
Production, Bioprocessing and Anti-Proliferative Activity of Camptothecin from Penicillium chrysogenum, “An Endozoic of Marine Sponge, Cliona sp.”, as a Metabolically Stable Camptothecin Producing Isolate


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Marine Sponges, Isolation and Morphological Identification of the Endozoic Fungi
2.2. Screening and Chromatographic Analyses of Camptothecin from the Potent Fungi
2.3. UV-Vis, FT-IR, and LC-MS Analyses
2.4. Molecular Identification of the Recovered Endozoic Fungi
2.5. Bioprocess Optimization of Camptothecin Production by the Potent Fungal Isolate with Plackett-Burman and Faced Central Composite Designs (FCCD)
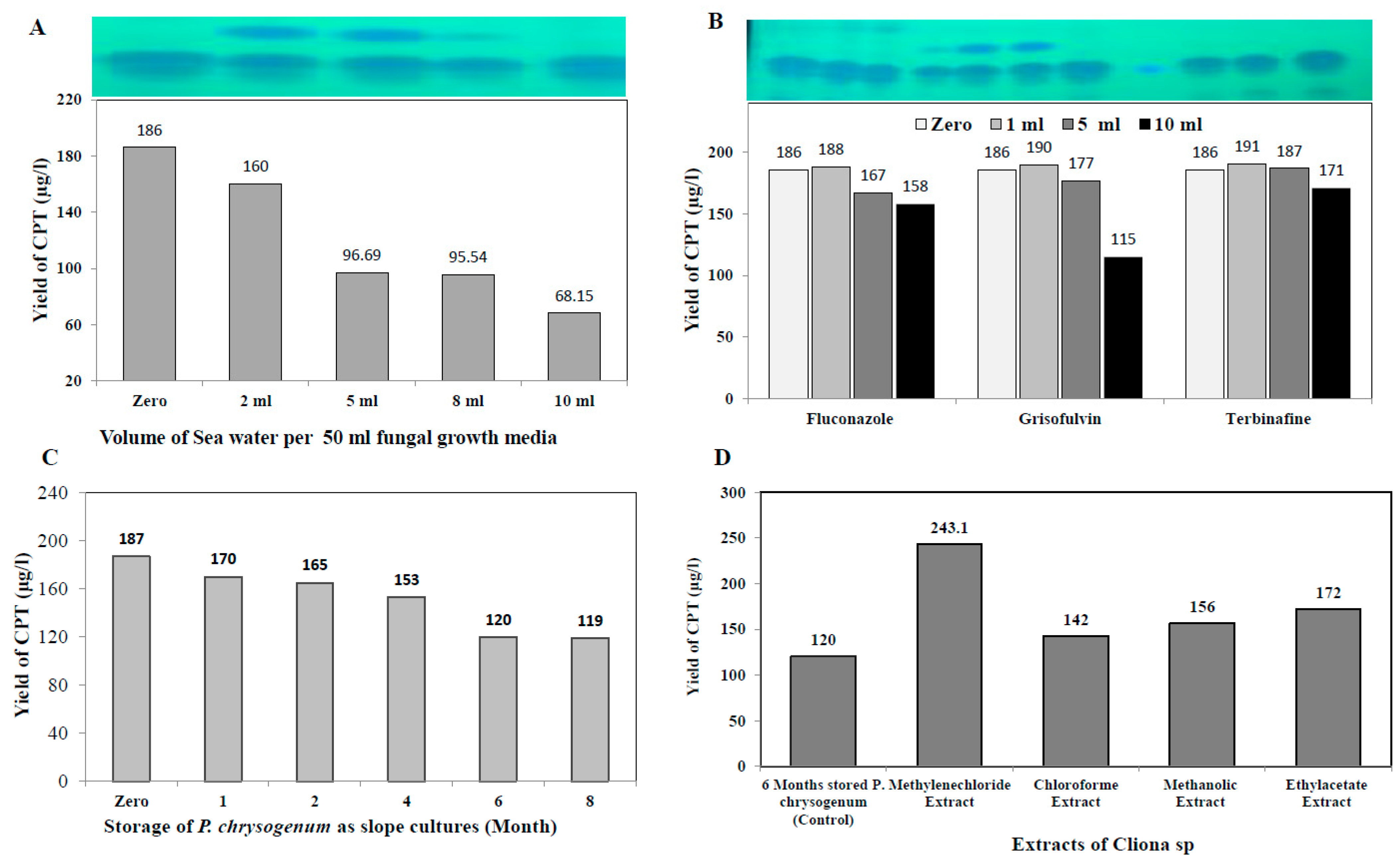
2.6. Effect of Fungal Growth Inhibitors/Elicitors on Camptothecin Production by the Potent Fungal Isolate
2.7. Stability of Camptothecin Productivity during Fungal Storage, and Effect of Sponge Extracts on Restoring Biosynthetic Machinery
2.8. Antiproliferative Activity of the Purified Camptothecin from Penicillium Chrysogenum
2.9. Internal Transcriped Spaer Fungal Deposition
2.10. Statistical Analysis
3. Results
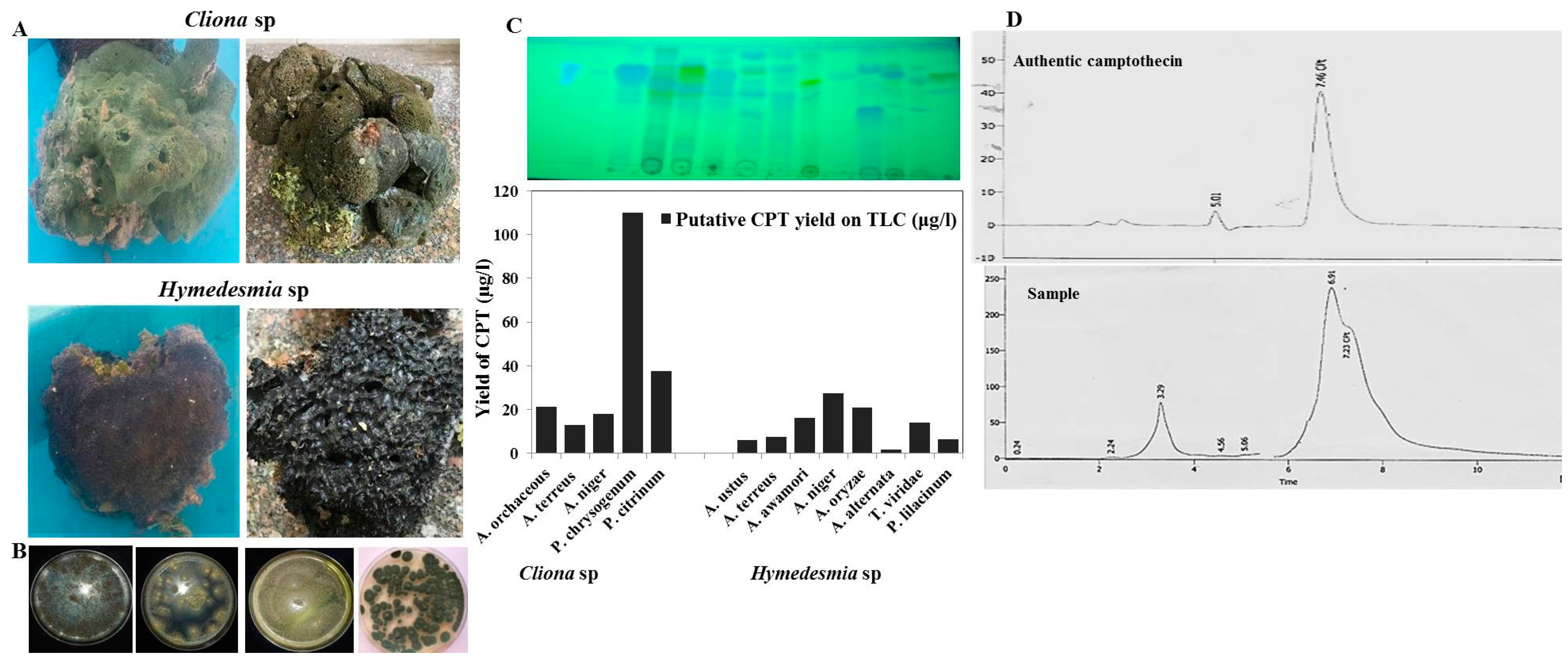
3.1. Isolation, and Screening for Camptothecin from Marine Sponges-Derived Fungi
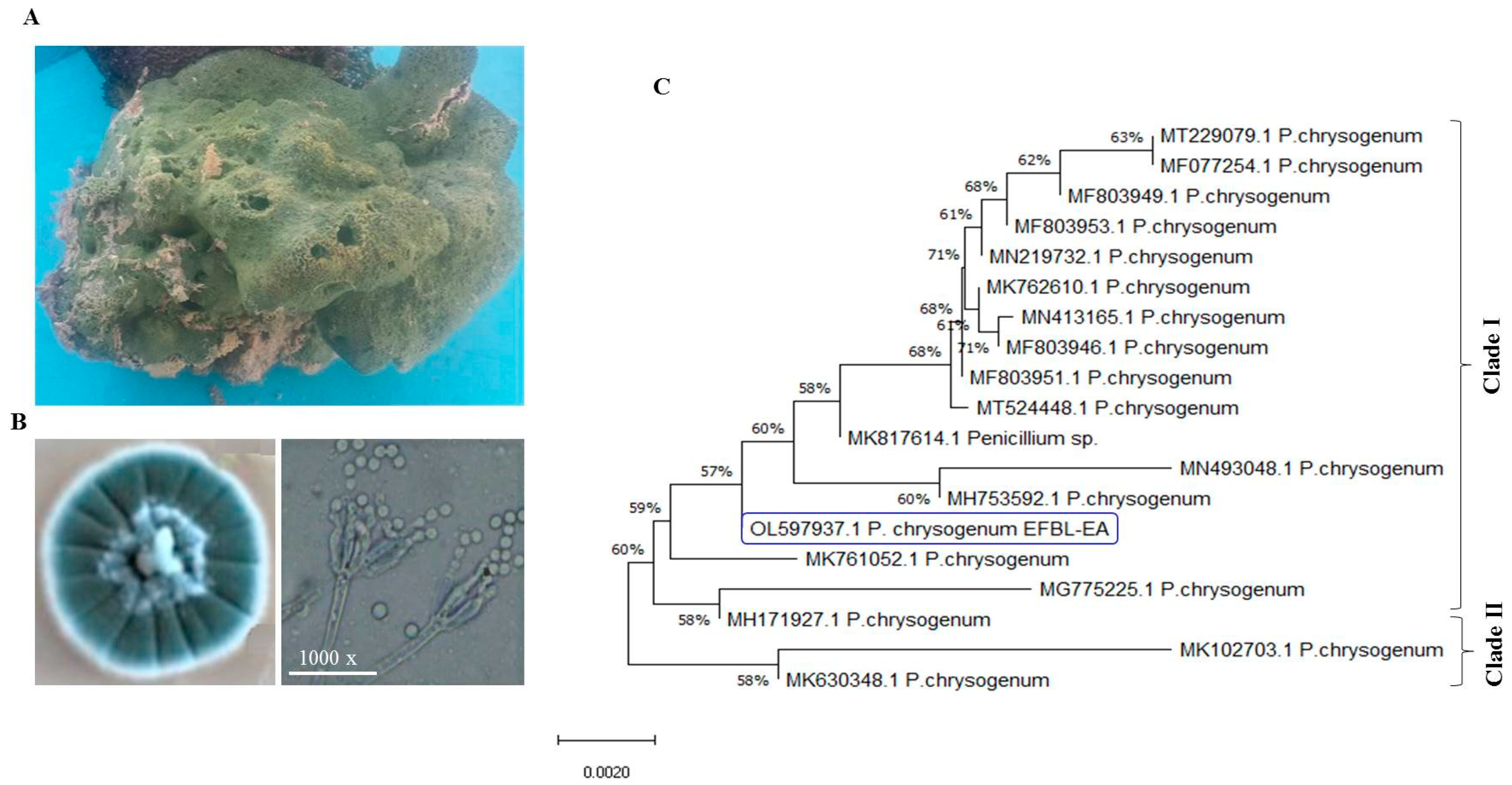
3.2. Molecular Identification of Potent Camptothecin Producing Fungi
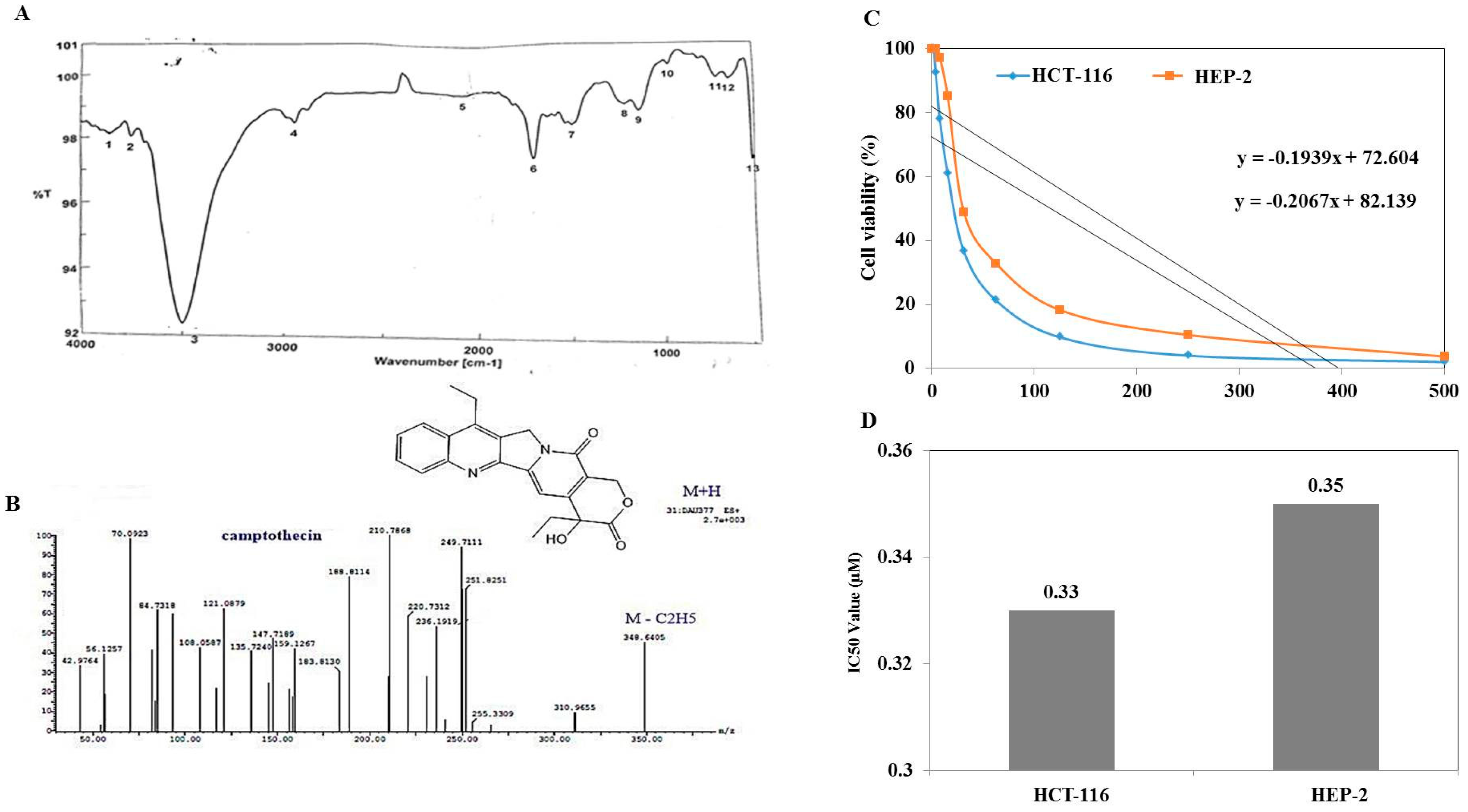
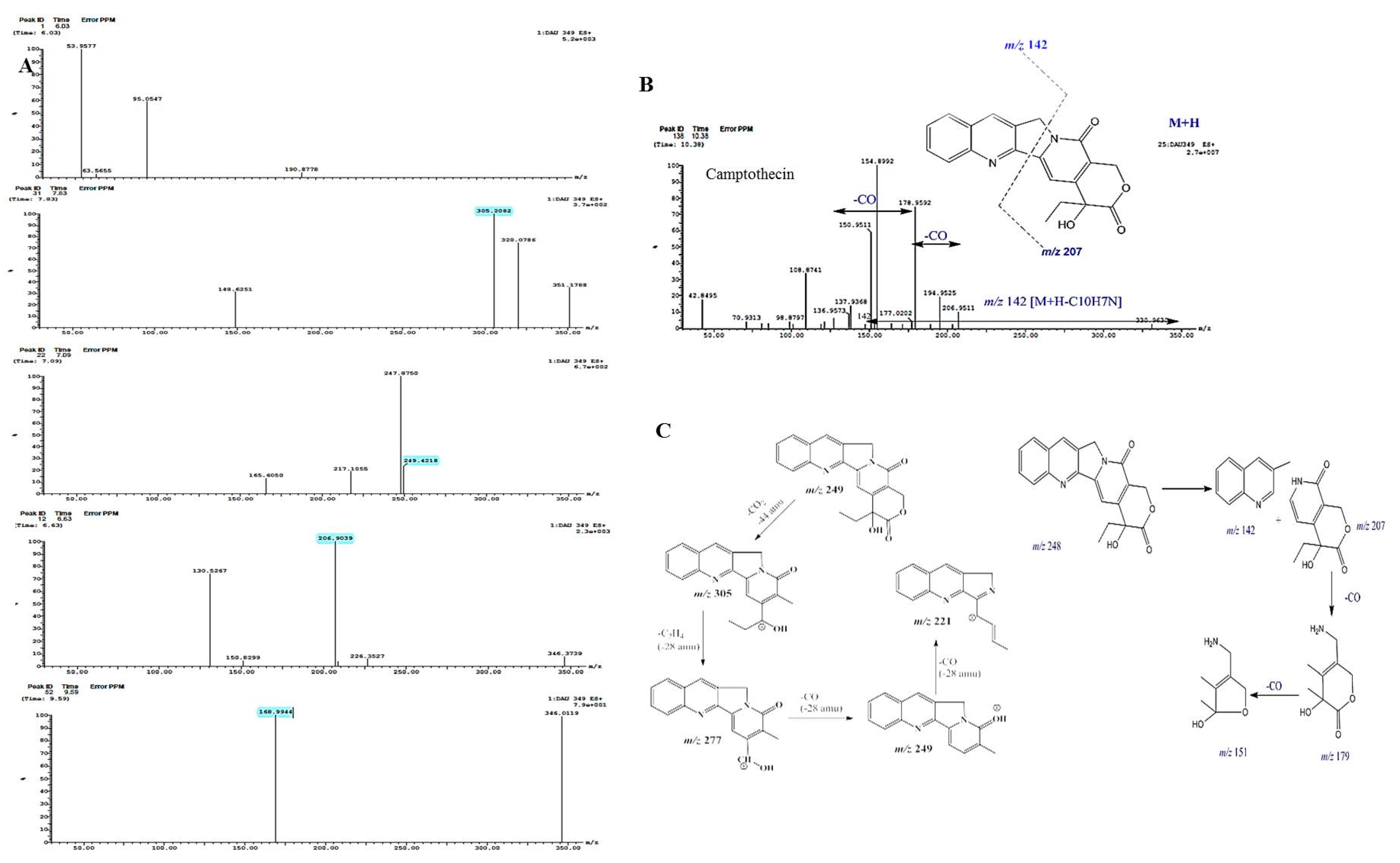
3.3. Chromatographic and Spectroscopic Analyses, and Antiproliferative Activity of Extracted Camptothecin
3.4. Bioprocess Optimization of Camptothecin Production by P. chrysogenum with Plackett-Burman Design
3.5. Effect of Fungal Growth Inhibitors on Camptothecin Yield by P. chrysogenum
3.6. Biosynthetic Stability of Camptothecin by P. chrysogenum in Response to Storage, and Effect of Sponge Extracts on Restoring Its Biosynthetic Machinery
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Wall, M.E.; Wani, M.C.; Cook, C.E.; Palmer, K.H.; McPhail, A.T.; Sim, G.A. Plant Antitumor Agents. I. The Isolation and Structure of Camptothecin, a Novel Alkaloidal Leukemia and Tumor Inhibitor from Camptotheca acuminata1,2. J. Am. Chem. Soc. 1966, 88, 3888–3890. [Google Scholar] [CrossRef]
- Liu, Y.Q.; Li, W.Q.; Morris-Natschke, S.L. Perspectives on biologically active camptothecin derivatives. Med. Res. Rev. 2015, 35, 753–789. [Google Scholar]
- Yamazaki, Y.; Kitajima, M.; Arita, M.; Takayama, H.; Sudo, H.; Yamazaki, M.; Aimi, N.; Saito, K. Biosynthesis of Camptothecin. In Silico and in Vivo Tracer Study from [1-13C]Glucose. Plant Physiol. 2004, 134, 161–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puri, S.C.; Handa, G.; Bhat, B.A.; Gupta, V.K.; Amna, T.; Verma, N.; Anand, R.; Dhar, K.L.; Qazi, G.N. Separation of 9-methoxycamptothecin and camptothecin from Nothapodytes foetida by semipreparative HPLC. J. Chromatogr. Sci. 2005, 43, 348–350. [Google Scholar] [CrossRef] [PubMed]
- Lorence, A.; Medina-Bolivar, F.; Nessler, C.L. Camptothecin and 10-hydroxycamptothecin from Camptotheca acuminata hairy roots. Plant Cell Rep. 2004, 22, 437–441. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Meyer, M.; Nessler, C.L.; McKnight, T.D. Sites of accumulation of the antitumor alkaloid camptothecin in Camptotheca acuminata. Planta Med. 1994, 60, 558–560. [Google Scholar] [CrossRef] [PubMed]
- Chavan, S.P.; Venkatraman, M.S. Synthesis of (+)-camptothecin. Arch. Org. Chem. 2005, 2005, 165–169. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, A.S.; Shindia, A.A. Characterization and immobilization of purified Aspergillus flavipes L-methioninase: Continuous production of methanethiol. J. Appl. Microbiol. 2011, 111, 54–69. [Google Scholar] [CrossRef]
