
Production of Large-Ring Cyclodextrins by Amylomaltases


Abstract
:1. Introduction
2. Sources and Biochemical Properties of Amylomaltase
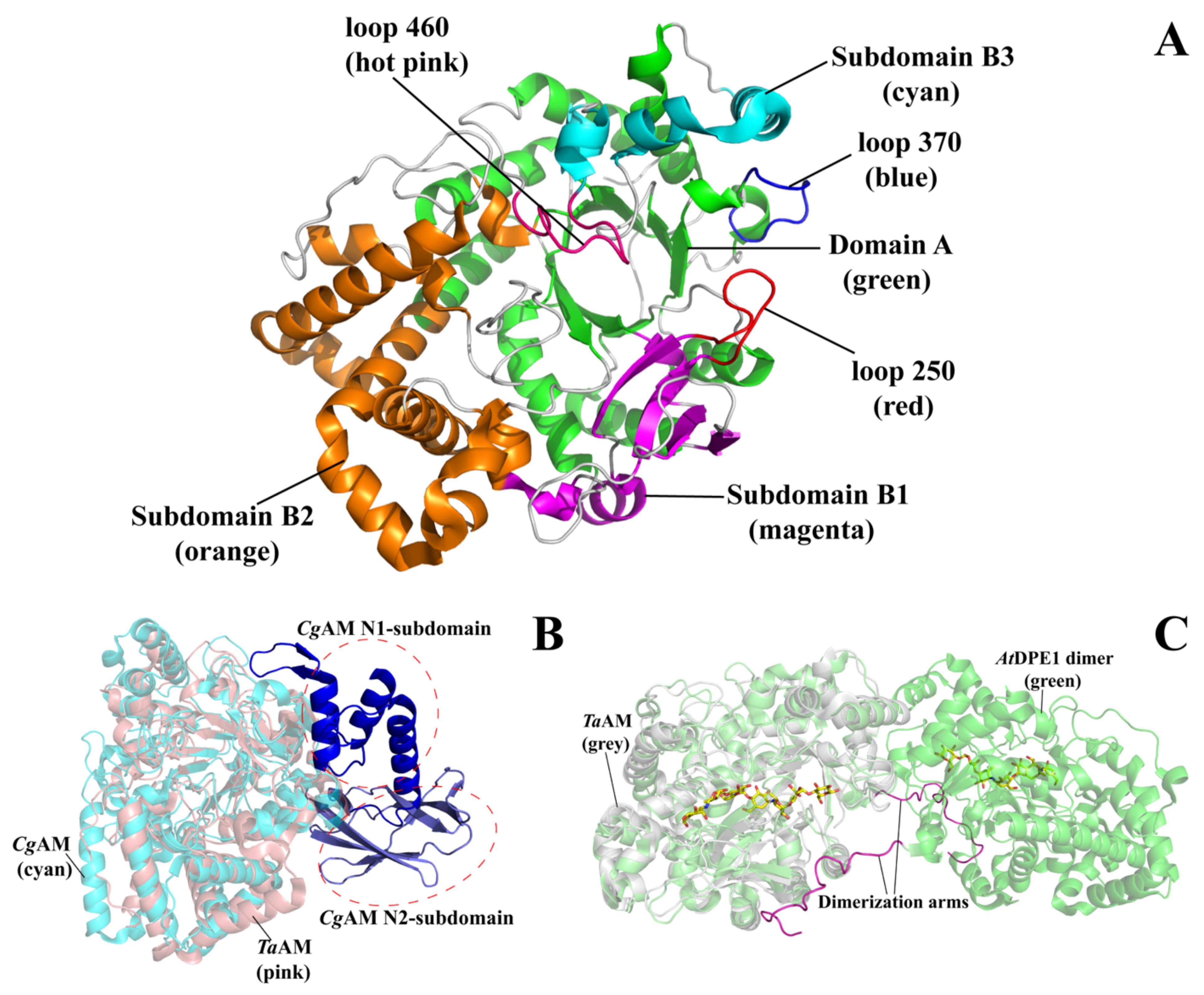
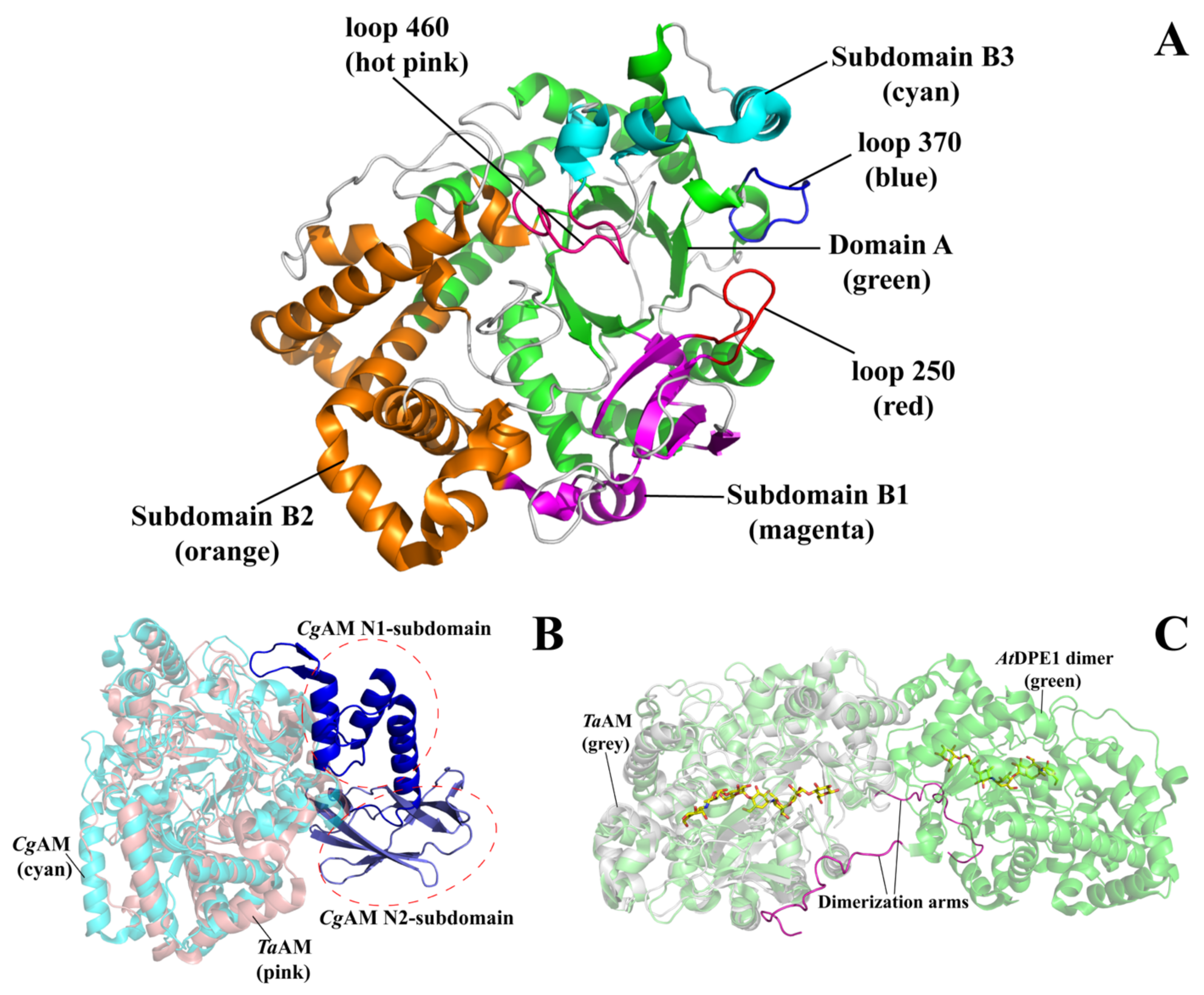
3. Overall Structure of Amylomaltase
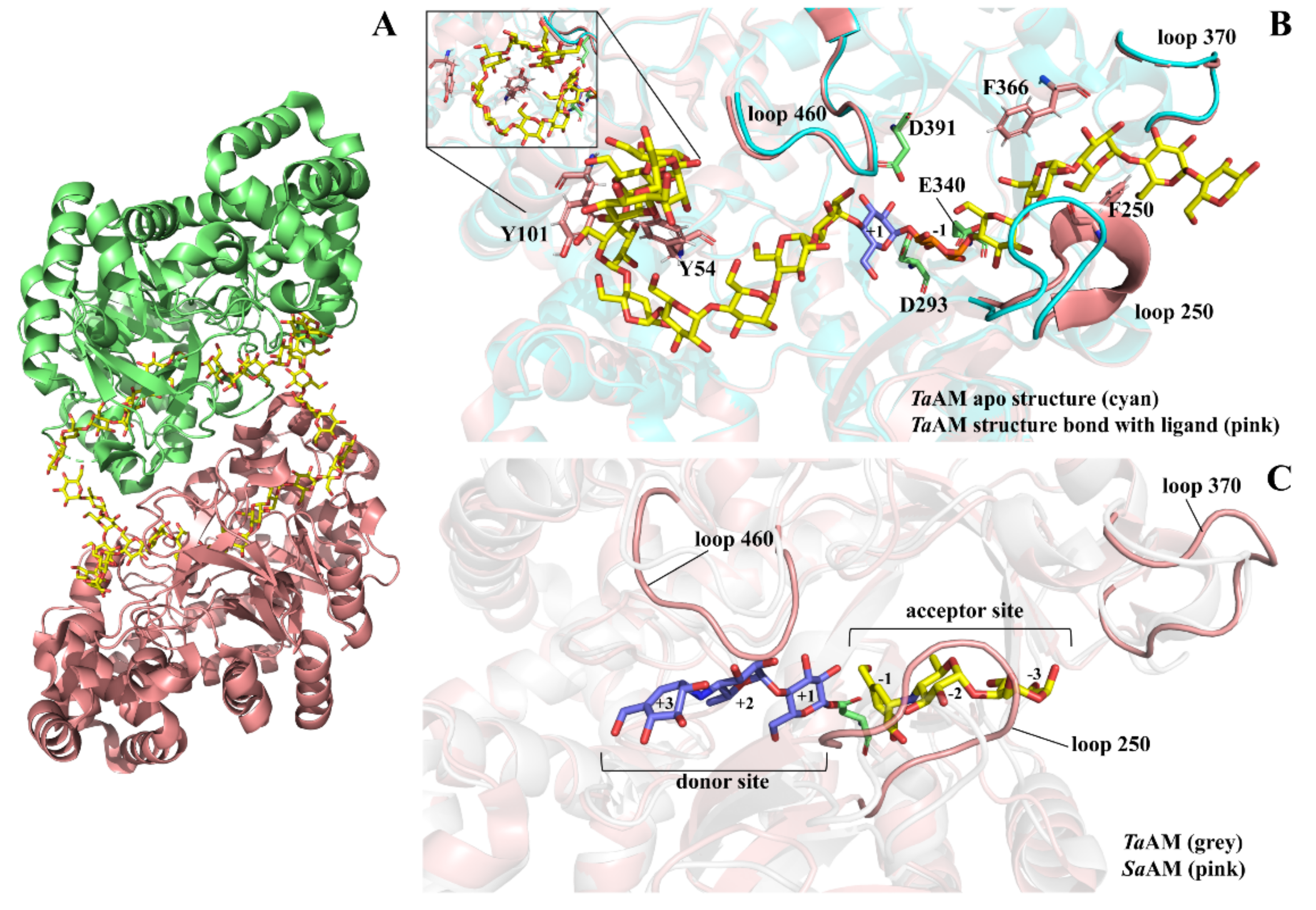
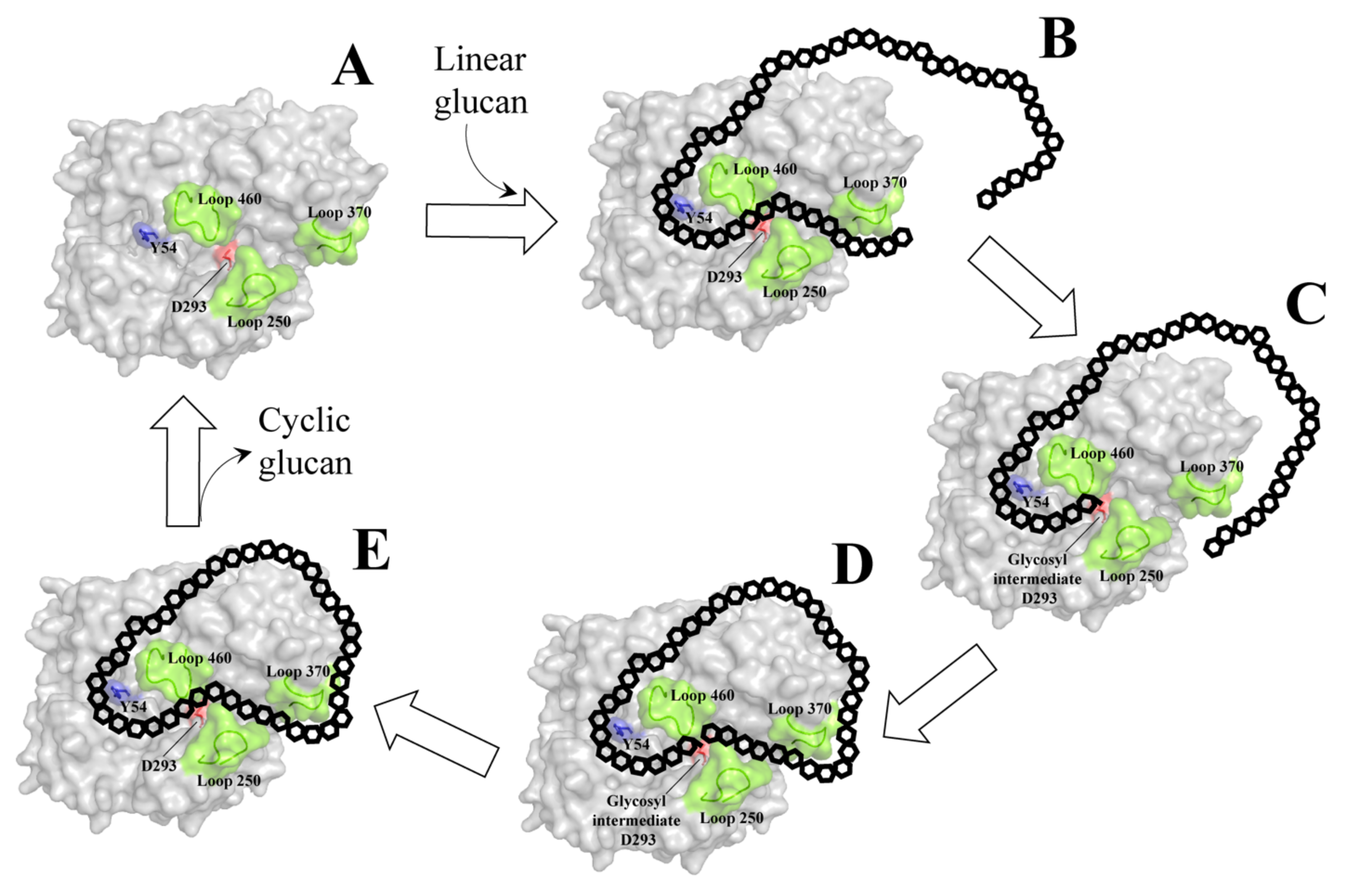
4. Cyclization Mechanism of Amylomaltase
5. Large-Ring Cyclodextrin Production by Amylomaltase
6. Applications of Large-Ring Cyclodextrins
7. Other Applications of Amylomaltase
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lombard, V.; Golaconda Ramulu, H.; Drula, E.; Coutinho, P.M.; Henrissat, B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 2014, 42, D490–D495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulley, A.O.; French, D. Studies on the Schardinger dextrins. XI. The isolation of new Schardinger dextrins. Biochem. Biophys Res. Commun. 1961, 5, 11–15. [Google Scholar] [CrossRef]
- Endo, T.; Zheng, M.; Zimmermann, W. Enzymatic synthesis and analysis of large-ring cyclodextrins. Aust. J. Chem. 2002, 55, 39–48. [Google Scholar]
- Terada, Y.; Sanbe, H.; Takaha, T.; Kitahata, S.; Koizumi, K.; Okada, S. Comparative study of the cyclization reactions of three bacterial cyclomaltodextrin glucanotransferases. Appl. Environ. Microbiol. 2001, 67, 1453–1460. [Google Scholar] [CrossRef] [Green Version]
- Sonnendecker, C.; Melzer, S.; Zimmermann, W. Engineered cyclodextrin glucanotransferases from Bacillus sp. G-825-6 produce largering cyclodextrins with high specificity. Microbiologyopen 2019, 8, e757. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.; Zhang, Y.; Niu, Y.; Ke, Q.; Kou, X. Cyclodextrins as carriers for volatile aroma compounds: A review. Carbohydr. Polym. 2021, 269, 118292. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sameen, D.E.; Ahmed, S.; Wang, Y.; Lu, R.; Dai, J.; Li, S.; Qin, W. Recent advances in cyclodextrin-based films for food packaging. Food Chem. 2022, 370, 131026. [Google Scholar] [CrossRef]
- Aiassa, V.; Garnero, C.; Longhi, M.R.; Zoppi, A. Cyclodextrin multicomponent complexes: Pharmaceutical applications. Pharmaceutics 2021, 13, 1099. [Google Scholar] [CrossRef]
- Perinelli, D.R.; Palmieri, G.F.; Cespi, M.; Bonacucina, G. Encapsulation of flavours and fragrances into polymeric capsules and cyclodextrins inclusion complexes: An update. Molecules 2020, 25, 5878. [Google Scholar] [CrossRef]
- Nakapong, S.; Tumhom, S.; Kaulpiboon, J.; Pongsawasdi, P. Heterologous expression of 4α-glucanotransferase: Overproduction and properties for industrial applications. World J. Microbiol. Biotechnol. 2022, 38, 36. [Google Scholar] [CrossRef]
- Leoni, C.; Gattulli, B.A.R.; Pesole, G.; Ceci, L.R.; Volpicella, M. Amylomaltases in extremophilic microorganisms. Biomolecules 2021, 11, 1335. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, N.; Mehboob, S.; Rashid, N. Starch-processing enzymes—Emphasis on thermostable 4-α-glucanotransferases. Biologia 2015, 70, 709–725. [Google Scholar] [CrossRef]
