
Carotenoids and Their Biosynthesis in Fungi

Abstract
:1. Introduction
2. Evolutionary Origin of Fungal Carotenoid Biosynthesis
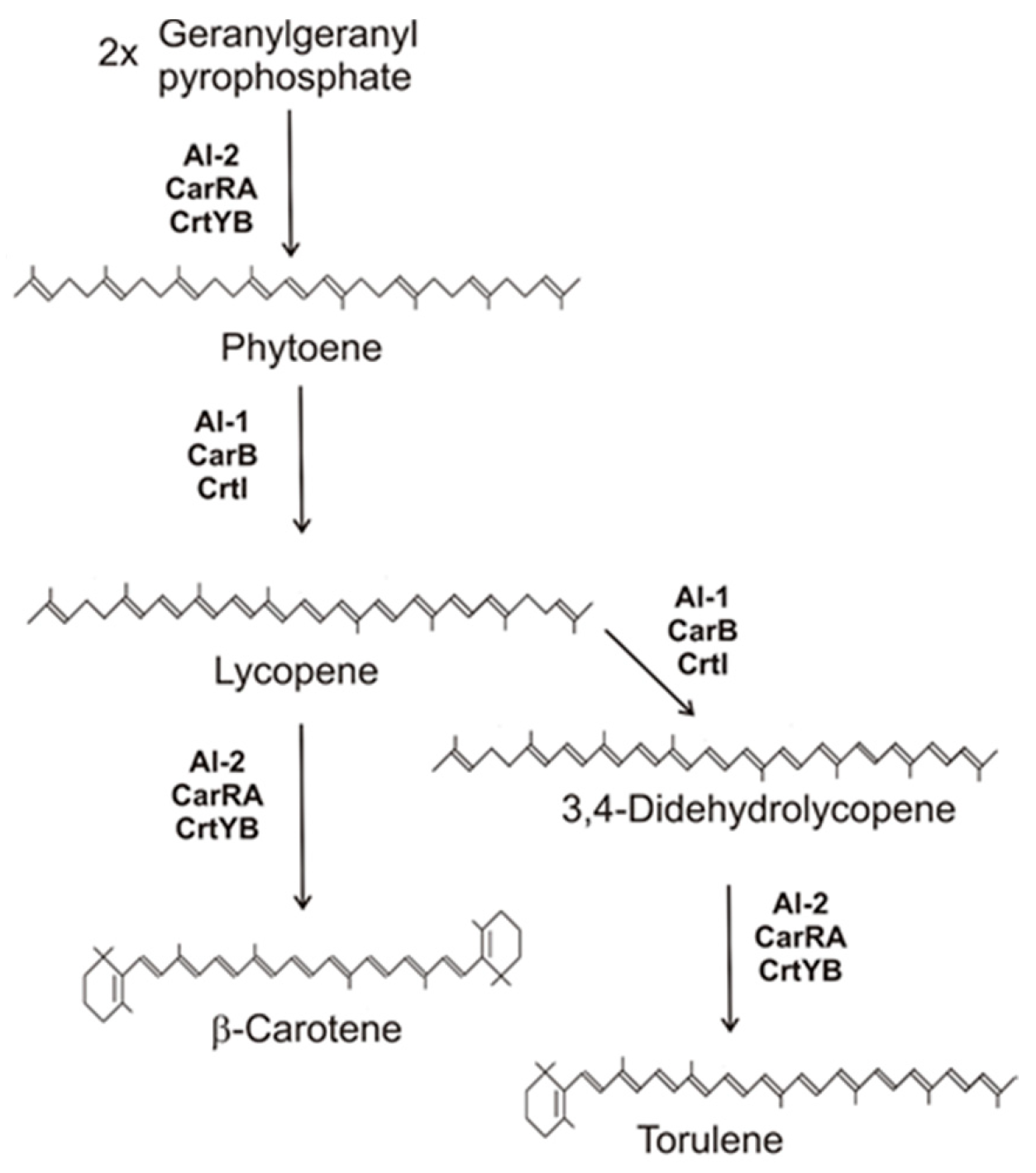
2.1. The Basic Pathway to β-Carotene
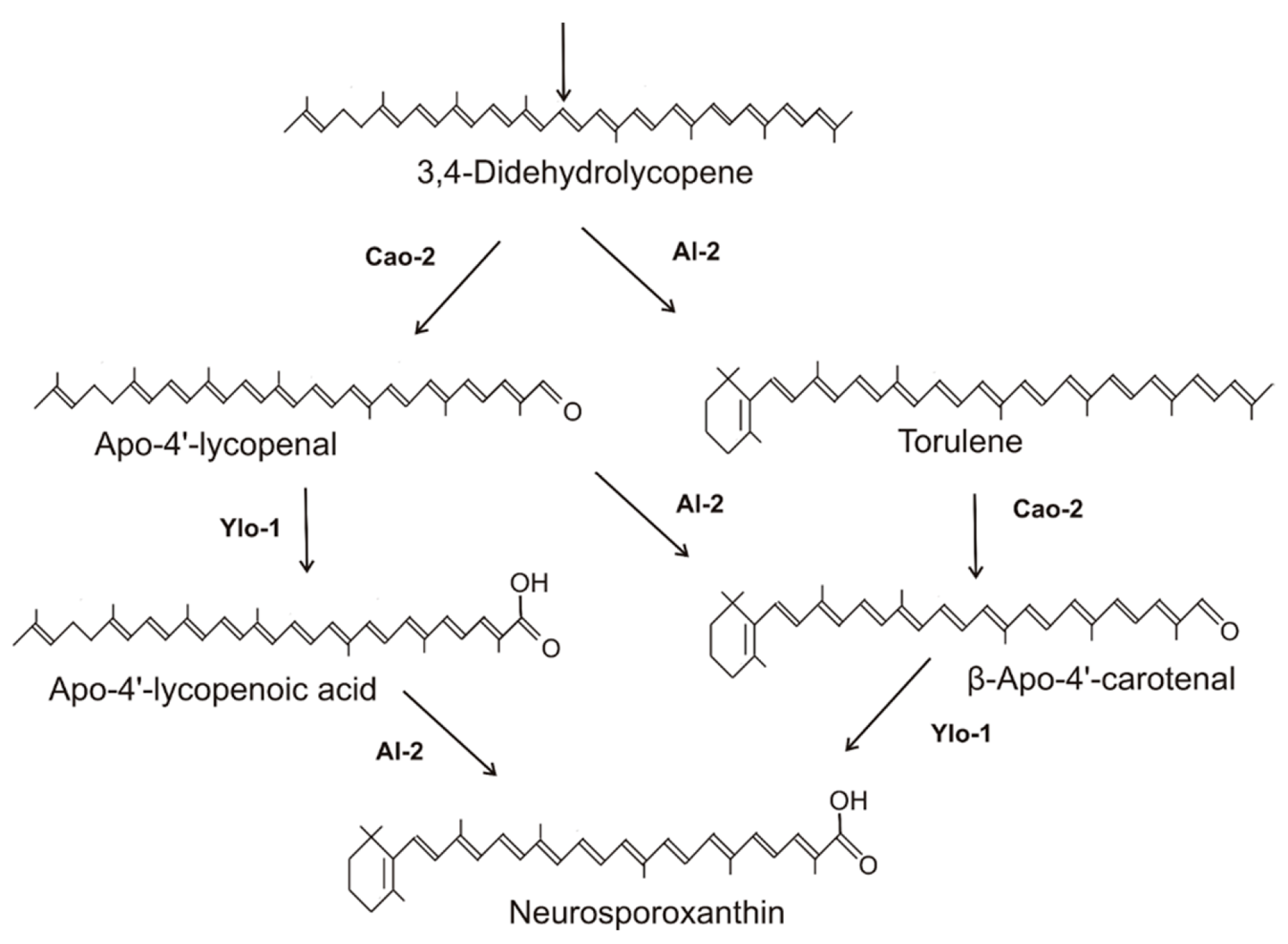
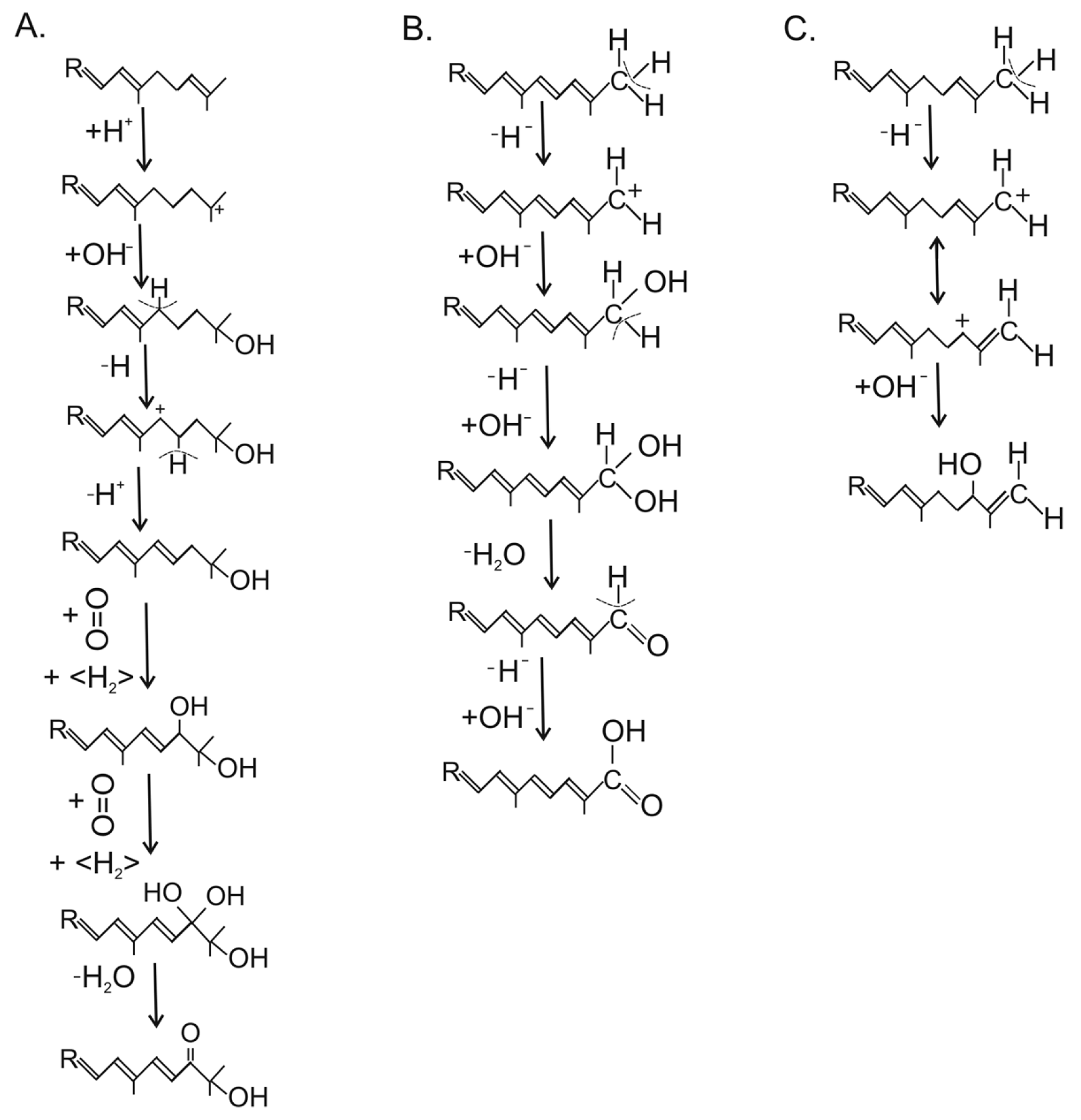
2.2. Synthesis of Neurosporaxanthin in Sordariomycetes
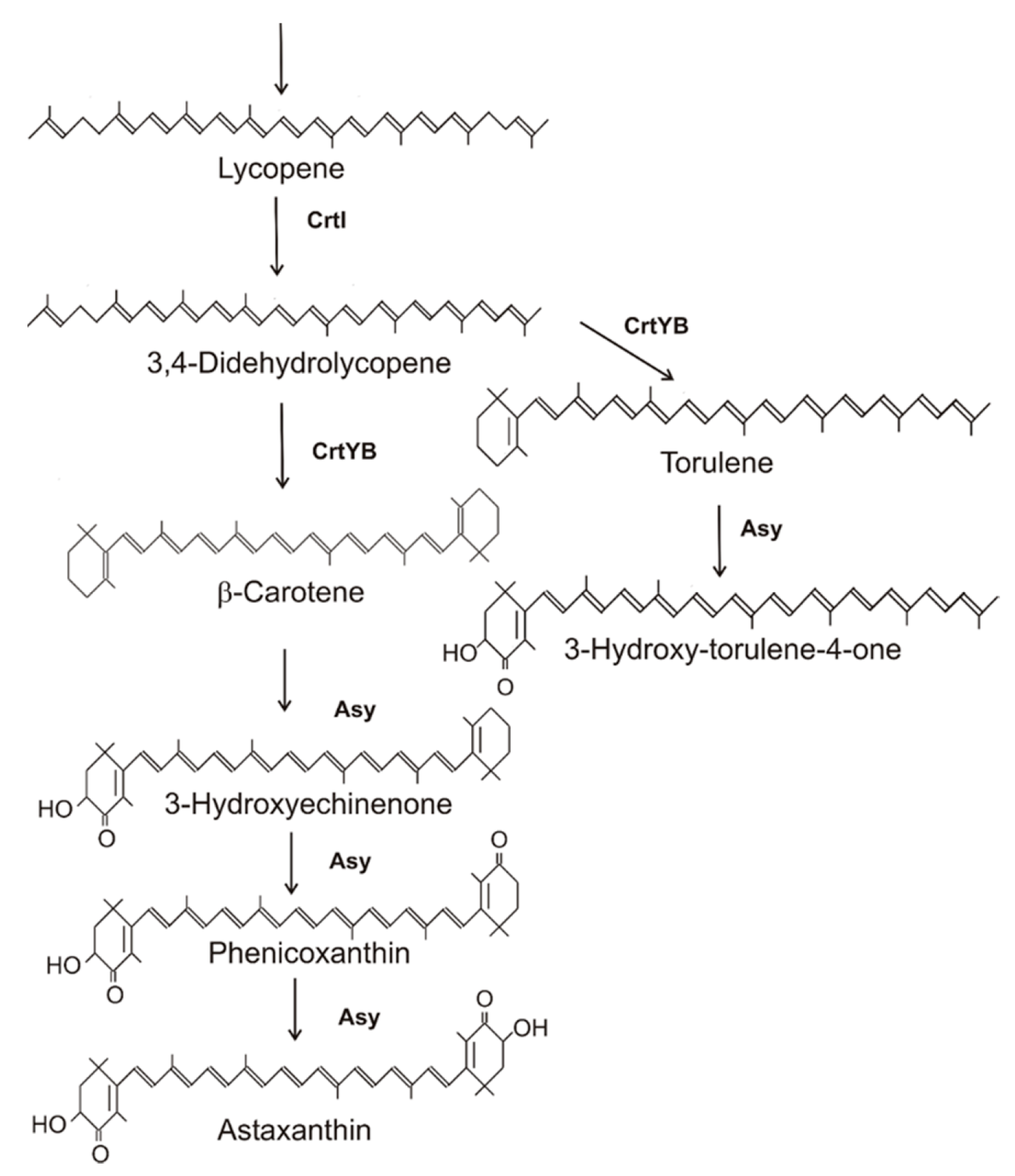
2.3. Synthesis of Astaxanthin in Xanthophyllomyces denrorhous
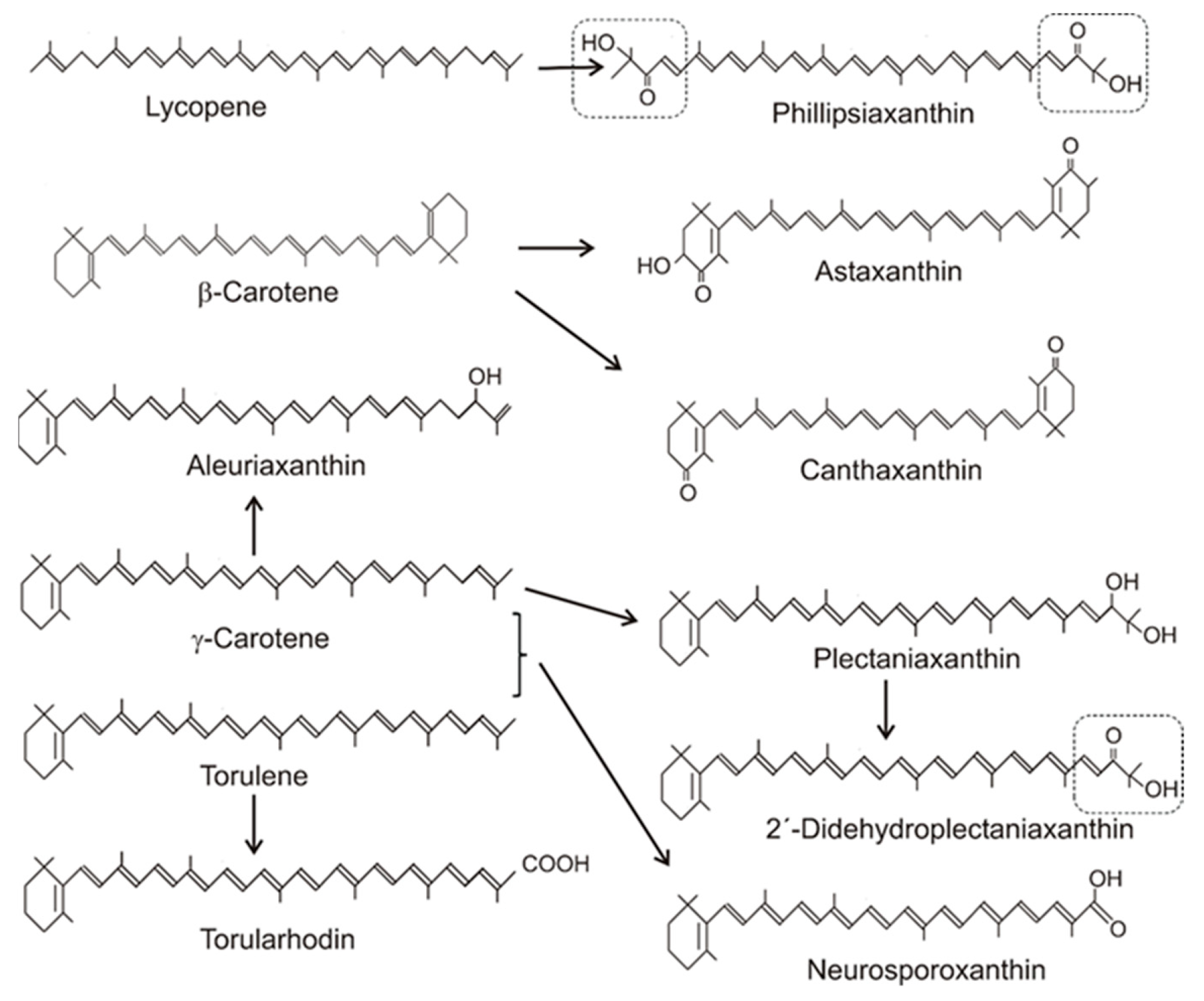
3. Formation of Carotenoid Structures and Their Distribution in Fungi
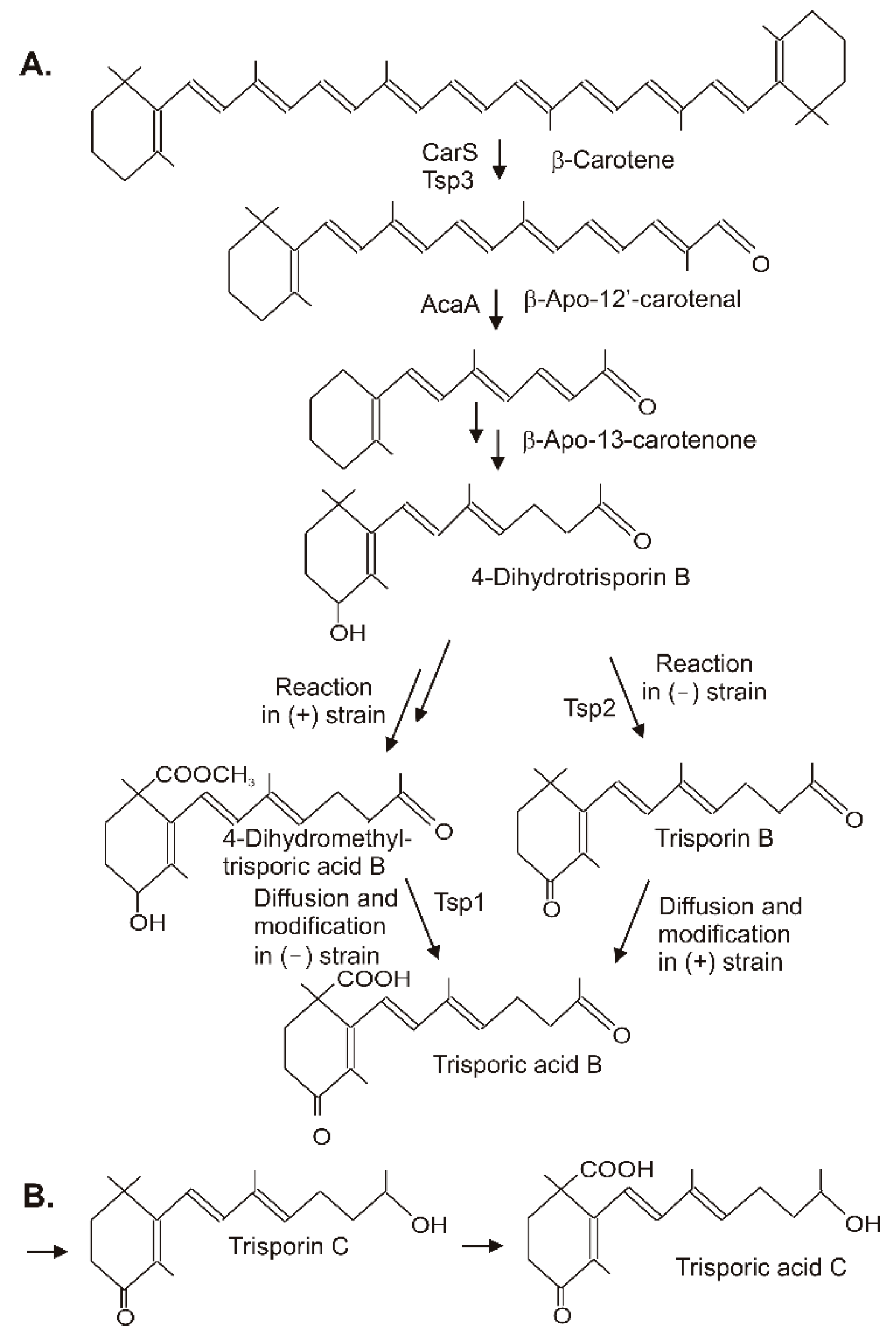
4. Conversion of β-Carotene to Trisporic Acid in the Mucorales
5. High-Yield Carotenoid Production
5.1. Classical Strain Development by Mutagenesis and Selection for Enhanced Carotenoid Production
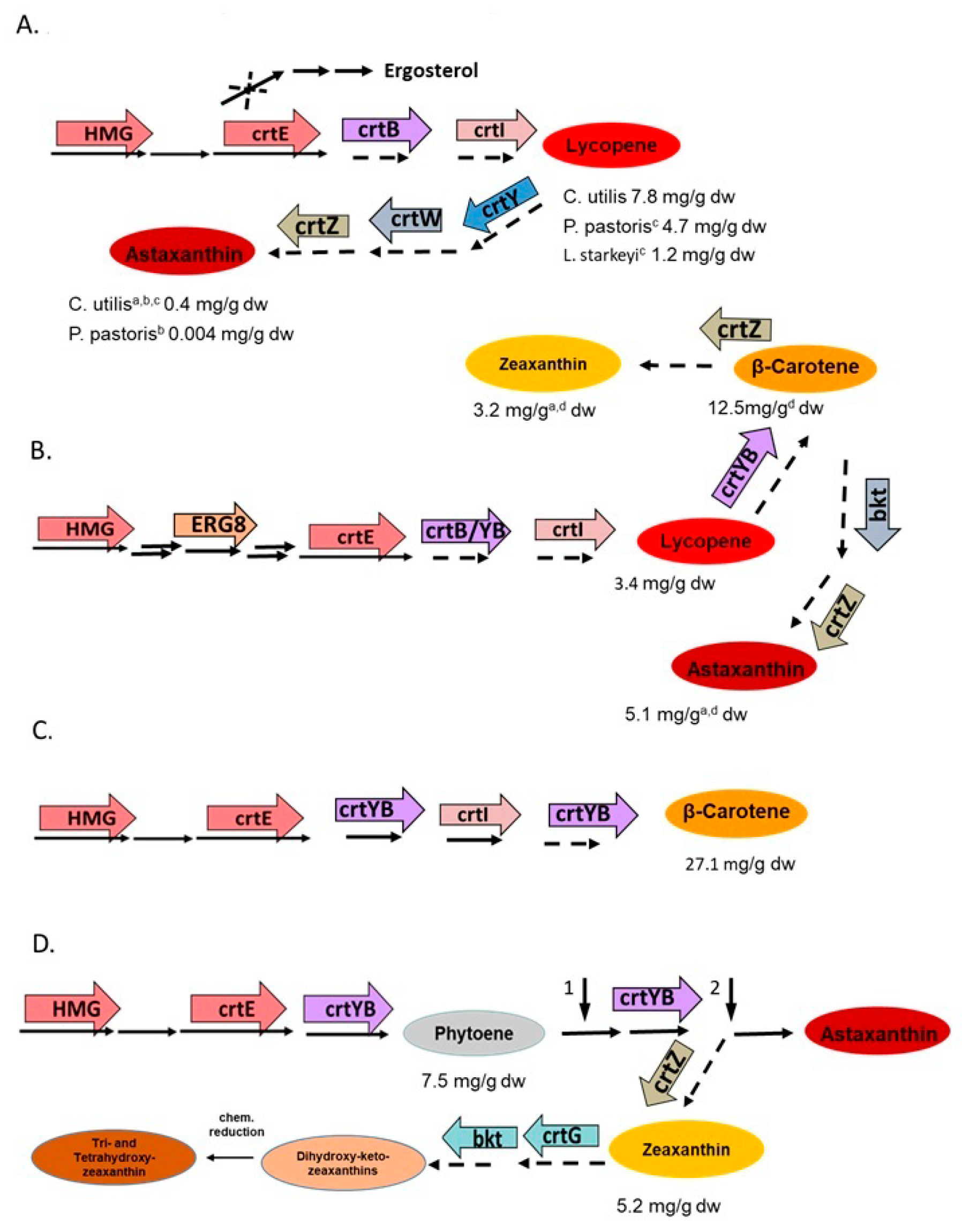
5.2. Metabolic Engineering of Carotenoid Pathways in Non-Conventional Yeasts
5.3. Pathway Engineering of Saccharomyces Cerevisiae for High-Yield Carotenoid Formation
6. Conclusions
Funding
Conflicts of Interest
Abbreviations
| CoA | coenzyme A |
| dw | dry weight |
| HMG | hydroxymethylglutaryl |
| NADPH | nicotinamide adenine dinucleotide phosphate |
References
- Goodwin, T.W. The Biochemistry of the Carotenoids, 2nd ed.; Chapman and Hall: London, UK; New York, NY, USA, 1980; Volumn I, Chapter 8; pp. 257–290. [Google Scholar]
- Valadon, L.R.G. Carotenoids as additional taxonomic characters in fungi: A review. Trans. Br. Mycol. Soc. 1976, 67, 1–15. [Google Scholar] [CrossRef]
- Sandmann, G.; Misawa, N. Fungal carotenoids. In The Mycota X. Industrial Applications; Osiewacz, H.D., Ed.; Springer: Berlin, Germany, 2002; pp. 247–262. [Google Scholar]
- Sandmann, G. Antioxidant protection from UV- and light-stress related to carotenoid structures. Antioxidants 2019, 8, 219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Will, O.H., III; Jankowski, P.; Kowacs, A.; Rossing, W.; Schneider, P.; Newland, N.A. A comparison of photo-killing among carotene and cytochrome c accumulating strains of the smut fungus Ustilaga violacea at specific wavelengths from 400 to 600 nm. Photochem. Photobiol. 1987, 45, 609–615. [Google Scholar] [CrossRef]
- Blanc, P.L.; Tuveson, R.W.; Sargent, M.L. Inactivation of carotenoid-producing and albino strains of Neurospora crassa by visible light, blacklight, and ultraviolet radiation. J. Bacteriol. 1976, 125, 616–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rau, W. Mechanism of photoregulation of carotenoid biosynthesis in plants. Pure Appl. Chem. 1985, 57, 777–784. [Google Scholar] [CrossRef]
- Schimek, C.; Wöstemeyer, J. Carotene derivatives in sexual communication of zygomycete fungi. Phytochemistry 2009, 70, 1867–1875. [Google Scholar] [CrossRef]
- Avalos, J.; Limón, M.C. Biological roles of fungal carotenoids. Curr. Genet. 2015, 61, 309–324. [Google Scholar] [CrossRef]
- Braithwaite, G.D.; Goodwin, T.W. Studies in Carotenogenesis 26. The incorporation of [14C] acetate, [14C] mevalonate and 14CO2 into β-carotene by the fungus Phycomyces blakesleeanus. Biochem. J. 1960, 76, 5–10. [Google Scholar] [CrossRef]
- Grob, C.; Butler, R. Über die Biosynthese des β-Carotins bei Mucor hiemalis Wehrner. Die Beteiligung der Essigsaure am Aufbau der Carotinmolekel, insbesondere in den Stellungen 3,4,6 bzw. 3’, 4’,6’, untersucht mit Hilfe von 14C-markierterEssigsäure. Helvet. Chim. Acta 1956, 39, 1775–1980. [Google Scholar] [CrossRef]
- Lee, T.C.; Chichester, C.O. Geranylgeranyl pyrophosphate as the condensing unit for enzymatic synthesis of carotenes. Phytochemistry 1969, 8, 603–609. [Google Scholar] [CrossRef]
