

Inhibitory Effect of Bisdemethoxycurcumin on DNCB-Induced Atopic Dermatitis in Mice

Abstract
:1. Introduction
2. Results
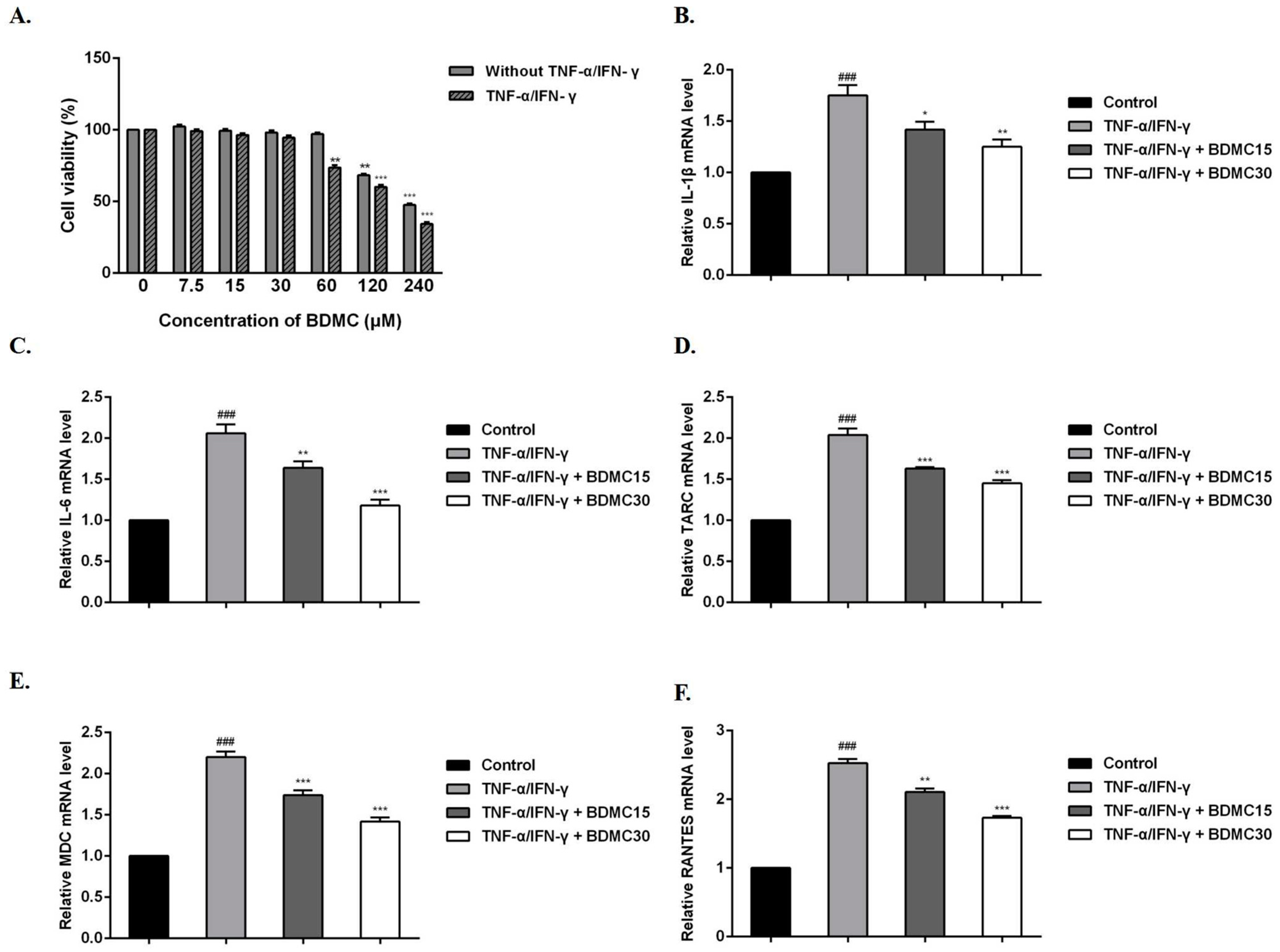
2.1. Effects of Different Concentrations of BDMC Treatment on the Viability of HaCaT Cells
2.2. BDMC Reduces the mRNA Expression Levels of Cytokines and Chemokines in HaCaT Cells
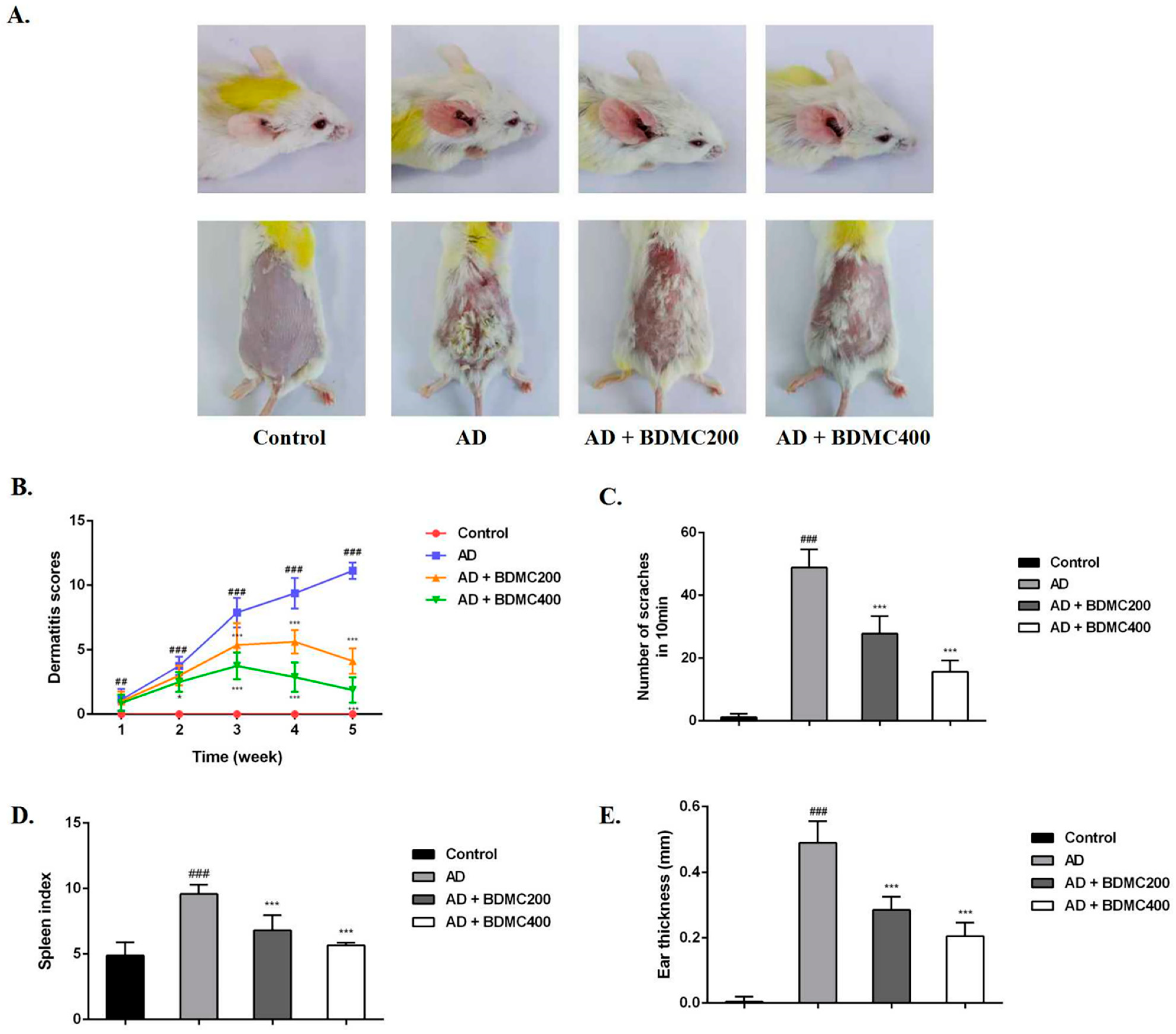
2.3. BDMC Alleviates the Clinical Symptoms of AD-like Skin Lesions in Mice
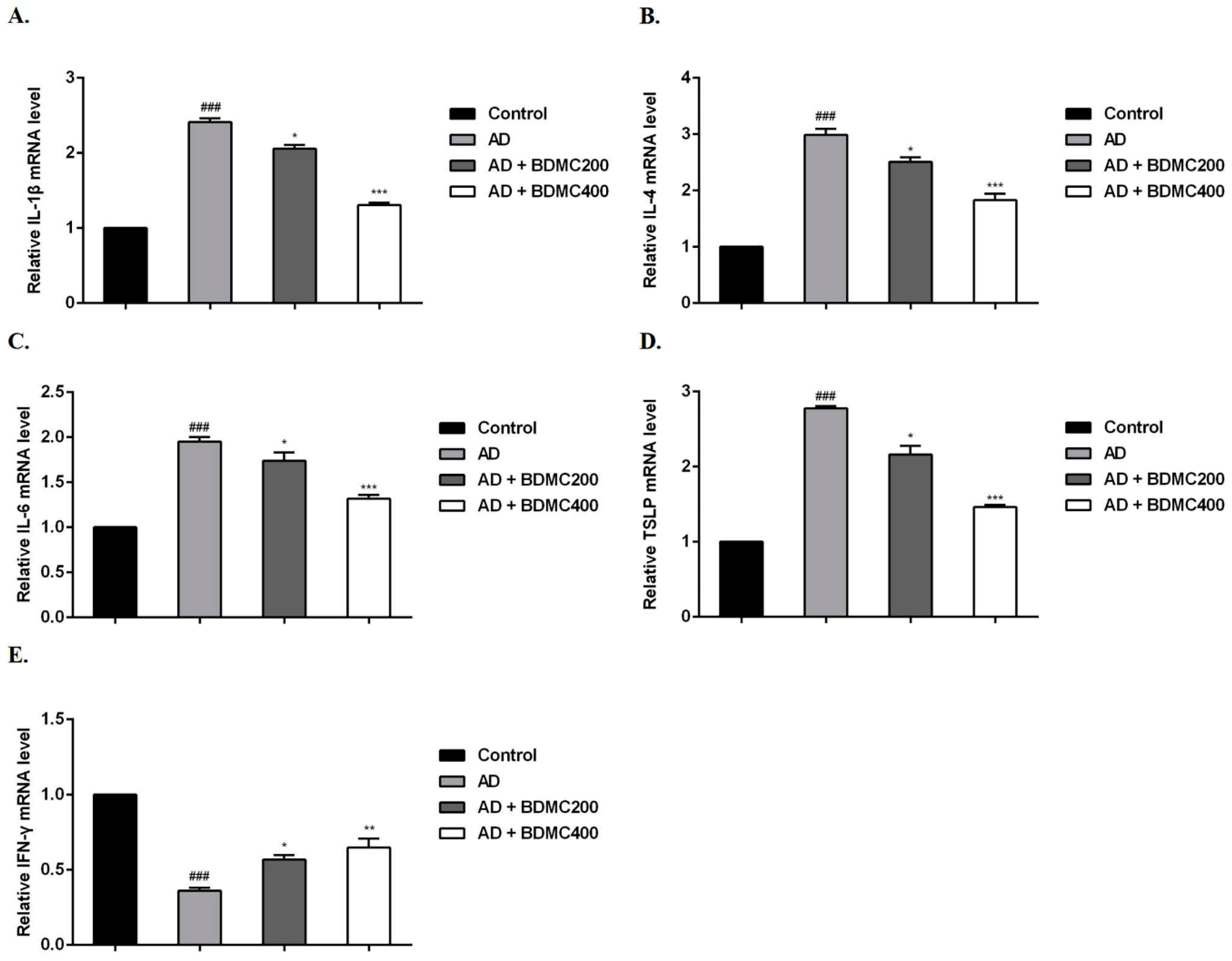
2.4. BDMC Decreases the Levels of Serum IgE, IL-4 and IFN-γ
2.5. BDMC Reduces Skin Hyperplasia and Mast Cell Infiltration in AD-like Skin Lesions
2.6. BDMC Reduces the mRNA Expression of Inflammatory Cytokines in AD-like Skin Lesions
