

Biological Investigation and Chemical Study of Brassica villosa subsp. drepanensis (Brassicaeae) Leaves

 ,
,  ,
,  ,
,  ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
2. Results
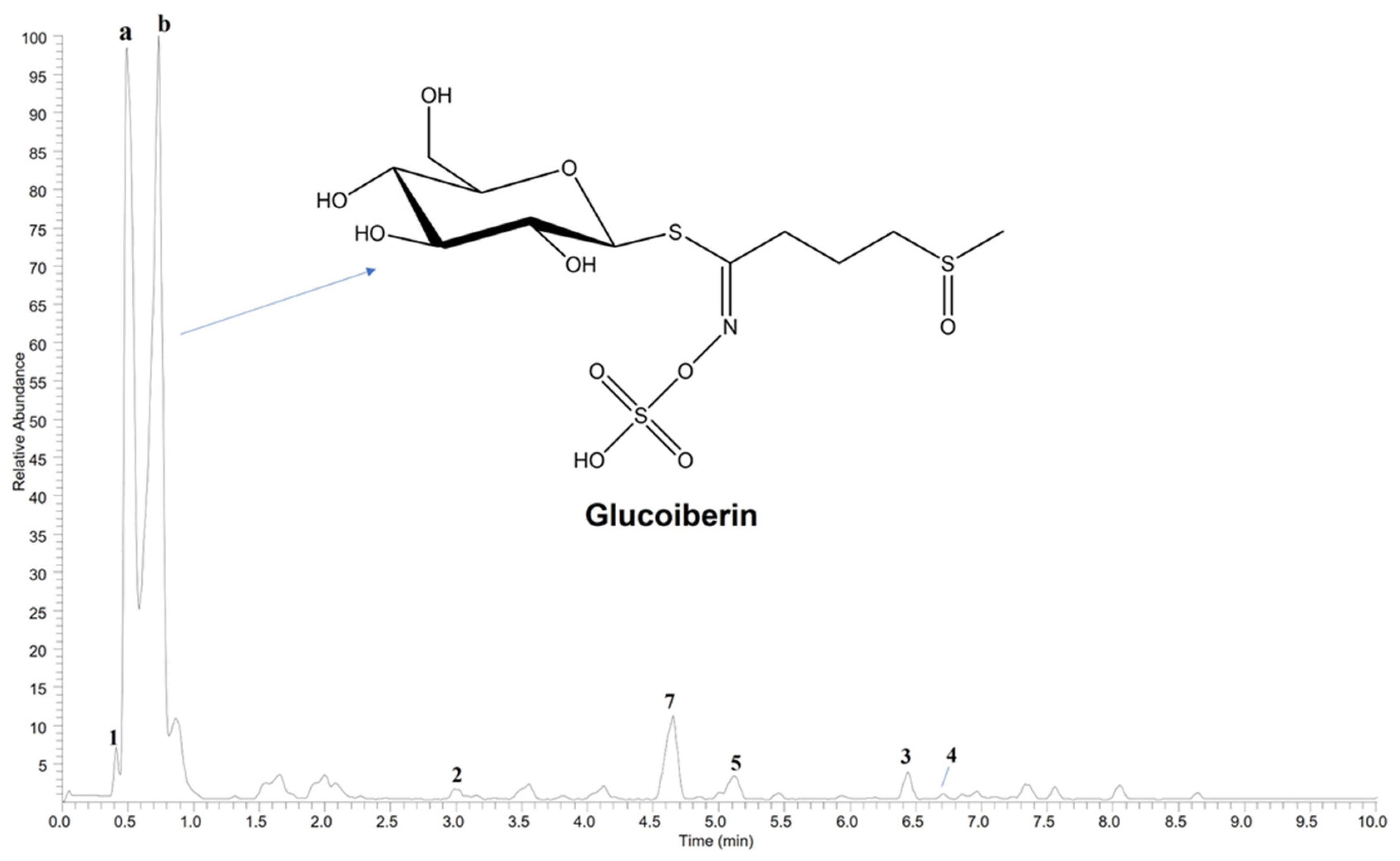
2.1. Phytochemical Analysis
2.2. In Vitro Cell Free Antioxidant Assay
2.3. Cell Viability
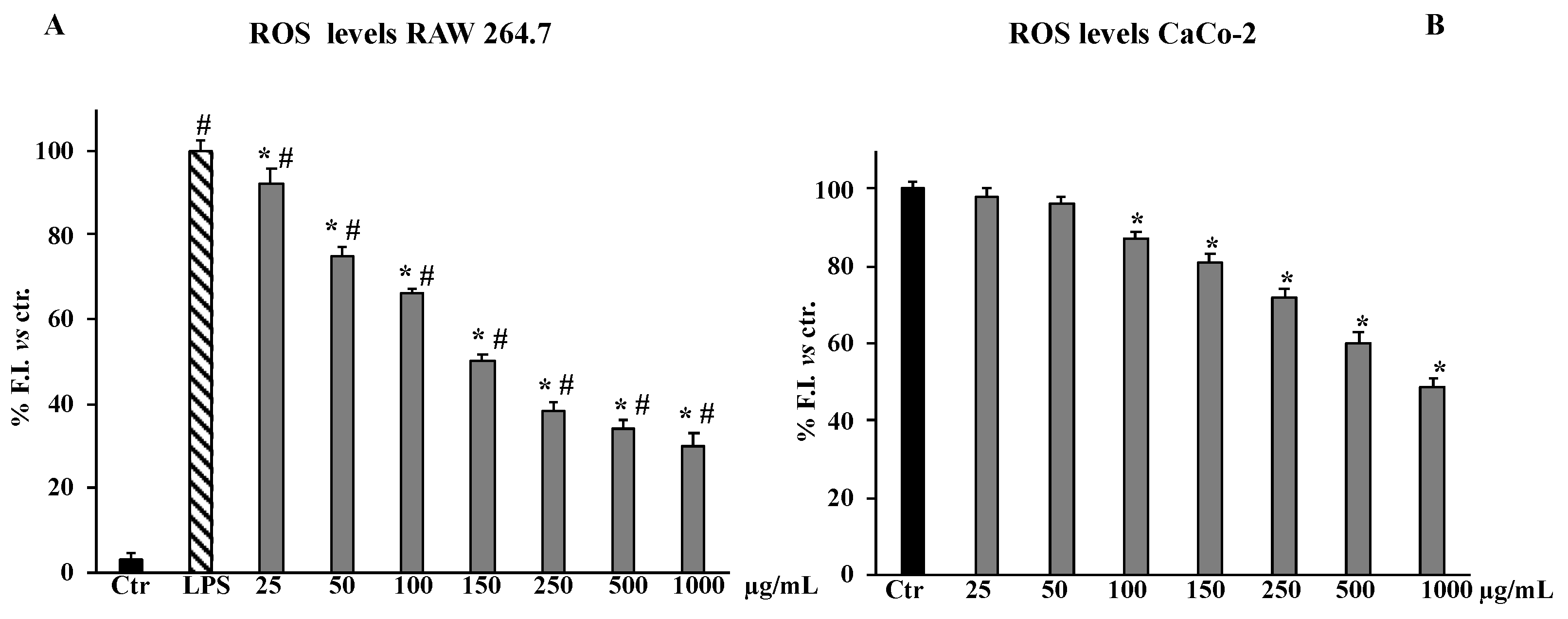
2.4. ROS Levels
2.5. Total Thiol Groups
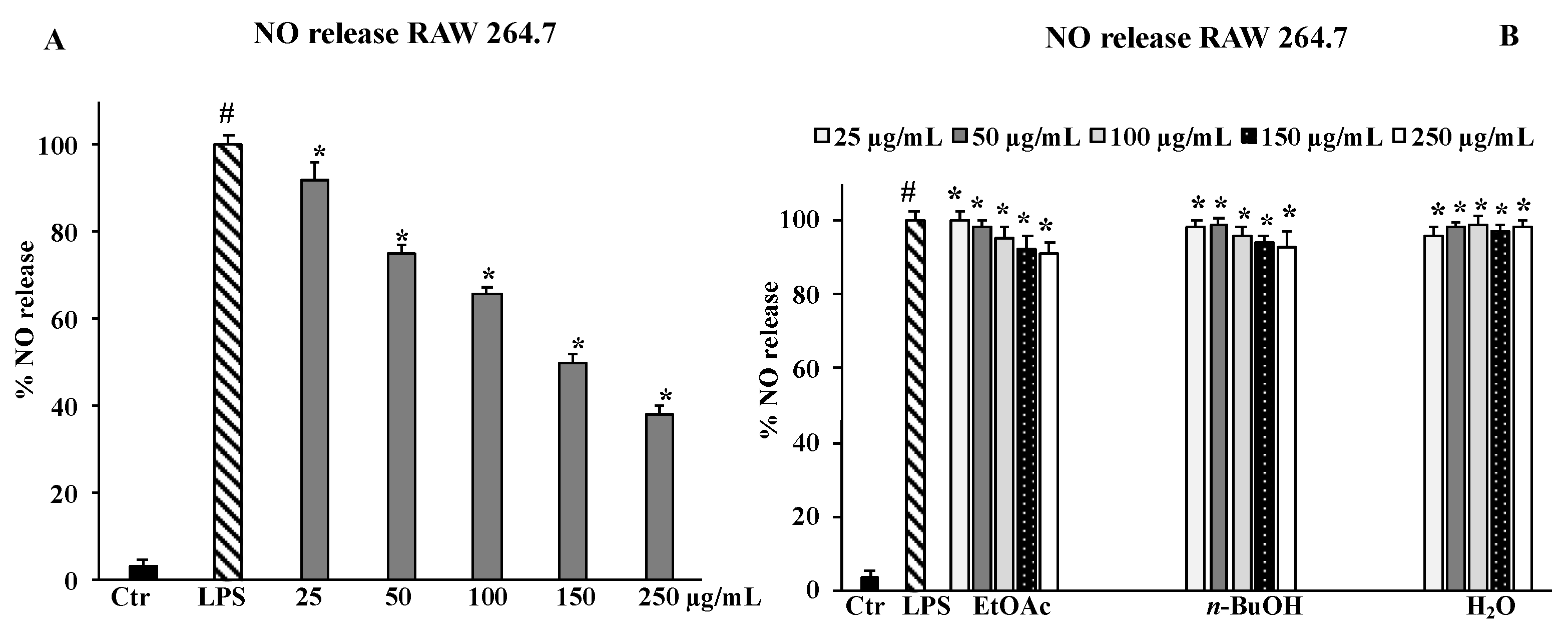
2.6. NO Release
2.7. Enzymatic Inhibitory Activity Test
2.8. Cytotoxicity in Cancer Cell Line
2.9. Total Phenol and Flavonoid Content
3. Discussion
4. Materials and Methods
4.1. Chemicals, Apparatus, and Reagents
4.2. Plant Collection and Extraction Procedure
4.3. Determination of Total Phenolic Content
4.4. Isolation of Pure Compounds
4.5. SOD-like Activity
4.6. DPPH Test
4.7. ABTS Assay
4.8. Ferric Reducing Activity Power (FRAP) Assay
4.9. Carotene Bleaching Assay
4.10. Cell Culture and Treatments
4.11. MTT Assay
4.12. Reactive Oxygen Species Assay
4.13. Total Thiol Group Determination
4.14. Measurement of NO· Release
4.15. Pancreatic Lipase Inhibitory Activity Test
4.16. Amylase Inhibitory Activity Test
4.17. α-Glucosidase Inhibitory Activity Test
4.18. LDH Assay
4.19. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mason, A.S.; Batley, J. Creating new interspecific hybrid and polyploid crops. Trends Biotechnol. 2015, 33, 436–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H. Discovery and domestication of new fruit trees in the 21st century. Plants 2022, 11, 2107. [Google Scholar] [CrossRef] [PubMed]
- Malfa, G.A.; Acquaviva, R.; Bucchini, A.A.E.; Ragusa, S.; Raimondo, F.M.; Spadaro, V. The Sicilian wild cabbages as biological resources: Taxonomic update and a review on chemical constituents and biological activities. Fl. Medit. 2020, 30, 245–260. [Google Scholar] [CrossRef]
- Kapusta-Duch, J.; Kopeć, A.; Piatkowska, E.; Borczak, B.; Leszczyńska, T. The beneficial effects of Brassica vegetables on human health. Rocz. Panstw. Zakl. Hig. 2012, 63, 389–395. [Google Scholar]
