
Bioactive Compounds from Pale Ale Beer Powder Attenuate Experimental Colitis in BALB/c Mice


 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results
2.1. PCs Identification and Quantification
2.2. Dextran Sodium Sulfate (DSS) Induction Leads to Clinical and Anatomic Signs of Inflammatory Activity
2.3. BPE and BPFPE Attenuate Microscopic Inflammation in DSS-Induced Acute Colitis
2.4. Pretreatment with BPE and BPFPE Decreases the Loss of Goblet Cells and Apoptotic Rates in the Colon of DSS-Induced Mice
2.5. Pretreatment with BPE and BPFPE Decreases the Accumulation of Immune Cells in the Colonic Lamina Propria of DSS-Induced Mice
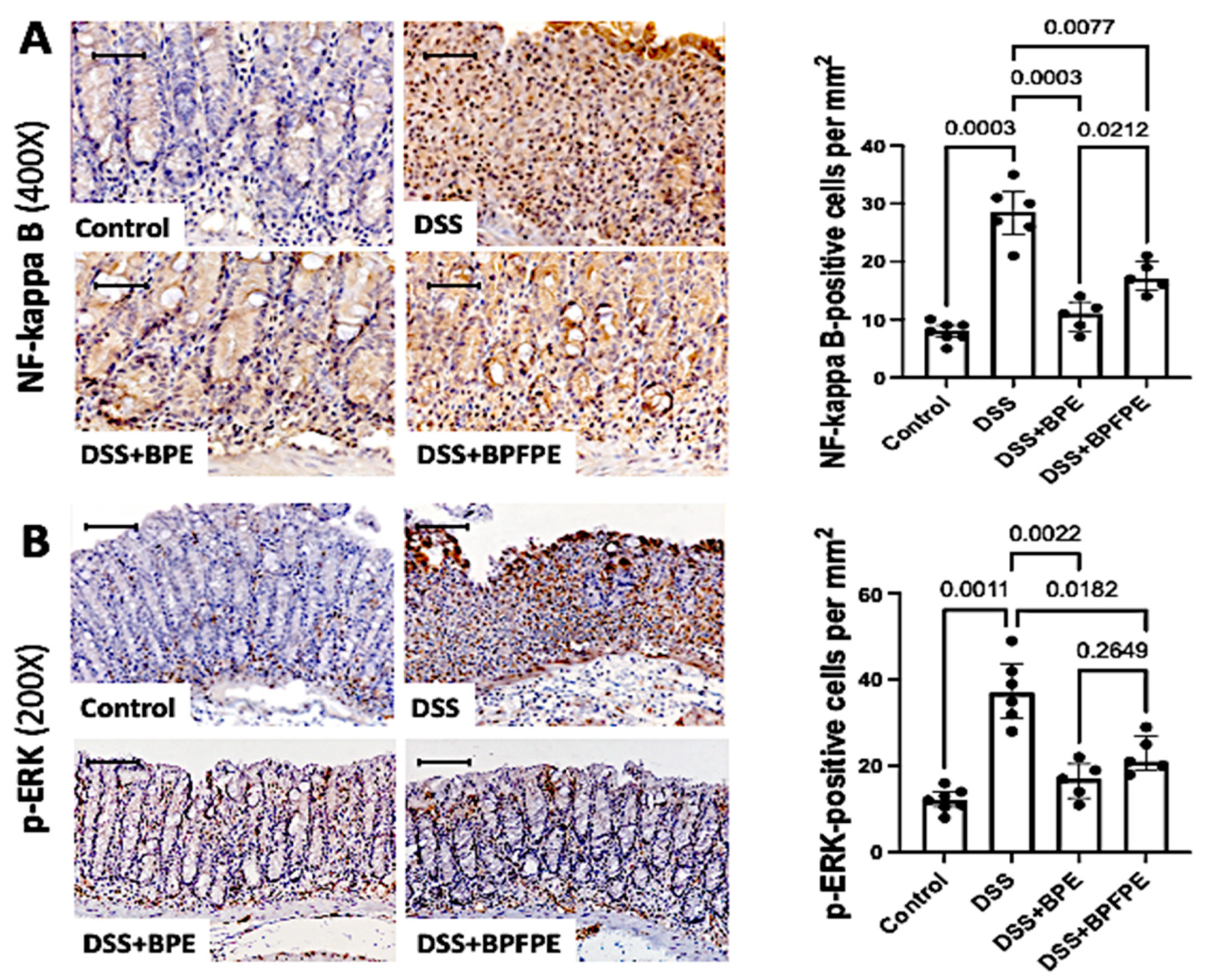
2.6. Pretreatment with BPE and BPFPE Blunts NF-κB and ERK Activation
3. Discussion
4. Methods
4.1. Powdered Pale Ale Craft Beer
4.2. Phenolic Extract Production
4.3. PCs Contents
4.4. In Vivo Bioactivity Assessment of Phenolic Extract and Powdered Pale Ale Craft Beer
Mice
4.5. Experimental Design
4.6. Histological Sample Preparation and Evaluation
4.7. Goblet Cell and Collagen Deposition Assessment in the Colon
4.8. Immunohistochemistry
4.9. Assessment of Apoptosis in the Colon
4.10. Quantitative Assessment of Colon Sections
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Ambra, R.; Pastore, G.; Lucchetti, S. The Role of Bioactive Phenolic Compounds on the Impact of Beer on Health. Molecules 2021, 26, 486. [Google Scholar] [CrossRef] [PubMed]
- Bamforth, C. Progress in Brewing Science and Beer Production. Annu. Rev. Chem. Biomol. Eng. 2017, 8, 161–176. [Google Scholar] [CrossRef]
- Rada, P.Q.; Vallverdu-Queralt, A.; Martínez-Huélamo, M.; Chiva-Blanch, G.; Jáuregui, O.; Estruch, R.; Lamuela-Raventós, R. A comprehensive characterisation of beer polyphenols by high resolution mass spectrometry (LC–ESI-LTQ-Orbitrap-MS). Food Chem. 2015, 169, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Karatzi, K.; Rontoyanni, V.G.; Protogerou, A.D.; Georgoulia, A.; Xenos, K.; Chrysou, J.; Sfikakis, P.P.; Sidossis, L.S. Acute effects of beer on endothelial function and hemodynamics: A single-blind, crossover study in healthy volunteers. Nutrients 2013, 29, 1122–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.; Je, Y. Flavonoid intake and mortality from cardiovascular disease and all causes: A meta-analysis of prospective cohort studies. Clin. Nutr. ESPEN 2017, 20, 68–77. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.E.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (Poly)phenolics in Human Health: Structures, Bioavailability, and Evidence of Protective Effects Against Chronic Diseases. Antioxid. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [Green Version]