- Bramley, P.M.; Davies, B. Carotene biosynthesis by cell extracts of mutants of Phycomyces blakesleeanus. Phytochemistry 1975, 14, 463–469. [Google Scholar] [CrossRef]
- Eme, L.; Spang, A.; Lombard, J.; Stairs, C.W.; Ettema, T.J.G. Archaea and the origin of eukaryotes. Nat. Rev. Microbiol. 2017, 15, 711–723. [Google Scholar] [CrossRef] [PubMed]
- Peck, R.F.; Johnson, E.A.; Krebs, M.P. Identification of a lycopene beta-cyclase required for bacteriorhodopsin biogenesis in the archaeon Halobacterium salinarum. J. Bacteriol. 2002, 184, 2889–2897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verdoes, J.C.; Krubasik, P.; Sandmann, G.; van Ooyen, A.J.J. Isolation and functional characterisation of a novel type of carotenoid biosynthetic gene from Xanthophyllomyces dendrorhous. Mol. Gen. Genet. 1999, 262, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Hemmi, H.; Ikejiri, S.; Nakayama, T.; Nishino, T. Fusion-type lycopene β-cyclase from a thermoacidophilic archaeon Sulfolobus solfataricus. Biochem. Biophys. Res. Commun. 2003, 305, 586–591. [Google Scholar] [CrossRef]
- Lombard, J.; Moreira, D. Origins and early evolution of the mevalonate pathway of isoprenoid biosynthesis in the three domains of life. Mol. Biol. Evol. 2011, 28, 87–99. [Google Scholar] [CrossRef] [Green Version]
- Lange, B.M.; Rujan, T.; Martin, W.; Croteau, R. Isoprenoid biosynthesis: The evolution of two ancient and distinct pathways across genomes. Proc. Natl. Acad. Sci. USA 2000, 97, 13172–13177. [Google Scholar] [CrossRef] [Green Version]
- Almeida, E.R.A.; Cerdá-Olmedo, E. Gene expression in the regulation of carotene biosynthesis in Phycomyces. Curr. Genet. 2008, 53, 129–137. [Google Scholar] [CrossRef]
- Moran, N.A.; Jarvik, T. Lateral transfer of genes from fungi underlies carotenoid production in aphids. Science 2010, 328, 624–627. [Google Scholar] [CrossRef] [Green Version]
- Schmidhauser, T.J.; Lauter, F.R.; Schumacher, M.; Zhou, W.; Russo, V.E.; Yanofsky, C. Characterization of al-2, the phytoene synthase gene of Neurospora crassa. Cloning, sequence analysis, and photoregulation. J. Biol. Chem. 1994, 269, 12060–12066. [Google Scholar] [CrossRef]
- Avalos, J.; Pardo-Medina, J.; Parra-Rivero, O.; Ruger-Herreros, M.; Rodríguez-Ortiz, R.; Hornero-Méndez, D.; Limón, M.C.J. Carotenoid biosynthesis in Fusarium. J. Fungi 2017, 3, 39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidhauser, T.J.; Lauter, F.R.; Russo, V.E.; Yanofsky, C. Cloning, sequence, and photoregulation of al-1, a carotenoid biosynthetic gene. Mol. Cell. Biol. 1990, 10, 5064–5070. [Google Scholar] [PubMed] [Green Version]
- Verdoes, J.C.; Misawa, N.; van Ooyen, A.J. Cloning and characterization of the astaxanthin biosynthetic gene encoding phytoene desaturase of Xanthophyllomyces dendrorhous. Biotechnol. Bioeng. 1999, 63, 750–755. [Google Scholar] [CrossRef]
- Breitenbach, J.; Fraser, P.D.; Sandmann, G. Carotenoid synthesis and phytoene synthase activity during mating of Blakeslea trispora. Phytochemistry 2012, 76, 40–45. [Google Scholar] [CrossRef]
- Ruiz-Hidalgo, M.R.; Benito, E.P.; Sandmann, G.; Eslava, A.P. The phytoene dehydrogenase gene of Phycomyces: Regulation of its expression by blue light and vitamin A. Mol. Gen. Gent. 1997, 253, 734–744. [Google Scholar] [CrossRef]
- Hausmann, A.; Sandmann, G. A single five-step desaturase is involved in the carotenoid biosynthesis pathway to γ-carotene and torulene in Neurospora crassa. Fungal Genet. Biol. 2000, 30, 147–153. [Google Scholar] [CrossRef]