- Chang, M.X.; Xiong, F. Astaxanthin and its effects in inflammatory responses and inflammation-associated diseases: Recent advances and future directions. Molecules 2020, 25, 5342. [Google Scholar] [CrossRef]
- Libby, P. Inflammatory mechanisms: The molecular basis of inflammation and disease. Nutr. Rev. 2007, 65, S140–S146. [Google Scholar] [CrossRef]
- Sharma, M.; Majumdar, P.K. Occupational lifestyle diseases: An emerging issue. Indian J. Occup. Environ. Med. 2009, 13, 109–112. [Google Scholar] [CrossRef] [Green Version]
- Venter, C.; Eyerich, S.; Sarin, T.; Klatt, K.C. Nutrition and the immune system: A complicated tango. Nutrients 2020, 12, 818. [Google Scholar] [CrossRef] [Green Version]
- Bordoni, A.; Danesi, F.; Dardevet, D.; Dupont, D.; Fernandez, A.S.; Gille, D.; dos Santos, C.D.; Pinto, P.; Re, R.; Rémond, D.; et al. Dairy products and inflammation: A review of the clinical evidence. Crit. Rev. Food Sci. Nutr. 2017, 57, 2497–2525. [Google Scholar] [CrossRef]
- Cartea, M.E.; Francisco, M.; Soengas, P.; Velasco, P. Phenolic compounds in Brassica vegetables. Molecules 2010, 16, 251–280. [Google Scholar] [CrossRef]
- Cartea, M.E.; Velasco, P. Glucosinolates in Brassica foods: Bioavailability in food and significance for human health. Phytochem. Rev. 2008, 7, 213–229. [Google Scholar] [CrossRef]
- Raimondo, F.M.; Mazzola, P.; Ottonello, D. On the taxonomy and distribution of Brassica sect. Brassica (Cruciferae) in Sicily. Fl. Medit. 1991, 1, 63–86. [Google Scholar]
- Faulkner, K.; Mithen, R.; Williamson, G. Selective increase of the potential anticarcinogen 4-methylsulphinylbutyl glucosinolate in broccoli. Carcinogenesis 1998, 19, 605–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, C.; Fu, R.; Hu, J.; Hou, L.; Ding, S. Carbon-13 chemical shift anisotropies of solid amino acids. Magn. Reason. Chem. 1993, 31, 699–704. [Google Scholar] [CrossRef]
- Otsuka, H.; Yao, M.; Kamada, K.; Takeda, Y. Alangionosides G-M: Glycosides of megastigmane derivatives from the leaves of Alangium premnifolium. Chem. Pharm. Bull. 1995, 43, 754–759. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.W.; Lee, D.Y.; Baek, D.R.; Jeong, R.H.; Lee, D.S.; Kim, Y.C.; Kim, G.S.; Baek, N.I.; Lee, Y.H. Phenylpropanoids from red kohlrabi sprouts inhibits nitric oxide production in RAW 264.7 macrophage cells. Food Sci. Biotechnol. 2014, 23, 965–969. [Google Scholar] [CrossRef]