- Ronksley, P.; Brien, E.S.; Turner, B.J.; Mukamal, K.J.; Ghali, A.W. Association of alcohol consumption with selected cardiovascular disease outcomes: A systematic review and meta-analysis. BMJ 2011, 342, d671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brien, E.S.; Ronksley, P.; Turner, B.J.; Mukamal, K.J.; Ghali, A.W. Effect of alcohol consumption on biological markers associated with risk of coronary heart disease: Systematic review and meta-analysis of interventional studies. BMJ 2011, 342, d636. [Google Scholar] [CrossRef] [Green Version]
- Mishima, Y.; Sartor, R.B. Manipulating resident microbiota to enhance regulatory immune function to treat inflammatory bowel diseases. J. Gastroenterol. 2020, 55, 4–14. [Google Scholar] [CrossRef] [Green Version]
- Cosnes, J.; Gower, C.; Seksik, P.; Cortot, A. Epidemiology and Natural History of Inflammatory Bowel Diseases. Gastroenterology 2011, 140, 1785–1794.e4. [Google Scholar] [CrossRef]
- Park, J.H.; Peyrin-Biroulet, L.; Eisenhut, M.; Shin, J.I. IBD immunopathogenesis: A comprehensive review of inflammatory molecules. Autoimmun. Rev. 2017, 16, 416–426. [Google Scholar] [CrossRef]
- Ananthakrishnan, A.N. Epidemiology and risk factors for IBD. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 205–217. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.C.; Tang, W.; Ching, J.Y.; Wong, M.; Chow, C.M.; Hui, A.J.; Wong, T.C.; Leung, V.K.; Tsang, S.W.; Yu, H.H.; et al. Incidence and Phenotype of Inflammatory Bowel Disease Based on Results from the Asia-Pacific Crohn’s and Colitis Epidemiology Study. Gastroenterology 2013, 145, 158–165.e2. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.C.; Shi, H.Y.; Hamidi, N.; Underwood, F.E.; Tang, W.; Benchimol, E.I.; Panaccione, R.; Ghosh, S.; Wu, J.C.Y.; Chan, F.K.L.; et al. Worldwide incidence and prevalence of inflammatory bowel disease in the 21st century: A systematic review of population-based studies. Lancet 2018, 390, 2769–2778. [Google Scholar] [CrossRef]
- Patel, K.R.; Brown, V.A.; Jones, D.J.; Britton, R.G.; Hemingway, D.; Miller, A.S.; West, K.P.; Booth, T.D.; Perloff, M.; Crowell, J.A.; et al. Clinical Pharmacology of Resveratrol and Its Metabolites in Colorectal Cancer Patients. Cancer Res. 2010, 70, 7392–7399. [Google Scholar] [CrossRef] [Green Version]
- Koosirirat, C.; Linpisarn, S.; Changsom, D.; Chawansuntati, K.; Wipasa, J. Investigation of the anti-inflammatory effect of Curcuma longa in Helicobacter pylori-infected patients. Int. Immunopharmacol. 2010, 10, 815–818. [Google Scholar] [CrossRef]
- Hoensch, H.; Groh, B.; Edler, L.; Kirch, W. Prospective cohort comparison of flavonoid treatment in patients with resected colorectal cancer to prevent recurrence. World J. Gastroenterol. 2008, 14, 2187–2193. [Google Scholar] [CrossRef]
- Graziani, G.; D’Argenio, G.; Tuccillo, C.; Loguercio, C.; Ritieni, A.; Morisco, F.; Blanco, C.D.V.; Fogliano, V.; Romano, M. Apple polyphenol extracts prevent damage to human gastric epithelial cells in vitro and to rat gastric mucosa in vivo. Gut 2005, 54, 193–200. [Google Scholar] [CrossRef]
