
Archaeal and Bacterial Content in a Two-Stage Anaerobic System for Efficient Energy Production from Agricultural Wastes

 ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setup
2.2. Inoculum Preparation
2.3. Analytical Methods
2.3.1. Biogas Volume Measurement
2.3.2. Microscopic Observation
2.3.3. Cellulose Concentration Estimation
2.3.4. Volatile Fatty Acid Determination
2.4. Metagenome Sequencing and Bioinformatics Analysis
2.5. Statistical Analysis
3. Results and Discussion
3.1. Biogas Production Using Cascade Bioreactors
3.2. Microbial Communities in the Two-Stage Anaerobic System Identified by Metagenomics
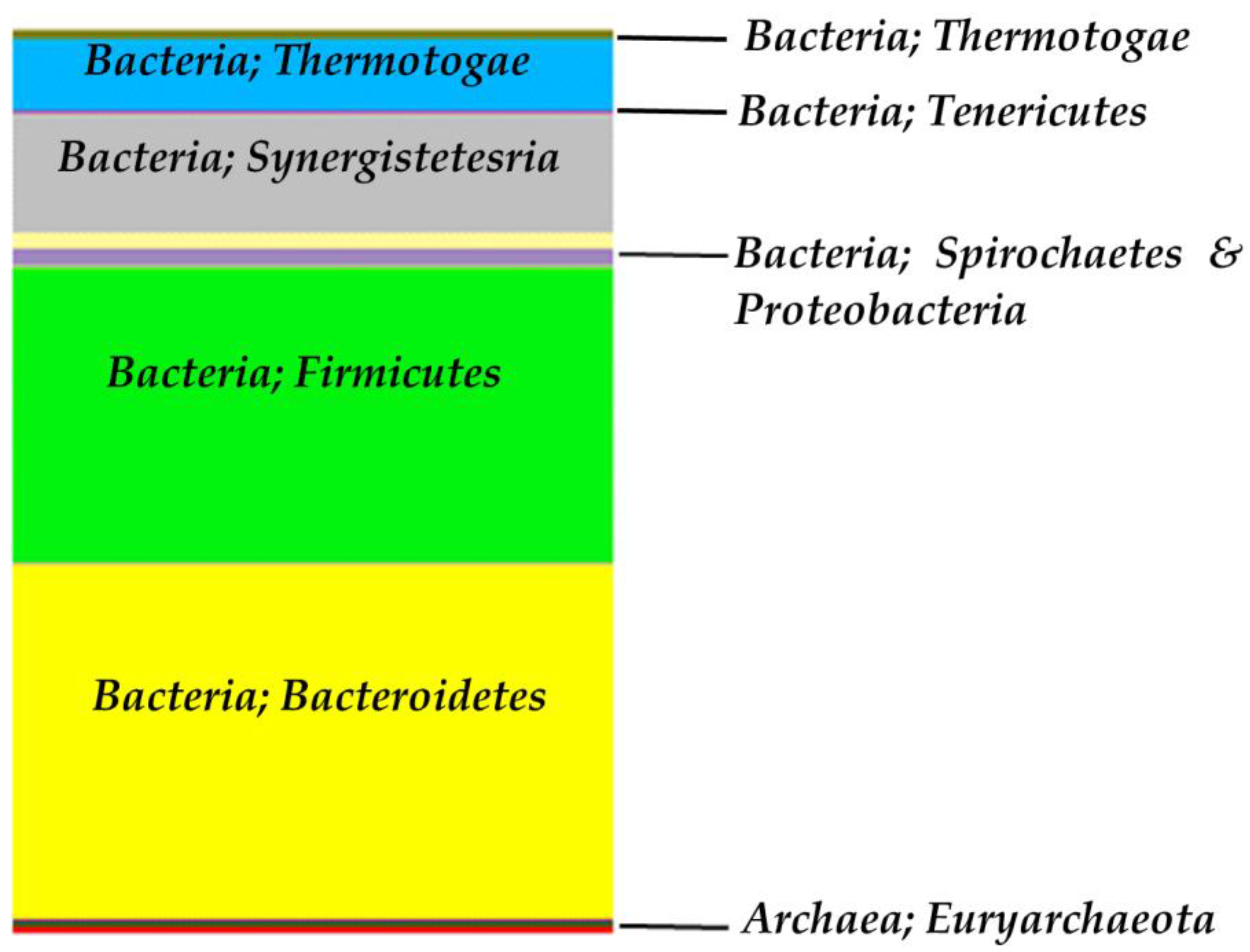
3.2.1. Bacterial Biodiversity
3.2.2. Archaeal Biodiversity
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| AD | anaerobic digestion |
| DGGE | denaturing gradient gel electrophoresis |
| FISH | fluorescence in situ hybridization |
| BR | bioreactor |
| Yp | Yield of energy carrier per ton wheat straw added, [m3·t−1] |
| LHV | lower heating value, [kWh·Nm−3] |
| Ee | primary energy obtained during the anaerobic digestion process, [kWh·t−1] |
| VFA | volatile fatty acids, [g/L] |
| Ac | acetate, [g/L] |
| Prop | propionate, [g/L] |
| i-But | isobutyrate, [g/L] |
| But | butyrate, [g/L] |
| i-Val | isovalerate, [g/L] |
| Val | valerate, [g/L] |
| Cap | capric acid, [g/L] |
| PASC | phosphoric acid-swollen cellulose |
| NCBI | National Center for Biotechnology Information |
| rDNA | ribosomal deoxyribonucleic acid |
| FID | flame ionization detector |
| SD | standard deviation |
References
- Sadh, P.K.; Duhan, S.; Duhan, J.S. Agro-industrial wastes and their utilization using solid state fermentation: A review. Bioresour. Bioprocess. 2018, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.; Ouyang, D.; Zhou, Z.; Page, J.S.; Liu, D.; Zhao, X. Lignocellulosic biomass as sustainable feedstock and materials for power generation and energy storage. J. Energy Chem. 2021, 57, 247–280. [Google Scholar] [CrossRef]
- Monlau, F.; Barakat, A.; Trably, E.; Dumas, C.; Steyer, J.-P.; Carrere, H. Lignocellulosic materials into biohydrogen and biomethane: Impact of structural features and pretreatment. Crit. Rev. Environ. Sci. Technol. 2012, 43, 260–322. [Google Scholar] [CrossRef]
- He, Y.; Pang, Y.; Liu, Y.; Li, X.; Wang, K. Physicochemical characterization of rice straw pretreated with sodium hydroxide in the solid state for enhancing biogas production. Energy Fuels 2008, 22, 2775–2781. [Google Scholar] [CrossRef]
- Naik, S.N.; Goud, V.V.; Rout, P.K.; Dalai, A.K. Production of first and second generation biofuels: A comprehensive review. Renew. Sustain. Energy Rev. 2010, 14, 578–597. [Google Scholar] [CrossRef]
- Ali Shah, F.; Mahmood, Q.; Shah, M.M.; Pervez, A.; Asad, S.A. Microbial Ecology of Anaerobic Digesters: The Key Players of Anaerobiosis. Hindawi Publishing Corporation. Sci. World J. 2014, 2014, 21. [Google Scholar] [CrossRef] [PubMed]