- Staker, B.L.; Hjerrild, K.; Feese, M.D.; Behnke, C.A.; Burgin, A.B.; Stewart, L. The mechanism of topoisomerase I poisoning by a camptothecin analog. Proc. Natl. Acad. Sci. USA 2002, 99, 15387–15392. [Google Scholar] [CrossRef] [Green Version]
- Matsumoto, Y.; Fujiwara, T.; Honjo, Y.; Sasaoka, N.; Tsuchida, T.; Nagao, S. Quantitative analysis of DNA topoisomerase I activity in human and rat glioma: Characterization and mechanism of resistance to antitopoisomerase chemical, camptothecin-11. J. Surg. Oncol. 1993, 53, 97–103. [Google Scholar] [CrossRef]
- Benderoth, M.; Textor, S.; Windsor, A.J.; Mitchell-Olds, T.; Gershenzon, J.; Kroymann, J. Positive selection driving diversification in plant secondary metabolism. Proc. Natl. Acad. Sci. USA 2006, 103, 9118–9123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takimoto, C.H.; Wright, J.; Arbuck, S.G. Clinical applications of the camptothecins. BBA-Gene. Struct. Expr. 1998, 1400, 107–119. [Google Scholar] [CrossRef]
- Del Poeta, M.; Chen, S.F.; Von Hoff, D.; Dykstra, C.C.; Wani, M.C.; Manikumar, G.; Heitman, J.; Wall, M.E.; Perfect, J.R. Comparison of in vitro activities of camptothecin and nitidine derivatives against fungal and cancer cells. Antimicrob. Agents Chemother. 1999, 43, 2862–2868. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhalkar, B.N.; Patil, S.M.; Govindwar, S.P. Camptothecine production by mixed fermentation of two endophytic fungi from Nothapodytes nimmoniana. Fungal Biol. 2016, 120, 873–883. [Google Scholar] [CrossRef] [PubMed]
- Puri, S.G.; Verma, V.; Amna, T.; Qazi, G.N.; Spiteller, M. An endophytic fungus from Nothapodytes foetida that produces camptothecin. J. Nat. Prod. 2005, 68, 1717–1719. [Google Scholar] [CrossRef]
- Upadhya, V.; Pai, S.R.; Sharma, A.K.; Hegde, H.V.; Kholkute, S.D.; Joshi, R.K. Compound specific extraction of camptothecin from nothapodytes nimmoniana and piperine from piper nigrum using accelerated solvent extractor. J. Anal. Methods Chem. 2014, 2014, 932036. [Google Scholar]
- Li, S.; Yi, Y.; Wang, Y.; Zhang, Z.; Beasley, R.S. Camptothecin accumulation and variations in Camptotheca. Planta Med. 2002, 68, 1010–1016. [Google Scholar] [CrossRef]
- Sirikantaramas, S.; Asano, T.; Sudo, H.; Yamazaki, M.; Saito, K. Camptothecin: Therapeutic Potential and Biotechnology. Curr. Pharm. Biotechnol. 2007, 8, 196–202. [Google Scholar] [CrossRef]
- Pu, X.; Chen, F.; Yang, Y.; Qu, X.; Zhang, G.; Luo, Y. Isolation and characterization of Paenibacillus polymyxa LY214, a camptothecin-producing endophytic bacterium from Camptotheca acuminata. J. Ind. Microbiol. Biot. 2015, 42, 1197–1202. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Yassin, M.A.; Ibrahim, H. Coimmobilization of l -methioninase and glutamate dehydrogenase: Novel approach for l -homoalanine synthesis. Biotechnol. Appl. Biochem. 2015, 62, 514–522. [Google Scholar] [CrossRef]
- Grudatt, P.S.; Priti, V.; Shweta, S.; Ramesha, B.T.; Ravikanth, G.; Vasudeva, R.; Amna, T.; Deepika, S.; Ganeshaiah, K.N.; Uma Shaanker, R.; et al. Attenuation of camptothecin production and negative relation between hyphal biomass and camptothecin content in endophytic fungal strains isolated from Nothapodytes nimmoniana Grahm (Icacinaceae). Curr. Sci. 2010, 98, 1006–1010. [Google Scholar]
- El-Sayed, A.S.A.; Khalaf, S.A.; Azez, H.A.; Hussein, H.A.; EL-Moslamy, S.H.; Sitohy, B.; El-Baz, A.F. Production, bioprocess optimization and anticancer activity of Camptothecin from Aspergillus terreus and Aspergillus flavus, endophytes of Ficus elastica. Process Biochem. 2021, 107, 59–73. [Google Scholar] [CrossRef]
