
Coptisine Alleviates Imiquimod-Induced Psoriasis-like Skin Lesions and Anxiety-like Behavior in Mice



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effects of Coptisine on Psoriasis-like Skin Symptoms in IMQ-Treated Mice
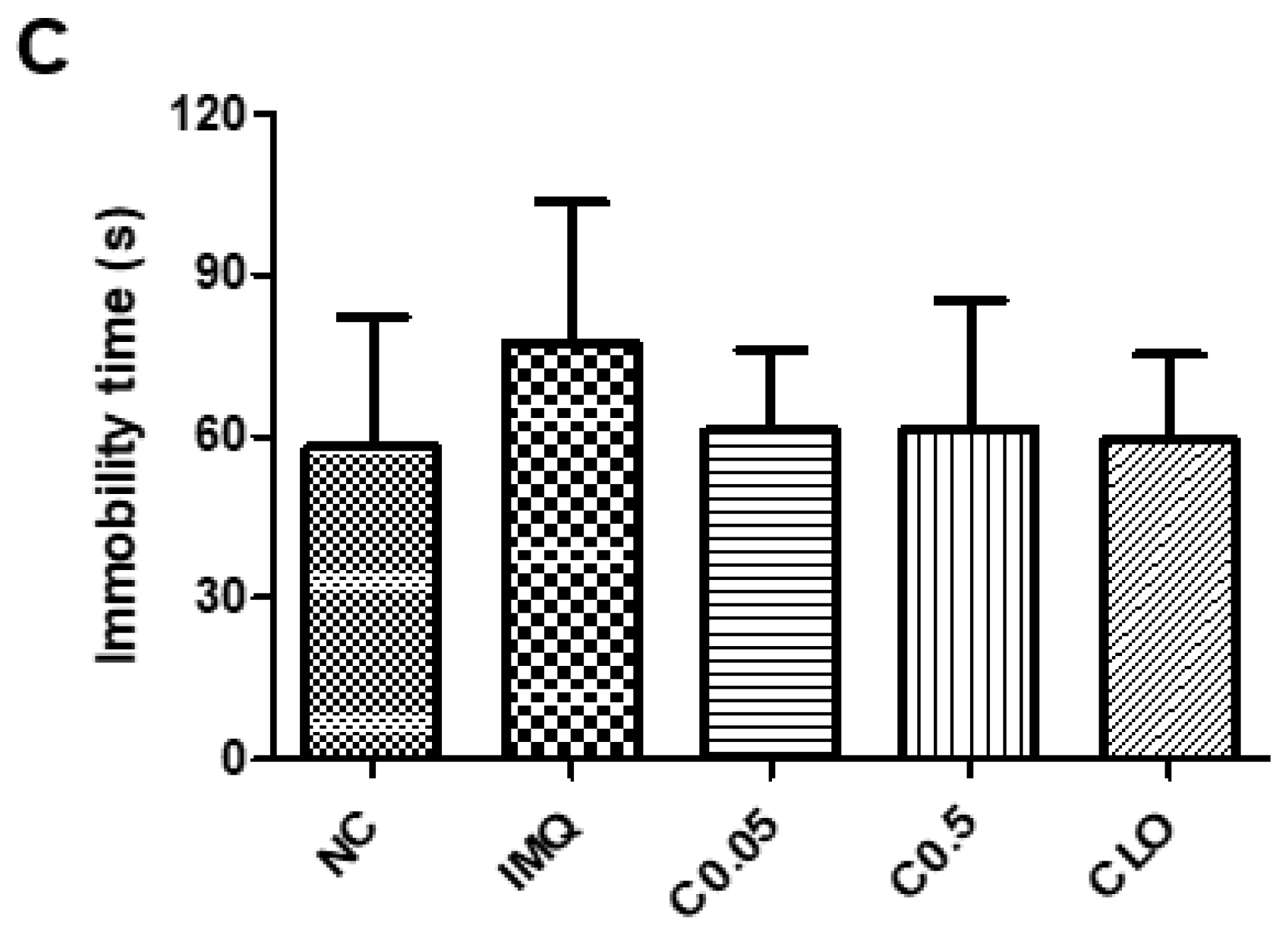
2.2. Effects of Coptisine on the Behavioral Changes in IMQ-Treated Mice
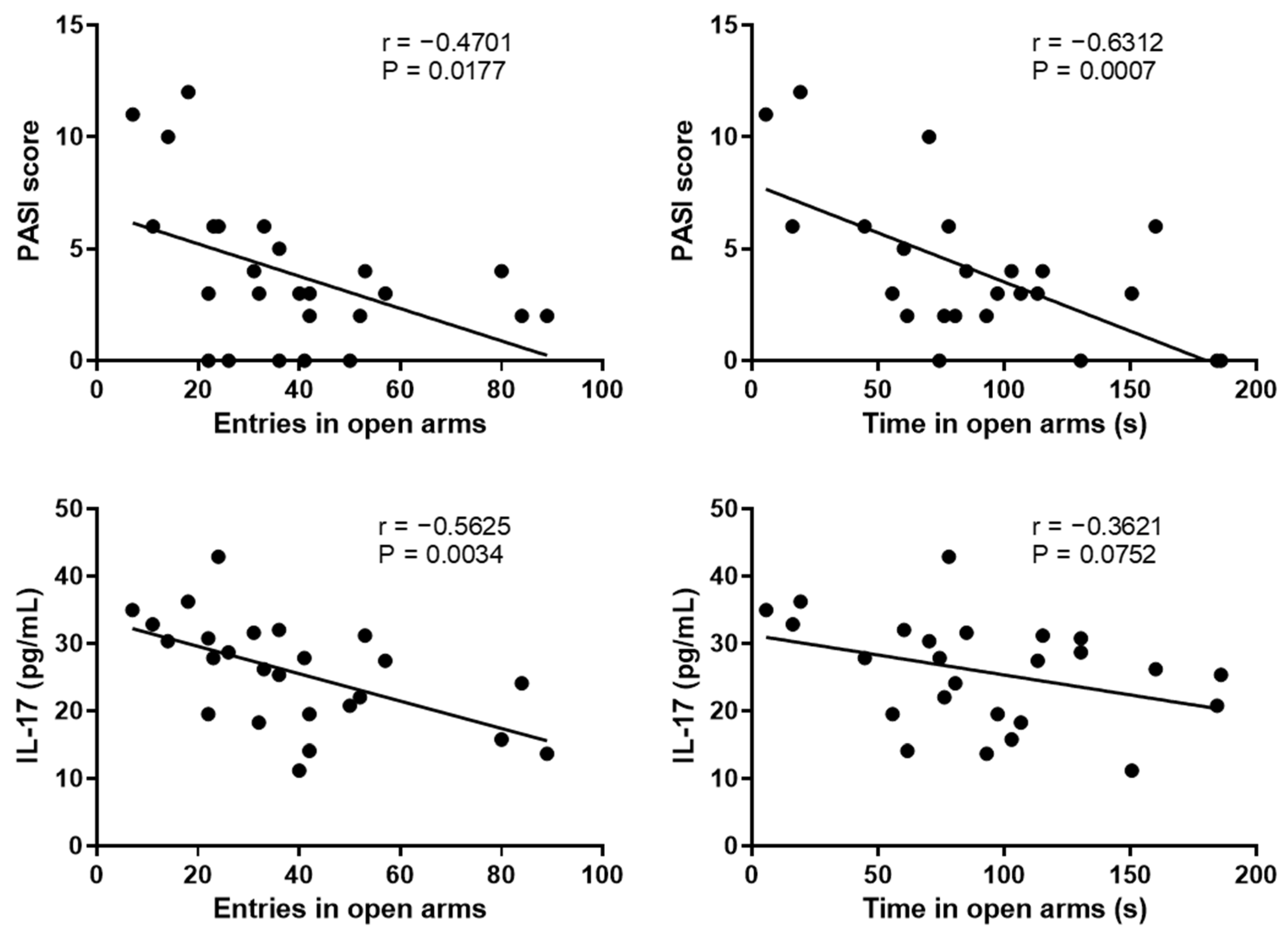
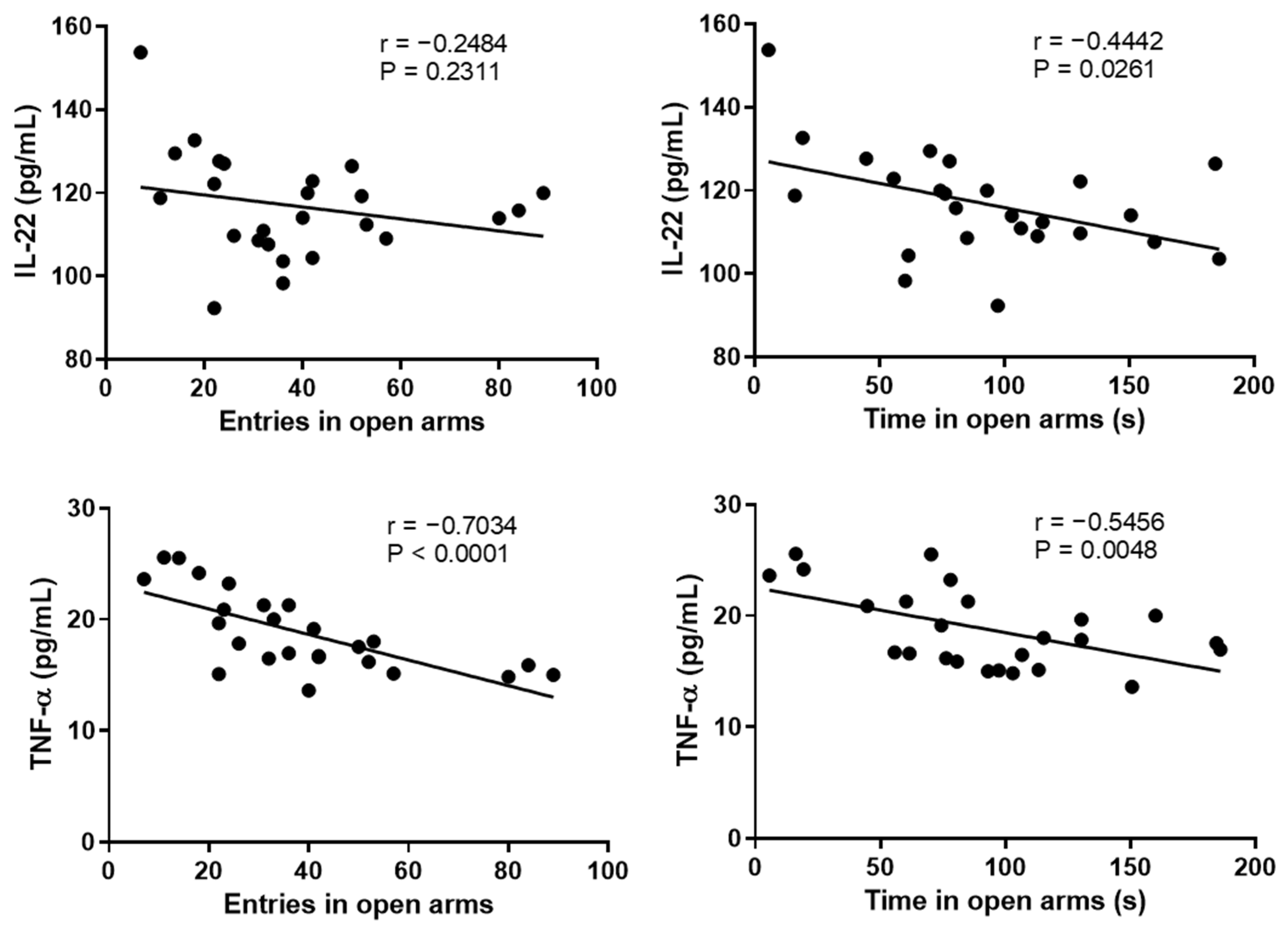
2.3. Correlation between the Anxiety-like Behavior and Psoriasis Severity
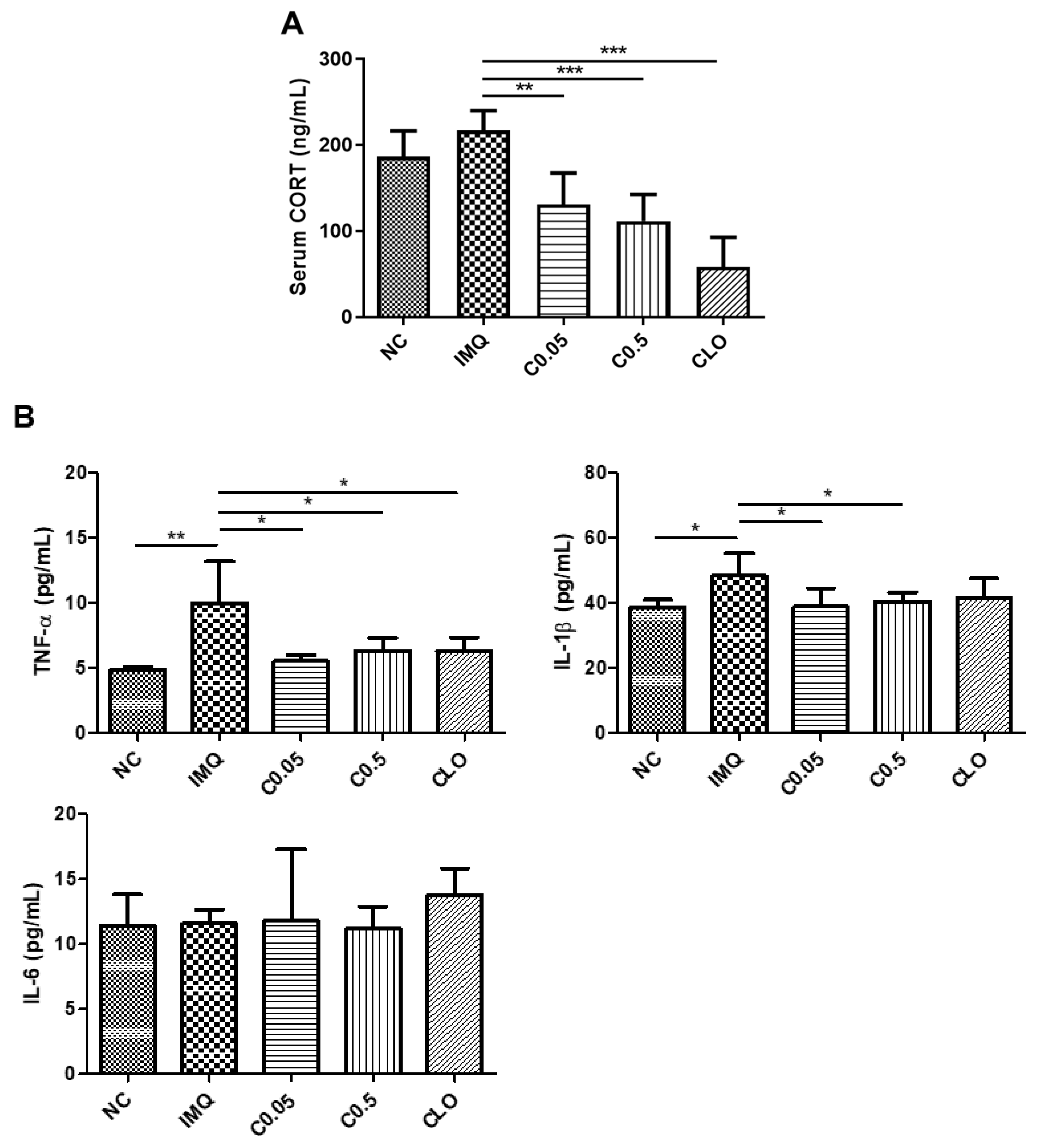
2.4. Effects of Coptisine on the Serum Corticosterone (CORT) Levels and Neuroinflammation in IMQ-Treated Mice
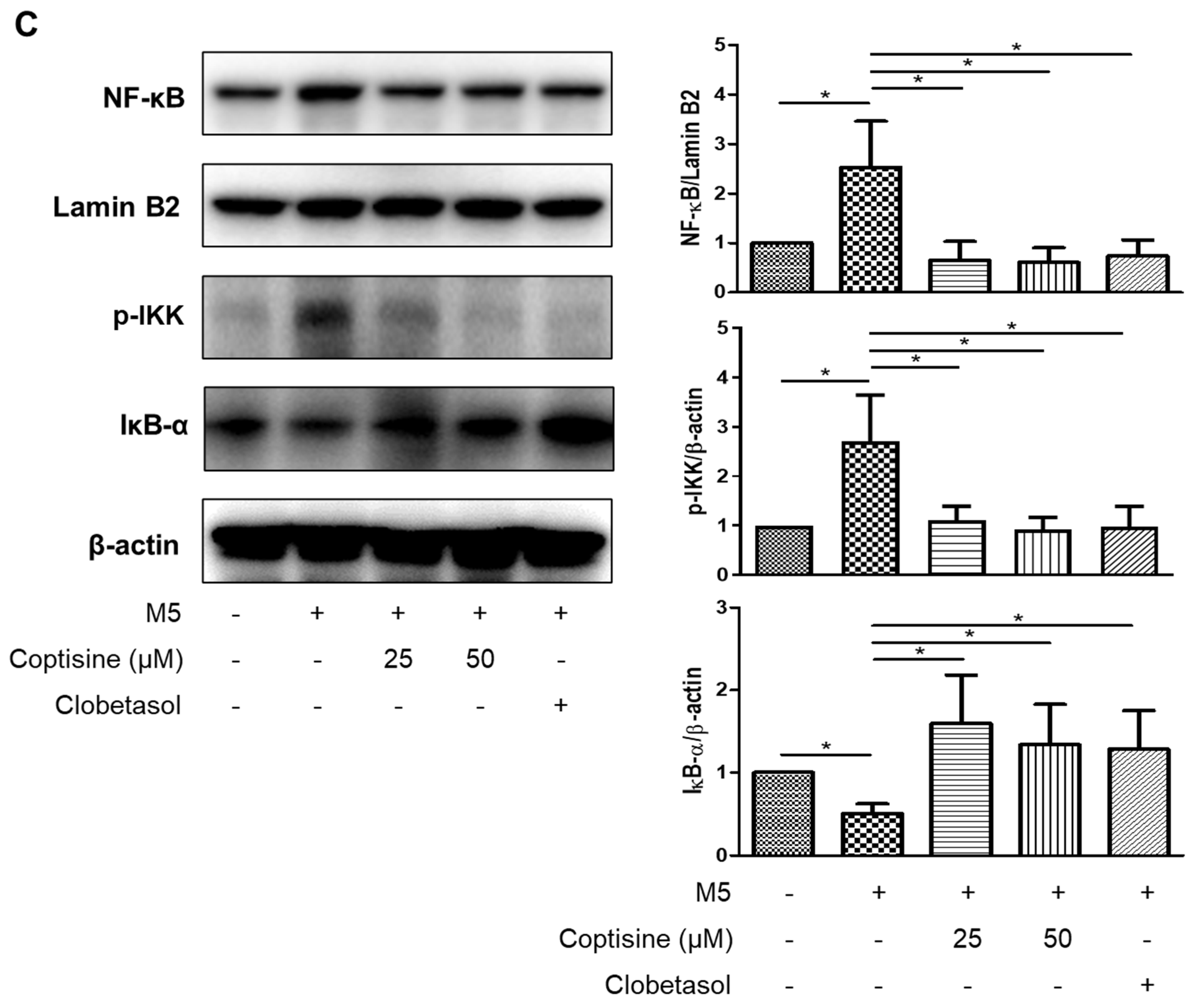
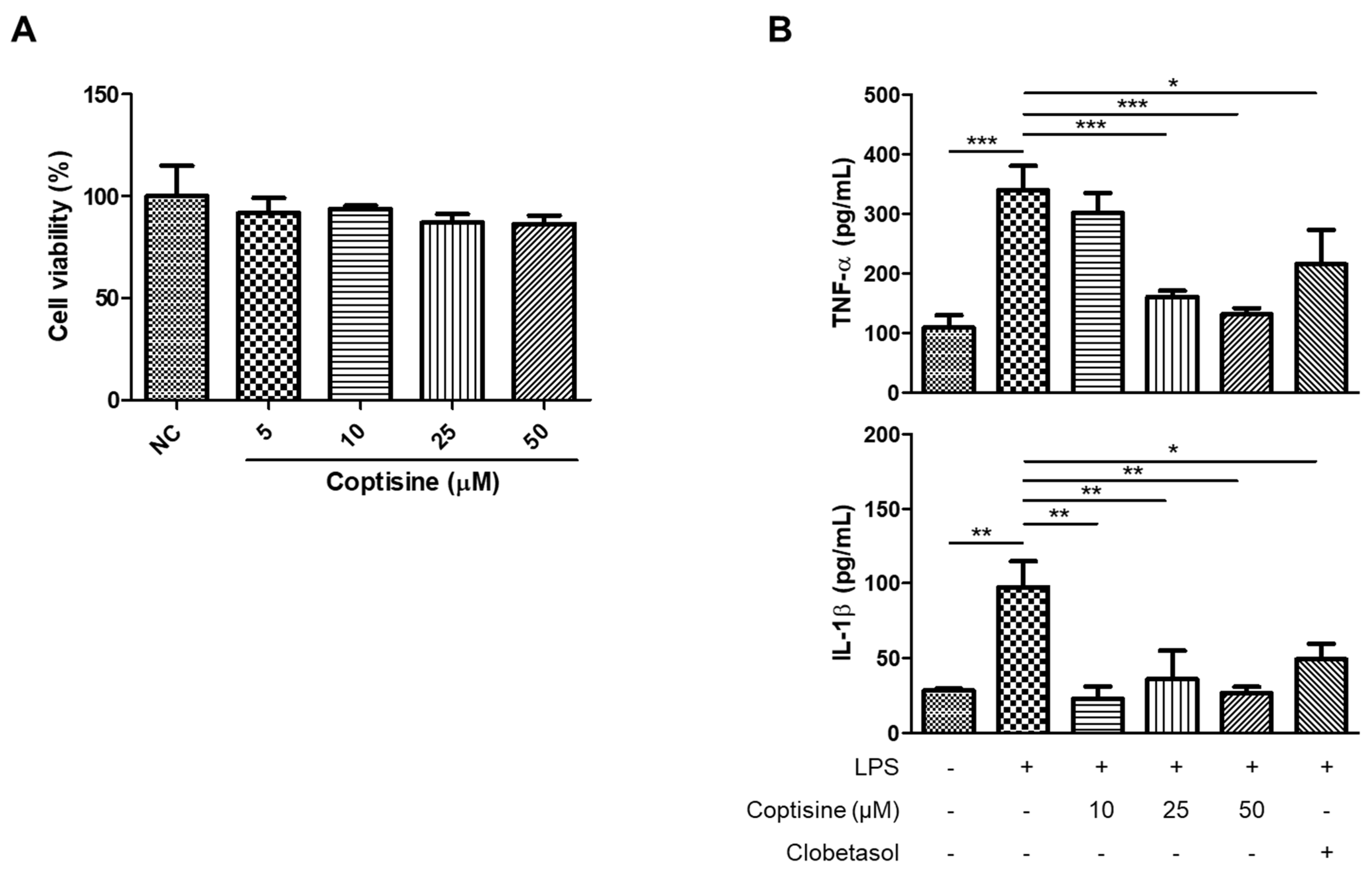
2.5. Effects of Coptisine on the Production of Inflammatory Cytokines and Chemokines in M5-Treated HaCaT Cells
2.6. Effects of Coptisine on the Production of Inflammatory Cytokines in LPS-Stimulated BV2 Cells
3. Discussion
4. Limitation
5. Materials and Methods
5.1. Animals
5.2. Elevated Plus Maze (EPM)
5.3. Tail Suspension Test (TST)
5.4. Histological Examination
5.5. Cell Culture
5.6. Cell Viability
5.7. Enzyme-Linked Immunosorbent Assay (ELISA)
5.8. Western Blot Analysis
5.9. Statistical Analysis
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| IMQ | imiquimod |
| TNF-α | tumor necrosis factor α |
| MIP-3α | macrophage inflammatory protein 3α |
| IP-10 | interferon γ–induced protein-10 |
| ICAM-1 | intercellular adhesion molecule-1 |
| LPS | lipopolysaccharide |
| CORT | corticosterone |
| PASI | Psoriasis Area Severity Index |
| NF-κB | nuclear factor kappa B |
| IκB-α | nuclear factor kappa B inhibitor alpha |
| IKK | IκB kinase |
References
- Perera, G.K.; Di Meglio, P.; Nestle, F.O. Psoriasis. Annu. Rev. Pathol. 2012, 7, 385–422. [Google Scholar] [CrossRef] [PubMed]
- Parisi, R.; Symmons, D.P.M.; Griffiths, C.E.M.; Ashcroft, D.M. Global Epidemiology of Psoriasis: A Systematic Review of Incidence and Prevalence. J. Investig. Dermatol. 2013, 133, 377–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Meglio, P.; Villanova, F.; Nestle, F.O. Psoriasis. Cold Spring Harb. Perspect. Med. 2014, 4, a015354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Albanesi, C.; Madonna, S.; Gisondi, P.; Girolomoni, G. The Interplay Between Keratinocytes and Immune Cells in the Pathogenesis of Psoriasis. Front. Immunol. 2018, 9, 1549. [Google Scholar] [CrossRef] [Green Version]
- Benhadou, F.; Mintoff, D.; del Marmol, V. Psoriasis: Keratinocytes or Immune Cells—Which Is the Trigger? Dermatology 2019, 235, 91–100. [Google Scholar] [CrossRef]