- Chandra, R.; Yadav, S.; Kumar, V. Microbial Degradation of Lignocellulosic Waste and Its Metabolic Products. In Environmental Waste Management, 1st ed.; CRC Press: Boca Raton, FL, USA, 2015. [Google Scholar]
- Sun, Y.; Zhuang, J.P.; Lin, L.; Ouyang, P.K. Clean conversion of cellulose into fermentable glucose. Biotechnol. Adv. 2009, 27, 625–632. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.L.; Zeng, G.M.; Feng, C.L.; Hu, S.; Zhao, M.H.; Lai, C.; Zhang, Y.; Jiang, X.Y.; Liu, H.L. Mycelial growth and solid-state fermentation of lignocellulosic waste by white-rot fungus Phanerochaete chrysosporium under lead stress. Chemosphere 2010, 81, 1091–1097. [Google Scholar] [CrossRef] [PubMed]
- Braz, G.H.R.; Fernandez-Gonzalez, N.; Lema, J.M.; Carballa, M. The time response of anaerobic digestion microbiome during an organic loading rate shock. Appl. Microbiol. Biotechnol. 2018, 102, 10285–10297. [Google Scholar] [CrossRef]
- Regueiro, L.; Veiga, P.; Figueroa, M.; Alonso-Gutierrez, J.; Stams, A.J.M.; Lema, J.M.; Carballa, M. Relationship between microbial activity and microbial community structure in six full-scale anaerobic digesters. Microbiol. Res. 2012, 167, 581–589. [Google Scholar] [CrossRef]
- Demirel, B.; Scherer, P. The roles of acetotrophic and hydrogenotrophic methanogens during anaerobic conversion of biomass to methane: A review. Rev. Environ. Sci. Biotechnol. 2008, 7, 173–190. [Google Scholar] [CrossRef]
- Cortes-Tolalpa, L.; Salles, J.F.; van Elsas, J.D. Bacterial Synergism in Lignocellulose Biomass Degradation–Complementary Roles of Degraders As Influenced by Complexity of the Carbon Source. Front. Microbiol. 2017, 8, 1628. [Google Scholar] [CrossRef]
- Rivière, D.; Desvignes, V.; Pelletier, E.; Chaussonnerie, S.; Guermazi, S.; Weissenbach, J.; Li, T.; Camacho, P.; Sghir, A. Towards the definition of a core of microorganisms involved in anaerobic digestion of sludge. ISME J. 2009, 3, 700–714. [Google Scholar] [CrossRef] [Green Version]
- Venkateswaran, S.; Deepa, S. Assessment of Groundwater Quality Using GIS Techniques in Vaniyar Watershed, Ponnaiyar River, Tamil Nadu. Aquat. Procedia 2015, 4, 1283–1290. [Google Scholar] [CrossRef]
- Seon, J.; Lee, T.; Lee, S.C.; Pham, H.D.; Woo, H.C.; Song, M. Bacterial community structure in maximum volatile fatty acids production from alginate in acidogenesis. Biores. Technol. 2014, 157, 22–27. [Google Scholar] [CrossRef]
- Imachi, H.; Aoi, K.; Tasumi, E.; Saito, Y.; Yamanaka, Y.; Saito, Y.; Yamaguchi, T.; Tomaru, H.; Takeuchi, R.; Morono, Y.; et al. Cultivation of methanogenic community from subseafloor sediments using a continuous-flow bioreactor. ISME J. 2011, 5, 1913–1925. [Google Scholar] [CrossRef] [Green Version]
- Amani, T.; Nosrati, M.; Sreekrishnan, T.R. Anaerobic digestion from the viewpoint of microbiological, chemical, and operational aspects—A review. Environ. Rev. 2010, 18, 1–20. [Google Scholar] [CrossRef]
