
Imperatorin Influences Depressive-like Behaviors: A Preclinical Study on Behavioral and Neurochemical Sex Differences


 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Spontaneous Locomotor Activity in Female Mice under the Influence of Imperatorin (1, 5, 10 mg/kg) and Imipramine (30 mg/kg)
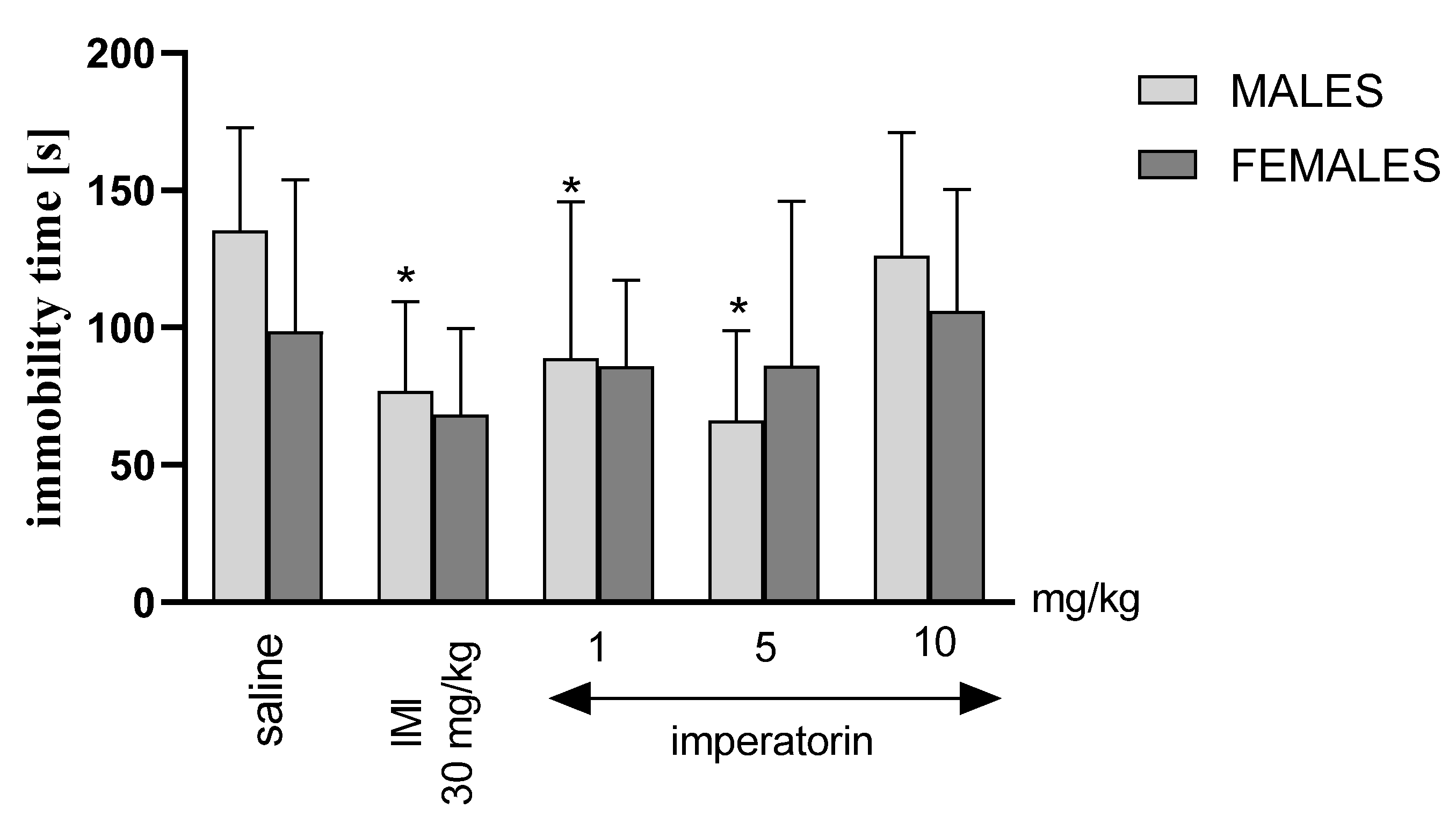
2.2. Imperatorin Decreases the Level of Depressive-like Behaviors in the Forced Swimming Test
2.3. Immobility Latency in the Forced Swim Test
2.4. Imperatorin Alters Serotonin Tissue Levels in the Hippocampus and Prefrontal Cortex
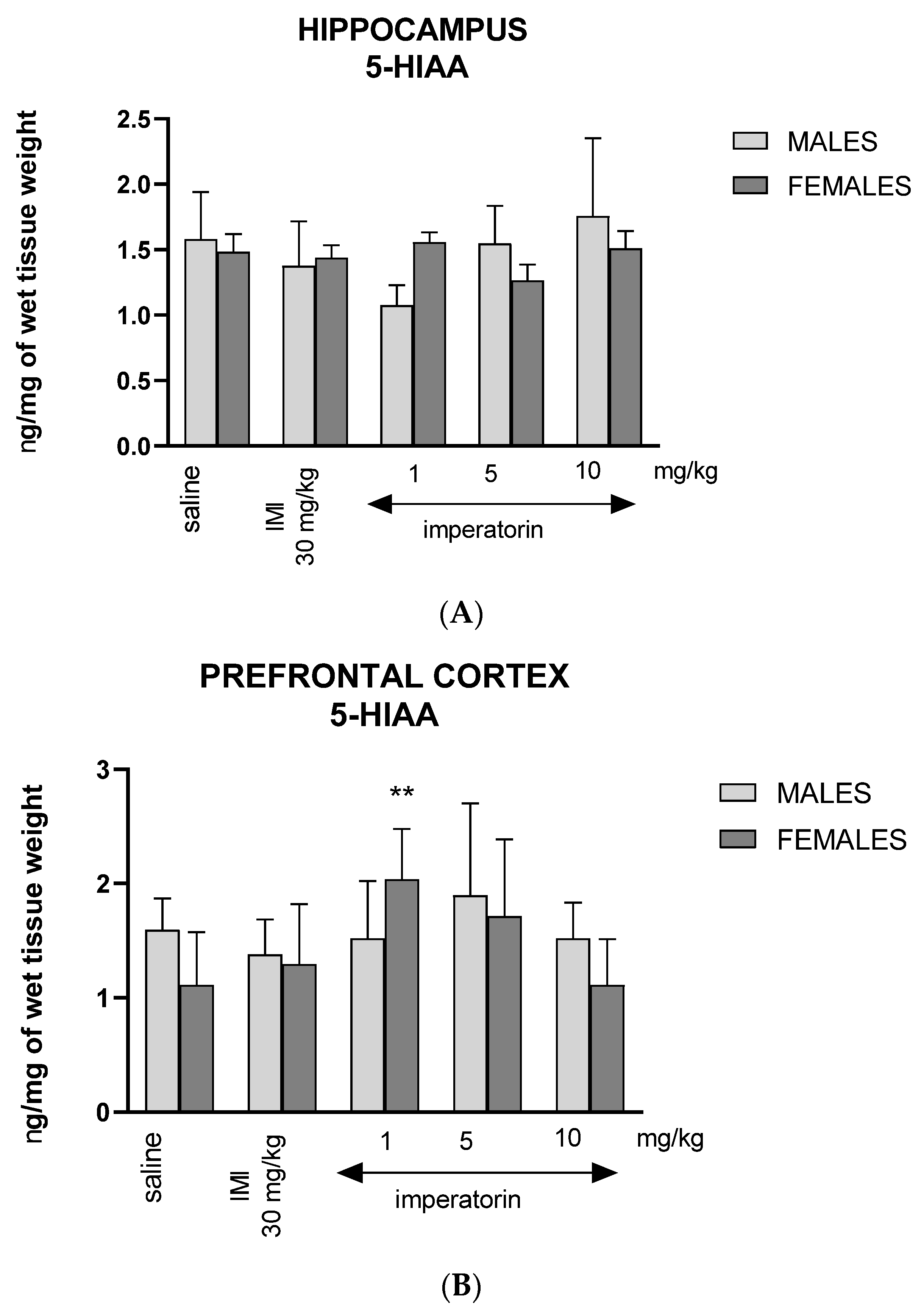
2.5. Imperatorin Alters 5-Hydroxyindoleacetic Acid Tissue Levels in the Hippocampus and Prefrontal Cortex
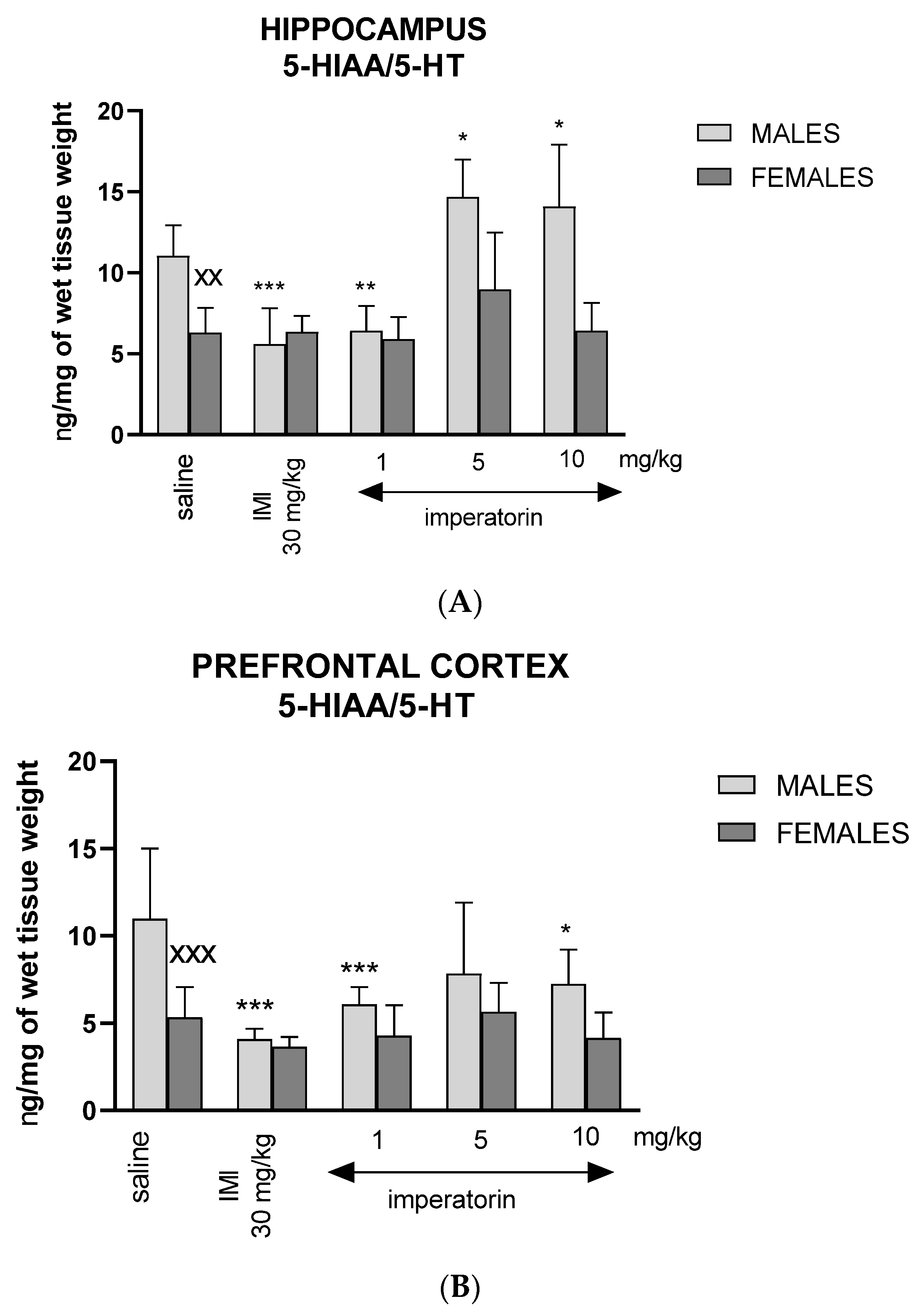
2.6. Imperatorin Modifies Serotonin Turnover Ratio
2.7. Imperatorin Influences Noradrenaline Tissue Levels
3. Discussion
4. Materials and Methods
4.1. Drugs
4.2. Animals
4.3. Spontaneous Locomotor Activity
4.4. Forced Swim Test (FST)
4.5. Neurochemical Assays
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, J.; Wu, X.; Lai, W.; Long, E.; Zhang, X.; Li, W.; Zhu, Y.; Chen, C.; Zhong, X.; Liu, Z.; et al. Prevalence of depression and depressive symptoms among outpatients: A systematic review and meta-analysis. BMJ Open 2017, 7, e017173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozioł, E.; Skalicka-Woźniak, K. Imperatorin-pharmacological meaning and analytical clues: Profound investigation. Phytochem. Rev. 2016, 15, 627–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tun, T.; Kang, Y.S. Imperatorin is Transported through Blood-Brain Barrier by Carrier-Mediated Transporters. Biomol. Ther. 2017, 25, 441–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budzynska, B.; Boguszewska-Czubara, A.; Kruk-Slomka, M.; Skalicka-Wozniak, K.; Michalak, A.; Musik, I.; Biala, G. Effects of imperatorin on scopolamine-induced cognitive impairment and oxidative stress in mice. Psychopharmacology 2015, 232, 931–942. [Google Scholar] [CrossRef]