- Romano, M.; Vitaglione, P.; Sellitto, S.; D’Argenio, G. Nutraceuticals for protection and healing of gastrointestinal mucosa. Curr. Med. Chem. 2012, 19, 109–117. [Google Scholar] [CrossRef]
- Arya, V.S.; Kanthlal, S.K.; Linda, G. The role of dietary polyphenols in inflammatory bowel disease: A possible clue on the molecular mechanisms involved in the prevention of immune and inflammatory reactions. J. Food Biochem. 2020, 44. [Google Scholar] [CrossRef]
- Biasi, F.; Astegiano, M.; Maina, M.; Leonarduzzi, G.M.; Poli, G. Polyphenol Supplementation as a Complementary Medicinal Approach to Treating Inflammatory Bowel Disease. Curr. Med. Chem. 2011, 18, 4851–4865. [Google Scholar] [CrossRef]
- Yang, Y.; Xiang, L.; He, J. Beverage intake and risk of Crohn disease. Medicine 2019, 98, e15795. [Google Scholar] [CrossRef] [PubMed]
- Zheng, T.; Wang, X.; Chen, Z.; He, A.; Zheng, Z.; Liu, G. Efficacy of adjuvant curcumin therapy in ulcerative colitis: A meta-analysis of randomized controlled trials. J. Gastroenterol. Hepatol. 2019, 35, 722–729. [Google Scholar] [CrossRef] [PubMed]
- Chandan, S. Curcumin use in ulcerative colitis: Is it ready for prime time? A systematic review and meta-analysis of clinical trials. Ann. Gastroenterol. 2019, 33, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Rosillo, M.A.; Hidalgo, M.S.; Cárdeno, A.; de la Lastra, C.A. Protective effect of ellagic acid, a natural polyphenolic compound, in a murine model of Crohn’s disease. Biochem. Pharmacol. 2011, 82, 737–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bora, A.F.M.; Ma, S.; Li, X.; Liu, L. Application of microencapsulation for the safe delivery of green tea polyphenols in food systems: Review and recent advances. Food Res. Int. 2018, 105, 241–249. [Google Scholar] [CrossRef] [PubMed]
- Sarabandi, K.; Jafari, S.M.; Mahoonak, A.S.; Mohammadi, A. Application of gum Arabic and maltodextrin for encapsulation of eggplant peel extract as a natural antioxidant and color source. Int. J. Biol. Macromol. 2019, 140, 59–68. [Google Scholar] [CrossRef]
- Zhao, H.; Chen, W.; Lu, J.; Zhao, M. Phenolic profiles and antioxidant activities of commercial beers. Food Chem. 2010, 119, 1150–1158. [Google Scholar] [CrossRef]
- Moura-Nunes, N.; Brito, T.C.; da Fonseca, N.D.; de Aguiar, P.F.; Monteiro, M.; Perrone, D.; Torres, A.G. Phenolic compounds of Brazilian beers from different types and styles and application of chemometrics for modeling antioxidant capacity. Food Chem. 2016, 199, 105–113. [Google Scholar] [CrossRef] [Green Version]
- Ellulu, M.S.; Patimah, I.; KhazáAi, H.; Rahmat, A.; Abed, Y. Obesity and inflammation: The linking mechanism and the complications. Arch. Med. Sci. 2017, 13, 851–863. [Google Scholar] [CrossRef]
