
Computational Modeling of O-Linked Glycan Biosynthesis in CHO Cells

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
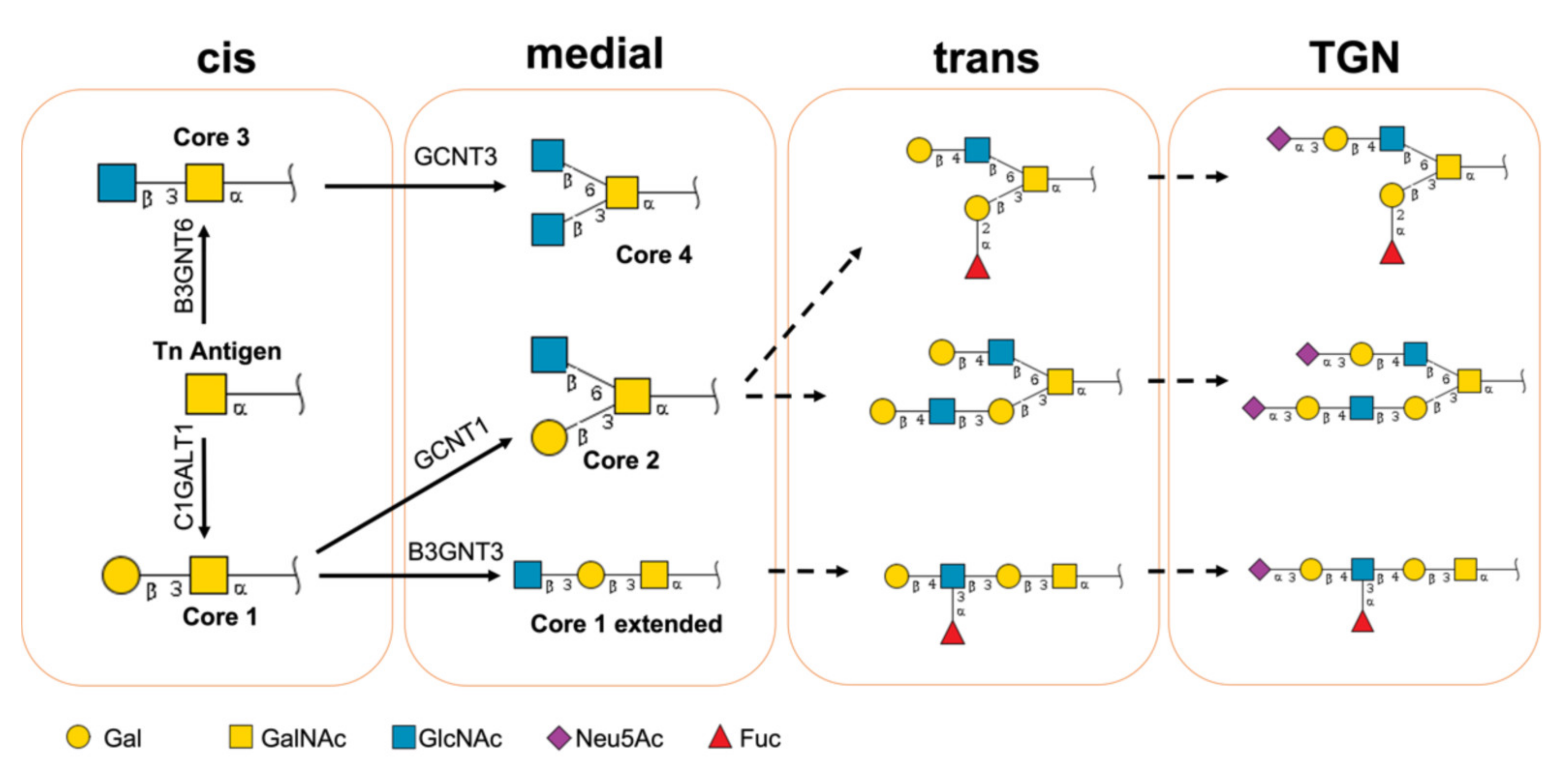
2.1. Glycan Profiles Modeled in this Study
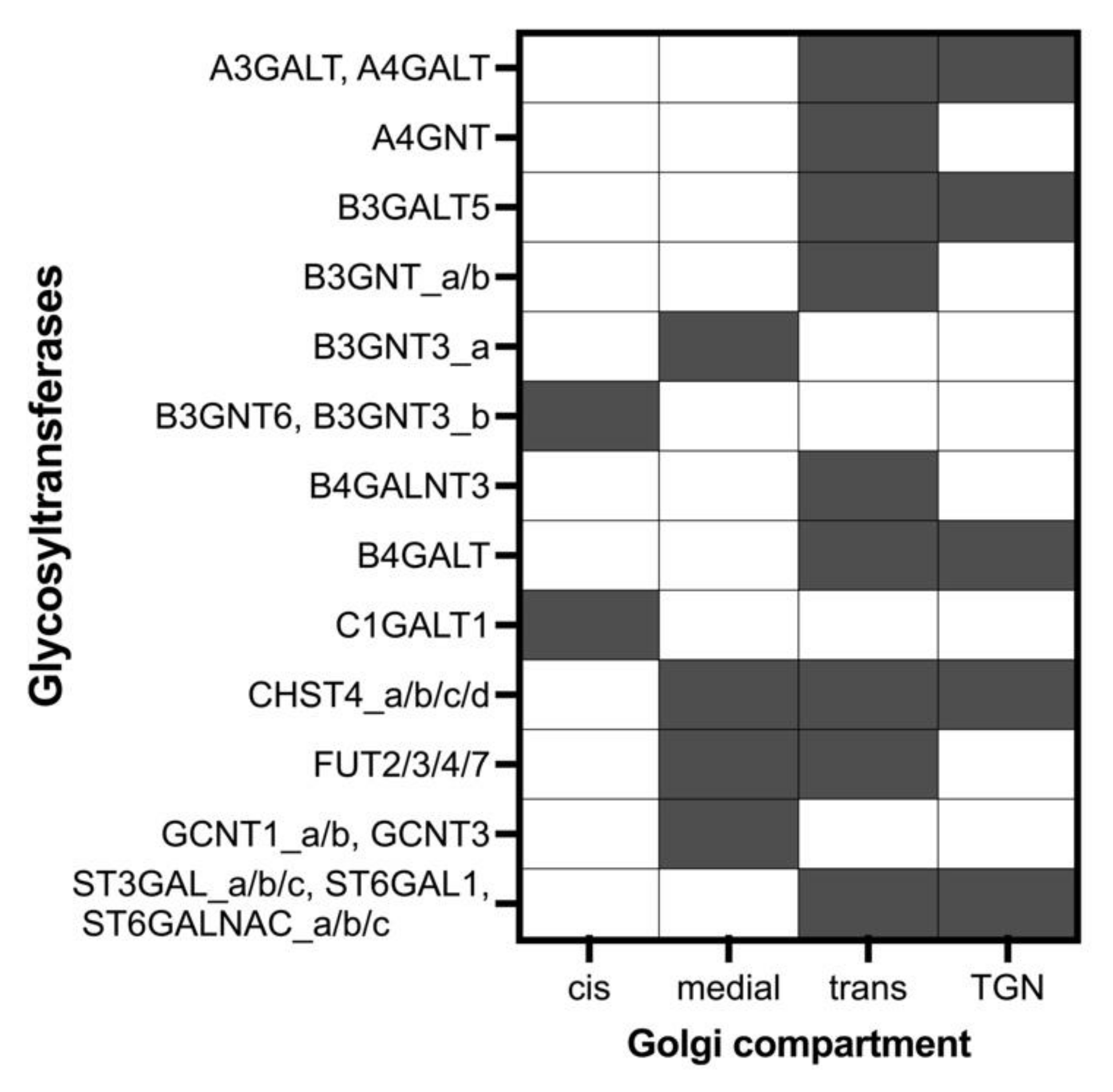
2.2. Building the Models
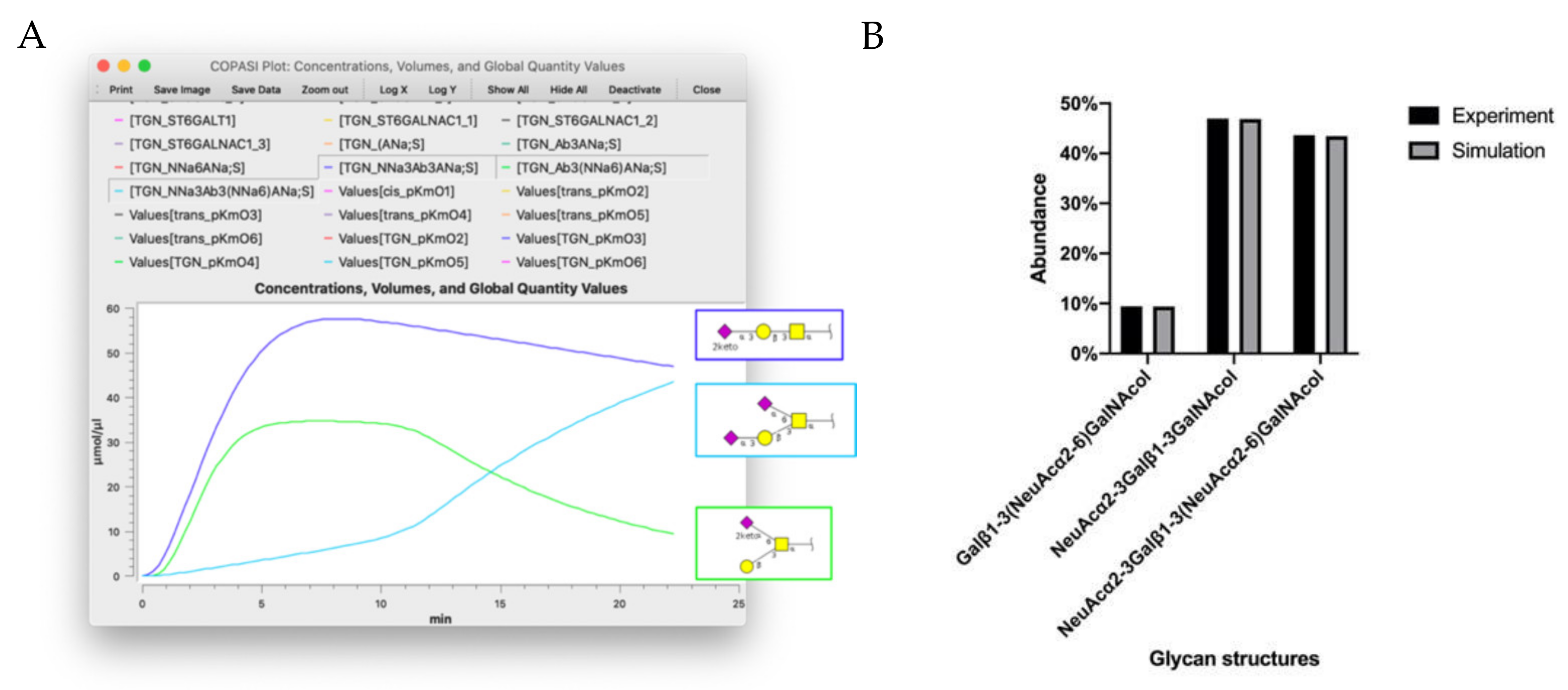
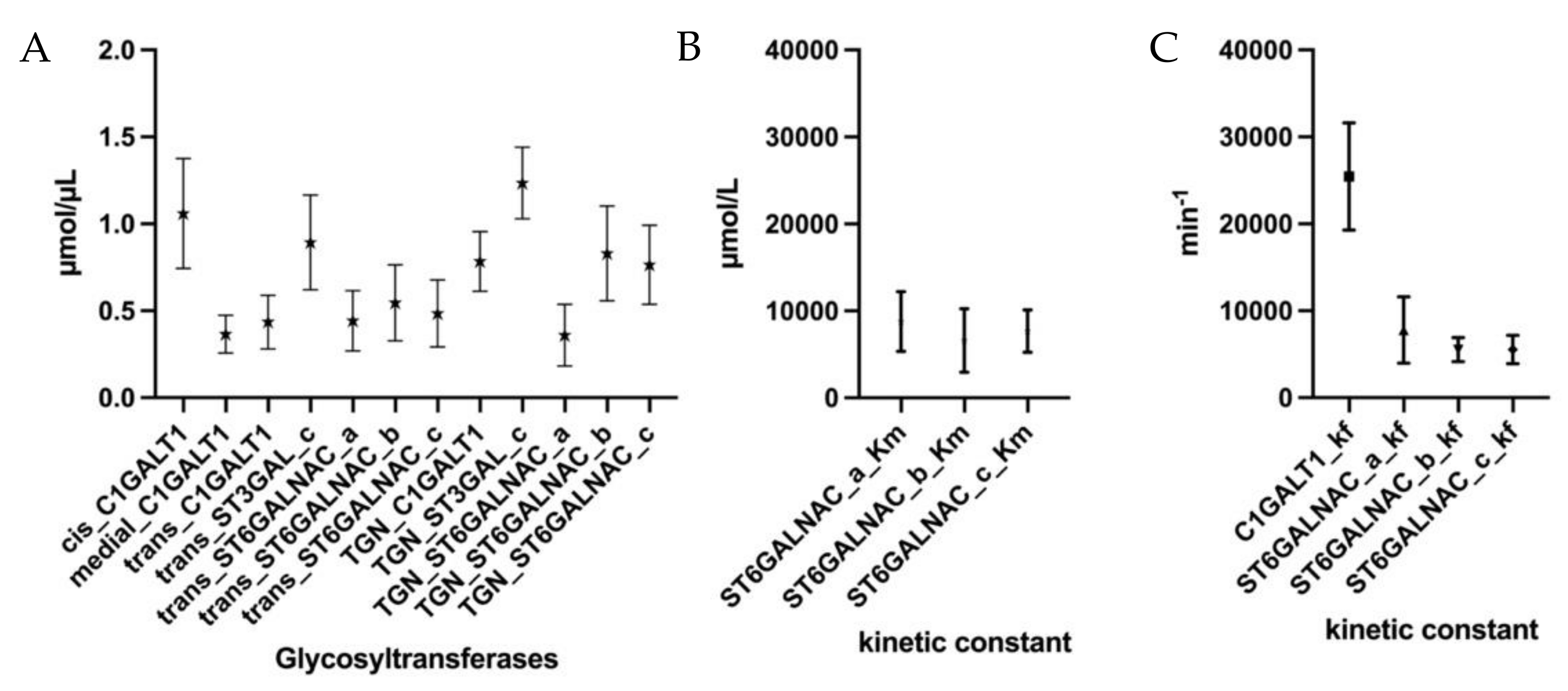
2.3. Parameter Estimation and Simulations
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Rini, J.M.; Esko, J.D. Glycosyltransferases and Glycan-Processing Enzymes. In Essentials of Glycobiology; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2017; pp. 65–77. ISBN 9780879697709. [Google Scholar]
- Varki, A.; Gagneux, P. Biological Functions of Glycans. In Essentials of Glycobiology; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2017; pp. 77–89. [Google Scholar]
- Zhang, L.; Tian, E.; Ten Hagen, K.G. UDP-N-acetyl-alpha-DGalactosamine: Polypeptide N-Acetylgalactosaminyltransferases (ppGalNAc-Ts). In Handbook of Glycosyltransferases and Related Genes, 2nd ed.; Springer: Tokyo, Japan, 2014; Volume 1, pp. 495–511. ISBN 9784431542407. [Google Scholar]
- Kim, J.Y.; Kim, Y.G.; Lee, G.M. CHO cells in biotechnology for production of recombinant proteins: Current state and further potential. Appl. Microbiol. Biotechnol. 2012, 93, 917–930. [Google Scholar] [CrossRef]
- Umaña, P.; Bailey, J.E. A mathematical model of N-linked glycoform biosynthesis. Biotechnol. Bioeng. 1997, 55, 890–908. [Google Scholar] [CrossRef]