- Ferrari, S.M.; Ruffilli, I.; Colaci, M.; Antonelli, A.; Ferri, C.; Fallahi, P. CXCL10 in psoriasis. Adv. Med. Sci. 2015, 60, 349–354. [Google Scholar] [CrossRef]
- Gottlieb, A.B.; Krueger, J.G.; Wittkowski, K.; Dedrick, R.; Walicke, P.A.; Garovoy, M. Psoriasis as a Model for T-Cell–Mediated Disease: Immunobiologic and Clinical Effects of Treatment With Multiple Doses of Efalizumab, an Anti–CD11a Antibody. Arch. Dermatol. 2002, 138, 591–600. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Laumonnier, Y.; Syrovets, T.; Simmet, T. Recruitment of CCR6-expressing Th17 cells by CCL20 secreted from plasmin-stimulated macrophages. Acta Biochim. Biophys. Sin. 2013, 45, 593–600. [Google Scholar] [CrossRef] [Green Version]
- Rendon, A.; Schäkel, K. Psoriasis Pathogenesis and Treatment. Int. J. Mol. Sci. 2019, 20, 1475. [Google Scholar] [CrossRef] [Green Version]
- Oliveira Mde, F.; Rocha Bde, O.; Duarte, G.V. Psoriasis: Classical and emerging comorbidities. An. Bras. Dermatol. 2015, 90, 9–20. [Google Scholar] [CrossRef] [Green Version]
- Ni, C.; Chiu, M.W. Psoriasis and comorbidities: Links and risks. Clin. Cosmet. Investig. Dermatol. 2014, 7, 119–132. [Google Scholar] [PubMed]
- Dalgard, F.J.; Gieler, U.; Tomas-Aragones, L.; Lien, L.; Poot, F.; Jemec, G.B.E.; Misery, L.; Szabo, C.; Linder, D.; Sampogna, F.; et al. The Psychological Burden of Skin Diseases: A Cross-Sectional Multicenter Study among Dermatological Out-Patients in 13 European Countries. J. Investig. Dermatol. 2015, 135, 984–991. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, R.; Garner, M.; Holmes, C.; Osmond, C.; Teeling, J.; Lau, L.; Baldwin, D.S. Peripheral inflammatory cytokines and immune balance in Generalised Anxiety Disorder: Case-controlled study. Brain. Behav. Immun. 2017, 62, 212–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Felger, J.C.; Lotrich, F.E. Inflammatory cytokines in depression: Neurobiological mechanisms and therapeutic implications. Neuroscience 2013, 246, 199–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warrington, T.P.; Bostwick, J.M. Psychiatric adverse effects of corticosteroids. Mayo Clin. Proc. 2006, 81, 1361–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taper, C. Systemic corticosteroid–associated psychiatric adverse effects. US Pharm. 2016, 41, 16–18. [Google Scholar]
- Savas, M.; Vinkers, C.H.; Rosmalen, J.G.M.; Hartman, C.A.; Wester, V.L.; van den Akker, E.L.T.; Iyer, A.M.; McEwen, B.S.; van Rossum, E.F.C. Systemic and Local Corticosteroid Use Is Associated with Reduced Executive Cognition, and Mood and Anxiety Disorders. Neuroendocrinology 2020, 110, 282–291. [Google Scholar] [CrossRef]