- Amna, T.; Puri, S.C.; Verma, V.; Sharma, J.P.; Khajuria, R.K.; Musarrat, J.; Spiteller, M.; Qazi, G.N. Bioreactor studies on the endophytic fungus Entrophospora infrequens for the production of an anticancer alkaloid camptothecin. Can. J. Microbiol. 2006, 52, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Isah, T.; Mujib, A. Camptothecin from Nothapodytes nimmoniana: Review on biotechnology applications. Acta Physiol. Plant. 2015, 37, 1–14. [Google Scholar]
- Venugopalan, A.; Srivastava, S. Enhanced camptothecin production by ethanol addition in the suspension culture of the endophyte, Fusarium solani. Bioresour. Technol. 2015, 188, 251–257. [Google Scholar] [CrossRef]
- Ran, X.; Zhang, G.; Li, S.; Wang, J. Characterization and antitumor activity of camptothecin from endophytic fungus Fusarium solani isolated from Camptotheca acuminate. Afr. Health Sci. 2017, 17, 566–574. [Google Scholar] [CrossRef] [Green Version]
- Shweta, S.; Bindu, J.H.; Raghu, J.; Suma, H.K.; Manjunatha, B.L.; Kumara, P.M.; Ravikanth, G.; Nataraja, K.N.; Ganeshaiah, K.N.; Uma Shaanker, R. Isolation of endophytic bacteria producing the anti-cancer alkaloid camptothecine from Miquelia dentata Bedd. (Icacinaceae). Phytomedicine 2013, 20, 913–917. [Google Scholar] [CrossRef]
- Bhalkar, B.N.; Bedekar, P.A.; Patil, S.M.; Patil, S.A.; Govindwar, S.P. Production of camptothecine using whey by an endophytic fungus: Standardization using response surface methodology. RSC Adv. 2015, 5, 620150122. [Google Scholar] [CrossRef]
- Kusari, S.; Lamshöft, M.; Spiteller, M. Aspergillus fumigatus Fresenius, an endophytic fungus from Juniperus communis L. Horstmann as a novel source of the anticancer pro-drug deoxypodophyllotoxin. J. Appl. Microbiol. 2009, 107, 1019–1030. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Hassan, A.E.A.; Shindia, A.A.; Mohamed, S.G.; Sitohy, M.Z. Aspergillus flavipes methionine γ-lyase-dextran conjugates with enhanced structural, proteolytic stability and anticancer efficiency (2016). J. Mol. Catal. B-Enzym. 2016, 133, S15–S24. [Google Scholar] [CrossRef]
- El-Kalyoubi, S.; Agili, F.; Zordok, W.A.; El-Sayed, A.S.A. Synthesis, in silico prediction and in vitro evaluation of antimicrobial activity, dft calculation and theoretical investigation of novel xanthines and uracil containing imidazolone derivatives. Int. J. Mol. Sci. 2021, 22, 10979. [Google Scholar] [CrossRef] [PubMed]
- Kusari, S.; Hertweck, C.; Spiteller, M. Chemical ecology of endophytic fungi: Origins of secondary metabolites. Chem. Bio. 2012, 19, 792–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Sayed, A.S.A.; Shindia, A.A.; Ali, G.S.; Yassin, M.A.; Hussein, H.; Awad, S.A.; Ammar, H.A. Production and bioprocess optimization of antitumor Epothilone B analogue from Aspergillus fumigatus, endophyte of Catharanthus roseus, with response surface methodology. Enzym. Microb. Technol. 2021, 143, 109718. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, S.K.; Prakash, V.; Ranjan, N. Marine fungi: A source of potential anticancer compounds. Front. Microbiol. 2018, 8, 2536. [Google Scholar] [CrossRef] [PubMed]
- Butler, M.S.; Robertson, A.A.B.; Cooper, M.A. Natural product and natural product derived drugs in clinical trials. Nat. Prod. Rep. 2014, 31, 1612–1661. [Google Scholar] [PubMed]