- Kato, S.; Haruta, S.; Cui, Z.J.; Ishii, M.; Igarashi, Y. Effective cellulose degradation by a mixed-culture system composed of a cellulolytic Clostridium and aerobic non-cellulolytic bacteria. FEMS Microbiol. Ecol. 2004, 51, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Kanokratana, P.; Wongwilaiwalin, S.; Mhuantong, W.; Tangphatsornruang, S.; Eurwilaichitr, L.; Champreda, V. Characterization of cellulolytic microbial consortium enriched on Napier grass using metagenomic approaches. J. Biosci. Bioeng. 2018, 125, 439–447. [Google Scholar] [CrossRef]
- Carlos, C.; Fan, H.; Currie, C.R. Substrate Shift Reveals Roles for Members of Bacterial Consortia in Degradation of Plant Cell Wall Polymers. Front. Microbiol. 2018, 9, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stolze, Y.; Zakrzewski, M.; Maus, I.; Eikmeyer, F.G.; Jaenicke, S.; Rottmann, N.; Siebner, C.; Pühler, A.; Schlüter, A. Comparative metagenomics of biogas-producing microbial communities from production-scale biogas plants operating under wet or dry fermentation conditions. Biotechnol. Biofuels 2015, 8, 14. [Google Scholar] [CrossRef] [Green Version]
- Monlau, F.; Barakat, A.; Steyer, J.P.; Carrere, H. Comparison of seven types of thermo-chemical pretreatments on the structural features and anaerobic digestion of sunflower stakls. Bioresour. Technol. 2012, 120, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Updegraff, M.D. Semimicro Determination of Cellulose in Biological Materials. Anal. Biochem. 1969, 32, 420–424. [Google Scholar] [CrossRef]
- Pausan, M.R.; Csorba, C.; Singer, G.; Till, H.; Schöpf, V.; Santigli, E.; Klug, B.; Högenauer, C.; Blohs, M.; Moissl-Eichinger, C. Exploring the Archaeome: Detection of Archaeal Signatures in the Human Body. Front. Microbiol. 2019, 10, 2796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Yin, Y. Fermentative hydrogen production using pretreated microalgal biomass as feedstock. Microb. Cell Fact. 2018, 17, 22. [Google Scholar] [CrossRef] [Green Version]
- Bertin, L.; Grilli, S.; Spagni, A.; Fava, F. Innovative two-stage anaerobic process for effective codigestion of cheese whey and cattle manure. Bioresour. Technol. 2013, 128, 779–783. [Google Scholar] [CrossRef]
- Baldi, F.; Pecorini, I.; Iannelli, R. Comparison of single-stage and two-stage anaerobic co-digestion of food waste and activated sludge for hydrogen and methane production. Renew. Energy 2019, 143, 1755–1765. [Google Scholar] [CrossRef]
- Markphan, W.; Mamimin, C.; Suksong, W.; Prasertsan, P.; O-Thong, S. Comparative assessment of single-stage and two-stage anaerobic digestion for biogas production from high moisture municipal solid waste. Peer J. 2020, 8, e9693. [Google Scholar] [CrossRef]
- Qyyum, M.A.; Ismail, S.; Ni, S.Q.; Ihsanullah, I.; Ahmad, R.; Khan, A.; Tawfik, A.; Nizami, A.-S.; Lee, M. Harvesting biohydrogen from industrial wastewater: Production potential, pilot-scale bioreactors, commercialization status, techno-economics, and policy analysis. J. Clean. Prod. 2022, 340, 130809. [Google Scholar] [CrossRef]