- Baumert, A.; Milkowski, C.; Schmidt, J.; Nimtz, M.; Wray, V.; Strack, D. Formation of a complex pattern of sinapate esters in Brassica napus seeds, catalyzed by enzymes of a serine carboxypeptidase-like acyltransferase family? Phytochemistry 2005, 66, 1334–1345. [Google Scholar] [CrossRef]
- Hruska, F.E.; Blonski, W.J.P. A 1H and 13C nuclear magnetic resonance study of nucleosides with methylated pyrimidine bases. Can. J. Chem. 1982, 60, 3026–3032. [Google Scholar] [CrossRef] [Green Version]
- Blackburn, B.J.; Lapper, R.D.; Smith, I.C. Proton magnetic resonance study of the conformations of 3′,5′-cyclic nucleotides. J. Chem. Soc. 1973, 95, 2873–2878. [Google Scholar] [CrossRef]
- Clarke, D.B. Glucosinolates, structures and analysis in food. Anal. Methods 2010, 2, 301–416. [Google Scholar] [CrossRef]
- Velasco, P.; Francisco, M.; Moreno, D.A.; Ferreres, F.; García-Viguera, C.; Cartea, M.E. Phytochemical fingerprinting of vegetable Brassica oleracea and Brassica napus by simultaneous identification of glucosinolates and phenolics. Phytochem. Anal. 2011, 22, 144–152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floegel, A.; Kim, D.O.; Chung, S.J.; Koo, S.I.; Chun, O.K. Comparison of ABTS/DPPH assays to measure antioxidant capacity in popular antioxidant-rich US foods. J. Food Compos. Anal. 2001, 24, 1043–1048. [Google Scholar] [CrossRef]
- Wang, J.; Fang, X.; Ge, L.; Cao, F.; Zhao, L.; Wang, Z.; Xiao, W. Antitumor, antioxidant and anti-inflammatory activities of kaempferol and its corresponding glycosides and the enzymatic preparation of kaempferol. PLoS ONE 2018, 13, e0197563. [Google Scholar] [CrossRef] [PubMed]
- Cavò, E.; Taviano, M.F.; Davì, F.; Cacciola, F.; Oulad El Majdoub, Y.; Mondello, L.; Ragusa, M.; Condurso, C.; Merlino, M.; Verzera, A.; et al. Phenolic and volatile composition and antioxidant properties of the leaf extract of Brassica fruticulosa subsp. fruticulosa (Brassicaceae) growing wild in Sicily (Italy). Molecules 2022, 27, 2768. [Google Scholar] [CrossRef] [PubMed]