- Takaha, T.; Yanase, M.; Takata, H.; Okada, S.; Smith, S.M. Potato D-enzyme catalyzes the cyclization of amylose to produce cycloamylose, a novel cyclic glucan. J. Biol. Chem. 1996, 271, 2902–2908. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.E.; Tran, P.L.; Ko, J.M.; Kim, S.R.; Kim, J.H.; Park, J.T. Comparison of catalyzing properties of bacterial 4-α-glucanotransferases focusing on their cyclizing activity. J. Microbiol. Biotechnol. 2021, 31, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Boos, W.; Shuman, H. Maltose/maltodextrin system of Escherichia coli: Transport, metabolism, and regulation. Microbiol. Mol. Biol. Rev. 1998, 62, 204–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.T.; Shim, J.H.; Tran, P.L.; Hong, I.H.; Yong, H.U.; Oktavina, E.F.; Nguyen, H.D.; Kim, J.W.; Lee, T.S.; Park, S.H.; et al. Role of maltose enzymes in glycogen synthesis by Escherichia coli. J. Bacteriol. 2011, 193, 2517–2526. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, D.H.D.; Park, S.H.; Tran, P.L.; Kim, J.W.; Le, Q.T.; Boos, W.; Park, J.T. Characterization of the transglycosylation reaction of 4-α-glucanotransferase (MalQ) and its role in glycogen breakdown in Escherichia coli. J. Microbiol. Biotechnol. 2019, 29, 357–366. [Google Scholar] [CrossRef]
- Leoni, C.; Manzari, C.; Tran, H.; Golyshin, P.N.; Pesole, G.; Volpicella, M.; Ceci, L.R. Identification of an amylomaltase from the halophilic archaeon Haloquadratum walsbyi by functional metagenomics: Structural and functional insights. Life 2022, 12, 85. [Google Scholar] [CrossRef]
- Takaha, T.; Yanase, M.; Okada, S.; Smith, S.M. Disproportionating enzyme (4-α-glucanotransferase; EC 2.4.1.25) of potato. Purification, molecular cloning, and potential role in starch metabolism. J. Biol. Chem. 1993, 268, 1391–1396. [Google Scholar] [CrossRef]
- Lutken, H.; Lloyd, J.R.; Glaring, M.A.; Baunsgaard, L.; Laursen, K.H.; Haldrup, A.; Kossmann, J.; Blennow, A. Repression of both isoforms of disproportionating enzyme leads to higher malto-oligosaccharide content and reduced growth in potato. Planta 2010, 232, 1127–1139. [Google Scholar] [CrossRef]
- Zeeman, S.C.; Smith, S.M.; Smith, A.M. The breakdown of starch in leaves. New Phytol. 2004, 163, 247–261. [Google Scholar] [CrossRef] [PubMed]
- Tafazoli, S.; Wong, A.W.; Akiyama, T.; Kajiura, H.; Tomioka, E.; Kojima, I.; Takata, H.; Kuriki, T. Safety evaluation of amylomaltase from Thermus aquaticus. Regul. Toxicol. Pharmacol. 2010, 57, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Kaper, T.; Talik, B.; Ettema, T.J.; Bos, H.; van der Maarel, M.J.; Dijkhuizen, L. Amylomaltase of Pyrobaculum aerophilum IM2 produces thermoreversible starch gels. Appl. Environ. Microbiol. 2005, 71, 5098–5106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehboob, S.; Ahmad, N.; Munir, S.; Ali, R.; Younas, H.; Rashid, N. Gene cloning, expression enhancement in Escherichia coli and biochemical characterization of a highly thermostable amylomaltase from Pyrobaculum calidifontis. Int. J. Biol. Macromol. 2020, 165, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Bhuiyan, S.H.; Kitaoka, M.; Hayashi, K. A cycloamylose-forming hyperthermostable 4-α-glucanotransferase of Aquifex aeolicus expressed in Escherichia coli. J. Mol. Catal. B Enzym. 2003, 22, 45–53. [Google Scholar] [CrossRef]
- Terada, Y.; Fujii, K.; Takaha, T.; Okada, S. Thermus aquaticus ATCC 33923 amylomaltase gene cloning and expression and enzyme characterization: Production of cycloamylose. Appl. Environ. Microbiol. 1999, 65, 910–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaewpathomsri, P.; Takahashi, Y.; Nakamura, S.; Kaulpiboon, J.; Kidokoro, S.-i.; Murakami, S.; Krusong, K.; Pongsawasdi, P. Characterization of amylomaltase from Thermus filiformis and the increase in alkaline and thermo-stability by E27R substitution. Process Biochem. 2015, 50, 1814–1824. [Google Scholar] [CrossRef]