- Strober, B.; Gooderham, M.; de Jong, E.M.; Kimball, A.B.; Langley, R.G.; Lakdawala, N.; Goyal, K.; Lawson, F.; Langholff, W.; Hopkins, L. Depressive symptoms, depression, and the effect of biologic therapy among patients in Psoriasis Longitudinal Assessment and Registry (PSOLAR). J. Am. Acad. Dermatol. 2018, 78, 70–80. [Google Scholar] [CrossRef] [Green Version]
- Vasilakis-Scaramozza, C.; Persson, R.; Hagberg, K.; Jick, S. The risk of treated anxiety and treated depression among patients with psoriasis and psoriatic arthritis treated with apremilast compared to biologics, DMARDs and corticosteroids: A cohort study in the United States MarketScan database. J. Eur. Acad. Dermatol. Venereol. 2020, 34, 1755–1763. [Google Scholar] [CrossRef]
- Wang, J.; Wang, L.; Lou, G.H.; Zeng, H.R.; Hu, J.; Huang, Q.W.; Peng, W.; Yang, X.B. Coptidis Rhizoma: A comprehensive review of its traditional uses, botany, phytochemistry, pharmacology and toxicology. Pharm. Biol. 2019, 57, 193–225. [Google Scholar] [CrossRef] [Green Version]
- Jung, A.R.; Ahn, S.H.; Jeong, H.S.; Kim, K.B. Effect of Coptidis rhizoma extract on atopic dermatitis-like skin lesions in NC/Nga mice. J. Physiol. Pathol. Korean. Med. 2019, 33, 102–108. [Google Scholar] [CrossRef]
- Meng, S.; Lin, Z.; Wang, Y.; Wang, Z.; Li, P.; Zheng, Y. Psoriasis therapy by Chinese medicine and modern agents. Chin. Med. 2018, 13, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Luo, Y.; Deng, D.; Su, S.; Li, S.; Xiang, L.; Hu, Y.; Wang, P.; Meng, X. Coptisine from Coptis chinensis exerts diverse beneficial properties: A concise review. J. Cell Mol. Med. 2019, 23, 7946–7960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, J.; Zhang, H.; Hu, B.; Yang, L.; Wang, P.; Wang, F.; Meng, X. Coptisine from Coptis chinensis inhibits production of inflammatory mediators in lipopolysaccharide-stimulated RAW 264.7 murine macrophage cells. Eur. J. Pharmacol. 2016, 780, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Wang, L.; Xiang, L.; Wu, J.; Huang, W.G.; Xu, C.; Meng, X.; Wang, P. Pharmacokinetic-Pharmacodynamic Modeling for Coptisine Challenge of Inflammation in LPS-Stimulated Rats. Sci. Rep. 2019, 9, 1450. [Google Scholar] [CrossRef]
- Zhou, K.; Hu, L.; Liao, W.; Yin, D.; Rui, F. Coptisine Prevented IL-[beta]-Induced Expression of Inflammatory Mediators in Chondrocytes. Inflammation 2016, 39, 1558–1565. [Google Scholar] [CrossRef]
- Friedemann, T.; Schumacher, U.; Tao, Y.; Leung, A.K.; Schröder, S. Neuroprotective Activity of Coptisine from Coptis chinensis (Franch). Evid.-Based Complement. Alternat. Med. 2015, 2015, 827308. [Google Scholar] [CrossRef] [Green Version]