- El-Sayed, A.S.; Khalaf, S.A.; Abdel-Hamid, G.; El-Batrik, M.I. Screening, morphological and molecular characterization of fungi producing cystathionine γ-lyase. Acta Biol. Hung. 2015, 66, 119–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sithranga Boopathy, N.; Kathiresan, K. Anticancer drugs from marine flora: An overview. J. Oncol. 2010, 2020, 214186. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, A.S.A.; Ali, G.S. Aspergillus flavipes is a novel efficient biocontrol agent of Phytophthora parasitica. Biol. Cont. 2020, 140, 104072. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; George, N.M.; Yassin, M.A.; Alaidaroos, B.A.; Bolbol, A.A.; Mohamed, M.S.; Rady, A.M.; Aziz, S.W.; Zayed, R.A.; Sitohy, M.Z. Purification and Characterization of Ornithine Decarboxylase from Aspergillus terreus; Kinetics of Inhibition by Various Inhibitors. Molecules 2019, 24, 2756. [Google Scholar] [CrossRef] [Green Version]
- Patel, J.S.; Vitoreli, A.; Palmateer, A.J.; El-Sayed, A.; Norman, D.J.; Goss, E.M.; Brennan, M.S.; Ali, G.S. Characterization of Phytophthora spp. Isolated from ornamental plants in Florida. Plant. Dis. 2016, 100, 500–509. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, A.S.A.; Shindia, A.A.; AbouZeid, A.; Koura, A.; Hassanein, S.E.; Ahmed, R.M. Triggering the biosynthetic machinery of Taxol by Aspergillus flavipes via cocultivation with Bacillus subtilis: Proteomic analyses emphasize the chromatin remodeling upon fungal-bacterial interaction. Environ. Sci. Pollut. Res. 2021, 28, 39866–39881. [Google Scholar]
- El-Sayed, A.S.A.; Ali, D.M.I.; Yassin, M.A.; Zayed, R.A.; Ali, G.S. Sterol inhibitor “Fluconazole” enhance the Taxol yield and molecular expression of its encoding genes cluster from Aspergillus flavipes. Process Biochem. 2019, 76, 55–67. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Akbar, A.; Iqrar, I.; Ali, R.; Norman, D.; Brennan, M.; Ali, G.S. A glucanolytic Pseudomonas sp. associated with Smilax bona-nox L. displays strong activity against Phytophthora parasitica. Microbiol. Res. 2018, 207, 140–152. [Google Scholar] [PubMed]
- El-Sayed, A.S.; Yassin, M.A.; Khalaf, S.A.; El-Batrik, M.; Ali, G.S.; Esener, S. Biochemical and Pharmacokinetic Properties of PEGylated Cystathionine γ-Lyase from Aspergillus carneus KF723837. J. Mol. Microbiol. Biotechnol. 2015, 25, 301–310. [Google Scholar] [CrossRef]
- Shaw, C.G. The Genus Aspergillus. Kenneth, B. Raper and Dorothy, I. Fennell. With a chapter by Peter, K.C. Austwick. Williams and Wilkins, Baltimore. Science 1965, 150, 736–737. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Samson, R.A. Polyphasic taxonomy of Penicillium subgenus Penicillium: A guide to identification of food and air-borne terverticillate Penicillia and their mycotoxins. Stud. Mycol. 2004, 2004, 1–173. [Google Scholar]
- Chen, A.J.; Frisvad, J.C.; Sun, B.D.; Varga, J.; Kocsubé, S.; Dijksterhuis, J.; Kim, D.H.; Hong, S.B.; Houbraken, J.; Samson, R.A. Aspergillus section Nidulantes (formerly Emericella): Polyphasic taxonomy, chemistry and biology. Stud. Mycol. 2016, 84, 1–118. [Google Scholar] [CrossRef] [Green Version]
- El Sayed, M.T.; El-Sayed, A.S.A. Biocidal activity of metal nanoparticles synthesized by Fusarium solani against multidrug-resistant bacteria and mycotoxigenic fungi. J. Microbiol. Biotechnol. 2020, 30, 226–236. [Google Scholar] [CrossRef]