- Krambeck, F.J.; Betenbaugh, M.J. A mathematical model of N-linked glycosylation. Biotechnol. Bioeng. 2005, 92, 711–728. [Google Scholar] [CrossRef]
- Krambeck, F.J.; Bennun, S.V.; Narang, S.; Choi, S.; Yarema, K.J.; Betenbaugh, M.J. A mathematical model to derive N-glycan structures and cellular enzyme activities from mass spectrometric data. Glycobiology 2009, 19, 1163–1175. [Google Scholar] [CrossRef]
- Krambeck, F.J.; Bennun, S.V.; Andersen, M.R.; Betenbaugh, M.J. Model-based analysis of N-glycosylation in Chinese hamster ovary cells. PLoS ONE 2017, 12, e0175376. [Google Scholar] [CrossRef]
- Liu, G.; Marathe, D.D.; Matta, K.L.; Neelamegham, S. Systems-level modeling of cellular glycosylation reaction networks: O-linked glycan formation on natural selectin ligands. Bioinformatics 2008, 24, 2740–2747. [Google Scholar] [CrossRef]
- McDonald, A.G.; Tipton, K.F.; Davey, G.P. A Knowledge-Based System for Display and Prediction of O-Glycosylation Network Behaviour in Response to Enzyme Knockouts. PLoS Comput. Biol. 2016, 12, e1004844. [Google Scholar] [CrossRef]
- Liu, J.; Jin, C.; Cherian, R.M.; Karlsson, N.G.; Holgersson, J. O-glycan repertoires on a mucin-type reporter protein expressed in CHO cell pools transiently transfected with O-glycan core enzyme cDNAs. J. Biotechnol. 2015, 199, 77–89. [Google Scholar] [CrossRef]
- Cherian, R.M.; Jin, C.; Liu, J.; Karlsson, N.G.; Holgersson, J. A panel of recombinant mucins carrying a repertoire of sialylated O-glycans based on different core chains for studies of glycan binding proteins. Biomolecules 2015, 5, 1810–1831. [Google Scholar] [CrossRef]