- Bang, B.Y.; Kim, H.J.; Kim, H.Y.; Baik, M.Y.; Ahn, S.C.; Kim, C.H.; Park, C.S. Cloning and overexpression of 4-α-glucanotransferase from Thermus brockianus (TBGT) in E. coli. J. Microbiol. Biotechnol. 2006, 16, 1809–1813. [Google Scholar]
- van der Maarel, M.J.E.C.; Capron, I.; Euverink, G.-J.W.; Bos, H.T.; Kaper, T.; Binnema, D.J.; Steeneken, P.A.M. A novel thermoreversible gelling product made by enzymatic modification of starch. Starch Stärke 2005, 57, 465–472. [Google Scholar] [CrossRef]
- Seo, N.S.; Roh, S.A.; Auh, J.H.; Park, J.H.; Kim, Y.R.; Park, K.H. Structural characterization of rice starch in rice cake modified by Thermus scotoductus 4-α-glucanotransferase (TS alpha GTase). J. Food Sci. 2007, 72, C331–C336. [Google Scholar] [CrossRef]
- Tumhom, S.; Nimpiboon, P.; Wangkanont, K.; Pongsawasdi, P. Streptococcus agalactiae amylomaltase offers insight into the transglycosylation mechanism and the molecular basis of thermostability among amylomaltases. Sci. Rep. 2021, 11, 6740. [Google Scholar] [CrossRef] [PubMed]
- Nimpiboon, P.; Kaulpiboon, J.; Krusong, K.; Nakamura, S.; Kidokoro, S.; Pongsawasdi, P. Mutagenesis for improvement of activity and thermostability of amylomaltase from Corynebacterium glutamicum. Int. J. Biol. Macromol. 2016, 86, 820–828. [Google Scholar] [CrossRef] [PubMed]
- Srisimarat, W.; Kaulpiboon, J.; Krusong, K.; Zimmermann, W.; Pongsawasdi, P. Altered large-ring cyclodextrin product profile due to a mutation at Tyr-172 in the amylomaltase of Corynebacterium glutamicum. Appl. Environ. Microbiol. 2012, 78, 7223–7228. [Google Scholar] [CrossRef] [PubMed]
- Godany, A.; Vidova, B.; Janecek, S. The unique glycoside hydrolase family 77 amylomaltase from Borrelia burgdorferi with only catalytic triad conserved. FEMS Microbiol. Lett. 2008, 284, 84–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nimpiboon, P.; Krusong, K.; Kaulpiboon, J.; Kidokoro, S.; Pongsawasdi, P. Roles of N287 in catalysis and product formation of amylomaltase from Corynebacterium glutamicum. Biochem. Biophys. Res. Commun. 2016, 478, 759–764. [Google Scholar] [CrossRef] [PubMed]
- Srisimarat, W.; Powviriyakul, A.; Kaulpiboon, J.; Krusong, K.; Zimmermann, W.; Pongsawasdi, P. A novel amylomaltase from Corynebacterium glutamicum and analysis of the large-ring cyclodextrin products. J. Incl. Phenom. Macrocycl. Chem. 2010, 70, 369–375. [Google Scholar] [CrossRef]
- Jiang, H.; Miao, M.; Ye, F.; Jiang, B.; Zhang, T. Enzymatic modification of corn starch with 4-α-glucanotransferase results in increasing slow digestible and resistant starch. Int. J. Biol. Macromol. 2014, 65, 208–214. [Google Scholar] [CrossRef]
- Wiesmeyer, H.; Cohn, M. The characterization of the pathway of maltose utilization by Escherichia coli. II. General properties and mechanism of action of amylomaltase. Biochim. Biophys. Acta 1960, 39, 427–439. [Google Scholar] [CrossRef]
- Hwang, S.; Choi, K.H.; Kim, J.; Cha, J. Biochemical characterization of 4-α-glucanotransferase from Saccharophagus degradans 2-40 and its potential role in glycogen degradation. FEMS Microbiol. Lett. 2013, 344, 145–151. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.H.; Oh, D.K.; Yoo, S.H. Characterization of 4-alpha-glucanotransferase from Synechocystis sp. PCC 6803 and its application to various corn starches. N. Biotechnol. 2009, 26, 29–36. [Google Scholar] [CrossRef]
- Tantanarat, K.; O’Neill, E.C.; Rejzek, M.; Field, R.A.; Limpaseni, T. Expression and characterization of 4-α-glucanotransferase genes from Manihot esculenta Crantz and Arabidopsis thaliana and their use for the production of cycloamyloses. Process Biochem. 2014, 49, 84–89. [Google Scholar] [CrossRef]
