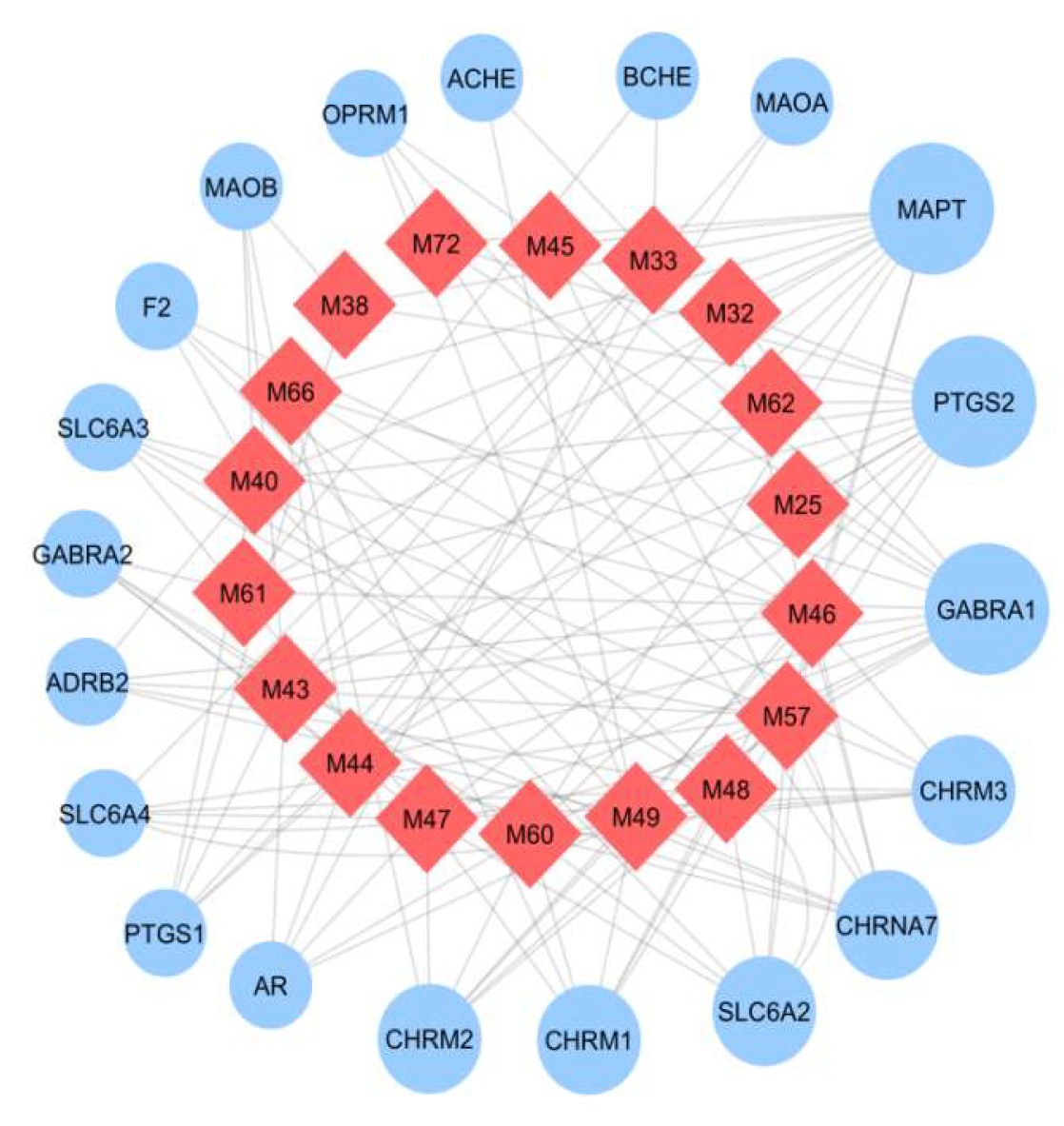
2.5.4. Target-Pathway (T-P) Network
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Data Set Construction
4.3. Active Compound Screening
4.4. Network Construction
4.5. Pathway Analysis
4.6. Cell Culture
4.7. Experimental Grouping and Drug Treatment
4.8. Cell Viability Assay (MTT)
4.9. Detection of MMP by Rhodamine 123 Staining
4.10. Measurement of Intracellular ROS
4.11. Western Blot Analysis (CYPD, CytC)
4.12. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- McKhann, G.; Drachman, D.; Folstein, M.; Katzman, R.; Price, D.; Stadlan, E.M. Clinical diagnosis of Alzheimer’s disease: Report of the NINCDS-ADRDA Work Group under the auspices of Department of Health and Human Services Task Force on Alzheimer’s Disease. Neurology 1984, 34, 939–944. [Google Scholar] [CrossRef] [Green Version]
- Erwin, W.G. Senile dementia of the Alzheimer type. Clin. Pharm. 1984, 3, 497–504. [Google Scholar] [CrossRef] [PubMed]
- Selkoe, D.; Podlisny, M.; Joachim, C.; Vickers, E.; Lee, G.; Fritz, L.; Oltersdorf, T. Beta-amyloid precursor protein of Alzheimer disease occurs as 110- to 135-kilodalton membrane-associated proteins in neural and nonneural tissues. Proc. Natl. Acad. Sci. USA 1988, 85, 7341–7345. [Google Scholar] [CrossRef] [Green Version]
- Selkoe, D.J. Aging, amyloid, and Alzheimer’s disease. N. Engl. J. Med. 1989, 320, 1484–1487. [Google Scholar] [CrossRef]
- Walsh, D.M.; Hartley, D.M.; Kusumoto, Y.; Fezoui, Y.; Condron, M.M.; Lomakin, A.; Benedek, G.B.; Selkoe, D.J.; Teplow, D.B. Amyloid beta-protein fibrillogenesis. Structure and biological activity of protofibrillar intermediates. J. Biol. Chem. 1999, 274, 25945–25952. [Google Scholar] [CrossRef] [Green Version]
- Goedert, M.; Spillantini, M.G. Molecular neuropathology of Alzheimer’s disease: In situ hybridization studies. Cell. Mol. Neurobiol. 1990, 10, 159–174. [Google Scholar] [CrossRef]
- Pereira, C.; Agostinho, P.; Moreira, P.I.; Cardoso, S.M.; Oliveira, C.R. Alzheimer’s disease-associated neurotoxic mechanisms and neuroprotective strategies. Curr. Drug Targets CNS Neurol. Disord. 2005, 4, 383–403. [Google Scholar] [CrossRef] [Green Version]
- Smith, M.A.; Drew, K.L.; Nunomura, A.; Takeda, A.; Hirai, K.; Zhu, X.; Atwood, C.S.; Raina, A.K.; Rottkamp, C.A.; Sayre, L.M.; et al. Amyloid-beta, tau alterations and mitochondrial dysfunction in Alzheimer disease: The chickens or the eggs? Neurochem. Int. 2002, 40, 527–531. [Google Scholar] [CrossRef] [PubMed]
- Thangavel, R.; Sahu, S.K.; Van Hoesen, G.W.; Zaheer, A. Loss of nonphosphorylated neurofilament immunoreactivity in temporal cortical areas in Alzheimer’s disease. Neuroscience 2009, 160, 427–433. [Google Scholar] [CrossRef] [Green Version]
- Ball, M.J. Neurofibrillary tangles in the dementia of “normal pressure” hydrocephalus. Can. J. Neurol. Sci. 1976, 3, 227–235. [Google Scholar] [CrossRef]