- Aasen, A.J.; Jensen, S.L. Fungal carotenoids II. The structure of the carotenoid acid neurosporaxanthin. Acta Chem. Scand. 1965, 19, 1843–1853. [Google Scholar] [CrossRef]
- Goldie, A.H.; Subden, R.E. The neutral carotenoids of wild-type and mutant strains of Neurospora crassa. Biochem. Genet. 1973, 10, 275–284. [Google Scholar] [CrossRef]
- Sandmann, G.; Takaichi, S.; Fraser, P.D. Novel C35-apocarotenoids in the yellow mutant Neurospora crassa YLO. Phytochemistry 2008, 69, 2886–2890. [Google Scholar] [CrossRef]
- Estrada, A.F.; Youssar, L.; Scherzinger, D.; Al-Babili, S.; Avalos, J. The ylo-1 gene encodes an aldehyde dehydrogenase responsible for the last reaction in the Neurospora carotenoid pathway. Mol. Microbiol. 2008, 69, 1207–1220. [Google Scholar] [CrossRef]
- Saelices, L.; Youssar, L.; Holdermann, I.; Al-Babili, S.; Avalos, J. Identification of the gene responsible for torulene cleavage in the Neurospora carotenoid pathway. Mol. Genet. Genom. 2007, 278, 527–537. [Google Scholar] [CrossRef] [PubMed]
- Prado-Cabrero, A.; Estrada, A.F.; Al-Babili, S.; Avalos, J. Identification and biochemical characterization of a novel carotenoid oxygenase: Elucidation of the cleavage step in the Fusarium carotenoid pathway. Mol. Microbiol. 2007, 64, 448–460. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Sánchez, V.; Estrada, A.F.; Trautmann, D.; Al-Babili, S.; Avalos, J. The gene carD encodes the aldehyde dehydrogenase responsible for neurosporaxanthin biosynthesis in Fusarium fujikuroi. FEBS J. 2011, 278, 3164–3176. [Google Scholar] [CrossRef]
- Jin, J.-M.; Lee, J.; Lee, Y.-W. Characterization of carotenoid biosynthetic genes in the ascomycete Gibberella zeae. FEMS Microbiol. Lett. 2010, 302, 197–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strobel, I.; Breitenbach, J.; Scheckhuber, C.Q.; Osiewacz, H.D.; Sandmann, G. Carotenoids and carotenogenic genes in Podospora anserina: Engineering of the carotenoid composition extends the life span of the mycelium. Curr. Genet. 2009, 55, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Andrewes, A.G.; Phaff, H.J.; Starr, M.P. Carotenoids of Phaffia rhodozyma, a red-pigmented fermenting yeast. Phytochemistry 1976, 15, 1003–1007. [Google Scholar] [CrossRef]
- Andrewes, A.G.; Starr, M.P. (3R,30R)-Astaxanthin from the yeast Phaffia rhodozyma. Phytochemistry 1976, 15, 1009–1011. [Google Scholar] [CrossRef]
- Ojima, K.; Breitenbach, J.; Visser, H.; Setoguchi, Y.; Tabata, K.; Hoshino, T.; van den Berg, J.; Sandmann, G. Cloning of the astaxanthin synthase gene from Xanthophyllomyces dendrorhous (Phaffia rhodozyma) and its assignment as a β-carotene 3-hydroxylase/4-ketolase. Mol. Genet. Genom. 2006, 275, 148–158. [Google Scholar] [CrossRef]
- Alvarez, V.; Rodríguez-Sáiz, M.; de la Fuente, J.L.; Gudiña, E.J.; Godio, R.P.; Martín, J.F.; Barredo, J.L. The crtS gene of Xanthophyllomyces dendrorhous encodes a novel cytochrome-P450 hydroxylase involved in the conversion of β-carotene into astaxanthin and other xanthophylls. Fungal Genet. Biol. 2006, 43, 261–272. [Google Scholar] [CrossRef]
- Alcaíno, J.; Barahona, S.; Carmona, M.; Lozano, C.; Marcoleta, A.; Niklitschek, M.; Sepúlveda, D.; Baeza, M.; Cifuentes, V. Cloning of the cytochrome p450 reductase (crtR) gene and its involvement in the astaxanthin biosynthesis of Xanthophyllomyces dendrorhous. BMC Microbiol. 2008, 8, 169. [Google Scholar] [CrossRef] [Green Version]