- Martini, D. Health benefits of Mediterranean diet. Nutrients 2019, 11, 1802. [Google Scholar] [CrossRef] [Green Version]
- Tosti, V.; Bertozzi, B.; Fontana, L. Health benefits of the Mediterranean diet: Metabolic and molecular mechanisms. J. Gerontol. A Biol. Sci. Med. Sci. 2018, 73, 318–326. [Google Scholar] [CrossRef] [Green Version]
- Alì, S.; Davinelli, S.; Accardi, G.; Aiello, A.; Caruso, C.; Duro, G.; Ligotti, M.E.; Pojero, F.; Scapagnini, G.; Candore, G. Healthy ageing and Mediterranean diet: A focus on hormetic phytochemicals. Mech. Ageing Dev. 2021, 200, 111592. [Google Scholar] [CrossRef]
- Aboussaleh, Y.; Capone, R.; Bilali, H.E. Mediterranean food consumption patterns: Low environmental impacts and significant health-nutrition benefits. Proc. Nutr. Soc. 2017, 76, 543–548. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khansari, N.; Shakiba, Y.; Mahmoudi, M. Chronic inflammation and oxidative stress as a major cause of age-related diseases and cancer. Recent Pat. Inflamm. Allergy Drug Discov. 2009, 3, 73–80. [Google Scholar] [CrossRef]
- Mattosinhos, P.D.S.; Sarandy, M.M.; Novaes, R.D.; Esposito, D.; Gonçalves, R.V. Anti-inflammatory, antioxidant, and skin regenerative potential of secondary metabolites from plants of the Brassicaceae family: A systematic review of in vitro and in vivo preclinical evidence (biological activities Brassicaceae skin diseases). Antioxidants 2022, 11, 1346. [Google Scholar] [CrossRef]
- Miceli, N.; Cavò, E.; Ragusa, M.; Cacciola, F.; Mondello, L.; Dugo, L.; Acquaviva, R.; Malfa, G.A.; Marino, A.; D’Arrigo, M.; et al. Brassica incana Ten. (Brassicaceae): Phenolic constituents, antioxidant and cytotoxic properties of the leaf and flowering top extracts. Molecules 2020, 25, 1461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Picchi, V.; Lo Scalzo, R.; Tava, A.; Doria, F.; Argento, S.; Toscano, S.; Treccarichi, S.; Branca, F. Phytochemical characterization and in vitro antioxidant properties of four Brassica wild species from Italy. Molecules 2020, 25, 3495. [Google Scholar] [CrossRef] [PubMed]