- Reitz, C.; Brayne, C.; Mayeux, R. Epidemiology of Alzheimer disease. Nat. Rev. Neurol. 2011, 7, 137–152. [Google Scholar] [CrossRef] [Green Version]
- Nelson, P.T.; Alafuzoff, I.; Bigio, E.H.; Bouras, C.; Braak, H.; Cairns, N.J.; Castellani, R.J.; Crain, B.J.; Davies, P.; Del Tredici, K.; et al. Correlation of Alzheimer disease neuropathologic changes with cognitive status: A review of the literature. J. Neuropathol. Exp. Neurol. 2012, 71, 362–381. [Google Scholar] [CrossRef]
- Yuyama, K.; Sun, H.; Usuki, S.; Sakai, S.; Hanamatsu, H.; Mioka, T.; Kimura, N.; Okada, M.; Tahara, H.; Furukawa, J.; et al. A potential function for neuronal exosomes: Sequestering intracerebral amyloid-β peptide. FEBS Lett. 2015, 589, 84–88. [Google Scholar] [CrossRef]
- Tillement, L.; Lecanu, L.; Papadopoulos, V. Alzheimer’s disease: Effects of β-amyloid on mitochondria. Mitochondrion 2011, 11, 13–21. [Google Scholar] [CrossRef]
- Ren, R.; Zhang, Y.; Li, B.; Wu, Y.; Li, B. Effect of β-amyloid (25-35) on mitochondrial function and expression of mitochondrial permeability transition pore proteins in rat hippocampal neurons. J. Cell. Biochem. 2011, 112, 1450–1457. [Google Scholar] [CrossRef] [PubMed]
- Sheehan, J.P.; Swerdlow, R.H.; Miller, S.W.; Davis, R.E.; Parks, J.K.; Parker, W.D.; Tuttle, J.B. Calcium homeostasis and reactive oxygen species production in cells transformed by mitochondria from individuals with sporadic Alzheimer’s disease. J. Neurosci. 1997, 17, 4612–4622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsujimoto, Y. Cell death regulation by the Bcl-2 protein family in the mitochondria. J. Cell. Physiol. 2003, 195, 158–167. [Google Scholar] [CrossRef]
- Lustbader, J.W.; Cirilli, M.; Lin, C.; Xu, H.W.; Takuma, K.; Wang, N.; Caspersen, C.; Chen, X.; Pollak, S.; Chaney, M.; et al. ABAD directly links Abeta to mitochondrial toxicity in Alzheimer’s disease. Science 2004, 304, 448–452. [Google Scholar] [CrossRef] [Green Version]
- Takuma, K.; Yao, J.; Huang, J.; Xu, H.; Chen, X.; Luddy, J.; Trillat, A.C.; Stern, D.M.; Arancio, O.; Yan, S.S. ABAD enhances Abeta-induced cell stress via mitochondrial dysfunction. Faseb J. 2005, 19, 597–598. [Google Scholar] [CrossRef]
- Du, H.; Guo, L.; Fang, F.; Chen, D.; Sosunov, A.A.; McKhann, G.M.; Yan, Y.; Wang, C.; Zhang, H.; Molkentin, J.D.; et al. Cyclophilin D deficiency attenuates mitochondrial and neuronal perturbation and ameliorates learning and memory in Alzheimer’s disease. Nat. Med. 2008, 14, 1097–1105. [Google Scholar] [CrossRef]
- Du, H.; Guo, L.; Zhang, W.; Rydzewska, M.; Yan, S. Cyclophilin D deficiency improves mitochondrial function and learning/memory in aging Alzheimer disease mouse model. Neurobiol. Aging 2011, 32, 398–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valasani, K.R.; Hu, G.; Chaney, M.O.; Yan, S.S. Structure-based design and synthesis of benzothiazole phosphonate analogues with inhibitors of human ABAD-Aβ for treatment of Alzheimer’s disease. Chem. Biol. Drug Des. 2013, 81, 238–249. [Google Scholar] [CrossRef]