- Visagie, C.M.; Houbraken, J.; Frisvad, J.C.; Hong, S.B.; Klaassen, C.H.W.; Perrone, G.; Seifert, K.A.; Varga, J.; Yaguchi, T.; Samson, R.A. Identification and nomenclature of the genus Penicillium. Stud. Mycol. 2014, 78, 343–371. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Fatah, S.S.; El-Batal, A.I.; El-Sherbiny, G.M.; Khalaf, M.A.; El-Sayed, A.S. Production, bioprocess optimization and γ-irradiation of Penicillium polonicum, as a new Taxol producing endophyte from Ginko biloba. Biotech. Rep. 2021, 30, e00623. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Shindia, A.A.; AbouZaid, A.A.; Yassin, A.M.; Ali, G.S.; Sitohy, M.Z. Biochemical characterization of peptidylarginine deiminase-like orthologs from thermotolerant Emericella dentata and Aspergillus nidulans. Enzym. Microb. Technol. 2019, 124, 41–53. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.S.A.; Shindia, A.A.; Zeid, A.A.A.; Yassin, A.M.; Sitohy, M.Z.; Sitohy, B. Aspergillus nidulans thermostable arginine deiminase-Dextran conjugates with enhanced molecular stability, proteolytic resistance, pharmacokinetic properties and anticancer activity. Enzym. Microb. Technol. 2019, 131, 109432. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.S.A.; Hassan, M.N.; Nada, H.M.S. Purification, immobilization, and biochemical characterization of L-arginine deiminase from thermophilic Aspergillus fumigatus KJ434941: Anticancer activity in vitro. Biotechnol. Prog. 2015, 31, 396–405. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Dudley, J.; Nei, M.; Kumar, S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 2007, 24, 1596–1599. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.A.; El-Sayed, A.; Akbar, A.; Mangravita-Novo, A.; Bibi, S.; Afzal, Z.; Norman, D.J.; Ali, G.S. A highly efficient ligation-independent cloning system for CRISPR/Cas9 based genome editing in plants. Plant Methods 2017, 13, 86. [Google Scholar] [CrossRef] [Green Version]
- Maamoun, H.S.; Rabie, G.H.; Shaker, I.; Alaidaroos, B.A.; Arabia, S. Biochemical and kinetic properties of tyrosinase from Aspergillus terreus and Penicillium copticola; Undecanoic acid is a potent enzyme inhibitor from Aspergillus flavus, endophyte of Moringa oleifera. Molecules 2020, 5, 1309. [Google Scholar]
- Abdella, A.; El-Baz, A.F.; Ibrahim, I.A.; Mahrous, E.E.; Yang, S.-T. Biotransformation of soy flour isoflavones by Aspergillus niger NRRL 3122 β-glucosidase enzyme. Nat. Prod. Res. 2018, 32, 2382–2391. [Google Scholar] [CrossRef]
- El-Naggar, N.E.A.; Moawad, H.; El-Shweihy, N.M.; El-Ewasy, S.M.; Elsehemy, I.A.; Abdelwahed, N.A.M. Process development for scale-up production of a therapeutic L-asparaginase by Streptomyces brollosae NEAE-115 from shake flasks to bioreactor. Sci. Rep. 2019, 9, 1–18. [Google Scholar]
- ElMekawy, A.; Hegab, H.M.; El-Baz, A.; Hudson, S.M. Kinetic properties and role of bacterial chitin deacetylase in the bioconversion of chitin to chitosan. Recent Pat. Biotechnol. 2013, 7, 234–241. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Abdel-Azeim, S.; Ibrahim, H.M.; Yassin, M.A.; Abdel-Ghany, S.E.; Esener, S.; Ali, G.S. Biochemical stability and molecular dynamic characterization of Aspergillus fumigatus cystathionine γ-lyase in response to various reaction effectors. Enzym. Microb. Technol. 2015, 81, 31–46. [Google Scholar] [CrossRef] [Green Version]
- Cory, A.H.; Owen, T.C.; Barltrop, J.A.; Cory, J.G. Use of an aqueous soluble tetrazolium/formazan assay for cell growth assays in culture. Cancer Commun. 1991, 3, 207–212. [Google Scholar] [CrossRef] [PubMed]
- El Baz, A.F.; Shetaia, Y.M.; Elkhouli, R.R. Kinetic behavior of Candida tropicalis during xylitol production using semi-synthetic and hydrolysate based media. Afr. J. Biotechnol. 2011, 10, 16617–16625. [Google Scholar] [CrossRef] [Green Version]
- Pitt, J.I. The genus Penicillium and its teleomorphic states Eupenicillium and Talaromyces; Academic Press: London, UK, 1979; ISBN 9780125577502. [Google Scholar]