- Verdoes, J.C.; Sandmann, G.; Visser, H.; Diaz, M.; van Mossel, M.; van Ooyen, A.J.J. Metabolic engineering of the carotenoid biosynthetic pathway in the yeast Xanthophyllomyces dendrorhous (Phaffia rhodozyma). Appl.Environ. Microbiol. 2003, 69, 3728–3738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vazquez, M.; Santos, V. 3-Hydroxy-3’, 4’-didehydro-beta-ψ-caroten-4-one (HDCO) from Xanthophyllomyces dendrorhous (Phaffia rhodozyma) cultivated on xylose media. Biotechnol. Lett. 1998, 20, 181–182. [Google Scholar] [CrossRef]
- Schrantz, J.P.; Lemoine, Y. Carotenoid composition of mycelium and apothecia in the discomycete Scutellinia umbrarum. Phytochemistry 1995, 40, 33–35. [Google Scholar] [CrossRef]
- Madhour, A.; Anke, H.; Mucci, A.; Davoli, P.; Weber, R.W.S. Biosynthesis of the xanthophyll plectaniaxanthin as a stress response in the red yeast Dioszegia (Tremellales, Heterobasidiomycetes, Fungi). Phytochemistry 2005, 66, 2617–2626. [Google Scholar] [CrossRef] [PubMed]
- Davoli, P.; Mierau, V.; Weber, R.W.S. Carotenoids and fatty acids in red yeasts Sporobolomyces roseus and Rhodotorula glutinis. Appl. Biochem. Microbiol. 2004, 40, 392–397. [Google Scholar] [CrossRef]
- Valadon, L.R.G. Carotenoid pigments of Protomyces inundates, Dangerard. Phytochemistry 1963, 2, 71–73. [Google Scholar] [CrossRef]
- Lederer, E. Sur les carotenoides des cryptogames. Bull. Soc. Chim. Fr. 1938, 20, 611–634. [Google Scholar]
- Smits, B.L.; Peterson, W.J. Carotenoids of telia galls of Gymnosporangium juniperi-virginianae LK. Science 1942, 96, 210–211. [Google Scholar] [CrossRef]
- Haxo, F. Carotenoids of the mushroom Cantharellus cinnabarinus. Bot. Gaz. 1950, 112, 228–232. [Google Scholar] [CrossRef]
- Arpin, N.; Kjφsen, H.; Francis, G.W.; Liaaen-Jensen, S. The structure of aleuriaxanthin. Phytochemistry 1973, 12, 2751–2758. [Google Scholar] [CrossRef]
- Arpin, N.; Liaaen-Jensen, S. Recherches chimotaxinomiques sur les champignons. Fungal carotenoids. III-Nouveaux carotenoides, notamment sous forme d’esters tertiaires, isoles de Plectania coccinea (Scop. Ex Fr) Fuck. Phytochemistry 1967, 6, 995–1005. [Google Scholar] [CrossRef]
- Molnár, P.; Ősz, E.; Turcsi, E.; Delia, J. Carotenoid composition of the mushroom Scarlet elf cup (Sarcoscypha coccinea). Heliyon 2019, 5, e01883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, M.; Lee, T.H.; Yokoyama, H.; Böttger, H.G.; Chichester, C.O. The occurrence of plectaniaxanthin in Cryptococcus laurentii. Phytochemistry 1971, 10, 625–629. [Google Scholar] [CrossRef]
- Arpin, N.; Liaaen-Jensen, S. Recherches chimiotaxinomiques sur le champignons. Sur la présence de l’ester méthylique de la torularhodine chez Cookeina sulcipes. C. R. Acad. Sci. Paris 1967, 265, 1083–1085. [Google Scholar]
- Herz, S.; Weber, R.W.S.; Anke, H.; Mucci, A.; Davoli, P. Intermediates in the oxidative pathway from torulene to torularhodin in the red yeasts Cystofilobasidium infirmominiatum and C. capitatum (Heterobasidiomycetes, Fungi). Phytochemistry 2007, 68, 2503–2511. [Google Scholar] [CrossRef]
- Fraser, P.D.; Miura, Y.; Misawa, N. In vitro characterization of astaxanthin biosynthetic enzymes. J. Biol. Chem. 1997, 272, 6128–6135. [Google Scholar] [CrossRef] [Green Version]
- Breitenbach, J.; Gerjets, T.; Sandmann, G. Catalytic properties and reaction mechanism of the CrtO carotenoid ketolase from the cyanobacterium Synechocystis sp. PCC 6803. Arch. Biochem. Biophys. 2013, 529, 86–91. [Google Scholar] [CrossRef]
- Takaichi, S.; Jung, D.O.; Madigan, M.T. Accumulation of unusual carotenoids in the spheroidene pathway, demethylspheroidene and demethylspheroidenone, in an alkaliphilic purple nonsulfur bacterium Rhodobaca bogoriensis. Photosynth. Res. 2001, 67, 207–214. [Google Scholar] [CrossRef]