- Budzynska, B.; Kruk-Slomka, M.; Skalicka-Wozniak, K.; Biala, G.; Glowniak, K. The effects of imperatorin on anxiety and memory-related behavior in male Swiss mice. Exp. Clin. Psychopharmacol. 2012, 20, 325–332. [Google Scholar] [CrossRef]
- Budzynska, B.; Boguszewska-Czubara, A.; Kruk-Slomka, M.; Skalicka-Wozniak, K.; Michalak, A.; Musik, I.; Biala, G.; Glowniak, K. Effects of imperatorin on nicotine-induced anxiety- and memory-related responses and oxidative stress in mice. Physiol. Behav. 2013, 122, 46–55. [Google Scholar] [CrossRef]
- Luszczki, J.J.; Glowniak, K.; Czuczwar, S.J. Imperatorin enhances the protective activity of conventional antiepileptic drugs against maximal electroshock-induced seizures in mice. Eur. J. Pharmacol. 2007, 574, 133–139. [Google Scholar] [CrossRef]
- Shaymaa, M.M.; Narayan, D.C.; Nesma, M.M.; Soad, A.L.B.; Babu, L.T.; Samir, A.R. Promising selective MAO-B inhibition by sesamin, a lignan from Zanthoxylum flavum stems. Saudi. Pharm. J. 2020, 28, 409–413. [Google Scholar] [CrossRef]
- Granica, S.; Kiss, A.K.; Jarończyk, M.; Maurin, J.K.; Mazurek, A.P.; Czarnocki, Z. Synthesis of imperatorin analogs and their evaluation as acetylcholinesterase and butyrylcholinesterase inhibitors. Arch. Pharm. 2013, 346, 775–782. [Google Scholar] [CrossRef]
- Huang, G.J.; Deng, J.S.; Liao, J.C.; Hou, W.C.; Wang, S.Y.; Sung, P.J.; Kuo, Y.H. Inducible nitric oxide synthase and cyclooxygenase-2 participate in anti-inflammatory activity of imperatorin from Glehnia littoralis. J. Agric. Food Chem. 2012, 60, 1673–1681. [Google Scholar] [CrossRef]
- Cao, Y.; Liu, J.; Wang, Q.; Liu, M.; Cheng, Y.; Zhang, X.; Lin, T.; Zhu, Z. Antidepressive-like effect of imperatorin from Angelica dahurica in prenatally stressed offspring rats through 5-hydroxytryptamine system. Neuroreport 2017, 28, 426–433. [Google Scholar] [CrossRef] [Green Version]