- Moore, K.L.; Eaton, S.F.; Lyons, D.E.; Lichenstein, H.S.; Cummings, R.D.; McEver, R.P. The P-selectin glycoprotein ligand from human neutrophils displays sialylated, fucosylated, O-linked poly-N-acetyllactosamine. J. Biol. Chem. 1994, 269, 23318–23327. [Google Scholar] [CrossRef]
- Liu, J.; Qian, Y.; Holgersson, J. Removal of xenoreactive human anti-pig antibodies by absorption on recombinant mucin-containing glycoproteins carrying the Galα1,3Gal epitope. Transplantation 1997, 63, 1673–1682. [Google Scholar] [CrossRef]
- Löfling, J.; Holgersson, J. Core saccharide dependence of sialyl Lewis X biosynthesis. Glycoconj. J. 2009, 26, 33–40. [Google Scholar] [CrossRef]
- Löfling, J.; Diswall, M.; Eriksson, S.; Borén, T.; Breimer, M.E.; Holgersson, J. Studies of Lewis antigens and H. pylori adhesion in CHO cell lines engineered to express Lewis b determinants. Glycobiology 2008, 18, 494–501. [Google Scholar] [CrossRef]
- Mthembu, Y.H.; Jin, C.; Padra, M.; Liu, J.; Edlund, J.O.; Ma, H.; Padra, J.; Oscarson, S.; Borén, T.; Karlsson, N.G.; et al. Recombinant mucin-type proteins carrying LacdiNAc on different O-glycan core chains fail to support H. pylori binding. Mol. Omics. 2020, 16, 243–257. [Google Scholar] [CrossRef]
- Cherian, R.M.; Jin, C.; Liu, J.; Karlsson, N.G.; Holgersson, J. Recombinant mucin-type fusion proteins with a Galα1,3Gal substitution as Clostridium difficile toxin A inhibitors. Infect. Immun. 2016, 84, 2842–2852. [Google Scholar] [CrossRef]