- Fleming, P.; Bai, J.W.; Pratt, M.; Sibbald, C.; Lynde, C.; Gulliver, W.P. The prevalence of anxiety in patients with psoriasis: A systematic review of observational studies and clinical trials. J. Eur. Acad. Dermatol. Venereol. 2017, 31, 798–807. [Google Scholar] [CrossRef]
- Kouris, A.; Christodoulou, C.; Efstathiou, V.; Tsatovidou, R.; Torlidi-Kordera, E.; Zouridaki, E.; Kontochristopoulos, G. Comparative study of quality of life and psychosocial characteristics in patients with psoriasis and leg ulcers. Wound Repair Regen. 2016, 24, 443–446. [Google Scholar] [CrossRef]
- Tafet, G.E.; Nemeroff, C.B. Pharmacological Treatment of Anxiety Disorders: The Role of the HPA Axis. Front. Psychiatry 2020, 11, 443. [Google Scholar] [CrossRef]
- Won, E.; Kim, Y.-K. Neuroinflammation-Associated Alterations of the Brain as Potential Neural Biomarkers in Anxiety Disorders. Int. J. Mol. Sci. 2020, 21, 6546. [Google Scholar] [CrossRef] [PubMed]
- Guilloteau, K.; Paris, I.; Pedretti, N.; Boniface, K.; Juchaux, F.; Huguier, V.; Guillet, G.; Bernard, F.-X.; Lecron, J.-C.; Morel, F. Skin inflammation induced by the synergistic action of IL-17A, IL-22, oncostatin M, IL-1α, and TNF-α recapitulates some features of psoriasis. J. Immunol. 2010, 184, 5263–5270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golpour, M.; Hosseini, S.H.; Khademloo, M.; Ghasemi, M.; Ebadi, A.; Koohkan, F.; Shahmohammadi, S. Depression and Anxiety Disorders among Patients with Psoriasis: A Hospital-Based Case-Control Study. Dermatol. Res. Pract. 2012, 2012, 381905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xie, J.; Huang, S.; Huang, H.; Deng, X.; Yue, P.; Lin, J.; Yang, M.; Han, L.; Zhang, D.-K. Advances in the Application of Natural Products and the Novel Drug Delivery Systems for Psoriasis. Front. Pharmacol. 2021, 12, 12. [Google Scholar] [CrossRef]
- Bonesi, M.; Rosa Loizzo, M.; Provenzano, E.; Menichini, F.; Tundis, R. Anti-Psoriasis Agents from Natural Plant Sources. Curr. Med. Chem. 2016, 23, 1250–1267. [Google Scholar] [CrossRef] [PubMed]
- Lagunin, A.; Filimonov, D.; Poroikov, V. Multi-Targeted Natural Products Evaluation Based on Biological Activity Prediction with PASS. Curr. Pharm. Des. 2010, 16, 1703–1717. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Xiao, Q.-Q.; Li, F.-L.; Xu, R.; Fan, B.; Wu, M.-F.; Zhou, M.; Li, S.; Chen, J.; Peng, S.-G.; et al. Immune Signatures in Patients with Psoriasis Vulgaris of Blood-Heat Syndrome: A Systematic Review and Meta-Analysis. Evid.-Based Complement. Alternat. Med. 2016, 2016, 9503652. [Google Scholar] [CrossRef] [Green Version]
