
Assessment of Murine Colon Inflammation Using Intraluminal Fluorescence Lifetime Imaging

 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Autofluorescence Properties Differ along Distinct Anatomical Regions within The Colon
2.2. The Epithelial Layer Influenced Tissue Autofluorescence Properties
2.3. Colon Fluorescence Lifetime Was Sensitive to Tissue Inflammation
3. Discussion
4. Materials and Methods
4.1. Animal Models
4.2. DSS Treatment
4.3. 5-ASA Treatment
4.4. Colonocyte Isolation
4.5. Lactate Measurements
4.6. ATP Measurements
4.7. Pdh Activity Measurements
4.8. Histology Images
4.9. Time-Resolved Fluorescence Spectrometer
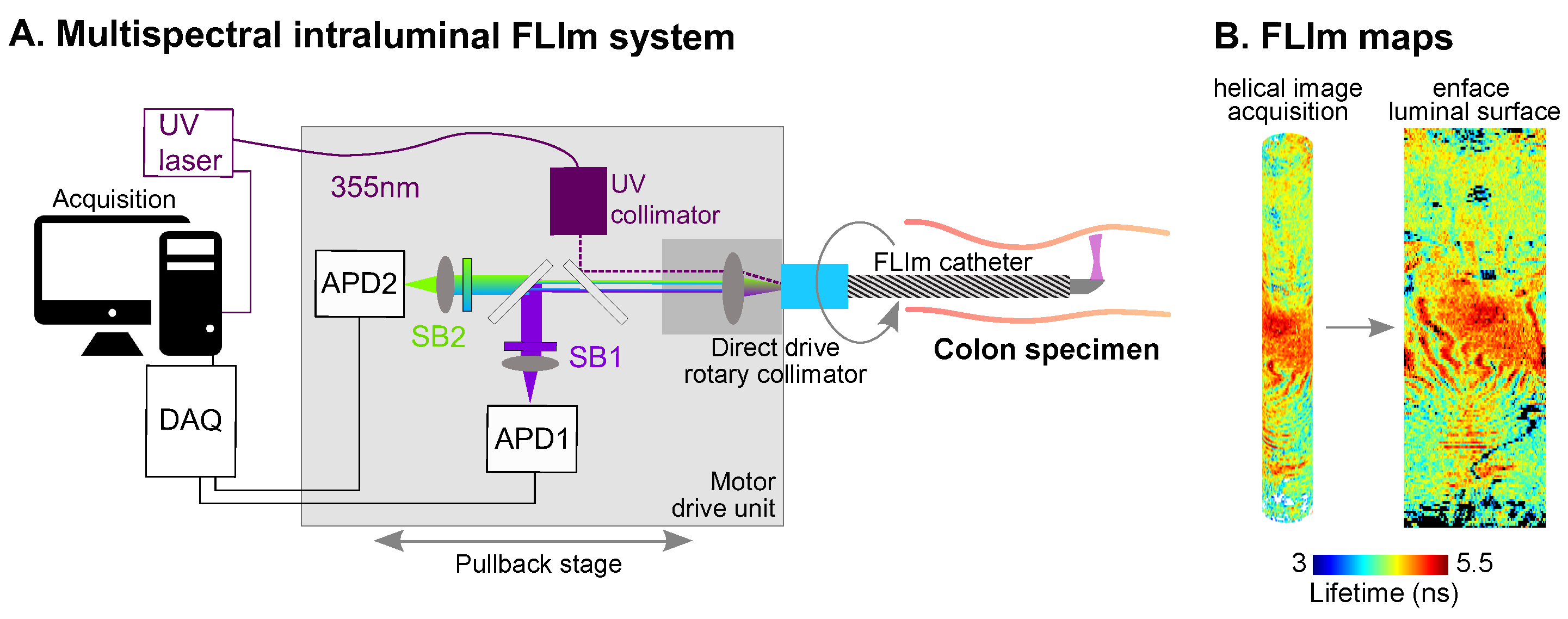
4.10. Fluorescence Lifetime Imaging (FLIm) Instruments
4.11. Spectral Measurements and Imaging of Colon Tissue
4.12. Image Analysis and Statistics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| FLIm | Fluorescence lifetime imaging |
| IBD | Inflammatory bowel disease |
| DSS | Dextran sodium sulfate |
| 5-ASA | 5-aminosalicylic acid |
| NADH | Nicotinamide adenine dinucleotide |
| FAD | Flavin adenine dinucleotide |
References
- Kaplan, G.G.; Windsor, J.W. The four epidemiological stages in the global evolution of inflammatory bowel disease. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 56–66. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, G.G. The global burden of IBD: From 2015 to 2025. Nat. Rev. Gastroenterol. Hepatol. 2015, 12, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Park, K.T.; Ehrlich, O.G.; Allen, J.I.; Meadows, P.; Szigethy, E.M.; Henrichsen, K.; Kim, S.C.; Lawton, R.C.; Murphy, S.M.; Regueiro, M.; et al. The Cost of Inflammatory Bowel Disease: An Initiative From the Crohn’s & Colitis Foundation. Inflamm. Bowel Dis. 2020, 26, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Mosli, M.; Beshir, M.A.; Al-Judaibi, B.; Al-Ameel, T.; Saleem, A.; Bessissow, T.; Ghosh, S.; Almadi, M. Advances in the Diagnosis and Management of Inflammatory Bowel Disease: Challenges and Uncertainties. Saudi J. Gastroenterol. Off. J. Saudi Gastroenterol. Assoc. 2014, 20, 81–101. [Google Scholar] [CrossRef]
- Parray, F.Q.; Wani, M.L.; Malik, A.A.; Wani, S.N.; Bijli, A.H.; Irshad, I.; Nayeem-UI-Hassan. Ulcerative Colitis: A Challenge to Surgeons. Int. J. Prev. Med. 2012, 3, 749. [Google Scholar] [PubMed]
- Kuipers, E.J.; Grady, W.M.; Lieberman, D.; Seufferlein, T.; Sung, J.J.; Boelens, P.G.; Velde, C.J.H.v.d.; Watanabe, T. Colorectal cancer. Nat. Rev. Dis. Prim. 2015, 1, 15065. [Google Scholar] [CrossRef] [Green Version]
- Terzić, J.; Grivennikov, S.; Karin, E.; Karin, M. Inflammation and Colon Cancer. Gastroenterology 2010, 138, 2101–2114.e5. [Google Scholar] [CrossRef]
- Dhyani, M.; Joshi, N.; Bemelman, W.A.; Gee, M.S.; Yajnik, V.; D’Hoore, A.; Traverso, G.; Donowitz, M.; Mostoslavsky, G.; Lu, T.K.; et al. Challenges in IBD Research: Novel Technologies. Inflamm. Bowel Dis. 2019, 25, S24–S30. [Google Scholar] [CrossRef]