- Liu, J.; Gustafsson, A.; Breimer, M.E.; Kussak, A.; Holgersson, J. Anti-pig antibody adsorption efficacy of α-Gal carrying recombinant P-selectin glycoprotein ligand-1/immunoglobulin chimeras increases with core 2 β1,6-N-acetylglucosaminyltransferase expression. Glycobiology 2005, 15, 571–583. [Google Scholar] [CrossRef]
- Olson, F.J. A MUC1 tandem repeat reporter protein produced in CHO-K1 cells has sialylated core 1 O-glycans and becomes more densely glycosylated if coexpressed with polypeptide-GalNAc-T4 transferase. Glycobiology 2005, 15, 177–191. [Google Scholar] [CrossRef]
- Bennett, E.P.; Mandel, U.; Clausen, H.; Gerken, T.A.; Fritz, T.A.; Tabak, L.A. Control of mucin-type O-glycosylation: A classification of the polypeptide GalNAc-transferase gene family. Glycobiology 2012, 22, 736–756. [Google Scholar] [CrossRef]
- Brockhausen, I.; Stanley, P. O-GalNAc Glycans. In Essentials of Glycobiology; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2017; pp. 113–125. [Google Scholar]
- Hawkins, C.J. Golgi Apparatus: Structure, Functions and Mechanisms; Hawkins, C.J., Ed.; Cell Biology Research Progress; Nova Science Publishers: New York, NY, USA, 2011; ISBN 9781611220513. [Google Scholar]
- Tu, L.; Banfield, D.K. Localization of Golgi-resident glycosyltransferases. Cell. Mol. Life Sci. 2010, 67, 29–41. [Google Scholar] [CrossRef]