- El-Sayed, A.S.A.; Mohamed, N.Z.; Safan, S.; Yassin, M.A.; Shaban, L.; Shindia, A.A.; Shad Ali, G.; Sitohy, M.Z. Restoring the Taxol biosynthetic machinery of Aspergillus terreus by Podocarpus gracilior Pilger microbiome, with retrieving the ribosome biogenesis proteins of WD40 superfamily. Sci. Rep. 2019, 9, 11534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desai, S.D.; Liu, L.F.; Vazquez-Abad, D.; D’Arpa, P. Ubiquitin-dependent destruction of topoisomerase I is stimulated by the antitumor drug camptothecin. J. Bio. Chem. 1997, 272, 24159–24164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wani, M.C.; Taylor, H.L.; Wall, M.E.; Coggon, P.; McPhail, A.T. Plant Antitumor Agents. VI. The Isolation and Structure of Taxol, a Novel Antileukemic and Antitumo Agent from Taxus bretvifolia. J. Am. Chem. Soc. 1971, 93, 2325–2327. [Google Scholar] [CrossRef]
- Guo, B.; Wang, Y.; Sun, X.; Tang, K. Bioactive natural products from endophytes: A review. Appl. Biochem. Micro. 2008, 44, 136–142. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Zayed, R.A.; El-Baz, A.F.; Ismaeil, W.M. Bioprocesses optimization and anticancer activity of camptothecin from Aspergillus flavus, an endophyte of in vitro cultured Astragalus fruticosus. Mol. Biol. Rep. 2022. [Google Scholar] [CrossRef]
- Hentschel, U.; Usher, K.M.; Taylor, M.W. Marine sponges as microbial fermenters. FEMS Microbiol. Ecol. 2006, 55, 167–177. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Wang, G. Diversity of fungal isolates from three Hawaiian marine sponges. Microbiol. Res. 2009, 164, 233–241. [Google Scholar] [CrossRef]
- Yarden, O. Fungal association with sessile marine invertebrates. Front. Microb. 2014, 5, 228. [Google Scholar] [CrossRef]
- Paz, Z.; Komon-Zelazowska, M.; Druzhinina, I.S.; Aveskamp, M.M.; Shnaiderman, A.; Aluma, Y.; Carmeli, S.; Ilan, M.; Yarden, O. Diversity and potential antifungal properties of fungi associated with a Mediterranean sponge. Fungal Divers. 2010, 42, 17–26. [Google Scholar] [CrossRef]
- El-Sayed, A.S.A.; Ruff, L.E.; Ghany, S.E.A.; Ali, G.S.; Esener, S. Molecular and Spectroscopic Characterization of Aspergillus flavipes and Pseudomonas putida L-Methionine γ-Lyase in Vitro. Appl. Biochem. Biot. 2017, 181, 1513–1532. [Google Scholar] [CrossRef] [PubMed]
- Namdeo, A.G.; Sharma, A. HPLC analysis of camptothecin content in various parts of Nothapodytes foetida collected on different periods. Asian Pac. J. Trop. Biomed. 2012, 2, 389–393. [Google Scholar] [CrossRef] [Green Version]
- Kai, G.; Wu, C.; Gen, L.; Zhang, L.; Cui, L.; Ni, X. Biosynthesis and biotechnological production of anti-cancer drug Camptothecin. Phytochem. Rev. 2015, 14, 525–539. [Google Scholar] [CrossRef]
- Zhao, K.; Sun, L.; Ma, X.; Li, X.; Wang, X.; Ping, W.; Zhou, D. Improved taxol production in Nodulisporium sylviforme derived from inactivated protoplast fusion. Afr. J. Biotechnol. 2011, 10, 4175–4182. [Google Scholar] [CrossRef]
- Wen, Y.; Wang, Y.; Liu, X.; Zhang, W.; Xiong, X.; Han, Z.; Liang, X. Camptothecin-based nanodrug delivery systems. Cancer Bio. Med. 2017, 14, 363–370. [Google Scholar]
- El-Sayed, A.S.A.; Sayed, M.T.E.; Nada, H.S.; Hassan, A.E.; Yousef, E.K. Production and Characterization of Taxol as Anticancer Agent from Aspergillus Terreus. J. Pure Appl. Microbiol. 2019, 13, 2055–2063. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, A.S.A.; Abdel-Ghany, S.E.; Ali, G.S. Genome editing approaches: Manipulating of lovastatin and taxol synthesis of filamentous fungi by CRISPR/Cas9 system. Appl. Microbiol. Biot. 2017, 101, 3953–3976. [Google Scholar]