- Balta, G.; Dalla, C.; Kokras, N. Women’s Psychiatry. Adv. Exp. Med. Biol. 2019, 1192, 225–249. [Google Scholar] [CrossRef] [PubMed]
- Zarrouf, F.A.; Artz, S.; Griffith, J.; Sirbu, C.; Kommor, M. Testosterone and depression: Systematic review and meta-analysis. J. Psychiatr. Pract. 2009, 15, 289–305. [Google Scholar] [CrossRef]
- Solomon, M.B.; Herman, J.P. Sex differences in psychopathology: Of gonads, adrenals and mental illness. Physiol. Behav. 2009, 97, 250–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parker, G.B.; Brotchie, H.L. From diathesis to dimorphism: The biology of gender differences in depression. J. Nerv. Ment. Dis. 2004, 192, 210–216. [Google Scholar] [CrossRef] [PubMed]
- Ahokas, A.; Kaukoranta, J.; Wahlbeck, K.; Aito, M. Estrogen deficiency in severe postpartum depression: Successful treatment with sublingual physiologic 17beta-estradiol: A preliminary study. J. Clin. Psychiatry 2001, 62, 332–336. [Google Scholar] [CrossRef]
- Moreno, F.A.; McGahuey, C.A.; Freeman, M.P.; Delgado, P.L. Sex differences in depressive response during monoamine depletions in remitted depressive subjects. J. Clin. Psychiatry 2006, 67, 1618–1623. [Google Scholar] [CrossRef]
- Szewczyk, B.; Albert, P.R.; Burns, A.M.; Czesak, M.; Overholser, J.C.; Jurjus, G.J.; Meltzer, H.Y.; Konick, L.C.; Dieter, L.; Herbst, N.; et al. Gender-specific decrease in NUDR and 5-HT1A receptor proteins in the prefrontal cortex of subjects with major depressive disorder. Int. J. Neuropsychopharmacol. 2009, 12, 155–168. [Google Scholar] [CrossRef] [Green Version]
- Mann, J.J.; Brent, D.A.; Arango, V. The neurobiology and genetics of suicide and attempted suicide: A focus on the serotonergic system. Neuropsychopharmacology 2001, 24, 467–477. [Google Scholar] [CrossRef]