- Kiesslich, R.; Neurath, M.F. Advanced endoscopy imaging in inflammatory bowel diseases. Gastrointest. Endosc. 2017, 85, 496–508. [Google Scholar] [CrossRef]
- Iacucci, M.; Furfaro, F.; Matsumoto, T.; Uraoka, T.; Smith, S.; Ghosh, S.; Kiesslich, R. Advanced endoscopic techniques in the assessment of inflammatory bowel disease: New technology, new era. Gut 2019, 68, 562. [Google Scholar] [CrossRef]
- Tontini, G.E.; Vecchi, M.; Pastorelli, L.; Neurath, M.F.; Neumann, H. Differential diagnosis in inflammatory bowel disease colitis: State of the art and future perspectives. World J. Gastroenterol. 2015, 21, 21–46. [Google Scholar] [CrossRef] [PubMed]
- Coda, S.; Siersema, P.D.; Stamp, G.W.H.; Thillainayagam, A.V. Biophotonic endoscopy: A review of clinical research techniques for optical imaging and sensing of early gastrointestinal cancer. Endosc. Int. Open 2015, 03, E380–E392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tjalma, J.J.J.; Koller, M.; Linssen, M.D.; Hartmans, E.; Jongh, S.D.; Jorritsma-Smit, A.; Karrenbeld, A.; Vries, E.G.D.; Kleibeuker, J.H.; Pennings, J.P.; et al. Quantitative fluorescence endoscopy: An innovative endoscopy approach to evaluate neoadjuvant treatment response in locally advanced rectal cancer. Gut 2019, 69, 406–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jongh, S.J.D.; Vrouwe, J.P.; Voskuil, F.J.; Schmidt, I.; Westerhof, J.; Koornstra, J.J.; Kam, M.L.D.; Karrenbeld, A.; Hardwick, J.C.; Robinson, D.J.; et al. The Optimal Imaging Window for Dysplastic Colorectal Polyp Detection Using c-Met–Targeted Fluorescence Molecular Endoscopy. J. Nucl. Med. 2020, 61, 1435–1441. [Google Scholar] [CrossRef]
- Bae, S.M.; Bae, D.J.; Do, E.J.; Oh, G.; Yoo, S.W.; Lee, G.J.; FChae, J.S.; Yun, Y.; Kim, S.; Kim, K.H.; et al. Multi-Spectral Fluorescence Imaging of Colon Dysplasia In Vivo Using a Multi-Spectral Endoscopy System. Transl. Oncol. 2019, 12, 226–235. [Google Scholar] [CrossRef]
- Maaser, C.; Sturm, A.; Vavricka, S.R.; Kucharzik, T.; Fiorino, G.; Annese, V.; Calabrese, E.; Baumgart, D.C.; Bettenworth, D.; Nunes, P.B.; et al. ECCO-ESGAR Guideline for Diagnostic Assessment in IBD Part 1: Initial diagnosis, monitoring of known IBD, detection of complications. J. Crohn’s Colitis 2018, 13, 144–164. [Google Scholar] [CrossRef] [Green Version]
- Zeng, M.Y.; Inohara, N.; Nuñez, G. Mechanisms of inflammation-driven bacterial dysbiosis in the gut. Mucosal Immunol. 2017, 10, 18. [Google Scholar] [CrossRef] [Green Version]
- Tiffany, C.R.; Baumler, A.J. Dysbiosis: From fiction to function. Am. J. Physiol. Gastrointest. Liver Physiol. 2019, 317, G602–G608. [Google Scholar] [CrossRef]
- Litvak, Y.; Byndloss, M.X.; Bäumler, A.J. Colonocyte metabolism shapes the gut microbiota. Science 2018, 362, eaat9076. [Google Scholar] [CrossRef] [Green Version]