- López-Meyer, M.; Nessler, C.L. Tryptophan decarboxylase is encoded by two autonomously regulated genes in Camptotheca acuminata which are differentially expressed during development and stress. Plant J. 1997, 11, 1167–1175. [Google Scholar] [CrossRef]
- Rehman, S.; Shawl, A.S.; Kour, A.; Sultan, P.; Ahmad, K.; Khajuria, R.; Qazi, G.N. Comparative studies and identification of camptothecin produced by an endophyte at shake flask and bioreactor. Nat. Prod. Res. 2009, 23, 1050–1057. [Google Scholar] [CrossRef]
- Pu, X.; Qu, X.; Chen, F.; Bao, J.; Zhang, G.; Luo, Y. Camptothecin-producing endophytic fungus Trichoderma atroviride LY357: Isolation, identification, and fermentation conditions optimization for camptothecin production. Appl. Microbiol. Biot. 2013, 97, 9365–9375. [Google Scholar] [CrossRef] [PubMed]
- Sudo, H.; Yamakawa, T.; Yamazaki, M.; Aimi, N.; Saito, K. Bioreactor production of camptothecin by hairy root cultures of ophiorrhiza pumila. Biotechnol. Lett. 2002, 24, 359–363. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CPT Yield on TLC (μg/L) | |||
|---|---|---|---|
| Cliona sp. | 1 | A. orchaceous | 21.2 |
| 2 | A. terreus | 12.8 | |
| 3 | A. niger | 18.2 | |
| 4 | P. chrysogenum | 110.1 | |
| 5 | P. citrinum | 37.8 | |
| Hymedesmia sp. | 1 | A. ustus | 5.9 |
| 2 | A. terreus | 7.6 | |
| 3 | A. awamori | 16.3 | |
| 4 | A. niger | 27.4 | |
| 5 | A. oryzae | 21.1 | |
| 6 | A. alternata | 1.7 | |
| 7 | T. viridae | 14.2 | |
| 8 | P. lilacinum | 6.6 |
| Codes | Factors | Levels * | |
|---|---|---|---|
| −1 | 1 | ||
| X1 | Acid Whey | 2 | 5 |
| X2 | Malt Extract | 1 | 3 |
| X3 | Potato starch | 5 | 10 |
| X4 | Methyl jasmonate | 0.1 | 0.5 |
| X5 | Tryptamine | 1 | 2 |
| X6 | Peptone | 2 | 1 |
| X7 | Dextrin | 1 | 2 |
| X8 | Tryptone | 5 | 10 |
| X9 | Glucose | 5 | 10 |
| X10 | Salicylic acid | 1 | 2 |
| X11 | Tryptophan | 2 | 5 |
| X12 | Serine | 1 | 4 |
| X13 | Cysteine | 2 | 5 |
| X14 | Pyruvate | 2 | 5 |
| X15 | Phenylalanine | 2 | 4 |
| X16 | Glutamate | 5 | 10 |
| Run | X1 | X2 | X3 | X4 | X5 | X6 | X7 | X8 | X9 | X10 | X11 | X12 | X13 | X14 | X15 | X16 | CPT Yield (µg/L) | Predicted CPT yield (µg/L) | Residuals |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | −1 | −1 | 1 | −1 | 1 | −1 | 1 | 70.6 | 100.2 | −29.6 |
| 2 | −1 | −1 | −1 | 1 | −1 | 1 | −1 | 1 | 1 | 1 | 1 | −1 | −1 | 1 | 1 | −1 | 80.2 | 40.8 | 39.4 |
| 3 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | −1 | −1 | 1 | −1 | 1 | −1 | 1 | 1 | 93.6 | 120 | −26.4 |
| 4 | 1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | −1 | −1 | 1 | −1 | 1 | −1 | 92.7 | 126.9 | −34.2 |
| 5 | 1 | −1 | 1 | 1 | 1 | 1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | −1 | 80.3 | 90.0 | −9.7 |
| 6 | −1 | 1 | −1 | 1 | 1 | 1 | 1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | 81.2 | 98.0 | −16.8 |
| 7 | 1 | −1 | 1 | 1 | −1 | −1 | −1 | −1 | 1 | −1 | 1 | −1 | 1 | 1 | 1 | 1 | 52.1 | 87.9 | −35.8 |
| 8 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | −1 | −1 | 1 | −1 | 1 | −1 | 1 | 1 | 1 | 82.2 | 79.0 | 3.2 |
| 9 | 1 | −1 | −1 | −1 | −1 | 1 | −1 | 1 | −1 | 1 | 1 | 1 | 1 | −1 | −1 | 1 | 96.1 | 96.4 | −0.3 |
| 10 | −1 | 1 | −1 | 1 | −1 | 1 | 1 | 1 | 1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | 89.2 | 92.6 | −3.4 |
| 11 | 1 | 1 | 1 | 1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | −1 | −1 | 1 | 100.6 | 98.9 | 1.7 |
| 12 | 1 | 1 | 1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | −1 | −1 | 1 | −1 | 103.9 | 95.0 | 8.9 |
| 13 | −1 | 1 | 1 | −1 | −1 | −1 | −1 | 1 | −1 | 1 | −1 | 1 | 1 | 1 | 1 | −1 | 120.8 | 110.9 | 9.9 |
| 14 | 1 | 1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | −1 | −1 | 1 | −1 | 1 | 199.1 | 203.9 | −4.8 |