- Schlesier, K.; Harwat, M.; Böhm, V.; Bitsch, R. Assessment of antioxidant activity by using different in vitro methods. Free Radic. Res. 2002, 36, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Abarca-Vargas, R.; Petricevich, V.L. Comparison of different extraction methods for the phenolic compounds recovery with the antioxidant activity of Bougainvillea × buttina. Curr. Anal. Chem. 2020, 16, 778–787. [Google Scholar] [CrossRef]
- Oishi, Y.; Manabe, I. Macrophages in inflammation, repair and regeneration. Int. Immunol. 2018, 30, 511–528. [Google Scholar] [CrossRef] [Green Version]
- Eming, S.A.; Wynn, T.A.; Martin, P. Inflammation and metabolism in tissue repair and regeneration. Science 2017, 356, 1026–1030. [Google Scholar] [CrossRef] [Green Version]
- Noworyta-Sokołowska, K.; Górska, A.; Gołembiowska, K. LPS-induced oxidative stress and inflammatory reaction in the rat striatum. Pharmacol. Rep. 2013, 65, 863–869. [Google Scholar] [CrossRef]
- Malfa, G.A.; Tomasello, B.; Acquaviva, R.; La Mantia, A.; Pappalardo, F.; Ragusa, M.; Renis, M.; Di Giacomo, C. The antioxidant activities of Betula etnensis Rafin. ethanolic extract exert protective and anti-diabetic effects on streptozotocin-induced diabetes in rats. Antioxidants 2020, 9, 847. [Google Scholar] [CrossRef]
- Nani, A.; Murtaza, B.; Sayed Khan, A.; Khan, N.A.; Hichami, A. Antioxidant and anti-inflammatory potential of polyphenols contained in Mediterranean diet in obesity: Molecular mechanisms. Molecules 2021, 26, 985. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [Green Version]
- Calabrese, G.; Zappalà, A.; Dolcimascolo, A.; Acquaviva, R.; Parenti, R.; Malfa, G.A. Phytochemical analysis and anti-inflammatory and anti-osteoarthritic bioactive potential of Verbascum thapsus L. (Scrophulariaceae) leaf extract evaluated in two in vitro models of inflammation and osteoarthritis. Molecules 2021, 26, 5392. [Google Scholar] [CrossRef] [PubMed]