- Shang, B.; Zhang, H.; Lu, Y.; Zhou, X.; Wang, Y.; Ma, M.; Ma, K. Insights from the Perspective of Traditional Chinese Medicine to Elucidate Association of Lily Disease and Yin Deficiency and Internal Heat of Depression. Evid.-Based Complement. Alternat. Med. 2020, 2020, 8899079. [Google Scholar] [CrossRef]
- van der Fits, L.; Mourits, S.; Voerman, J.S.A.; Kant, M.; Boon, L.; Laman, J.D.; Cornelissen, F.; Mus, A.-M.; Florencia, E.; Prens, E.P.; et al. Imiquimod-Induced Psoriasis-Like Skin Inflammation in Mice Is Mediated via the IL-23/IL-17 Axis. J. Immunol. 2009, 182, 5836. [Google Scholar] [CrossRef]
- Jariwala, S.P. The role of dendritic cells in the immunopathogenesis of psoriasis. Arch. Dermatol. Res. 2007, 299, 359–366. [Google Scholar] [CrossRef] [Green Version]
- Lew, W.; Bowcock, A.M.; Krueger, J.G. Psoriasis vulgaris: Cutaneous lymphoid tissue supports T-cell activation and ‘Type 1’ inflammatory gene expression. Trends Immunol. 2004, 25, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Huang, L.; Lv, P.; Li, X.; Liu, G.; Chen, Y.; Wang, Z.; Qian, X.; Shen, Y.; Li, Y.; et al. The role of Th17 cells in psoriasis. Immunol. Res. 2020, 68, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.-B.; Luo, C.-D.; Liang, J.-L.; Zhang, Z.-B.; Lin, G.-S.; Wu, J.-Z.; Li, C.-L.; Tan, L.-H.; Yang, X.-B.; Su, Z.-R.; et al. Anti-inflammatory activity of coptisine free base in mice through inhibition of NF-κB and MAPK signaling pathways. Eur. J. Pharmacol. 2017, 811, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Uva, L.; Miguel, D.; Pinheiro, C.; Antunes, J.; Cruz, D.; Ferreira, J.; Filipe, P. Mechanisms of Action of Topical Corticosteroids in Psoriasis. Int. J. Endocrinol. 2012, 2012, 561018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, T.; Shao, S.; Li, B.; Jin, L.; Lei, J.; Qiao, H.; Wang, G. Up-regulation of Interferon-inducible protein 16 contributes to psoriasis by modulating chemokine production in keratinocytes. Sci. Rep. 2016, 6, 25381. [Google Scholar] [CrossRef] [Green Version]
- Fielding, C.A.; McLoughlin, R.M.; McLeod, L.; Colmont, C.S.; Najdovska, M.; Grail, D.; Ernst, M.; Jones, S.A.; Topley, N.; Jenkins, B.J. IL-6 Regulates Neutrophil Trafficking during Acute Inflammation via STAT3. J. Immunol. 2008, 181, 2189. [Google Scholar] [CrossRef]
- Blauvelt, A. IL-6 Differs from TNF-α: Unpredicted Clinical Effects Caused by IL-6 Blockade in Psoriasis. J. Investig. Dermatol. 2017, 137, 541–542. [Google Scholar] [CrossRef] [Green Version]