- Georgakoudi, I.; Quinn, K.P. Optical Imaging Using Endogenous Contrast to Assess Metabolic State. Annu. Rev. Biomed. Eng. 2012, 14, 351–367. [Google Scholar] [CrossRef]
- Stringari, C.; Edwards, R.A.; Pate, K.T.; Waterman, M.L.; Donovan, P.J.; Gratton, E. Metabolic trajectory of cellular differentiation in small intestine by Phasor Fluorescence Lifetime Microscopy of NADH. Sci. Rep. 2012, 2, srep00568. [Google Scholar] [CrossRef] [Green Version]
- Alfonso-García, A.; Smith, T.D.; Datta, R.; Luu, T.U.; Gratton, E.; Potma, E.O.; Liu, W.F. Label-free identification of macrophage phenotype by fluorescence lifetime imaging microscopy. J. Biomed. Opt. 2016, 21, 46005. [Google Scholar] [CrossRef] [PubMed]
- Friedman, J.E.; Dobrinskikh, E.; Alfonso-Garcia, A.; Fast, A.; Janssen, R.C.; Soderborg, T.K.; Anderson, A.L.; Reisz, J.A.; D’Alessandro, A.; Frank, D.N.; et al. Pyrroloquinoline quinone prevents developmental programming of microbial dysbiosis and macrophage polarization to attenuate liver fibrosis in offspring of obese mice. Hepatol. Commun. 2018, 2, 313–328. [Google Scholar] [CrossRef] [Green Version]
- Hedde, P.N.; Cinco, R.; Malacrida, L.; Kamaid, A.; Gratton, E. Phasor-based hyperspectral snapshot microscopy allows fast imaging of live, three-dimensional tissues for biomedical applications. Commun. Biol. 2021, 4, 721. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, B.; Miedema, B.; Chandrasekhar, H.R. Emission Spectra of Colonic Tissue and Endogenous Fluorophores. Am. J. Med. Sci. 1998, 316, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Deal, J.; Mayes, S.; Browning, C.; Hill, S.; Rider, P.; Boudreaux, C.; Rich, T.C.; Leavesley, S.J. Identifying molecular contributors to autofluorescence of neoplastic and normal colon sections using excitation-scanning hyperspectral imaging. J. Biomed. Opt. 2019, 24, 21207. [Google Scholar] [CrossRef] [PubMed]
- Coda, S.; Thompson, A.J.; Kennedy, G.T.; Roche, K.L.; Ayaru, L.; Bansi, D.S.; Stamp, G.W.; Thillainayagam, A.V.; French, P.M.W.; Dunsby, C. Fluorescence lifetime spectroscopy of tissue autofluorescence in normal and diseased colon measured ex vivo using a fiber-optic probe. Biomed. Opt. Express 2014, 5, 515–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coda, S.; Thillainayagam, A. State of the art in advanced endoscopic imaging for the detection and evaluation of dysplasia and early cancer of the gastrointestinal tract. Clin. Exp. Gastroenterol. 2014, 7, 133–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munro, I.; McGinty, J.; Galletly, N.; Requejo-Isidro, J.; Lanigan, P.M.P.; Elson, D.S.; Dunsby, C.; Neil, M.A.A.; Lever, M.J.; Stamp, G.W.H.; et al. Toward the clinical application of time-domain fluorescence lifetime imaging. J. Biomed. Opt. 2005, 10, 051403. [Google Scholar] [CrossRef] [Green Version]