- Gill, D.J.; Clausen, H.; Bard, F. Location, location, location: New insights into O-GalNAc protein glycosylation. Trends Cell Biol. 2011, 21, 149–158. [Google Scholar] [CrossRef]
- Ju, T.; Cummings, R.D. Core 1 β3galactosyltransferase (C1GalT1, T-synthase) and its specific molecular chaperone cosmc (C1GalT1C1). In Handbook of Glycosyltransferases and Related Genes, 2nd ed.; Springer: Tokyo, Japan, 2014; Volume 1, pp. 149–169. ISBN 9784431542407. [Google Scholar]
- Togayachi, A.; Narimatsu, H. UDP-GlcNAc: Beta-gal beta1,3-NAcetylglucosaminyltransferase 6 (B3GNT6) (core 3 synthase, C3GnT). In Handbook of Glycosyltransferases and Related Genes, 2nd ed.; Springer: Tokyo, Japan, 2014; Volume 1, pp. 321–330. ISBN 9784431542407. [Google Scholar]
- Yeh, J.C.G.; Fukuda, M. Beta-1,3-galactosyl-O-glycosyl- glycoprotein beta-1,6-NAcetylglucosaminyltransferase 3 (GCNT3). In Handbook of Glycosyltransferases and Related Genes, 2nd ed.; Springer: Tokyo, Japan, 2014; Volume 1, pp. 367–377. ISBN 9784431542407. [Google Scholar]
- Tsuboi, S.; Fukuda, M. Beta-1,3-galactosyl-O-glycosyl-glycoprotein beta-1,6-NAcetylglucosaminyltransferase 1 (GCNT1) (C2GnT-L) and beta-1,3-galactosyl-oglycosyl-glycoprotein beta-1,6-NAcetylglucosaminyltransferase 3 (GCNT4) (C2GnT-T). In Handbook of Glycosyltransferases and Related Genes, 2nd ed.; Springer: Tokyo, Japan, 2014; Volume 1, pp. 355–366. ISBN 9784431542407. [Google Scholar]
- KEGG: Kyoto Encyclopedia of Genes and Genomes. Available online: https://www.genome.jp/kegg/ (accessed on 10 January 2020).