- Hosseinzadeh, A.; Zhou, J.L.; Li, X.; Afsari, M.; Altaee, A. Techno-economic and environmental impact assessment of hydrogen production processes using bio-waste as renewable energy resource. Renew. Sustain. Energy Rev. 2022, 156, 111991. [Google Scholar] [CrossRef]
- La Licata, B.; Sagnelli, F.; Boulanger, A.; Lanzini, A.; Leone, P.; Zitella, P.; Santarelli, M. Bio-hydrogen production from organic wastes in a pilot plant reactor and its use in a SOFC. Int. J. Hydrog. Energy 2011, 36, 7861–7865. [Google Scholar] [CrossRef]
- Schink, B. Energetics of syntrophic cooperation in methanogenic degradation. Microbiol. Molec. Biol. Rev. 1997, 61, 262–280. [Google Scholar]
- Hwang, S.; Lee, Y.; Yang, K. Maximization of acetic acid production in partial acidogenesis of swine wastewater. Biotechnol. Bioeng. 2001, 75, 521–529. [Google Scholar] [CrossRef]
- Maspolim, Y.; Zhou, Y.; Guo, C.; Xiao, K.; Jern Ng, W. Comparison of single-stage and two-phase anaerobic sludge digestion systems–Performance and microbial community dynamics. Chemosphere 2015, 140, 54–62. [Google Scholar] [CrossRef]
- Xu, R.; Zhang, K.; Liu, P.; Khan, A.; Xiong, J.; Tian, F.; Li, X. A critical review on the interaction of substrate nutrient balance and microbial community structure and function in anaerobic co-digestion. Bioresour. Technol. 2018, 247, 1119–1127. [Google Scholar] [CrossRef]
- Guo, B.; Li, L.; Liu, Y. Role of syntrophic acetate oxidation and hydrogenotrophic methanogenesis in co-digestion of blackwater with food waste. J. Clean. Prod. 2021, 283, 125393. [Google Scholar] [CrossRef]
- Hahnke, S.; Langer, T.; Koeck, D.E.; Klocke, M. Description of Proteiniphilum saccharofermentans sp. nov., Petrimonas mucosa sp. nov. and Fermentimonas caenicola gen. nov., sp. nov., isolated from mesophilic laboratory-scale biogas reactors, and emended description of the genus Proteiniphilum. Int. J. Syst. Evol. Microbiol. 2016, 66, 1466–1475. [Google Scholar] [CrossRef] [PubMed]
- Tomazetto, G.; Hahnke, S.; Wibberg, D.; Pühler, A.; Klocke, M.; Schlüter, A. Proteiniphilum saccharofermentans str. M3/6T isolated from a laboratory biogas reactor is versatile in polysaccharide and oligopeptide utilization as deduced from genome-based metabolic reconstructions. Biotechnol. Rep. 2018, 18, e00254. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Mamuad, L.L.; Islam, M.; Lee, S.S. Reductive acetogens isolated from ruminants and their effect on in vitro methane mitigation and milk performance in Holstein cows. J. Anim. Sci. Technol. 2020, 62, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Feng, K.; Wang, Q.; Li, H.; Du, X.; Zhang, Y. Microbial mechanism of enhancing methane production from anaerobic digestion of food waste via phase separation and pH control. J. Environ. Manag. 2021, 288, 112460. [Google Scholar] [CrossRef]
- Chen, S.; Dong, X. Proteiniphilum acetatigenes gen. nov., sp. nov., from a UASB reactor treating brewery wastewater. Int. J. Syst. Evol. Microbiol. 2005, 55, 2257–2261. [Google Scholar] [CrossRef] [Green Version]