- Mycek, M.A.; Schomacker, K.T.; Nishioka, N.S. Colonic polyp differentiation using time-resolved autofluorescence spectroscopy. Gastrointest. Endosc. 1998, 48, 390–394. [Google Scholar] [CrossRef]
- McGinty, J.; Galletly, N.P.; Dunsby, C.; Munro, I.; Elson, D.S.; Requejo-Isidro, J.; Cohen, P.; Ahmad, R.; Forsyth, A.; Thillainayagam, A.V.; et al. Wide-field fluorescence lifetime imaging of cancer. Biomed. Opt. Express 2010, 1, 627–640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bec, J.; Li, C.; Marcu, L. Broadband, freeform focusing micro-opticsfor a side-viewing imaging catheter. Opt. Lett. 2019, 44, 4961–4964. [Google Scholar] [CrossRef] [PubMed]
- Abdelrahman, M.A.; Marston, G.; Hull, M.A.; Markham, A.F.; Jones, P.F.; Evans, J.A.; Coletta, P.L. High-Frequency Ultrasound for In Vivo Measurement of Colon Wall Thickness in Mice. Ultrasound Med. Biol. 2012, 38, 432–442. [Google Scholar] [CrossRef] [PubMed]
- Chassaing, B.; Aitken, J.D.; Malleshappa, M.; Vijay-Kumar, M. Dextran Sulfate Sodium (DSS)-Induced Colitis in Mice. Curr. Protoc. Immunol. 2014, 104, 15–25. [Google Scholar] [CrossRef]
- Lakowicz, J.R. Principles of Fluorescence Spectroscopy, 3rd ed.; Springer: New York, NY, USA, 2006. [Google Scholar] [CrossRef]
- Berezin, M.Y.; Achilefu, S. Fluorescence Lifetime Measurements and Biological Imaging. Chem. Rev. 2010, 110, 2641–2684. [Google Scholar] [CrossRef] [Green Version]
- Eichele, D.D.; Kharbanda, K.K. Dextran sodium sulfate colitis murine model: An indispensable tool for advancing our understanding of inflammatory bowel diseases pathogenesis. World J. Gastroenterol. 2017, 23, 6016–6029. [Google Scholar] [CrossRef]
- Cevallos, S.A.; Lee, J.Y.; Tiffany, C.R.; Byndloss, A.J.; Johnston, L.; Byndloss, M.X.; Bäumler, A.J. Increased Epithelial Oxygenation Links Colitis to an Expansion of Tumorigenic Bacteria. mBio 2019, 10, e02244-19. [Google Scholar] [CrossRef] [Green Version]
- Cevallos, S.A.; Lee, J.Y.; Velazquez, E.M.; Foegeding, N.J.; Shelton, C.D.; Tiffany, C.R.; Parry, B.H.; Stull-Lane, A.R.; Olsan, E.E.; Savage, H.P.; et al. 5-Aminosalicylic Acid Ameliorates Colitis and Checks Dysbiotic Escherichia coli Expansion by Activating PPAR-γ Signaling in the Intestinal Epithelium. mBio 2021, 12, e03227-20. [Google Scholar] [CrossRef]
- Ramírez-Alcántara, V.; Montrose, M.H. Acute murine colitis reduces colonic 5-aminosalicylic acid metabolism by regulation of N-acetyltransferase-2. Am. J. Physiol.-Gastrointest. Liver Physiol. 2014, 306, G1002–G1010. [Google Scholar] [CrossRef] [Green Version]
- Matthis, A.L.; Zhang, B.; Denson, L.A.; Yacyshyn, B.R.; Aihara, E.; Montrose, M.H. Importance of the Evaluation of N-Acetyltransferase Enzyme Activity Prior to 5-Aminosalicylic Acid Medication for Ulcerative Colitis. Inflamm. Bowel Dis. 2016, 22, 1793–1802. [Google Scholar] [CrossRef] [Green Version]