- Caesar, L.K.; Cech, N.B. Synergy and antagonism in natural product extracts: When 1 + 1 does not equal 2. Nat. Prod. Rep. 2019, 36, 869–888. [Google Scholar] [CrossRef] [PubMed]
- Rohm, T.V.; Meier, D.T.; Olefsky, J.M.; Donath, M.Y. Inflammation in obesity, diabetes, and related disorders. Immunity 2022, 55, 31–55. [Google Scholar] [CrossRef] [PubMed]
- Zielińska-Bliźniewska, H.; Sitarek, P.; Merecz-Sadowska, A.; Malinowska, K.; Zajdel, K.; Jabłońska, M.R.; Śliwiński, T.; Zajdel, R. Plant extracts and reactive oxygen species as two counteracting agents with anti- and pro-obesity properties. Int. J. Mol. Sci. 2019, 20, 4556. [Google Scholar] [CrossRef] [Green Version]
- Nallamuthu, I.; Jain, A.; Anand, T. Comparative evaluation of Brassica oleracea, Ocimum basilicum, and Moringa oleifera leaf extracts on lipase inhibition and adipogenesis in 3T3-L1 adipocytes. J. Food Biochem. 2022, 46, e14158. [Google Scholar] [CrossRef]
- Connolly, E.L.; Sim, M.; Travica, N.; Marx, W.; Beasy, G.; Lynch, G.S.; Bondonno, C.P.; Lewis, J.R.; Hodgson, J.M.; Blekkenhorst, L.C. Glucosinolates from cruciferous vegetables and their potential role in chronic disease: Investigating the preclinical and clinical evidence. Front. Pharmacol. 2021, 12, 767975. [Google Scholar] [CrossRef]
- Malfa, G.A.; Tomasello, B.; Acquaviva, R.; Genovese, C.; La Mantia, A.; Cammarata, F.P.; Ragusa, M.; Renis, M.; Di Giacomo, C. Betula etnensis Raf. (Betulaceae) extract induced HO-1 expression and ferroptosis cell death in human colon cancer cells. Int. J. Mol. Sci. 2019, 20, 2723. [Google Scholar] [CrossRef] [Green Version]
- Forman, H.J.; Zhang, H. Targeting oxidative stress in disease: Promise and limitations of antioxidant therapy. Nat. Rev. Drug Discov. 2021, 20, 689–709. [Google Scholar] [CrossRef]
- Cháirez-Ramírez, M.H.; de la Cruz-López, K.G.; García-Carrancá, A. Polyphenols as antitumor agents targeting key players in cancer-driving signaling pathways. Front. Pharmacol. 2021, 12, 710304. [Google Scholar] [CrossRef]