- Kanda, N.; Hau, C.S.; Tada, Y.; Tatsuta, A.; Sato, S.; Watanabe, S. Visfatin Enhances CXCL8, CXCL10, and CCL20 Production in Human Keratinocytes. Endocrinology 2011, 152, 3155–3164. [Google Scholar] [CrossRef] [Green Version]
- Furue, K.; Ito, T.; Tsuji, G.; Nakahara, T.; Furue, M. The CCL20 and CCR6 axis in psoriasis. Scand. J. Immunol. 2020, 91, e12846. [Google Scholar] [CrossRef] [Green Version]
- Ban, D.-H.; Koo, S.-W.; Kim, Y.-K.; Choi, G.-S.; Lee, J.-H. Expression of Intercellular Adhesion Molecule-l (ICAM-1) in Vascular Endothelium and Keratinocytes of Psoriatic Skin. Ann. Dermatol. 2000, 12, 259–263. [Google Scholar] [CrossRef] [Green Version]
- Goldminz, A.M.; Au, S.C.; Kim, N.; Gottlieb, A.B.; Lizzul, P.F. NF-κB: An essential transcription factor in psoriasis. J. Dermatol. Sci. 2013, 69, 89–94. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, Y.; Gaynor, R.B. IκB kinases: Key regulators of the NF-κB pathway. Trends Biochem. Sci. 2004, 29, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Zhang, Y.; Zhang, H.; Zhang, Y.; Gao, L.; Tong, X.; Xie, Y.; Hu, Q.; Chen, C.; Ding, S.; et al. RPL22 Overexpression Promotes Psoriasis-Like Lesion by Inducing Keratinocytes Abnormal Biological Behavior. Front. Immunol. 2021, 12, 12. [Google Scholar] [CrossRef] [PubMed]
- Moorchung, N.; Kulaar, J.S.; Chatterjee, M.; Vasudevan, B.; Tripathi, T.; Dutta, V. Role of NF-κB in the pathogenesis of psoriasis elucidated by its staining in skin biopsy specimens. Int. J. Dermatol. 2014, 53, 570–574. [Google Scholar] [CrossRef]
- Andrés, R.M.; Montesinos, M.C.; Navalón, P.; Payá, M.; Terencio, M.C. NF-κB and STAT3 Inhibition as a Therapeutic Strategy in Psoriasis: In Vitro and In Vivo Effects of BTH. J. Investig. Dermatol. 2013, 133, 2362–2371. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Chen, F.; Fang, H.; Mi, J.; Qi, Q.; Yang, M. Daphnetin inhibits proliferation and inflammatory response in human HaCaT keratinocytes and ameliorates imiquimod-induced psoriasis-like skin lesion in mice. Biol. Res. 2020, 53, 48. [Google Scholar] [CrossRef]
- Bakar, R.S.; Jaapar, S.Z.S.; Azmi, A.F.; Aun, Y.C. Depression and anxiety among patients with psoriasis: A correlation with quality of life and associated factors. J. Taibah Univ. Med. Sci. 2021, 16, 491–496. [Google Scholar] [CrossRef]
- Walf, A.A.; Frye, C.A. The use of the elevated plus maze as an assay of anxiety-related behavior in rodents. Nat. Protoc. 2007, 2, 322–328. [Google Scholar] [CrossRef] [Green Version]