- Acosta-Estrada, B.A.; Gutiérrez-Uribe, J.A.; Serna-Saldívar, S.O. Bound phenolics in foods, a review. Food Chem. 2014, 152, 46–55. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Zamora-Ros, R.; Chan, S.S.M.; Cross, A.J.; Ward, H.; Jakszyn, P.; Luben, R.; Opstelten, J.L.; Oldenburg, B.; Hallmans, G.; et al. Dietary Polyphenols in the Aetiology of Crohnʼs Disease and Ulcerative Colitis—A Multicenter European Prospective Cohort Study (EPIC). Inflamm. Bowel Dis. 2017, 23, 2072–2082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hagan, M.; Hayee, B.; Rodriguez-Mateos, A. (Poly)phenols in Inflammatory Bowel Disease and Irritable Bowel Syndrome: A Review. Molecules 2021, 26, 1843. [Google Scholar] [CrossRef]
- Goulart, R.D.A.; Barbalho, S.M.; Lima, V.M.; De Souza, G.A.; Matias, J.N.; Araújo, A.C.; Rubira, C.J.; Buchaim, R.L.; Buchaim, D.V.; De Carvalho, A.C.A.; et al. Effects of the Use of Curcumin on Ulcerative Colitis and Crohn’s Disease: A Systematic Review. J. Med. Food 2020, 24, 675–685. [Google Scholar] [CrossRef] [PubMed]
- Granado-Lorencio, F.; Hernández-Álvarez, E. Functional Foods and Health Effects: A Nutritional Biochemistry Perspective. Curr. Med. Chem. 2016, 23, 2929–2957. [Google Scholar] [CrossRef] [PubMed]
- Clifford, M.N. Diet-Derived Phenols in Plasma and Tissues and their Implications for Health. Planta Med. 2004, 70, 1103–1114. [Google Scholar] [CrossRef] [Green Version]
- El-Mekkawy, S.; Shahat, A.A.; Alqahtani, A.S.; Alsaid, M.S.; Abdelfattah, M.A.; Ullah, R.; Emam, M.; Yasri, A.; Sobeh, M. A Polyphenols-Rich Extract from Moricandia sinaica Boiss. Exhibits Analgesic, Anti-Inflammatory and Antipyretic Activities In Vivo. Molecules 2020, 25, 5049. [Google Scholar] [CrossRef]
- Mitsunari, K.; Miyata, Y.; Matsuo, T.; Mukae, Y.; Otsubo, A.; Harada, J.; Kondo, T.; Matsuda, T.; Ohba, K.; Sakai, H. Pharmacological Effects and Potential Clinical Usefulness of Polyphenols in Benign Prostatic Hyperplasia. Molecules 2021, 26, 450. [Google Scholar] [CrossRef]
- Zhuang, Y.; Wu, H.; Wang, X.; He, J.; He, S.; Yin, Y. Resveratrol Attenuates Oxidative Stress-Induced Intestinal Barrier Injury through PI3K/Akt-Mediated Nrf2 Signaling Pathway. Oxidative Med. Cell. Longev. 2019, 2019, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Guan, Y.; Tan, Y.; Liu, W.; Yang, J.; Wang, D.; Pan, D.; Sun, Y.; Zheng, C. NF-E2-Related Factor 2 Suppresses Intestinal Fibrosis by Inhibiting Reactive Oxygen Species-Dependent TGF-β1/SMADs Pathway. Am. J. Dig. Dis. 2017, 63, 366–380. [Google Scholar] [CrossRef]
- Sergent, T.; Piront, N.; Meurice, J.; Toussaint, O.; Schneider, Y.-J. Anti-inflammatory effects of dietary phenolic compounds in an in vitro model of inflamed human intestinal epithelium. Chem. Interact. 2010, 188, 659–667. [Google Scholar] [CrossRef] [PubMed]