- Steiger, S.; Mazet, A.; Sandmann, G. Heterologous expression, purification, and enzymatic characterization of the acyclic carotenoid 1,2-hydratase from Rubrivivax gelatinosus. Arch. Biochem. Biophys. 2003, 414, 51–58. [Google Scholar] [CrossRef]
- Albrecht, M.; Ruther, A.; Sandmann, G. Purification and biochemical characterization of a hydroxyneursporene desaturase involved in the biosynthetic pathway of the carotenoid spheroidene in Rhodobacter spheroides. J. Bacteriol. 1997, 179, 7462–7467. [Google Scholar] [CrossRef] [Green Version]
- Gerjets, T.; Steiger, S.; Sandmann, G. Catalytic properties of the expressed acyclic carotenoid 2-ketolases from Rhodobacter capsulatus and Rubrivivax gelatinosus. Biochim. Biophys. Acta. 2009, 1791, 125–131. [Google Scholar] [CrossRef] [PubMed]
- Graham, J.E.; Bryant, D.A. The biosynthetic pathway for myxol-2’ fucoside (myxoxanthophyll) in the cyanobacterium Synechococcus sp. strain PCC 7002. J. Bacteriol. 2009, 191, 3292–3300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steiger, S.; Perez-Fons, L.; Cutting, S.M.; Fraser, P.D.; Sandmann, G. Annotation and functional assignment of the genes for the C30 carotenoid pathways from the genomes of two bacteria Bacillus indicus and Bacillus firmus. Microbiology 2015, 161, 194–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandmann, G. Diversity and origin of carotenoid biosynthesis: Its history of coevolution towards plant photosynthesis. New Phytol. 2021, 232, 479–493. [Google Scholar] [CrossRef]
- Fitzpatrick, D.A. Horizontal gene transfer in fungi. FEMS Microbiol. Lett. 2012, 329, 1–8. [Google Scholar] [CrossRef]
- Gooday, D.W.; Carlile, M.J. The discovery of fungal sex hormones. III. Trisporic acid and its precorsors. Mcologist 1997, 11, 126–130. [Google Scholar] [CrossRef]
- Barreroa, A.F.; Herrador, M.M.; Arteaga, J.F.; González-Delgado, J.A. Occurrence and chemical synthesis of apocarotenoids from Mucorales: A review. Nat. Prod. Commun. 2017, 12, 733–741. [Google Scholar] [CrossRef] [Green Version]
- Sutter, R.P.; Capage, D.A.; Harrison, T.L.; Keen, W.A. Trisporic acid biosynthesis in separate plus and minus cultures of Blakeslea trispora: Identification by Mucor assay of two mating-type-specific components. J. Bacteriol. 1973, 114, 1074–1082. [Google Scholar] [CrossRef] [Green Version]
- Ahrazem, O.; Gómez-Gómez, L.; Rodrigo, M.R.; Avalos, J.; Limón, M.C. Carotenoid cleavage oxygenases from microbes and photosynthetic organisms: Features and functions. Int. J. Mol. Sci. 2016, 17, 1781. [Google Scholar] [CrossRef] [Green Version]
- Burmester, A.; Richter, M.; Schultze, K.; Voelz, K.; Schachtschabel, D.; Boland, W.; Wöstemeyer, J.; Schimek, C. Cleavage of β-carotene as the first step in sexual hormone synthesis in zygomycetes is mediated by a trisporic acid regulated β-carotene oxygenase. Fungal Genet. Biol. 2007, 44, 1096–1108. [Google Scholar] [CrossRef]
- Medina, H.R.; Cerdá-Olmedo, E.; Al-Babili, S. Cleavage oxygenases for the biosynthesis of trisporoids and other apocarotenoids in Phycomyces. Mol. Microbiol. 2011, 82, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Heitman, J. Sex in the Mucoralean fungi. Mycoses 2014, 57, 18–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wetzel, J.; Scheibner, O.; Burmester, A.; Schimek, C.; Wöstemeyer, J. 4-Dihydrotrisporin-dehydrogenase, an enzyme of the sex hormone pathway of Mucor mucedo: Purification, cloning of the corresponding gene, and developmental expression. Eukaryot. Cell 2009, 8, 88–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schachtschabel, D.; David, A.; Menzel, K.-D.; Schimek, C.; Wöstemeyer, J.; Boland, W. Cooperative biosynthesis of trisporoids by the (+) and (-) mating types of the zygomycete Blakeslea trispora. ChemBioChem 2008, 9, 3004–3012. [Google Scholar] [CrossRef] [PubMed]