- JiaWen, W.; Hong, S.; ShengXiang, X.; Jing, L. Depression- and anxiety-like behaviour is related to BDNF/TrkB signalling in a mouse model of psoriasis. Clin. Exp. Dermatol. 2018, 43, 254–261. [Google Scholar] [CrossRef]
- Guo, J.; Liu, Y.; Guo, X.; Meng, Y.; Qi, C.; Zhao, J.; Di, T.; Zhang, L.; Guo, X.; Wang, Y.; et al. Depressive-like behaviors in mice with Imiquimod-induced psoriasis. Int. Immunopharmacol. 2020, 89, 107057. [Google Scholar] [CrossRef]
- Krishnan, V.; Nestler, E.J. Animal models of depression: Molecular perspectives. Curr. Top. Behav. Neurosci. 2011, 7, 121–147. [Google Scholar] [PubMed] [Green Version]
- Fourrier, C.; Bosch-Bouju, C.; Boursereau, R.; Sauvant, J.; Aubert, A.; Capuron, L.; Ferreira, G.; Layé, S.; Castanon, N. Brain tumor necrosis factor-α mediates anxiety-like behavior in a mouse model of severe obesity. Brain Behav. Immun. 2019, 77, 25–36. [Google Scholar] [CrossRef] [PubMed]
- Alshammari, M.A.; Khan, M.R.; Majid Mahmood, H.; Alshehri, A.O.; Alasmari, F.F.; Alqahtani, F.M.; Alasmari, A.F.; Alsharari, S.D.; Alhossan, A.; Ahmad, S.F.; et al. Systemic TNF-α blockade attenuates anxiety and depressive-like behaviors in db/db mice through downregulation of inflammatory signaling in peripheral immune cells. Saudi Pharm. J. 2020, 28, 621–629. [Google Scholar] [CrossRef] [PubMed]
- Rossi, S.; Sacchetti, L.; Napolitano, F.; De Chiara, V.; Motta, C.; Studer, V.; Musella, A.; Barbieri, F.; Bari, M.; Bernardi, G.; et al. Interleukin-1β Causes Anxiety by Interacting with the Endocannabinoid System. J. Neurosci. 2012, 32, 13896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.G.; Huh, E.; Lee, W.; Kim, Y.-K.; Lee, T.-H.; Oh, M.S. Anti-inflammatory effects of Coptidis Rhizoma in chronic cold stress-exposed mice. Kor. J. Herbol. 2018, 33, 35–42. [Google Scholar]
- Almeida, P.G.C.; Nani, J.V.; Oses, J.P.; Brietzke, E.; Hayashi, M.A.F. Neuroinflammation and glial cell activation in mental disorders. Brain Behav. Immun. 2020, 2, 100034. [Google Scholar] [CrossRef]
- Stein, D.J.; Vasconcelos, M.F.; Albrechet-Souza, L.; Ceresér, K.M.M.; de Almeida, R.M.M. Microglial Over-Activation by Social Defeat Stress Contributes to Anxiety- and Depressive-Like Behaviors. Front. Behav. Neurosci. 2017, 11, 207. [Google Scholar] [CrossRef] [Green Version]
- Nam, H.Y.; Nam, J.H.; Yoon, G.; Lee, J.-Y.; Nam, Y.; Kang, H.-J.; Cho, H.-J.; Kim, J.; Hoe, H.-S. Ibrutinib suppresses LPS-induced neuroinflammatory responses in BV2 microglial cells and wild-type mice. J. Neuroinflamm. 2018, 15, 271. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Yuan, C.; Wang, G.; Luo, J.; Ma, H.; Xu, L.; Mu, Y.; Li, Y.; Seeram, N.P.; Huang, X. Urolithins attenuate LPS-induced neuroinflammation in BV2Microglia via MAPK, Akt, and NF-κB signaling pathways. J. Agric. Food Chem. 2018, 66, 571–580. [Google Scholar] [CrossRef]