- Muñoz-Páez, K.M.; Alvarado-Michi, E.L.; Moreno-Andrade, I.; Buitrón, G.; Valdez-Vazquez, I. Comparison of suspended and granular cell anaerobic bioreactors for hydrogen production from acid agave bagasse hydrolyzates. Int. J. Hydrog. Energy 2020, 45, 275–285. [Google Scholar] [CrossRef]
- Castelló, E.; Perna, V.; Wenzel, J.; Borzacconi, L.; Etchebehere, C. Microbial community composition and reactor performance during hydrogen production in a UASB reactor fed with raw cheese whey inoculated with compost. Water Sci. Technol. 2011, 64, 2265–2273. [Google Scholar] [CrossRef] [PubMed]
- Mariakakis, I.; Bischoff, P.; Krampe, J.; Meyer, C.; Steinmetz, H. Effect of organic loading rate and solids retention time on microbial population during bio-hydrogen production by dark fermentation in large lab-scale. Int. J. Hydrog. Energy 2011, 36, 10690–10700. [Google Scholar] [CrossRef]
- Takahashi, N.; Yamada, T. Glucose metabolism by Prevotella intermedia and Prevotella nigrescens. Oral Microbiol. Immunol. 2000, 15, 188–195. [Google Scholar] [CrossRef]
- Nishiyama, T.; Ueki, A.; Kaku, N.; Watanabe, K.; Ueki, K. Bacteroides graminisolvens sp. nov., a xylanolytic anaerobe isolated from a methanogenic reactor treating cattle waste. Int. J. Syst. Evol. Microbiol. 2009, 59, 1901–1907. [Google Scholar] [CrossRef] [Green Version]
- Hatamoto, M.; Kaneshige, M.; Nakamura, A.; Yamaguchi, T. Bacteroides luti sp. nov., an anaerobic, cellulolytic and xylanolytic bacterium isolated from methanogenic sludge. Int. J. Syst. Evol. Microbiol. 2014, 64, 1770–1774. [Google Scholar] [CrossRef]
- Sarkar, O.; Kumar, A.N.; Dahiya, S.; Krishna, K.V.; Yeruva, D.K.; Mohan, S.V. Regulation of acidogenic metabolism towards enhanced short chain fatty acid biosynthesis from waste: Metagenomic profiling. RSC Adv. 2016, 6, 18641–18653. [Google Scholar] [CrossRef]
- Ueki, A.; Ohtaki, Y.; Kaku, N.; Ueki, K. Descriptions of Anaerotaenia torta gen. nov., sp. nov. and Anaerocolumna cellulosilytica gen. nov., sp. nov. isolated from a methanogenic reactor of cattle waste and reclassification of Clostridium aminovalericum, Clostridium jejuense and Clostridium xylanovorans as Anaerocolumna species. Int. J. Syst. Evol. Microbiol. 2016, 66, 2936–2943. [Google Scholar]
- Kim, S.; Jeong, H.; Kim, S.; Chun, J. Clostridium ganghwense sp. nov., isolated from tidal flat sediment. Int. J. Syst. Evol. Microbiol. 2006, 56, 691–693. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.; Jeon, B.S.; Sang, B.I. An efficient new process for the selective production of odd-chain carboxylic acids by simple carbon elongation using Megasphaera hexanoica. Sci. Rep. 2019, 9, 11999. [Google Scholar] [CrossRef] [Green Version]
- Kim, B.C.; Jeon, B.S.; Kim, S.; Kim, H.; Um, Y.; Sang, B.I. Caproiciproducens galactitolivorans gen. nov., sp. nov., a bacterium capable of producing caproic acid from galactitol, isolated from a wastewater treatment plant. Int. J. Syst. Evol. Microbiol. 2015, 65, 4902–4908. [Google Scholar] [CrossRef]