- Boussenna, A.; Cholet, J.; Goncalves-Mendes, N.; Joubert-Zakeyh, J.; Fraisse, D.; Vasson, M.-P.; Texier, O.; Felgines, C. Polyphenol-rich grape pomace extracts protect against dextran sulfate sodium-induced colitis in rats. J. Sci. Food Agric. 2015, 96, 1260–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin, D.A.; Bolling, B.W. A review of the efficacy of dietary polyphenols in experimental models of inflammatory bowel diseases. Food Funct. 2015, 6, 1773–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahimi, R.; Abdollahi, M. The Role of Dietary Polyphenols in the Management of Inflammatory Bowel Disease. Curr. Pharm. Biotechnol. 2015, 16, 196–210. [Google Scholar] [CrossRef]
- Ambriz-Pérez, D.L.; Bang, W.Y.; Nair, V.; Angulo-Escalante, M.A.; Cisneros-Zevallos, L.; Heredia, J.B. Protective Role of Flavonoids and Lipophilic Compounds from Jatropha platyphylla on the Suppression of Lipopolysaccharide (LPS)-Induced Inflammation in Macrophage Cells. J. Agric. Food Chem. 2016, 64, 1899–1909. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, L.R.; Wang, J.; Le, T. Caffeic acid phenethyl ester, an inhibitor of nuclear factor-kappaB, attenuates bacterial peptidoglycan polysaccharide-induced colitis in rats. J. Pharmacol. Exp. Ther. 2001, 299, 915–920. [Google Scholar]
- Ye, Z.; Liu, Z.; Henderson, A.; Lee, K.-W.; Hostetter, J.; Wannemuehler, M.; Hendrich, S. Increased CYP4B1 mRNA Is Associated with the Inhibition of Dextran Sulfate Sodium–Induced Colitis by Caffeic Acid in Mice. Exp. Biol. Med. 2009, 234, 605–616. [Google Scholar] [CrossRef] [Green Version]
- Vallabhapurapu, S.; Karin, M. Regulation and Function of NF-κB Transcription Factors in the Immune System. Annu. Rev. Immunol. 2009, 27, 693–733. [Google Scholar] [CrossRef]
- Li, H.; Christman, L.M.; Li, R.; Gu, L. Synergic interactions between polyphenols and gut microbiota in mitigating inflammatory bowel diseases. Food Funct. 2020, 11, 4878–4891. [Google Scholar] [CrossRef]
- Kaulmann, A.; Bohn, T. Bioactivity of Polyphenols: Preventive and Adjuvant Strategies toward Reducing Inflammatory Bowel Diseases—Promises, Perspectives, and Pitfalls. Oxidative Med. Cell. Longev. 2016, 2016, 1–29. [Google Scholar] [CrossRef] [Green Version]
- Bengmark, S. Curcumin, An Atoxic Antioxidant and Natural NFκB, Cyclooxygenase-2, Lipooxygenase, and Inducible Nitric Oxide Synthase Inhibitor: A Shield Against Acute and Chronic Diseases. J. Parenter. Enter. Nutr. 2006, 30, 45–51. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, A.S.; Porawski, M.; Alonso, M.; Marroni, N.; Collado, P.S.; González-Gallego, J. Quercetin Decreases Oxidative Stress, NF-κB Activation, and iNOS Overexpression in Liver of Streptozotocin-Induced Diabetic Rats. J. Nutr. 2005, 135, 2299–2304. [Google Scholar] [CrossRef] [Green Version]
- Bilotta, S.; Paruchuru, L.; Feilhauer, K.; Köninger, J.; Lorentz, A. Resveratrol Is a Natural Inhibitor of Human Intestinal Mast Cell Activation and Phosphorylation of Mitochondrial ERK1/2 and STAT3. Int. J. Mol. Sci. 2021, 22, 7640. [Google Scholar] [CrossRef]