2.7. BDMC Inhibits the Activation of MAPK and NF-κB Pathways in AD-like Skin Lesions
3. Discussion
4. Methods
4.1. Mice
4.2. Cell Culture and Treatment
4.3. Cell Viability Assay
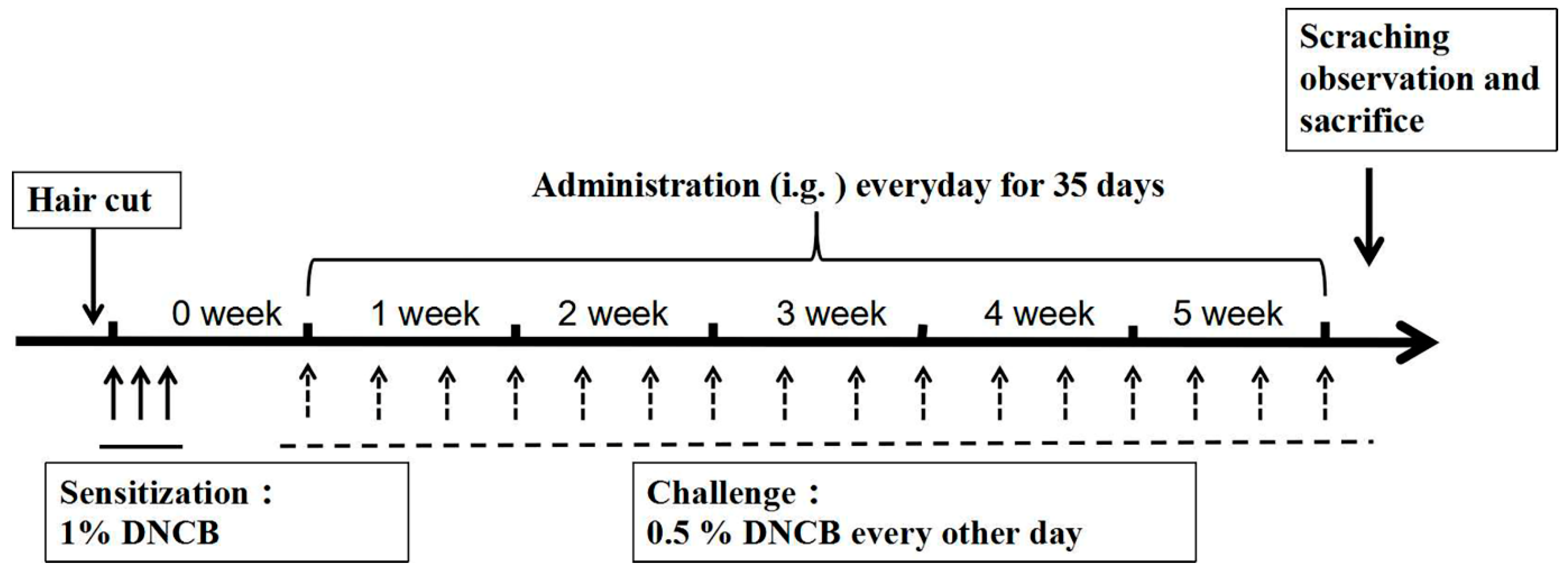
4.4. Induction of AD Mouse Model
4.5. Drug Treatment
4.6. Atopic Dermatitis Score, Scratches, Ear Thickness and Spleen Index
4.7. Histological Analysis
4.8. ELISA
4.9. Real-Time Polymerase Chain Reaction
4.10. Western Blot Assay
4.11. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Patient Consent for Publication
References
- Puar, N.; Chovatiya, R.; Paller, A.S. New treatments in atopic dermatitis. Ann. Allergy Asthma Immunol. 2021, 126, 21–31. [Google Scholar] [CrossRef] [PubMed]