- Valasani, K.R.; Chaney, M.O.; Day, V.W.; Shidu Yan, S. Acetylcholinesterase inhibitors: Structure based design, synthesis, pharmacophore modeling, and virtual screening. J. Chem. Inf. Model. 2013, 53, 2033–2046. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.A.; Perry, G.; Richey, P.L.; Sayre, L.M.; Anderson, V.E.; Beal, M.F.; Kowall, N. Oxidative damage in Alzheimer’s. Nature 1996, 382, 120–121. [Google Scholar] [CrossRef]
- Ferreiro, E.; Oliveira, C.R.; Pereira, C.M.F. The release of calcium from the endoplasmic reticulum induced by amyloid-beta and prion peptides activates the mitochondrial apoptotic pathway. Neurobiol. Dis. 2008, 30, 331–342. [Google Scholar] [CrossRef] [Green Version]
- Kroemer, G. The proto-oncogene Bcl-2 and its role in regulating apoptosis. Nat. Med. 1997, 3, 614–620. [Google Scholar] [CrossRef]
- Petrosillo, G.; Ruggiero, F.M.; Pistolese, M.; Paradies, G. Ca2+-induced reactive oxygen species production promotes cytochrome c release from rat liver mitochondria via mitochondrial permeability transition (MPT)-dependent and MPT-independent mechanisms: Role of cardiolipin. J. Biol. Chem. 2004, 279, 53103–53108. [Google Scholar] [CrossRef] [Green Version]
- Rosenstock, T.R.; Carvalho, A.C.; Jurkiewicz, A.; Frussa-Filho, R.; Smaili, S.S. Mitochondrial calcium, oxidative stress and apoptosis in a neurodegenerative disease model induced by 3-nitropropionic acid. J. Neurochem. 2004, 88, 1220–1228. [Google Scholar] [CrossRef]
- Reddy, P.H.; Beal, M.F. Amyloid beta, mitochondrial dysfunction and synaptic damage: Implications for cognitive decline in aging and Alzheimer’s disease. Trends Mol. Med. 2008, 14, 45–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rui, Y.; Tiwari, P.; Xie, Z.; Zheng, J.Q. Acute impairment of mitochondrial trafficking by beta-amyloid peptides in hippocampal neurons. J. Neurosci. 2006, 26, 10480–10487. [Google Scholar] [CrossRef] [Green Version]
- Du, H.; Guo, L.; Wu, X.; Sosunov, A.A.; McKhann, G.M.; Chen, J.X.; Yan, S.S. Cyclophilin D deficiency rescues Aβ-impaired PKA/CREB signaling and alleviates synaptic degeneration. Biochim. Biophys. Acta 2014, 1842, 2517–2527. [Google Scholar] [CrossRef] [Green Version]
- Chin, J.H.; Tse, F.W.; Harris, K.; Jhamandas, J.H. Beta-amyloid enhances intracellular calcium rises mediated by repeated activation of intracellular calcium stores and nicotinic receptors in acutely dissociated rat basal forebrain neurons. Brain Cell Biol. 2006, 35, 173–186. [Google Scholar] [CrossRef]
- Yang, X.; Yang, Y.; Li, G.; Wang, J.; Yang, E.S. Coenzyme Q10 attenuates beta-amyloid pathology in the aged transgenic mice with Alzheimer presenilin 1 mutation. J. Mol. Neurosci. 2008, 34, 165–171. [Google Scholar] [CrossRef]