- Qin, Y.; Li, L.; Wu, J.; Xiao, B.; Hojo, T.; Kubota, K.; Cheng, J.; Li, Y.Y. Co-production of biohydrogen and biomethane from food waste and paper waste via recirculated two-phase anaerobic digestion process: Bioenergy yields and metabolic distribution. Bioresour. Technol. 2019, 276, 325–334. [Google Scholar] [CrossRef]
- Najdenski, H.; Hubenov, V.; Simeonov, I.; Kussovski, V.; Dimitrova, L.; Petrova, P.; Angelov, P.; Kabaivanova, L. Microbial biodegradation as an option for waste utilization during long-term manned space missions. Bulg. Chem. Commun. 2020, 52, 379–386. [Google Scholar]
- Kong, X.; Du, J.; Ye, X.; Xi, Y.; Jin, H.; Zhang, M.; Guo, D. Enhanced methane production from wheat straw with the assistance of lignocellulolytic microbial consortium TC-5. Bioresour. Technol. 2018, 263, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Rettenmaier, R.; Kowollik, M.L.; Klingl, A.; Liebl, W.; Zverlov, V. Ruminiclostridium herbifermentans sp. nov., a mesophilic and moderately thermophilic cellulolytic and xylanolytic bacterium isolated from a lab-scale biogas fermenter fed with maize silage. Int. J. Syst. Evol. Microbiol. 2021, 71, 004692. [Google Scholar] [CrossRef] [PubMed]
- Goud, R.K.; Arunasri, K.; Yeruva, D.K.; Krishna, K.V.; Dahiya, S.; Mohan, S.V. Impact of selectively enriched microbial communities on long-term fermentative biohydrogen production. Bioresour. Technol. 2017, 242, 253–264. [Google Scholar] [CrossRef]
- Tarlera, S.; Muxí, L.; Soubes, M.; Stams, A.J. Caloramator proteoclasticus sp. nov., a new moderately thermophilic anaerobic proteolytic bacterium. Int. J. Syst. Evol. Microbiol. 1997, 47, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Prasertsan, P.; Sompong, O.; Birkeland, N.K. Optimization and microbial community analysis for production of biohydrogen from palm oil mill effluent by thermophilic fermentative process. Int. J. Hydrog. Energy 2009, 34, 7448–7459. [Google Scholar] [CrossRef]
- Hawkes, F.R.; Dinsdale, R.; Hawkes, D.L.; Hussy, I. Sustainable fermentative hydrogen production: Challenges for process optimisation. Int. J. Hydrog. Energy 2002, 27, 1339–1347. [Google Scholar] [CrossRef]
- Rettenmaier, R.; Schneider, M.; Munk, B.; Lebuhn, M.; Jünemann, S.; Sczyrba, A.; Liebl, W. Importance of Defluviitalea raffinosedens for hydrolytic biomass degradation in co-culture with Hungateiclostridium thermocellum. Microorganisms 2020, 8, 915. [Google Scholar] [CrossRef] [PubMed]
- Rettenmaier, R.; Duerr, C.; Neuhaus, K.; Liebl, W.; Zverlov, V.V. Comparison of sampling techniques and different media for the enrichment and isolation of cellulolytic organisms from biogas fermenters. Syst. Appl. Microbiol. 2019, 42, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Finneran, K.T.; Forbush, H.M.; VanPraagh, C.V.G.; Lovley, D.R. Desulfitobacterium metallireducens sp. nov., an anaerobic bacterium that couples growth to the reduction of metals and humic acids as well as chlorinated compounds. Int. J. Syst. Evol. Microbiol. 2002, 52, 1929–1935. [Google Scholar] [PubMed] [Green Version]