- Frey, B.N.; Skelin, I.; Sakai, Y.; Nishikawa, M.; Diksic, M. Gender differences in alpha-[(11)C]MTrp brain trapping, an index of serotonin synthesis, in medication-free individuals with major depressive disorder: A positron emission tomography study. Psychiatry Res. 2010, 183, 157–166. [Google Scholar] [CrossRef]
- Khan, A.; Brodhead, A.E.; Schwartz, K.A.; Kolts, R.L.; Brown, W.A. Sex differences in antidepressant response in recent antidepressant clinical trials. J. Clin. Psychopharmacol. 2005, 4, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Joyce, P.R.; Mulder, R.T.; Luty, S.E.; McKenzie, J.M.; Rae, A.M. A differential response to nortriptyline and fluoxetine in melancholic depression: The importance of age and gender. Acta Psychiatr. Scand. 2003, 108, 20–23. [Google Scholar] [CrossRef]
- Kornstein, S.G.; Schatzberg, A.F.; Thase, M.E.; Yonkers, K.A.; McCullough, J.P.; Keitner, G.I.; Gelenberg, A.J.; Davis, S.M.; Harrison, W.M.; Keller, M.B. Gender differences in treatment response to sertraline versus imipramine in chronic depression. Am. J. Psychiatry 2000, 157, 1445–1452. [Google Scholar] [CrossRef]
- Carrier, N.; Kabbaj, M. Testosterone and imipramine have antidepressant effects in socially isolated male but not female rats. Horm. Behav. 2012, 61, 678–685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, G.; Peng, C.; Du, W.; Wang, S. Simultaneous determination of imperatorin and its metabolites in vitro and in vivo by a GC-MS method: Application to a bioavailability and protein binding ability study in rat plasma. Biomed. Chromatogr. 2014, 28, 947–956. [Google Scholar] [CrossRef] [PubMed]
- He, W.; Chen, W.; Zhou, Y.; Tian, Y.; Liao, F. Xanthotoxol exerts neuroprotective effects via suppression of the inflammatory response in a rat model of focal cerebral ischemia. Cell. Mol. Neurobiol. 2013, 33, 715–722. [Google Scholar] [CrossRef]
- Liu, J.; Lian, Q.; Zhou, L.; Zhou, Q.; He, W.; Zhu, Z.; Lai, F. Calcium antagonistic effect of Xanthotoxol on isolated guinea pig atria. Zhong Yao Cai 2005, 28, 319–321. (In Chinese) [Google Scholar]