- Ruzanski, C.; Smirnova, J.; Rejzek, M.; Cockburn, D.; Pedersen, H.L.; Pike, M.; Willats, W.G.; Svensson, B.; Steup, M.; Ebenhoh, O.; et al. A bacterial glucanotransferase can replace the complex maltose metabolism required for starch to sucrose conversion in leaves at night. J. Biol. Chem. 2013, 288, 28581–28598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akdogan, G.; Kubota, J.; Kubo, A.; Takaha, T.; Kitamura, S. Expression and characterization of rice disproportionating enzymes. J. Appl. Glycosci. 2011, 58, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Mareček, F.; Møller, M.S.; Svensson, B.; Janeček, Š. A putative novel starch-binding domain revealed by in silico analysis of the N-terminal domain in bacterial amylomaltases from the family GH77. 3 Biotech 2021, 11, 229. [Google Scholar] [CrossRef] [PubMed]
- Janeček, Š.; Martinovičová, M. New groups of protein homologues in the α-amylase family GH57 closely related to α-glucan branching enzymes and 4-α-glucanotransferases. Genetica 2020, 148, 77–86. [Google Scholar] [CrossRef]
- Imamura, H.; Fushinobu, S.; Yamamoto, M.; Kumasaka, T.; Jeon, B.-S.; Wakagi, T.; Matsuzawa, H. Crystal structures of 4-α-glucanotransferase from Thermococcus litoralis and its complex with an inhibitor. J. Biol. Chem. 2003, 278, 19378–19386. [Google Scholar] [CrossRef] [Green Version]
- Paul, C.J.; Leemhuis, H.; Dobruchowska, J.M.; Grey, C.; Önnby, L.; van Leeuwen, S.S.; Dijkhuizen, L.; Karlsson, E.N. A GH57 4-α-glucanotransferase of hyperthermophilic origin with potential for alkyl glycoside production. Appl. Microbiol. Biotechnol. 2015, 99, 7101–7113. [Google Scholar] [CrossRef]
- Przylas, I.; Tomoo, K.; Terada, Y.; Takaha, T.; Fujii, K.; Saenger, W.; Strater, N. Crystal structure of amylomaltase from Thermus aquaticus, a glycosyltransferase catalysing the production of large cyclic glucans. J. Mol. Biol. 2000, 296, 873–886. [Google Scholar] [CrossRef]
- Uitdehaag, J.C.; Kalk, K.H.; van der Veen, B.A.; Dijkhuizen, L.; Dijkstra, B.W. The cyclization mechanism of cyclodextrin glycosyltransferase (CGTase) as revealed by a γ-cyclodextrin-CGTase complex at 1.8-Å resolution. J. Biol. Chem. 1999, 274, 34868–34876. [Google Scholar] [CrossRef] [Green Version]
- Uitdehaag, J.C.; Van Alebeek, G.-J.W.; Van Der Veen, B.A.; Dijkhuizen, L.; Dijkstra, B.W. Structures of maltohexaose and maltoheptaose bound at the donor sites of cyclodextrin glycosyltransferase give insight into the mechanisms of transglycosylation activity and cyclodextrin size specificity. Biochemistry 2000, 39, 7772–7780. [Google Scholar] [CrossRef] [Green Version]
- Weiss, S.C.; Skerra, A.; Schiefner, A. Structural basis for the interconversion of maltodextrins by MalQ, the amylomaltase of Escherichia coli. J. Biol. Chem. 2015, 290, 21352–21364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joo, S.; Kim, S.; Seo, H.; Kim, K.J. Crystal structure of amylomaltase from Corynebacterium glutamicum. J. Agric. Food Chem. 2016, 64, 5662–5670. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, E.C.; Stevenson, C.E.; Tantanarat, K.; Latousakis, D.; Donaldson, M.I.; Rejzek, M.; Nepogodiev, S.A.; Limpaseni, T.; Field, R.A.; Lawson, D.M. Structural dissection of the maltodextrin disproportionation cycle of the Arabidopsis plastidial disproportionating enzyme 1 (DPE1). J. Biol. Chem. 2015, 290, 29834–29853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imamura, K.; Matsuura, T.; Nakagawa, A.; Kitamura, S.; Kusunoki, M.; Takaha, T.; Unno, H. Structural analysis and reaction mechanism of the disproportionating enzyme (D-enzyme) from potato. Protein. Sci. 2020, 29, 2085–2100. [Google Scholar] [CrossRef]