- Politi, M.; Ferrante, C.; Menghini, L.; Angelini, P.; Flores, G.A.; Muscatello, B.; Braca, A.; De Leo, M. Hydrosols from Rosmarinus officinalis, Salvia officinalis, and Cupressus sempervirens: Phytochemical analysis and bioactivity evaluation. Plants 2022, 11, 349. [Google Scholar] [CrossRef]
- Acquaviva, R.; D’Angeli, F.; Malfa, G.A.; Ronsisvalle, S.; Garozzo, A.; Stivala, A.; Ragusa, S.; Nicolosi, D.; Salmeri, M.; Genovese, C. Antibacterial and anti-biofilm activities of walnut pellicle extract (Juglans regia L.) against coagulase-negative staphylococci. Nat. Prod. Res. 2021, 35, 2076–2081. [Google Scholar] [CrossRef] [PubMed]
- Salerno, L.; Modica, M.N.; Pittalà, V.; Romeo, G.; Siracusa, M.A.; Di Giacomo, C.; Sorrenti, V.; Acquaviva, R. Antioxidant activity and phenolic content of microwave-assisted Solanum melongena extracts. Sci. World J. 2014, 2014, 315473. [Google Scholar] [CrossRef]
- Loizzo, M.R.; Pacetti, D.; Lucci, P.; Oscar Núñez, O.; Menichini, F.; Frega, N.G.; Tundis, R. Prunus persica var. platycarpa (Tabacchiera peach): Bioactive compounds and antioxidant activity of pulp, peel and seed ethanolic extracts. Plant Foods Hum. Nutr. 2015, 70, 331–337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tundis, R.; Conidi, C.; Loizzo, M.R.; Sicari, V.; Romeo, R.; Cassano, A. Concentration of bioactive phenolic compounds in olive mill wastewater by direct contact membrane distillation. Molecules 2021, 26, 1808. [Google Scholar] [CrossRef] [PubMed]
- Tenuta, M.C.; Deguin, B.; Loizzo, M.R.; Dugay, A.; Acquaviva, R.; Malfa, G.A.; Bonesi, M.; Bouzidi, C.; Tundis, R. Contribution of flavonoids and iridoids to the hypoglycaemic, antioxidant, and nitric oxide (NO) inhibitory activities of Arbutus unedo L. Antioxidants 2020, 9, 184. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| B. drepanensis Extract | ABTS Test IC50 (μg/mL) | FRAP Test IC50 (μMfe (II)/g) | DPPH Test IC50 (μg/mL) | SOD-like Activity IC50 (μg/mL) | β-Carotene Bleaching Test IC50 (μg/mL) | |
|---|---|---|---|---|---|---|
| EtOH-H2O | 3.32 ± 0.88 * | 10.46 ± 1.17 * | 368 ± 1.7 * | 66.66 ± 1.1 * | 38.34 ± 2.51 * | 47.95 ± 2.64 * |
| n-BuOH | 2.5 ± 1.1 * | 7.09 ± 1.8 * | 301 ± 2.3 * | 40.03 ± 1.2 * | 29.04 ± 3.1 * | 35.60 ± 1.4 * |