- Todo, H. Transdermal Permeation of Drugs in Various Animal Species. Pharmaceutics 2017, 9, 33. [Google Scholar] [CrossRef] [Green Version]
- Su, J.; Miao, Q.; Miao, P.; Zhao, Y.; Zhang, Y.; Chen, N.; Zhang, Y.; Ma, S. Pharmacokinetics and Brain Distribution and Metabolite Identification of Coptisine, a Protoberberine Alkaloid with Therapeutic Potential for CNS Disorders, in Rats. Biol. Pharm. Bull. 2015, 38, 1518–1528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.; Lyga, J. Brain-skin connection: Stress, inflammation and skin aging. Inflamm. Allergy Drug Targets 2014, 13, 177–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Feng, J.; Lv, W.; Huang, Q.; Fu, M.; Cai, M.; He, Q.; Shang, J. Developmental Neurotoxic Effects of Percutaneous Drug Delivery: Behavior and Neurochemical Studies in C57BL/6 Mice. PLoS ONE 2016, 11, e0162570. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Meng, X.; Wang, C.; Zhang, H.; Chen, H.; Deng, P.; Liu, J.; Huandike, M.; Wei, J.; Chai, L. Coptidis alkaloids extracted from Coptis chinensis Franch attenuate IFN-γ-induced destruction of bone marrow cells. PLoS ONE 2020, 15, e0236433. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Li, X.; Deng, P.; Wei, Y.; Liu, J.; Chen, M.; Xu, Y.; Zhang, D.; Zhu, L.; Lou, L.; et al. Chinese Herbal Formula, Modified Danggui Buxue Tang, Attenuates Apoptosis of Hematopoietic Stem Cells in Immune-Mediated Aplastic Anemia Mouse Model. J. Immunol. Res. 2017, 2017, 9786972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Can, A.; Dao, D.T.; Terrillion, C.E.; Piantadosi, S.C.; Bhat, S.; Gould, T.D. The tail suspension test. J. Vis. Exp. 2012, 59, e3769. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguyen, L.T.H.; Choi, M.-J.; Shin, H.-M.; Yang, I.-J. Coptisine Alleviates Imiquimod-Induced Psoriasis-like Skin Lesions and Anxiety-like Behavior in Mice. Molecules 2022, 27, 1412. https://doi.org/10.3390/molecules27041412
Nguyen LTH, Choi M-J, Shin H-M, Yang I-J. Coptisine Alleviates Imiquimod-Induced Psoriasis-like Skin Lesions and Anxiety-like Behavior in Mice. Molecules. 2022; 27(4):1412. https://doi.org/10.3390/molecules27041412
Chicago/Turabian StyleNguyen, Ly Thi Huong, Min-Jin Choi, Heung-Mook Shin, and In-Jun Yang. 2022. "Coptisine Alleviates Imiquimod-Induced Psoriasis-like Skin Lesions and Anxiety-like Behavior in Mice" Molecules 27, no. 4: 1412. https://doi.org/10.3390/molecules27041412