- Márquez, N.; Sancho, R.; Ballero, M.; Bremner, P.; Appendino, G.; Fiebich, B.L.; Heinrich, M.; Muñoz, E. Imperatorin inhibits T-cell proliferation by targeting the transcription factor NFAT. Planta Med. 2004, 70, 1016–1021. [Google Scholar] [CrossRef]
- Kowalczyk, J.; Nakos-Bimpos, M.; Polissidis, A.; Dalla, C.; Kokras, N.; Skalicka-Wozniak, K.; Budzynska, B. Xanthotoxin affects depression-related behavior and neurotransmitters content in a sex-dependent manner in mice. Behav. Brain Res. 2021, 399, 112985. [Google Scholar] [CrossRef]
- Kokras, N.; Antoniou, K.; Mikail, H.G.; Kafetzopoulos, V.; Papadopoulou-Daifoti, Z.; Dalla, C. Forced swim test: What about females? Neuropharmacology 2015, 99, 408–421. [Google Scholar] [CrossRef]
- Kokras, N.; Dalla, C.; Papadopoulou-Daifoti, Z. Sex differences in pharmacokinetics of antidepressants. Expert Opin. Drug Metab. Toxicol. 2011, 7, 213–226. [Google Scholar] [CrossRef]
- Mikail, H.G.; Dalla, C.; Kokras, N.; Kafetzopoulos, V.; Papadopoulou-Daifoti, Z. Sertraline behavioral response associates closer and dose-dependently with cortical rather than hippocampal serotonergic activity in the rat forced swim stress. Physiol. Behav. 2012, 107, 201–206. [Google Scholar] [CrossRef]
- Barton, D.A.; Esler, M.D.; Dawood, T.; Lambert, E.A.; Haikerwal, D.; Brenchley, C.; Socratous, F.; Hastings, J.; Guo, L.; Wiesner, G.; et al. Elevated Brain Serotonin Turnover in Patients With Depression: Effect of Genotype and Therapy. Arch. Gen. Psychiatry 2008, 65, 38–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leopoldo, M.; Lacivita, E.; Berardi, F.; Perrone, R.; Hedlund, P.B. Serotonin 5-HT7 receptor agents: Structure-activity relationships and potential therapeutic applications in central nervous system disorders. Pharmacol. Ther. 2011, 129, 120–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delgado, P.L. Depression: The case for a monoamine deficiency. J. Clin. Psychiatry 2000, 61 (Suppl. 6), 7–11. [Google Scholar] [PubMed]