| EtOAc | >50 | >150 | >700 | >200 | N.D. | N.D. |
| H2O | >50 | >150 | >700 | >200 | N.D. | N.D. |
| Positive control | ||||||
| Ascorbic acid | 1.73 ± 0.06 | |||||
| BHT | 63.24 ± 2.34 | |||||
| Trolox | 15 μM ± 0.62 | |||||
| SOD | 40 mU ± 0.85 | |||||
| Propyl gallate | 0.09 ± 0.04 | 0.09 ± 0.04 | ||||
| Sample | α-Amylase | α-Glucosidase | Lipase |
|---|---|---|---|
| B. drepanensis whole extract | 504.25 ± 5.11 * | 364.15 ± 3.14 * | 74.84 ± 3.67 * |
| Positive control | |||
| Acarbose | 35.52 ± 0.93 | 50.14 ± 1.31 | |
| Orlistat | 37.45 ± 1.08 |
| B. drepanensis Whole Extract | Total Phenol Content (TPC) mg GAE/g Extract | Total Flavonoid Content (TFC) mg CE/g Extract |
|---|---|---|
| 29.87 ± 0.14 * | 26.12 ± 0.09 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malfa, G.A.; De Leo, M.; Tundis, R.; Braca, A.; Loizzo, M.R.; Di Giacomo, C.; Raimondo, F.M.; Bucchini, A.E.A.; Acquaviva, R. Biological Investigation and Chemical Study of Brassica villosa subsp. drepanensis (Brassicaeae) Leaves. Molecules 2022, 27, 8447. https://doi.org/10.3390/molecules27238447
Malfa GA, De Leo M, Tundis R, Braca A, Loizzo MR, Di Giacomo C, Raimondo FM, Bucchini AEA, Acquaviva R. Biological Investigation and Chemical Study of Brassica villosa subsp. drepanensis (Brassicaeae) Leaves. Molecules. 2022; 27(23):8447. https://doi.org/10.3390/molecules27238447
Chicago/Turabian StyleMalfa, Giuseppe Antonio, Marinella De Leo, Rosa Tundis, Alessandra Braca, Monica Rosa Loizzo, Claudia Di Giacomo, Francesco Maria Raimondo, Anahi Elena Ada Bucchini, and Rosaria Acquaviva. 2022. "Biological Investigation and Chemical Study of Brassica villosa subsp. drepanensis (Brassicaeae) Leaves" Molecules 27, no. 23: 8447. https://doi.org/10.3390/molecules27238447
APA StyleMalfa, G. A., De Leo, M., Tundis, R., Braca, A., Loizzo, M. R., Di Giacomo, C., Raimondo, F. M., Bucchini, A. E. A., & Acquaviva, R. (2022). Biological Investigation and Chemical Study of Brassica villosa subsp. drepanensis (Brassicaeae) Leaves. Molecules, 27(23), 8447. https://doi.org/10.3390/molecules27238447