- Zhdanov, A.V.; Okkelman, I.A.; Golubeva, A.V.; Doerr, B.; Hyland, N.P.; Melgar, S.; Shanahan, F.; Cryan, J.F.; Papkovsky, D.B. Quantitative analysis of mucosal oxygenation using ex vivo imaging of healthy and inflamed mammalian colon tissue. Cell. Mol. Life Sci. 2017, 74, 141–151. [Google Scholar] [CrossRef] [PubMed]
- Brückner, M.; Lenz, P.; Mücke, M.M.; Gohar, F.; Willeke, P.; Domagk, D.; Bettenworth, D. Diagnostic imaging advances in murine models of colitis. World J. Gastroenterol. 2016, 22, 996–1007. [Google Scholar] [CrossRef] [PubMed]
- Yankelevich, D.R.; Ma, D.; Liu, J.; Sun, Y.; Sun, Y.; Bec, J.; Elson, D.S.; Marcu, L. Design and evaluation of a device for fast multispectral time-resolved fluorescence spectroscopy and imaging. Rev. Sci. Instrum. 2014, 85, 34303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfonso-Garcia, A.; Shklover, J.; Sherlock, B.E.; Panitch, A.; Griffiths, L.G.; Marcu, L. Fiber-based fluorescence lifetime imaging of recellularization processes on vascular tissue constructs. J. Biophotonics 2018, 11, e201700391. [Google Scholar] [CrossRef]
- Zhou, X.; Bec, J.; Yankelevich, D.; Marcu, L. Multispectral fluorescence lifetime imaging device with a silicon avalanche photodetector. Opt. Express 2021, 29, 20105. [Google Scholar] [CrossRef]
- American National Standards Institute. American National Standard for Safe Use of Lasers in Health Care ANSI Z136.1; Laser Institute of America: Orlando, FL, USA, 2007. [Google Scholar]
- Liu, J.; Sun, Y.; Qi, J.; Marcu, L. A novel method for fast and robust estimation of fluorescence decay dynamics using constrained least-squares deconvolution with Laguerre expansion. Phys. Med. Biol. 2012, 57, 843–865. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alfonso-Garcia, A.; Cevallos, S.A.; Lee, J.-Y.; Li, C.; Bec, J.; Bäumler, A.J.; Marcu, L. Assessment of Murine Colon Inflammation Using Intraluminal Fluorescence Lifetime Imaging. Molecules 2022, 27, 1317. https://doi.org/10.3390/molecules27041317
Alfonso-Garcia A, Cevallos SA, Lee J-Y, Li C, Bec J, Bäumler AJ, Marcu L. Assessment of Murine Colon Inflammation Using Intraluminal Fluorescence Lifetime Imaging. Molecules. 2022; 27(4):1317. https://doi.org/10.3390/molecules27041317
Chicago/Turabian StyleAlfonso-Garcia, Alba, Stephanie A. Cevallos, Jee-Yon Lee, Cai Li, Julien Bec, Andreas J. Bäumler, and Laura Marcu. 2022. "Assessment of Murine Colon Inflammation Using Intraluminal Fluorescence Lifetime Imaging" Molecules 27, no. 4: 1317. https://doi.org/10.3390/molecules27041317
APA StyleAlfonso-Garcia, A., Cevallos, S. A., Lee, J.-Y., Li, C., Bec, J., Bäumler, A. J., & Marcu, L. (2022). Assessment of Murine Colon Inflammation Using Intraluminal Fluorescence Lifetime Imaging. Molecules, 27(4), 1317. https://doi.org/10.3390/molecules27041317