- Matos, M.J.; Vilar, S.; Vazquez-Rodriguez, S.; Kachler, S.; Klotz, K.N.; Buccioni, M.; Delogu, G.; Santana, L.; Uriarte, E.; Borges, F. Structure-Based Optimization of Coumarin hA3 Adenosine Receptor Antagonists. J. Med. Chem. 2020, 63, 2577–2587. [Google Scholar] [CrossRef] [PubMed]
- Yu-Taeger, L.; Ott, T.; Bonsi, P.; Tomczak, C.; Wassouf, Z.; Martella, G.; Sciamanna, G.; Imbriani, P.; Ponterio, G.; Tassone, A.; et al. Impaired dopamine- and adenosine-mediated signaling and plasticity in a novel rodent model for DYT25 dystonia. Neurobiol. Dis. 2020, 134, 104634. [Google Scholar] [CrossRef] [PubMed]
- López-Cruz, L.; Salamone, J.D.; Correa, M. Caffeine and Selective Adenosine Receptor Antagonists as New Therapeutic Tools for the Motivational Symptoms of Depression. Front. Pharmacol. 2018, 9, 526. [Google Scholar] [CrossRef]
- Kokras, N.; Baltas, D.; Theocharis, F.; Dalla, C. Kinoscope: An Open-Source Computer Program for Behavioral Pharmacologists. Front. Behav. Neurosci. 2017, 11, 88. [Google Scholar] [CrossRef] [Green Version]
- Polissidis, A.; Koronaiou, M.; Kollia, V.; Koronaiou, E.; Nakos-Bimpos, M.; Bogiongko, M.; Vrettou, S.; Karali, K.; Casadei, N.; Riess, O.; et al. Psychosis-Like Behavior and Hyperdopaminergic Dysregulation in Human α-Synuclein BAC Transgenic Rats. Mov. Disord. 2021, 36, 716–728. [Google Scholar] [CrossRef]
- Kokras, N.; Pastromas, N.; Papasava, D.; de Bournonville, C.; Cornil, C.A.; Dalla, C. Sex differences in behavioral and neurochemical effects of gonadectomy and aromatase inhibition in rats. Psychoneuroendocrinology 2018, 87, 93–107. [Google Scholar] [CrossRef] [PubMed]
- Kokras, N.; Antoniou, K.; Dalla, C.; Bekris, S.; Xagoraris, M.; Ovestreet, D.H.; Papadopoulou-Daifoti, Z. Sex-related differential response to clomipramine treatment in a rat model of depression. J. Psychopharmacol. 2009, 8, 945–956. [Google Scholar] [CrossRef] [PubMed]