- Lee, H.C.; Jenner, A.M.; Low, C.S.; Lee, Y.K. Effect of tea phenolics and their aromatic fecal bacterial metabolites on intestinal microbiota. Res. Microbiol. 2006, 157, 876–884. [Google Scholar] [CrossRef]
- Smith, A.; Mackie, R.I. Effect of Condensed Tannins on Bacterial Diversity and Metabolic Activity in the Rat Gastrointestinal Tract. Appl. Environ. Microbiol. 2004, 70, 1104–1115. [Google Scholar] [CrossRef] [Green Version]
- Dolara, P.; Luceri, C.; De Filippo, C.; Femia, A.P.; Giovannelli, L.; Caderni, G.; Cecchini, C.; Silvi, S.; Orpianesi, C.; Cresci, A. Red wine polyphenols influence carcinogenesis, intestinal microflora, oxidative damage and gene expression profiles of colonic mucosa in F344 rats. Mutat. Res. Mol. Mech. Mutagen. 2005, 591, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Domínguez-Avila, J.A.; Villa-Rodriguez, J.A.; Montiel-Herrera, M.; Pacheco-Ordaz, R.; Roopchand, D.E.; Venema, K.; González-Aguilar, G.A. Phenolic Compounds Promote Diversity of Gut Microbiota and Maintain Colonic Health. Am. J. Dig. Dis. 2021, 66, 3270–3289. [Google Scholar] [CrossRef]
- Maia, P.D.D.S.; dos Santos Baião, D.; da Silva, V.P.F.; Lemos Miguel, M.A.; Quirino Lacerda, E.C.; de Araújo Calado, V.M.; da Silva Carneiro, C.; Finotelli, P.V.; Pierucci, A.P.T.R. Microencapsulation of a craft beer, nutritional composition, antioxidant stability, and drink acceptance. LWT 2020, 133, 110104. [Google Scholar] [CrossRef]
- Maia, P.D.D.S.; Baião, D.D.S.; Da Silva, V.P.F.; Calado, V.M.D.A.; Queiroz, C.; Pedrosa, C.; Valente-Mesquita, V.L.; Pierucci, A.P.T.R. Highly Stable Microparticles of Cashew Apple (Anacardium occidentale L.) Juice with Maltodextrin and Chemically Modified Starch. Food Bioprocess Technol. 2019, 12, 2107–2119. [Google Scholar] [CrossRef]
- Inada, K.O.P.; Oliveira, A.A.; Revorêdo, T.B.; Martins, A.B.N.; Lacerda, E.C.Q.; Freire, A.S.; Braz, B.F.; Santelli, R.; Torres, A.G.; Perrone, D.; et al. Screening of the chemical composition and occurring antioxidants in jabuticaba (Myrciaria jaboticaba) and jussara (Euterpe edulis) fruits and their fractions. J. Funct. Foods 2015, 17, 422–433. [Google Scholar] [CrossRef] [Green Version]
- Baião, D.D.S.; De Freitas, C.S.; Gomes, L.P.; Da Silva, D.; Correa, A.C.N.T.F.; Pereira, P.R.; Del Aguila, E.M.; Paschoalin, V.M.F. Polyphenols from Root, Tubercles and Grains Cropped in Brazil: Chemical and Nutritional Characterization and Their Effects on Human Health and Diseases. Nutrients 2017, 9, 1044. [Google Scholar] [CrossRef] [PubMed]