- Miners, J.S.; Baig, S.; Palmer, J.; Palmer, L.E.; Kehoe, P.G.; Love, S. Abeta-degrading enzymes in Alzheimer’s disease. Brain Pathol. 2008, 18, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Schroeter, S.; Khan, K.; Barbour, R.; Doan, M.; Chen, M.; Guido, T.; Gill, D.; Basi, G.; Schenk, D.; Seubert, P.; et al. Immunotherapy reduces vascular amyloid-beta in PDAPP mice. J. Neurosci. 2008, 28, 6787–6793. [Google Scholar] [CrossRef] [Green Version]
- Chipana, C.; Camarasa, J.; Pubill, D.; Escubedo, E. Memantine prevents MDMA-induced neurotoxicity. Neurotoxicology 2008, 29, 179–183. [Google Scholar] [CrossRef]
- Diehl-Schmid, J.; Förstl, H.; Perneczky, R.; Pohl, C.; Kurz, A. A 6-month, open-label study of memantine in patients with frontotemporal dementia. Int. J. Geriatr. Psychiatry 2008, 23, 754–759. [Google Scholar] [CrossRef]
- Yang, X.; Wang, Y.; Luo, J.; Liu, S.; Yang, Z. Protective effects of YC-1 against glutamate induced PC12 cell apoptosis. Cell. Mol. Neurobiol. 2011, 31, 303–311. [Google Scholar] [CrossRef]
- Wang, K.; Zhu, X.; Zhang, K.; Zhou, F.; Zhu, L. Gas1 Knockdown Increases the Neuroprotective Effect of Glial Cell-Derived Neurotrophic Factor Against Glutamate-Induced Cell Injury in Human SH-SY5Y Neuroblastoma Cells. Cell. Mol. Neurobiol. 2016, 36, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.T.; Chen, L.G.; Chou, D.S.; Liang, W.L.; Wang, C.C. Anti-Oxidative Abilities of Essential Oils from Atractylodes ovata Rhizome. Evid.-Based Complement. Altern. Med. 2011, 2011, 204892. [Google Scholar] [CrossRef]
- Wang, K.T.; Chen, L.G.; Wu, C.H.; Chang, C.C.; Wang, C.C. Gastroprotective activity of atractylenolide III from Atractylodes ovata on ethanol-induced gastric ulcer in vitro and in vivo. J. Pharm. Pharm. 2010, 62, 381–388. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Jia, Z.; Dong, L.; Wang, R.; Qiu, G. A randomized pilot study of atractylenolide I on gastric cancer cachexia patients. Evid.-Based Complement. Altern. Med. 2008, 5, 337–344. [Google Scholar] [CrossRef]
- Zhu, L.; Ning, N.; Li, Y.; Zhang, Q.F.; Xie, Y.C.; Irshad, M.; Feng, X.; Tao, X.J. Biatractylolide Modulates PI3K-Akt-GSK3β-Dependent Pathways to Protect against Glutamate-Induced Cell Damage in PC12 and SH-SY5Y Cells. Evid.-Based Complement. Altern. Med. 2017, 2017, 1291458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, Y.C.; Ning, N.; Zhu, L.; Li, D.N.; Feng, X.; Yang, X.P. Primary Investigation for the Mechanism of Biatractylolide from Atractylodis Macrocephalae Rhizoma as an Acetylcholinesterase Inhibitor. Evid.-Based Complement. Altern. Med. 2016, 2016, 7481323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sams-Dodd, F. Target-based drug discovery: Is something wrong? Drug Discov. Today 2005, 10, 139–147. [Google Scholar] [CrossRef]
- Roth, B.L.; Sheffler, D.J.; Kroeze, W.K. Magic shotguns versus magic bullets: Selectively non-selective drugs for mood disorders and schizophrenia. Nat. Rev. Drug Discov. 2004, 3, 353–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Zhang, B. Traditional Chinese medicine network pharmacology: Theory, methodology and application. Chin. J. Nat. Med. 2013, 11, 110–120. [Google Scholar] [CrossRef]