- Steichen, J.M.; Petty, R.V.; Sharkey, T.D. Domain characterization of a 4-α-glucanotransferase essential for maltose metabolism in photosynthetic leaves. J. Biol. Chem. 2008, 283, 20797–20804. [Google Scholar] [CrossRef] [Green Version]
- Kuchtová, A.; Janeček, Š. In silico analysis of family GH77 with focus on amylomaltases from borreliae and disproportionating enzymes DPE2 from plants and bacteria. Biochim. Biophys. Acta Proteins Proteom. 2015, 1854, 1260–1268. [Google Scholar] [CrossRef]
- Barends, T.R.; Bultema, J.B.; Kaper, T.; van der Maarel, M.J.; Dijkhuizen, L.; Dijkstra, B.W. Three-way stabilization of the covalent intermediate in amylomaltase, an alpha-amylase-like transglycosylase. J. Biol. Chem. 2007, 282, 17242–17249. [Google Scholar] [CrossRef] [Green Version]
- Roth, C.; Weizenmann, N.; Bexten, N.; Saenger, W.; Zimmermann, W.; Maier, T.; Strater, N. Amylose recognition and ring-size determination of amylomaltase. Sci. Adv. 2017, 3, e1601386. [Google Scholar] [CrossRef] [Green Version]
- Terada, Y.; Yanase, M.; Takata, H.; Takaha, T.; Okada, S. Cyclodextrins are not the major cyclic alpha-1,4-glucans produced by the initial action of cyclodextrin glucanotransferase on amylose. J. Biol. Chem. 1997, 272, 15729–15733. [Google Scholar] [CrossRef] [Green Version]
- Qi, Q.; Mokhtar, M.N.; Zimmermann, W. Effect of ethanol on the synthesis of large-ring cyclodextrins by cyclodextrin glucanotransferases. J. Incl. Phenom. Macrocycl. Chem. 2007, 57, 95–99. [Google Scholar] [CrossRef]
- Sonnendecker, C.; Thurmann, S.; Przybylski, C.; Zitzmann, F.D.; Heinke, N.; Krauke, Y.; Monks, K.; Robitzki, A.A.; Belder, D.; Zimmermann, W. Large-ring cyclodextrins as chiral selectors for enantiomeric pharmaceuticals. Angew. Chem. 2019, 58, 6411–6414. [Google Scholar] [CrossRef] [PubMed]
- Sonnendecker, C.; Zimmermann, W. Change of the product specificity of a cyclodextrin glucanotransferase by semi-rational mutagenesis to synthesize large-ring cyclodextrins. Catalysts 2019, 9, 242. [Google Scholar] [CrossRef] [Green Version]
- Kuttiyawong, K.; Saehu, S.; Ito, K.; Pongsawasdi, P. Synthesis of large-ring cyclodextrin from tapioca starch by amylomaltase and complex formation with Vitamin E acetate for solubility enhancement. Process Biochem. 2015, 50, 2168–2176. [Google Scholar] [CrossRef]
- Kim, J.H.; Wang, R.; Lee, W.H.; Park, C.S.; Lee, S.; Yoo, S.H. One-pot synthesis of cycloamyloses from sucrose by dual enzyme treatment: Combined reaction of amylosucrase and 4-α-glucanotransferase. J. Agric. Food Chem. 2011, 59, 5044–5051. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhen, C.; Liu, L.; Wu, L. Streptomyces ST66 amylomaltase gene cloning and expression and production of cycloamylose. In Proceedings of the 2009 3rd International Conference on Bioinformatics and Biomedical Engineering, Beijing, China, 11–13 June 2009. [Google Scholar]
- Vongpichayapaiboon, T.; Pongsawasdi, P.; Krusong, K. Optimization of large-ring cyclodextrin production from starch by amylomaltase from Corynebacterium glutamicum and effect of organic solvent on product size. J. Appl. Microbiol. 2016, 120, 912–920. [Google Scholar] [CrossRef]