- James, M.O. Enzyme Kinetics of Conjugating Enzymes: PAPS Sulfotransferase. Methods Mol. Biol. 2014, 1113, 187–201. [Google Scholar] [CrossRef]
- Axelsson, M.A.B.; Karlsson, N.G.; Steel, D.M.; Ouwendijk, J.; Nilsson, T.; Hansson, G.C. Neutralization of pH in the Golgi apparatus causes redistribution of glycosyltransferases and changes in the O-glycosylation of mucins. Glycobiology 2001, 11, 633–644. [Google Scholar] [CrossRef]
- Kellokumpu, S. Golgi pH, ion and redox homeostasis: How much do they really matter? Front. Cell Dev. Biol. 2019, 7, 93. [Google Scholar] [CrossRef]
- Khoder-Agha, F.; Sosicka, P.; Escriva Conde, M.; Hassinen, A.; Glumoff, T.; Olczak, M.; Kellokumpu, S. N-acetylglucosaminyltransferases and nucleotide sugar transporters form multi-enzyme–multi-transporter assemblies in golgi membranes in vivo. Cell. Mol. Life Sci. 2019, 76, 1821–1832. [Google Scholar] [CrossRef]
- Akase, S. Simulation Analysis and Web Tool Development for Glycan Synthesis Prediction; Graduate School of Engineering, Soka University: Tokyo, Japan, 2020. [Google Scholar]
- GlycoSim. Available online: https://rings.glycoinfo.org/glycosims/index (accessed on 10 January 2020).
- Hoops, S.; Sahle, S.; Gauges, R.; Lee, C.; Pahle, J.; Simus, N.; Singhal, M.; Xu, L.; Mendes, P.; Kummer, U. COPASI—A COmplex PAthway SImulator. Bioinformatics 2006, 22, 3067–3074. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
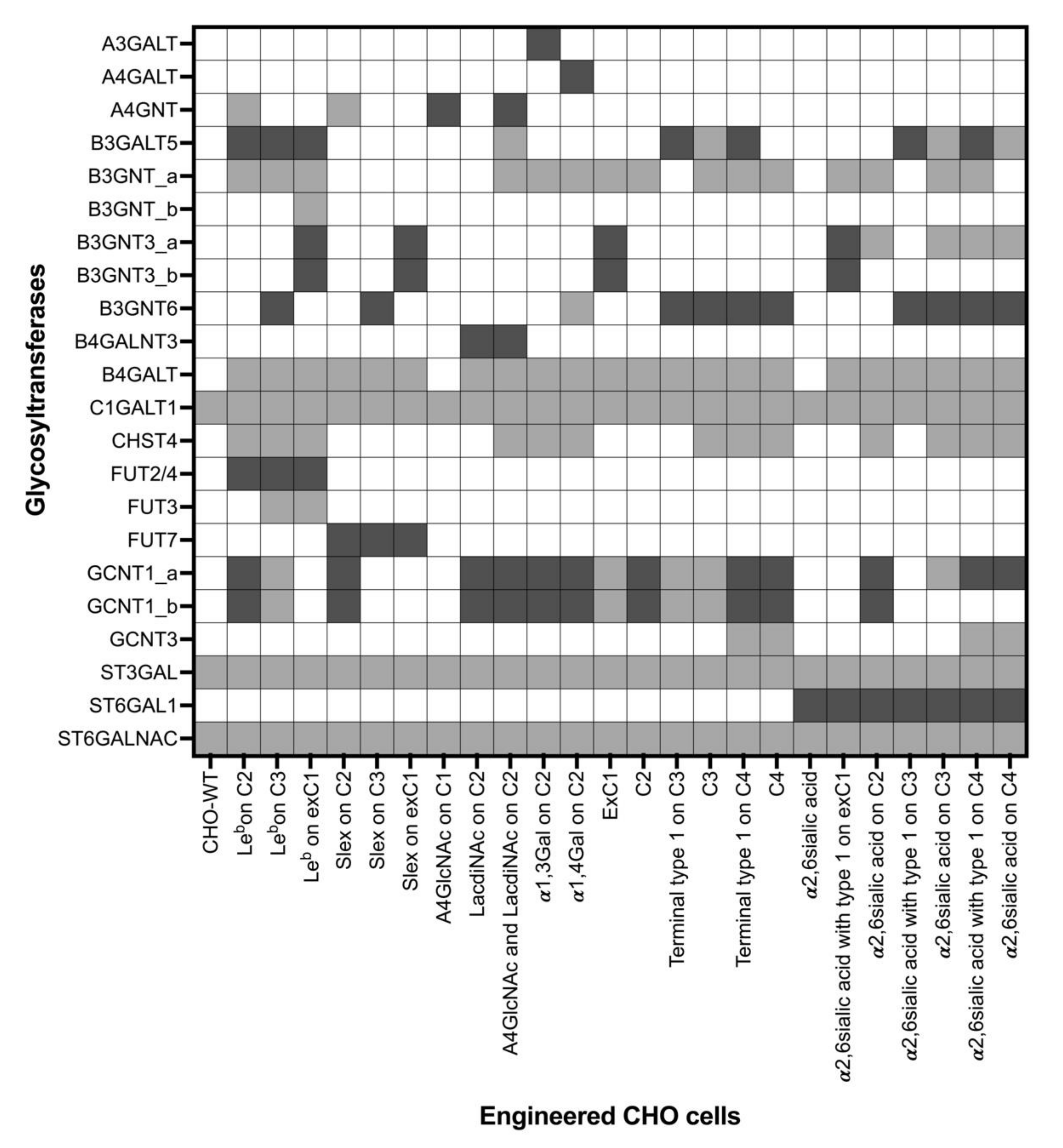
| Engineered CHO Cells’ Names | Transfected Glycosyltransferases | Reference |
|---|---|---|
| CHO/CHO-WT | - | [11,14] |
| Leb on C2 | GCNT1, B3GalT5, FUT2, FUT4 | [15,16], Unpublished data |
| Leb on C3 | B3GNT6, B3GalT5, FUT2, FUT4 | [15,16], Unpublished data |
| Leb on exC1 | B3GNT3, B3GalT5, FUT2, FUT4 | [15,16], Unpublished data |
| Slex on C2 | GCNT1, FUT7 | [15], Unpublished data |
| Slex on C3 | B3GNT6, FUT7 | [15], Unpublished data |