- Czempinski, K.; Kruft, V.; Wöstemeyer, J.; Burmester, A. 4- Dihydromethyltrisporate dehydrogenase from Mucor mucedo, an enzyme of the sexual hormone pathway: Purification, and cloning of the corresponding gene. Microbiology 1966, 142, 2647–2654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Werkman, B. Localization and partial characterization of a sex-specific enzyme in homothallic and heterothallic Mucorales. Arch. Microbiol. 1976, 109, 209–213. [Google Scholar] [CrossRef]
- Sandmann, G. Carotenoids of biotechnological importance. In Biotechnology of Isoprenoids, Advances in Biochemical Engineering/Biotechnology; Schrader, J., Bohlmann, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; Volume 148, pp. 449–467. [Google Scholar]
- Kanamoto, H.; Nakamura, K.; Misawa, N. Carotenoid production in oleaginous yeasts. Adv. Exp. Med. Biol. 2021, 1261, 153–163. [Google Scholar]
- Miura, Y.; Kondo, K.; Saito, T.; Shimada, H.; Fraser, P.D.; Misawa, N. Production of the carotenoids lycopene, ß-carotene, and astaxanthin in the food yeast Candida utilis. Appl. Environ. Microbiol. 1998, 64, 1226–1229. [Google Scholar] [CrossRef] [Green Version]
- Shimada, H.; Kondo, K.; Fraser, P.D.; Miura, Y.; Saito, T.; Misawa, N. Increased carotenoid production by the food yeast Candida utilis through metabolic engineering of the isoprenoid pathway. Appl. Environ. Microbiol. 1998, 64, 2676–2680. [Google Scholar] [CrossRef] [Green Version]
- Bhataya, A.; Schmidt-Dannert, C.; Lee, P.C. Metabolic engineering of Pichia pastoris X-33 for lycopene production. Process. Biochem. 2009, 44, 1095–1102. [Google Scholar] [CrossRef]
- Araya-Garay, J.M.; Ageitos, J.M.; Vallejo, J.A.; Veiga-Crespo, P.; Sánchez-Pérez, A.; Villa, T.G. Construction of a novel Pichia pastoris strain for production of xanthophylls. AMB Express 2012, 2, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schwartz, C.; Shabbir-Hussain, M.; Frogue, K.; Blenner, M.; Wheeldon, I. Standardized markerless gene integration for pathway engineering in Yarrowia lipolytica. ACS Synth. Biol. 2017, 6, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Qu, Y.L.; Dong, G.R.; Wang, J.; Hu, C.Y.; Meng, Y.H. Elevated β-carotene production using codon-adapted CarRA&B and metabolic balance in engineered Yarrowia lipolytica. Front. Microbiol. 2021, 12, 627150. [Google Scholar] [PubMed]
- Xie, Y.; Chen, S.; Xiong, X. Metabolic engineering of non-carotenoid-producing yeast Yarrowia lipolytica for the biosynthesis of zeaxanthin. Front. Microbiol. 2021, 12, 699235. [Google Scholar] [CrossRef] [PubMed]
- Tramontin, L.R.R.; Kildegaard, K.R.; Sudarsan, S.; Borodina, I. Enhancement of astaxanthin biosynthesis in oleaginous yeast Yarrowia lipolytica via microalgal pathway. Microorganisms 2019, 7, 472. [Google Scholar] [CrossRef] [Green Version]
- Pi, H.-W.; Anandharaj, M.; Kao, Y.-Y.; Lin, Y.-J.; Chang, J.-J.; Li, W.-H. Engineering the oleaginous red yeast Rhodotorula glutinis for simultaneous β-carotene and cellulase production. Sci. Rep. 2018, 8, 10850. [Google Scholar] [CrossRef]
- Gassel, S.; Breitenbach, J.; Sandmann, G. Genetic engineering of the complete carotenoid pathway towards enhanced astaxanthin formation in Xanthophyllomyces dendrorhous starting from a high-yield mutant. Appl. Microbiol. Biotechnol. 2014, 98, 345–350. [Google Scholar] [CrossRef]
- Pollmann, H.; Breitenbach, J.; Sandmann, G. Development of Xanthophyllomyces dendrorhous as a production system for the colorless carotene phytoene. J. Biotechnol. 2017, 247, 34–41. [Google Scholar] [CrossRef]