- Nimpiboon, P.; Tumhom, S.; Nakapong, S.; Pongsawasdi, P. Amylomaltase from Thermus filiformis: Expression in Saccharomyces cerevisiae and its use in starch modification. J. Appl. Microbiol. 2020, 129, 1287–1296. [Google Scholar] [CrossRef]
- Tumhom, S.; Krusong, K.; Pongsawasdi, P. Y418 in 410s loop is required for high transglucosylation activity and large-ring cyclodextrin production of amylomaltase from Corynebacterium glutamicum. Biochem. Biophys. Res. Commun. 2017, 488, 516–521. [Google Scholar] [CrossRef]
- Suksiri, P.; Ismail, A.; Sirirattanachatchawan, C.; Wangpaiboon, K.; Muangsin, N.; Tananuwong, K.; Krusong, K. Enhancement of large ring cyclodextrin production using pretreated starch by glycogen debranching enzyme from Corynebacterium glutamicum. Int. J. Biol. Macromol. 2021, 193, 81–87. [Google Scholar] [CrossRef]
- Xu, Y.; Zhou, X.; Bai, Y.; Wang, J.; Wu, C.; Xu, X.; Jin, Z. Cycloamylose production from amylomaize by isoamylase and Thermus aquaticus 4-α-glucanotransferase. Carbohydr. Polym. 2014, 102, 66–73. [Google Scholar] [CrossRef]
- Chu, S.; Hong, J.S.; Rho, S.J.; Park, J.; Han, S.I.; Kim, Y.W.; Kim, Y.R. High-yield cycloamylose production from sweet potato starch using Pseudomonas isoamylase and Thermus aquaticus 4-α-glucanotransferase. Food Sci. Biotechnol. 2016, 25, 1413–1419. [Google Scholar] [CrossRef]
- Park, J.; Rho, S.-J.; Kim, Y.-R. Feasibility and characterization of the cycloamylose production from high amylose corn starch. Cereal Chem. 2018, 95, 838–848. [Google Scholar] [CrossRef]
- Ueda, H. Physicochemical properties and complex formation abilities of large-ring cyclodextrins. J. Incl. Phenom. 2002, 44, 53–56. [Google Scholar] [CrossRef]
- Saokham, P.; Muankaew, C.; Jansook, P.; Loftsson, T. Solubility of cyclodextrins and drug/cyclodextrin complexes. Molecules 2018, 23, 1161. [Google Scholar] [CrossRef] [Green Version]
- Barclay, T.G.; Day, C.M.; Petrovsky, N.; Garg, S. Review of polysaccharide particle-based functional drug delivery. Carbohydr. Polym. 2019, 221, 94–112. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; Kerdpol, K.; Rungrotmongkol, T.; Tananuwong, K.; Ueno, T.; Ekasit, S.; Muangsin, N.; Krusong, K. Solubility enhancement of poorly water soluble domperidone by complexation with the large ring cyclodextrin. Int. J. Pharm. 2021, 606, 120909. [Google Scholar] [CrossRef] [PubMed]
- Rho, S.J.; Mun, S.; Hong, J.S.; Kim, Y.L.; Do, H.V.; Kim, Y.W.; Han, S.I.; Kim, Y.R. Physicochemical interactions of cycloamylose with phenolic compounds. Carbohydr. Polym. 2017, 174, 980–989. [Google Scholar] [CrossRef]
- Kerdpol, K.; Nutho, B.; Krusong, K.; Poo-arporn, R.P.; Rungrotmongkol, T.; Hannongbua, S. Encapsulation of α-tocopherol in large-ring cyclodextrin containing 26 α-d-glucopyranose units: A molecular dynamics study. J. Mol. Liq. 2021, 339, 116802. [Google Scholar] [CrossRef]
- Machida, S.; Ogawa, S.; Xiaohua, S.; Takaha, T.; Fujii, K.; Hayashi, K. Cycloamylose as an efficient artificial chaperone for protein refolding. FEBS Lett. 2000, 486, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Tahara, Y.; Yasuoka, J.; Sawada, S.; Sasaki, Y.; Akiyoshi, K. Effective CpG DNA delivery using amphiphilic cycloamylose nanogels. Biomater. Sci. 2015, 3, 256–264. [Google Scholar] [CrossRef]
- Fujii, H.; Shin-Ya, M.; Takeda, S.; Hashimoto, Y.; Mukai, S.A.; Sawada, S.; Adachi, T.; Akiyoshi, K.; Miki, T.; Mazda, O. Cycloamylose-nanogel drug delivery system-mediated intratumor silencing of the vascular endothelial growth factor regulates neovascularization in tumor microenvironment. Cancer Sci. 2014, 105, 1616–1625. [Google Scholar] [CrossRef] [Green Version]