| Slex on exC1 | B3GNT3, FUT7 | [15], Unpublished data |
| A4GlcNAc on C1 | A4GNT | Unpublished data |
| LacdiNAc on C2 | GCNT1, B4GALNT | [17] |
| A4GlcNAc and LacdiNAc on C2 | GCNT1, A4GNT, B4GALNT | Unpublished data |
| α1,3Gal on C2 | GCNT1, A3GALT | [18] |
| α1,4Gal on C2 | GCNT1, A4GALT | [18] |
| ExC1 | B3GNT3 | [11] |
| C2 | GCNT1 | [11,19] |
| Terminal type 1 on C3 | B3GNT6, B3GalT5 | [12,16] |
| Terminal type 2 on C3 | B3GNT6 | [11,16] |
| Terminal type 1 on C4 | GCNT1, B3GNT6, B3GalT5 | [12,16] |
| C4 | GCNT1, B3GNT6 | [12] |
| α2,6sialic acid | ST6GAL1 | [11,12] |
| α2,6sialic acid with type 1 on exC1 | ST6GAL1, B3GNT3 | [11,12] |
| α2,6sialic acid on C2 | ST6GAL1, GCNT1 | [11,12] |
| α2,6sialic acid with type 1 on C3 | ST6GAL1, B3GNT6, B3GALT5 | [12,16] |
| α2,6sialic acid on C3 | ST6GAL1, B3GNT6 | [11,12] |
| α2,6sialic acid with type 1 on C4 | ST6GAL1, B3GNT3, GCNT1, B3GALT5 | [12,16] |
| α2,6sialic acid on C4 | ST6GAL1, B3GNT3, GCNT1 | [12] |
| Abbreviation | Glycosyltransferase | EC Number | Cores or Terminal Epitopes |
|---|---|---|---|
| A3GALT * | Alpha-1,3-galactosyltransferase | 2.4.1.- | αGal |
| A4GALT * | Alpha-1,4-galactosyltransferase | 2.4.1.228 | P1 antigen |
| A4GNT * | Alpha-1,4-N-Acetylglucosaminyltransferase | 2.4.1.- | α1,4GlcNAc |
| B3GALT5 * | Beta-1,3-galactosyltransferase 5 | 2.4.1.- | Type 1 chain |
| B3GNT | Beta-1,3-N-Acetylglucosaminyltransferase | 2.4.1.149 | |
| B3GNT3 * | Beta-1,3-N-Acetylglucosaminyltransferase 3 | 2.4.1.146 | Extended core 1 |
| B3GNT6 * | Beta-1,3-N-Acetylglucosaminyltransferase 6 | 2.4.1.147 | Core 3 |
| B4GALNT3 * | Beta-1,4-N-acetylgalactosaminyltransferase 3 | 2.4.1.244 | LacdiNAc |
| B4GALT | BetBeta-1,4-galactosyltransferase | 2.4.1.38 | Type 2 chain |
| C1GALT1 | Glycoprotein-N-acetylgalactosamine beta-1,3-galactosyltransferase | 2.4.1.122 | Core 1 |
| CHST4 | Carbohydrate sulfotransferase 4 | 2.8.2.- | 6SGlcNAc |