- Breitenbach, J.; Pollmann, H.; Sandmann, G. Genetic modification of the carotenoid pathway in the red yeast Xanthophyllomyces dendrorhous: Engineering of a high-yield zeaxanthin strain. J. Biotechnol. 2019, 289, 112–117. [Google Scholar] [CrossRef]
- Salgado, L.; Bejarano, E.R.; Cerda-Olmedo, W. Carotene-superproducing mutants of Phycomyces blakesleeanus. Exp. Mycol. 1989, 13, 332–336. [Google Scholar] [CrossRef]
- Avalos, J.; Cerdá-Olmedo, E. Carotenoid mutants of Gibberella fujikuroi. Curr. Genet. 1987, 25, 1837–1841. [Google Scholar] [CrossRef]
- Schmidt, I.; Schewe, H.; Gassel, S.; Jin, C.; Buckingham, J.; Hümbelin, M.; Sandmann, G.; Schrader, J. Biotechnological production of astaxanthin with Phaffia rhodozyma/Xanthophyllomyces dendrorhous. Appl. Microbiol. Biotechnol. 2010, 8, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Gassel, S.; Schewe, H.; Schmidt, I.; Schrader, J.; Sandmann, G. Multiple improvement of astaxanthin biosynthesis in Xanthophyllomyces dendrorhous by a combination of conventional mutagenesis and metabolic pathway engineering. Biotechnol. Lett. 2013, 35, 565–569. [Google Scholar] [CrossRef] [PubMed]
- Mehta, B.J.; Obraztsova, I.N.; Cerdá-Olmedo, E. Mutants and intersexual heterokaryons of Blakeslea trispora for production of β-carotene and lycopene. Appl. Environ. Microbiol. 2003, 69, 4043–4048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mawgoud, A.M.; Markham, K.A.; Palmer, C.M.; Liu, N.; Stephanopoulos, G.; Alper, H.S. Metabolic engineering in the host Yarrowia lipolytica. Metab. Eng. 2018, 50, 192–208. [Google Scholar] [CrossRef]
- Sandmann, G.; Pollmann, H.; Gassel, S.; Breitenbach, J. Xanthophyllomyces dendrorhous, a versatile platform for the production of carotenoids and other acetyl-CoA-derived compounds. J. Adv. Exp. Med. Biol. 2021, 1261, 137–151. [Google Scholar]
- Pollmann, H.; Breitenbach, J.; Wolff, H.; Bode, H.B.; Sandmann, G. Combinatorial biosynthesis of novel multi-hydroxy carotenoids in the red yeast Xanthophyllomyces dendrorhous. J. Fungi 2017, 3, 9. [Google Scholar] [CrossRef]
- Larroude, M.; Celinska, E.; Back, A.; Thomas, S.; Nicaud, J.-M.; Ledesma-Amaro, R. A synthetic biology approach to transform Yarrowia lipolytica into a competitive biotechnological producer of β-carotene. Biotechnol. Bioeng. 2017, 115, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Matthäus, F.; Ketelhot, M.; Gatter, M.; Barth, G. Production of lycopene in the non-carotenoid-producing yeast Yarrowia lipolytica. Appl. Environ. Microbiol. 2014, 80, 1660–1669. [Google Scholar] [CrossRef] [Green Version]
- Spagnuolo, M.; Hussain, M.S.; Gambill, L.; Blenner, M. Alternative substrate metabolism in Yarrowia lipolytica. Front. Microbiol. 2018, 9, 1077. [Google Scholar] [CrossRef]
- Li, C.; Swofford, C.A.; Sinskey, A.J. Modular engineering for microbial production of carotenoids. Metab. Eng. Commun. 2020, 10, e00118. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.; Ma, T.; Ye, Z.; Li, X.; Huang, Y.; Zhou, Z.; Ding, Y.; Deng, Z.; Liu, T. Systematic metabolic engineering of Saccharomyces cerevisiae for lycopene overproduction. J. Agric. Food Chem. 2019, 67, 11148–11157. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.; Shi, S.; Ye, Z.; Li, X.; Liu, M.; Chen, Y.; Xia, J.; Nielsen, J.; Deng, Z.; Liu, T. Lipid engineering combined with systematic metabolic engineering of Saccharomyces cerevisiae for high-yield production of lycopene. Metab. Eng. 2019, 52, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Xiao, W.; Wang, Y.; Liu, H.; Li, X.; Yuan, Y. Lycopene overproduction in Saccharomyces cerevisiae through combining pathway engineering with host engineering. Microb. Cell Fact. 2016, 15, 113. [Google Scholar] [CrossRef] [Green Version]