- Laukens, D.; Brinkman, B.M.; Raes, J.; De Vos, M.; Vandenabeele, P. Heterogeneity of the gut microbiome in mice: Guidelines for optimizing experimental design. FEMS Microbiol. Rev. 2016, 40, 117–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boussenna, A.; Gonçalves-Mendes, N.; Joubert-Zakeyh, J.; Pereira, B.; Fraisse, D.; Vasson, M.-P.; Texier, O.; Felgines, C. Impact of basal diet on dextran sodium sulphate (DSS)-induced colitis in rats. Eur. J. Nutr. 2014, 54, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Jin, B.-R.; Chung, K.-S.; Hwang, S.; Hwang, S.N.; Rhee, K.-J.; Lee, M.; An, H.-J. Rosmarinic acid represses colitis-associated colon cancer: A pivotal involvement of the TLR4-mediated NF-κB-STAT3 axis. Neoplasia 2021, 23, 561–573. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BPFPE (mg·g−1 fwb) | BPE (mg·g−1 fwb) | |
|---|---|---|
| Soluble Phenolic Compounds | ||
| Chlorogenic acid | 16.04 ± 0.08 a,* | 12.28 ± 1.08 a |
| Di-OH-benzoic acid | 14.81 ± 1.12 b | 11.39 ± 2.82 a,b |
| Ferulic acid | 10.52 ± 0.20 c | 8.09 ± 2.20 b |
| Gallic acid | 14.41 ± 0.01 b,* | 11.08 ± 1.07 a,b |
| m-Coumaric | 12.11 ± 2.09 c,* | 9.32 ± 1.59 b |
| p-Coumaric | 6.67 ± 1.37 e | 5.12 ± 2.37 c |
| Rosmarinic acid | 8.59 ± 0.68 d,e,* | 6.61 ± 0.58 c |
| Syringic acid | 9.58 ± 1.12 c,d | 7.37 ± 2.12 b,c |
| Vanillic acid | 15.37 ± 1.05 a,b | 11.82 ± 1.05 a |
| 4-OH-phenylacetic acid | 11.97 ± 1.69 b,c | 9.21 ± 1.89 a,b |
| Insoluble phenolic compounds | ||
| Gallic acid | 9.87 ± 0.21 a,* | 7.59 ± 0.11 a |
| Rosmarinic acid | 9.67 ± 1.01 a | 7.44 ± 1.51 a |
| Vanillic acid | 10.82 ± 1.19 a | 8.32 ± 1.39 a |
| 4-OH-phenylacetic acid | 9.78 ± 0.99 a,* | 7.52 ± 0.29 a |
| Total | 160.21 ± 2.17 * | 123.16 ± 2.17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maia, P.D.D.S.; Baião, D.d.S.; Nanini, H.F.; da Silva, V.P.F.; Frambach, L.B.; Cabral, I.M.; Pêgo, B.; Ribeiro, B.E.; Pavão, M.S.G.; Paschoalin, V.M.F.; et al. Bioactive Compounds from Pale Ale Beer Powder Attenuate Experimental Colitis in BALB/c Mice. Molecules 2022, 27, 1194. https://doi.org/10.3390/molecules27041194
Maia PDDS, Baião DdS, Nanini HF, da Silva VPF, Frambach LB, Cabral IM, Pêgo B, Ribeiro BE, Pavão MSG, Paschoalin VMF, et al. Bioactive Compounds from Pale Ale Beer Powder Attenuate Experimental Colitis in BALB/c Mice. Molecules. 2022; 27(4):1194. https://doi.org/10.3390/molecules27041194
Chicago/Turabian StyleMaia, Paola D. D. S., Diego dos Santos Baião, Hayandra F. Nanini, Victor Paulo F. da Silva, Lissa Bantim Frambach, Iuri Matheus Cabral, Beatriz Pêgo, Beatriz E. Ribeiro, Mauro Sérgio Gonçalves Pavão, Vania M. F. Paschoalin, and et al. 2022. "Bioactive Compounds from Pale Ale Beer Powder Attenuate Experimental Colitis in BALB/c Mice" Molecules 27, no. 4: 1194. https://doi.org/10.3390/molecules27041194
APA StyleMaia, P. D. D. S., Baião, D. d. S., Nanini, H. F., da Silva, V. P. F., Frambach, L. B., Cabral, I. M., Pêgo, B., Ribeiro, B. E., Pavão, M. S. G., Paschoalin, V. M. F., de Souza, H. S. P., & Pierucci, A. P. T. R. (2022). Bioactive Compounds from Pale Ale Beer Powder Attenuate Experimental Colitis in BALB/c Mice. Molecules, 27(4), 1194. https://doi.org/10.3390/molecules27041194