- Karataş, S.; Hasar, H.; Taşkan, E.; Özkaya, B.; Şahinkaya, E. Bio-reduction of tetrachloroethene using a H2-based membrane biofilm reactor and community fingerprinting. Water Res. 2014, 58, 21–28. [Google Scholar] [CrossRef]
- Koelschbach, J.S.; Mouttaki, H.; Pickl, C.; Heipieper, H.J.; Rachel, R.; Lawson, P.A.; Meckenstock, R.U. Rectinema cohabitants gen. nov., sp. nov., a rod-shaped spirochaete isolated from an anaerobic naphthalene-degrading enrichment culture. Int. J. Syst. Evol. Microbiol. 2017, 67, 1288–1295. [Google Scholar] [CrossRef]
- Lim, J.W.; Park, T.; Tong, Y.W.; Yu, Z. The microbiome driving anaerobic digestion and microbial analysis. Adv. Bioenergy 2020, 5, 1–61. [Google Scholar]
- Schauer, N.L.; Ferry, J.G. Metabolism of Formate in Methanobacterium formicicum. J. Bacteriol. 1980, 142, 800–807. [Google Scholar] [CrossRef] [Green Version]
- Ziganshin, A.M.; Ziganshina, E.E.; Kleinsteuber, S.; Nikolausz, M. Comparative Analysis of Methanogenic Communities in Different Laboratory-Scale Anaerobic Digesters. Archaea 2016, 2016, 3401272. [Google Scholar] [CrossRef] [PubMed]
- Ma, K.; Liu, X.; Dong, X. Methanobacterium beijingense sp. nov., a novel methanogen isolated from anaerobic digesters. Int. J. Syst. Evol. Microbiol. 2005, 55, 325–329. [Google Scholar] [CrossRef] [Green Version]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| System | Energy Carrier | Cumulative Yield (cm3) | Yp *(m3·t−1) | LHV * (kWh·Nm−3) | Total Energy (kWh·t−1) | Total Energy (per System) (kWh·t−1) |
|---|---|---|---|---|---|---|
| Single-phase | Methane | 29104.0 | 36.38 | 9.94 | 361.62 | 361.62 |
| Two-phase | Hydrogen | 134.7 | 4.49 | 2.99 | 13.43 | 1195.89 |
| Methane | 95168.0 | 118.96 | 9.94 | 1182.46 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kabaivanova, L.; Hubenov, V.; Dimitrova, L.; Simeonov, I.; Wang, H.; Petrova, P. Archaeal and Bacterial Content in a Two-Stage Anaerobic System for Efficient Energy Production from Agricultural Wastes. Molecules 2022, 27, 1512. https://doi.org/10.3390/molecules27051512
Kabaivanova L, Hubenov V, Dimitrova L, Simeonov I, Wang H, Petrova P. Archaeal and Bacterial Content in a Two-Stage Anaerobic System for Efficient Energy Production from Agricultural Wastes. Molecules. 2022; 27(5):1512. https://doi.org/10.3390/molecules27051512
Chicago/Turabian StyleKabaivanova, Lyudmila, Venelin Hubenov, Lyudmila Dimitrova, Ivan Simeonov, Haoping Wang, and Penka Petrova. 2022. "Archaeal and Bacterial Content in a Two-Stage Anaerobic System for Efficient Energy Production from Agricultural Wastes" Molecules 27, no. 5: 1512. https://doi.org/10.3390/molecules27051512
APA StyleKabaivanova, L., Hubenov, V., Dimitrova, L., Simeonov, I., Wang, H., & Petrova, P. (2022). Archaeal and Bacterial Content in a Two-Stage Anaerobic System for Efficient Energy Production from Agricultural Wastes. Molecules, 27(5), 1512. https://doi.org/10.3390/molecules27051512