| FUT2 * | Fucosyltransferase 2 | 2.4.1.- | α1,2Fuc |
| FUT3 | Fucosyltransferase 3 | 2.4.1.65 | α1,3/4Fuc |
| FUT4 * | Fucosyltransferase 4 | 2.4.1. | α1,3Fuc |
| FUT7 * | Fucosyltransferase 7 | 2.4.1. | α1,3Fuc |
| GCNT1 * | Beta-1,6-N-acetylglucosaminyltransferase 1 | 2.4.1.102 | Core 2/4 |
| GCNT3 | Beta-1,6-N-acetylglucosaminyltransferase 3 | 2.4.1.148 | Core 2/4 |
| ST3GAL | Beta-galactoside alpha-2,3-sialyltransferase | 2.4.99.4, 2.4.99.6 | α2,3Sia |
| ST6GAL1 * | Beta-galactoside alpha-2,6-sialyltransferase 1 | 2.4.99.1 | α2,6Sia |
| ST6GALNAC | ST6 N-acetylgalactosaminide alpha-2,6-sialyltransferase | 2.4.99.3 | α2,6Sia |
| Donor | cis | Medial | Trans | TGN |
|---|---|---|---|---|
| CMP_NeuAc | 0 | 0 | 3000 | 3000 |
| CMP_NeuGc | 0 | 0 | 3000 | 3000 |
| GDP_Fuc | 0 | 5000 | 5000 | 0 |
| UDP_GlcNAc | 9143 | 9143 | 9143 | 0 |
| UDP_Gal | 3810 | 0 | 3810 | 3810 |
| UDP_GalNAc | 0 | 0 | 3000 | 0 |
| PAP_S | 0 | 920 | 920 | 920 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kouka, T.; Akase, S.; Sogabe, I.; Jin, C.; Karlsson, N.G.; Aoki-Kinoshita, K.F. Computational Modeling of O-Linked Glycan Biosynthesis in CHO Cells. Molecules 2022, 27, 1766. https://doi.org/10.3390/molecules27061766
Kouka T, Akase S, Sogabe I, Jin C, Karlsson NG, Aoki-Kinoshita KF. Computational Modeling of O-Linked Glycan Biosynthesis in CHO Cells. Molecules. 2022; 27(6):1766. https://doi.org/10.3390/molecules27061766
Chicago/Turabian StyleKouka, Thukaa, Sachiko Akase, Isami Sogabe, Chunsheng Jin, Niclas G. Karlsson, and Kiyoko F. Aoki-Kinoshita. 2022. "Computational Modeling of O-Linked Glycan Biosynthesis in CHO Cells" Molecules 27, no. 6: 1766. https://doi.org/10.3390/molecules27061766
APA StyleKouka, T., Akase, S., Sogabe, I., Jin, C., Karlsson, N. G., & Aoki-Kinoshita, K. F. (2022). Computational Modeling of O-Linked Glycan Biosynthesis in CHO Cells. Molecules, 27(6), 1766. https://doi.org/10.3390/molecules27061766