- Verwaal, R.; Wang, J.; Meijnen, J.-P.; Visser, H.; Sandmann, G.; van den Berg, J.A.; van Ooyen, J.J. High-level production of beta-carotene in Saccharomyces cerevisiae by successive transformation with carotenogenic genes from Xanthophyllomyces dendrorhous. Appl. Environ. Microbiol. 2007, 73, 4342–4350. [Google Scholar] [CrossRef] [Green Version]
- Godara, A.; Gomez-Rodriguez, M.A.; Weatherston, J.D.; Peabody, G.L.; Wu, H.-J.; Kao, K.C. Beneficial mutations for carotenoid production identified from laboratory-evolved Saccharomyces cerevisiae. J. Ind. Microbiol. Biotechnol. 2019, 46, 1793–1804. [Google Scholar] [CrossRef]
- Fathi, Z.; Tramontin, L.R.R.; Ebrahimipour, G.; Borodina, I.; Darvishi, F. Metabolic engineering of Saccharomyces cerevisiae for production of β-carotene from hydrophobic substrates. FEMS Yeast Res. 2021, 21, foaa068. [Google Scholar] [CrossRef]
- Jin, J.; Wang, Y.; Yao, M.; Gu, X.; Li, B.; Liu, H.; Ding, M.; Xiao, W.; Yuan, Y. Astaxanthin overproduction in yeast by strain engineering and new gene target uncovering. Biotechnol. Biofuels 2018, 11, 230. [Google Scholar] [CrossRef]
- Jin, E.; Feth, B.; Melis, A. A mutant of the green alga Dunaliella salina constitutively accumulates zeaxanthin under all growth conditions. Biotechnol. Bioeng. Symp. 2003, 81, 115–124. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Carotenoids | Selected Species | References |
|---|---|---|
| Lycopene ---> γ-Carotene | Chytridiomycota: | [2] |
| Cladochytrium replicatum | ||
| Blastocladiomycota | ||
| Allomyces arbuscula | ||
| Lycopene ---> β-Carotene | Mucoromycotina: | [1] |
| Blakeslea trispora | ||
| Phycomyces blakesleanus | ||
| Ascomycota: | [48] | |
| Protomyces inundates | ||
| Basidiomycota: | [49] | |
| Tremella mesenterica | [50] | |
| Gymnosporangium juniperi-virginianae | ||
| Lycopene ---> Phillipsiaxanthin | Ascomycota: | [2] |
| Phillipsia carminea | ||
| β-Carotene ---> Canthaxanthin | Basidiomycota: | [51] |
| Cantharellus species | ||
| β-Carotene ---> Astaxanthin | Basidiomycota: | [38] |
| Xanthophyllomyces dendrorhous | ||
| γ-Carotene ---> Aleuriaxanthin | Ascomycota: | |
| Aleuria aurantiaca | [52] | |
| Scutellina umbrarum | [45] | |
| γ-Carotene and/or torulene ---> Neurosporaxanthin | Ascomycota: | |
| Fusarium species | [23] | |
| Neurospora crassa | [30] | |
| γ-Carotene and/or torulene ---> Plectaniaxanthin and 2′-didehyroplectaniaxanthin | Ascomycota: | |
| Plectania coccinea | [53] | |
| Sarcoscypha coccinea | [54] | |
| Basidiomycota: | ||
| Cryptococcus laurentii | [55] | |
| Dioszegia species | [46] | |
| Torulene ---> Torularhodin | Ascomycota: | [56] |
| Cookeina sulcipes | ||
| Basidiomycota: | [57] | |
| Cystofilobasidium species | [47] | |
| Rhodotorula glutinis |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandmann, G. Carotenoids and Their Biosynthesis in Fungi. Molecules 2022, 27, 1431. https://doi.org/10.3390/molecules27041431
Sandmann G. Carotenoids and Their Biosynthesis in Fungi. Molecules. 2022; 27(4):1431. https://doi.org/10.3390/molecules27041431
Chicago/Turabian StyleSandmann, Gerhard. 2022. "Carotenoids and Their Biosynthesis in Fungi" Molecules 27, no. 4: 1431. https://doi.org/10.3390/molecules27041431
APA StyleSandmann, G. (2022). Carotenoids and Their Biosynthesis in Fungi. Molecules, 27(4), 1431. https://doi.org/10.3390/molecules27041431