- Novais, A.; Ferreira, A.C.; Marques, F.; Pêgo, J.M.; Cerqueira, J.J.; David-Pereira, A.; Campos, F.L.; Dalla, C.; Kokras, N.; Sousa, N.; et al. Neudesin is involved in anxiety behavior: Structural and neurochemical correlates. Front. Behav. Neurosci. 2013, 7, 119. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MEANS ± SD Photocell Beam Breaks /30 min | |||||
|---|---|---|---|---|---|
| Saline | IMI 30 mg/kg | IMP 1 mg/kg | IMP 5 mg/kg | IMP 10 mg/kg | |
| Females | 7734 ± 3618 | 6623 ± 5156 | 10,242 ± 7766 | 7390 ± 3906 | 7960 ± 4662 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalczyk, J.; Nakos-Bimpos, M.; Polissidis, A.; Dalla, C.; Kokras, N.; Skalicka-Woźniak, K.; Budzyńska, B. Imperatorin Influences Depressive-like Behaviors: A Preclinical Study on Behavioral and Neurochemical Sex Differences. Molecules 2022, 27, 1179. https://doi.org/10.3390/molecules27041179
Kowalczyk J, Nakos-Bimpos M, Polissidis A, Dalla C, Kokras N, Skalicka-Woźniak K, Budzyńska B. Imperatorin Influences Depressive-like Behaviors: A Preclinical Study on Behavioral and Neurochemical Sex Differences. Molecules. 2022; 27(4):1179. https://doi.org/10.3390/molecules27041179
Chicago/Turabian StyleKowalczyk, Joanna, Modestos Nakos-Bimpos, Alexia Polissidis, Christina Dalla, Nikolaos Kokras, Krystyna Skalicka-Woźniak, and Barbara Budzyńska. 2022. "Imperatorin Influences Depressive-like Behaviors: A Preclinical Study on Behavioral and Neurochemical Sex Differences" Molecules 27, no. 4: 1179. https://doi.org/10.3390/molecules27041179
APA StyleKowalczyk, J., Nakos-Bimpos, M., Polissidis, A., Dalla, C., Kokras, N., Skalicka-Woźniak, K., & Budzyńska, B. (2022). Imperatorin Influences Depressive-like Behaviors: A Preclinical Study on Behavioral and Neurochemical Sex Differences. Molecules, 27(4), 1179. https://doi.org/10.3390/molecules27041179