- Lin, C.I.; Chang, Y.C.; Kao, N.J.; Lee, W.J.; Cross, T.W.; Lin, S.H. 1,25(OH)(2)D(3) Alleviates Aβ(25-35)-Induced Tau Hyperphosphorylation, Excessive Reactive Oxygen Species, and Apoptosis Through Interplay with Glial Cell Line-Derived Neurotrophic Factor Signaling in SH-SY5Y Cells. Int. J. Mol. Sci. 2020, 21, 4215. [Google Scholar] [CrossRef]
- Zhang, L.; Han, L.; Wang, X.; Wei, Y.; Zheng, J.; Zhao, L.; Tong, X. Exploring the mechanisms underlying the therapeutic effect of Salvia miltiorrhiza in diabetic nephropathy using network pharmacology and molecular docking. Biosci. Rep. 2021, 41, BSR20203520. [Google Scholar] [CrossRef] [PubMed]
- Iyengar, R.; Zhao, S.; Chung, S.W.; Mager, D.E.; Gallo, J.M. Merging systems biology with pharmacodynamics. Sci. Transl. Med. 2012, 4, 126ps127. [Google Scholar] [CrossRef]
- Su, Q.; Peng, M.; Zhang, Y.; Xu, W.; Darko, K.O.; Tao, T.; Huang, Y.; Tao, X.; Yang, X. Quercetin induces bladder cancer cells apoptosis by activation of AMPK signaling pathway. Am. J. Cancer Res. 2016, 6, 498–508. [Google Scholar] [PubMed]
- Modi, K.K.; Jana, A.; Ghosh, S.; Watson, R.; Pahan, K. A physically-modified saline suppresses neuronal apoptosis, attenuates tau phosphorylation and protects memory in an animal model of Alzheimer’s disease. PLoS ONE 2014, 9, e103606. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.S.; Liu, W.; Lu, G.X. miR-200a-3p promotes b-Amyloid-induced neuronal apoptosis through down-regulation of SIRT1 in Alzheimer’s disease. J. Biosci. 2017, 42, 397–404. [Google Scholar] [CrossRef]
- Siedlak, S.L.; Casadesus, G.; Webber, K.M.; Pappolla, M.A.; Atwood, C.S.; Smith, M.A.; Perry, G. Chronic antioxidant therapy reduces oxidative stress in a mouse model of Alzheimer’s disease. Free Radic. Res. 2009, 43, 156–164. [Google Scholar] [CrossRef] [Green Version]
- Smoot, M.E.; Ono, K.; Ruscheinski, J.; Wang, P.L.; Ideker, T. Cytoscape 2.8: New features for data integration and network visualization. Bioinformatics 2011, 27, 431–432. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, Q.; Wang, J.; Irshad, M.; Mao, S.; Chen, H.; Song, Y.; Xu, X.; Feng, X. Neuroprotective Effects of the Psychoactive Compound Biatractylolide (BD) in Alzheimer’s Disease. Molecules 2022, 27, 8294. https://doi.org/10.3390/molecules27238294
Hu Q, Wang J, Irshad M, Mao S, Chen H, Song Y, Xu X, Feng X. Neuroprotective Effects of the Psychoactive Compound Biatractylolide (BD) in Alzheimer’s Disease. Molecules. 2022; 27(23):8294. https://doi.org/10.3390/molecules27238294
Chicago/Turabian StyleHu, Qianmei, Jixiang Wang, Maida Irshad, Siyu Mao, Hongting Chen, Yujiao Song, Xuan Xu, and Xing Feng. 2022. "Neuroprotective Effects of the Psychoactive Compound Biatractylolide (BD) in Alzheimer’s Disease" Molecules 27, no. 23: 8294. https://doi.org/10.3390/molecules27238294