- Toita, S.; Morimoto, N.; Akiyoshi, K. Functional cycloamylose as a polysaccharide-based biomaterial: Application in a gene delivery system. Biomacromolecules 2010, 11, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Toita, S.; Soma, Y.; Morimoto, N.; Akiyoshi, K. Cycloamylose-based biomaterial: Nanogel of cholesterol-bearing cationic cycloamylose for siRNA delivery. Chem. Lett. 2009, 38, 1114–1115. [Google Scholar] [CrossRef]
- Saehu, S.; Srisimarat, W.; Prousoontorn, M.H.; Pongsawasdi, P. Transglucosylation reaction of amylomaltase for the synthesis of anticariogenic oligosaccharides. J. Mol. Catal. B Enzym. 2013, 88, 77–83. [Google Scholar] [CrossRef]
- Kaulpiboon, J.; Poomipark, N.; Watanasatitarpa, S. Expression and characterization of amylomaltase gene involved in the large-ring cyclodextrin and isomalto-oligosaccharide production. Sci. Technol. Asia 2016, 21, 18–28. [Google Scholar]
- Kaulpiboon, J.; Rudeekulthamrong, P.; Watanasatitarpa, S.; Ito, K.; Pongsawasdi, P. Synthesis of long-chain isomaltooligosaccharides from tapioca starch and an in vitro investigation of their prebiotic properties. J. Mol. Catal. B Enzym. 2015, 120, 127–135. [Google Scholar] [CrossRef]
- Lee, H.S.; Auh, J.H.; Yoon, H.G.; Kim, M.J.; Park, J.H.; Hong, S.S.; Kang, M.H.; Kim, T.J.; Moon, T.W.; Kim, J.W.; et al. Cooperative action of alpha-glucanotransferase and maltogenic amylase for an improved process of isomaltooligosaccharide (IMO) production. J. Agric. Food Chem. 2002, 50, 2812–2817. [Google Scholar] [CrossRef]
- Kaper, T.; van der Maarel, M.J.; Euverink, G.J.; Dijkhuizen, L. Exploring and exploiting starch-modifying amylomaltases from thermophiles. Biochem. Soc. Trans. 2004, 32, 279–282. [Google Scholar] [CrossRef] [Green Version]
- van der Maarel, M.J.; Leemhuis, H. Starch modification with microbial alpha-glucanotransferase enzymes. Carbohydr. Polym. 2013, 93, 116–121. [Google Scholar] [CrossRef] [Green Version]
- Euverink, G.J.W.; Binnema, D.J. Use of Modified Starch as an Agent for Forming a Thermoreversible Gel. U.S. Patent No. 6,864,063, 8 March 2005. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
| Enzyme Name | Source | Optimum Condition | References | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Cyclization | Disproportionation | Hydrolysis | Coupling | |||||||||||
| T °C | pH | Specific Activity (U/mg) | T °C | pH | Specific Activity (U/mg) | T °C | pH | Specific Activity (U/mg) | T °C | pH | Specific Activity (U/mg) | |||
| Archaea | ||||||||||||||
| AM | Pyrobaculum aerophilum IM2 | N/A | N/A | N/A | 95 | 6.7 | 450 | N/A | N/A | N/A | N/A | N/A | N/A | [23] |
| AM | Pyrobaculum calidifontis A3MU77 | N/A | N/A | N/A | 80 | 6.9 | 690 | N/A | N/A | N/A | N/A | N/A | N/A | [24] |
| Bacteria | ||||||||||||||
| AM | Aquifex aeolicus VF5 | N/A | N/A | N/A | 90 | 6.6 | 44.2 | N/A | N/A | N/A | N/A | N/A | N/A | [25] |
| AM | Corynebacterium glutamicum ATCC 13032 | 30 | 6.0 | 0.50 | 30–45 | 6.0 | 21.8–44.3 | N/A | N/A | 0.02–8.05 | N/A | N/A | 0.03 | [32,33,36] |
| AM | Streptococcus agalactiae YZ1605 | 30 | 6.0 | 0.9 | 40 | 6.0 | 54 | N/A | N/A | 0.05 | N/A | N/A | 0.19 | [31] |
| AM | Thermus aquaticus ATCC 33923 | N/A | N/A | N/A | 75 | 5.5–6.0 | 2.9 | N/A | N/A | N/A | N/A | N/A | N/A | [26] |
| AM | Thermus filiformis | 70 | 5.0 | 0.64 | 60 | 6.5 | 159 | N/A | N/A | 1.86 × 10−2 | N/A | N/A | 6.91 × 10−2 | [27] |