| 15 | −1 | −1 | −1 | −1 | 1 | −1 | 1 | −1 | 1 | 1 | 1 | 1 | −1 | −1 | 1 | 1 | 102.2 | 98.7 | 3.5 |
| 16 | 1 | −1 | 1 | −1 | 1 | 1 | 1 | 1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | 181.2 | 128.3 | 52.9 |
| 17 | −1 | 1 | 1 | 1 | 1 | −1 | −1 | 1 | 1 | −1 | 1 | 1 | −1 | −1 | −1 | −1 | 180.4 | 160.2 | 20.2 |
| 18 | −1 | −1 | 1 | −1 | 1 | −1 | 1 | 1 | 1 | 1 | −1 | −1 | 1 | 1 | −1 | 1 | 115.4 | 94.9 | 20.5 |
| 19 | 1 | 1 | −1 | −1 | −1 | −1 | 1 | −1 | 1 | −1 | 1 | 1 | 1 | 1 | −1 | −1 | 120.4 | 101.6 | 18.8 |
| 20 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | −1 | 125.5 | 109.0 | 16.5 |
| Source | Sum of Squares | df | Mean Square | F Value | p-Value Prob > F | |
|---|---|---|---|---|---|---|
| Model | 66586.05 | 7 | 9512.29 | 10.85 | 0.0002 | significant |
| D-Phenylalanine | 12872.74 | 1 | 12872.74 | 14.68 | 0.0024 | |
| E-Pyruvate | 13833.8 | 1 | 13833.8 | 15.78 | 0.0019 | |
| F-Methyl Jasmonate | 6244.58 | 1 | 6244.58 | 7.12 | 0.0205 | |
| M-Salicylic acid | 14161.84 | 1 | 14161.84 | 16.15 | 0.0017 | |
| P-Cysteine | 5294.26 | 1 | 5294.26 | 6.04 | 0.0302 | |
| R-Phenylalanine | 10089.03 | 1 | 10089.03 | 11.51 | 0.0053 | |
| T-Fluconazole | 4089.8 | 1 | 4089.8 | 4.67 | 0.0517 | |
| Residual | 10520.32 | 12 | 876.69 | |||
| Cor Total | 77106.37 | 19 | ||||
| Coefficient | Standard Error | 95% CI | VIF | |||
| Factor | Estimate | df | Low | High | ||
| Intercept | 47.95 | 1 | 6.620776 | 33.52457 | 62.37543 | |
| D-Pyruvate | −25.37 | 1 | 6.620776 | −39.7954 | −10.9446 | 1 |
| F-Methyl Jasmonate | 17.67 | 1 | 6.620776 | 3.244568 | 32.09543 | 1 |
| M-Salicylic acid | −26.61 | 1 | 6.620776 | −41.0354 | −12.1846 | 1 |
| P-Cysteine | −16.27 | 1 | 6.620776 | −30.6954 | −1.84457 | 1 |
| R-Phenylalanine | −22.46 | 1 | 6.620776 | −36.8854 | −8.03457 | 1 |
| T-Fluconazole | −14.3 | 1 | 6.620776 | −28.7254 | 0.125432 | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Sayed, A.S.A.; Hassan, W.H.B.; Sweilam, S.H.; Alqarni, M.H.S.; El Sayed, Z.I.; Abdel-Aal, M.M.; Abdelsalam, E.; Abdelaziz, S. Production, Bioprocessing and Anti-Proliferative Activity of Camptothecin from Penicillium chrysogenum, “An Endozoic of Marine Sponge, Cliona sp.”, as a Metabolically Stable Camptothecin Producing Isolate. Molecules 2022, 27, 3033. https://doi.org/10.3390/molecules27093033
El-Sayed ASA, Hassan WHB, Sweilam SH, Alqarni MHS, El Sayed ZI, Abdel-Aal MM, Abdelsalam E, Abdelaziz S. Production, Bioprocessing and Anti-Proliferative Activity of Camptothecin from Penicillium chrysogenum, “An Endozoic of Marine Sponge, Cliona sp.”, as a Metabolically Stable Camptothecin Producing Isolate. Molecules. 2022; 27(9):3033. https://doi.org/10.3390/molecules27093033
Chicago/Turabian StyleEl-Sayed, Ashraf S. A., Wafaa H. B. Hassan, Sherouk Hussein Sweilam, Mohammed Hamed Saeed Alqarni, Zeinab I. El Sayed, Mahmoud M. Abdel-Aal, Eman Abdelsalam, and Sahar Abdelaziz. 2022. "Production, Bioprocessing and Anti-Proliferative Activity of Camptothecin from Penicillium chrysogenum, “An Endozoic of Marine Sponge, Cliona sp.”, as a Metabolically Stable Camptothecin Producing Isolate" Molecules 27, no. 9: 3033. https://doi.org/10.3390/molecules27093033
APA StyleEl-Sayed, A. S. A., Hassan, W. H. B., Sweilam, S. H., Alqarni, M. H. S., El Sayed, Z. I., Abdel-Aal, M. M., Abdelsalam, E., & Abdelaziz, S. (2022). Production, Bioprocessing and Anti-Proliferative Activity of Camptothecin from Penicillium chrysogenum, “An Endozoic of Marine Sponge, Cliona sp.”, as a Metabolically Stable Camptothecin Producing Isolate. Molecules, 27(9), 3033. https://doi.org/10.3390/molecules27093033