| 4αGTase/AM (TBGT) | Thermus brockianus | N/A | N/A | N/A | 70 | 6.0 | 70734 | N/A | N/A | N/A | N/A | N/A | N/A | [28] |
| 4αGTase/AM | Thermus thermophilus | N/A | N/A | N/A | 72–75 | 5.5–6.3 | 400 | N/A | N/A | N/A | N/A | N/A | N/A | [29] |
| 4αGTase | Acidothermus cellulolyticus 11B | N/A | N/A | N/A | 75 | 7.5 | N/A | N/A | N/A | N/A | N/A | N/A | N/A | [37] |
| 4αGTase | Borrelia burgdorferi | N/A | N/A | N/A | 37 | 5.5 | N/A | N/A | N/A | N/A | N/A | N/A | N/A | [34] |
| 4αGTase | Escherichia coli str. K-12 | N/A | N/A | N/A | 28 | 6.9 | 9400 | N/A | N/A | N/A | N/A | N/A | N/A | [38] |
| 4αGTase | Saccharophagus degradans 2-40 | N/A | N/A | N/A | 35 | 8.5 | N/A | N/A | N/A | N/A | N/A | N/A | N/A | [39] |
| 4αGTase | Synechocystis sp. PCC 6803 | N/A | N/A | N/A | 45 | 7.0 | 5.84 | N/A | N/A | N/A | N/A | N/A | N/A | [40] |
| 4αGTase | Thermus scotoductus | N/A | N/A | N/A | 75 | 7.5 | N/A | N/A | N/A | N/A | N/A | N/A | N/A | [30] |
| Plant | ||||||||||||||
| D-enzyme (AtDPE1) | Arabidopsis thaliana | N/A | N/A | N/A | 37 | 6.0–8.0 | N/A | N/A | N/A | N/A | N/A | N/A | N/A | [41] |
| D-enzyme (AtDPE2) | Arabidopsis thaliana | N/A | N/A | N/A | 42 | 7.0 | N/A | N/A | N/A | N/A | N/A | N/A | N/A | [42] |
| D-enzyme (MeDPE1) | Manihot esculenta Crantz | N/A | N/A | N/A | 37 | 6.0–8.0 | N/A | N/A | N/A | N/A | N/A | N/A | N/A | [41] |
| D-enzyme (StDPE) | Solanum tuberosum | N/A | N/A | N/A | 45 | 6.7 | 47.5 | N/A | N/A | N/A | N/A | N/A | N/A | [13,19] |
| D-enzyme (OsDPE1 and OsDPE2) | Oryza sativa L. | N/A | N/A | N/A | 30–39 | 6.0–7.0 | N/A | N/A | N/A | N/A | N/A | N/A | N/A | [43] |
| Enzyme | Origin | Substrate | Degree of Polymerization | References |
|---|---|---|---|---|
| AM | Streptococcus agalactiae | Pea starch | DP22–DP50 | [31] |
| AM | Thermus aquaticus | Synthetic amylose | DP22–DP > 60 | [26] |
| AM | Corynebacterium glutamicum | Pea starch | DP19–DP50 | [63] |
| AM | Corynebacterium glutamicum | Tapioca starch | DP22–DP54 | [36] |
| AM | Thermus filiformis | Pea starch | DP22–DP60 | [27] |
| AM | Streptomyces ST66 | Potato amylose | N/A | [65] |
| 4αGTase | Synechocystis sp. PCC 6803 | Sucrose | DP24–DP284 | [64] |
| 4αGTase | Synechocystis sp. PCC 6803 | Corn starch | DP12–DP36 | [40] |
| 4αGTase | Deinococcus geothermalis | Amylose | DP5–DP37 | [14] |
| 4αGTase | Aquifex aeolicus | Amylose | DP16–DP50 | [25] |
| D-enzyme | Manihot esculenta Crantz | Potato amylose | DP16–DP > 60 | [41] |
| D-enzyme | Arabidopsis thaliana (AtDPE1) | Potato amylose | DP16–DP50 | [41] |
| D-enzyme | Potato tuber | Synthetic amylose | DP17–DP > 100 | [13] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krusong, K.; Ismail, A.; Wangpaiboon, K.; Pongsawasdi, P. Production of Large-Ring Cyclodextrins by Amylomaltases. Molecules 2022, 27, 1446. https://doi.org/10.3390/molecules27041446
Krusong K, Ismail A, Wangpaiboon K, Pongsawasdi P. Production of Large-Ring Cyclodextrins by Amylomaltases. Molecules. 2022; 27(4):1446. https://doi.org/10.3390/molecules27041446
Chicago/Turabian StyleKrusong, Kuakarun, Abbas Ismail, Karan Wangpaiboon, and Piamsook Pongsawasdi. 2022. "Production of Large-Ring Cyclodextrins by Amylomaltases" Molecules 27, no. 4: 1446. https://doi.org/10.3390/molecules27041446
APA StyleKrusong, K., Ismail, A., Wangpaiboon, K., & Pongsawasdi, P. (2022). Production of Large-Ring Cyclodextrins by Amylomaltases. Molecules, 27(4), 1446. https://doi.org/10.3390/molecules27041446