- Czarnowicki, T.; He, H.; Krueger, J.G.; Guttman-Yassky, E. Atopic dermatitis endotypes and implications for targeted therapeutics. J. Allergy Clin. Immunol. 2019, 143, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Leung, D.; Guttman-Yassky, E. Assessing the current treatment of atopic dermatitis: Unmet needs. J. Allergy Clin. Immunol. 2017, 139, S47–S48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silverberg, J.I.; Gelfand, J.M.; Margolis, D.J.; Boguniewicz, M.; Fonacier, L.; Grayson, M.H.; Simpson, E.L.; Ong, P.Y.; Fuxench, Z.C.C. Association of atopic dermatitis with allergic, autoimmune, and cardiovascular comorbidities in US adults. Ann. Allergy Asthma Immunol. 2018, 121, 604–612e3. [Google Scholar] [CrossRef] [Green Version]
- Silverberg, J.I. Comorbidities and the impact of atopic dermatitis. Ann. Allergy Asthma Immunol. 2019, 123, 144–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paller, A.; Jaworski, J.C.; Simpson, E.L.; Boguniewicz, M.; Russell, J.J.; Block, J.K.; Tofte, S.; Dunn, J.D.; Feldman, S.R.; Clark, A.R.; et al. Major Comorbidities of Atopic Dermatitis: Beyond Allergic Disorders. Am. J. Clin. Dermatol. 2018, 19, 821–838. [Google Scholar] [CrossRef]
- Homey, B.; Steinhoff, M.; Ruzicka, T.; Leung, D.Y. Cytokines and chemokines orchestrate atopic skin inflammation. J. Allergy Clin. Immunol. 2006, 118, 178–189. [Google Scholar] [CrossRef]
- Paoletti, I.; De Gregorio, V.; Baroni, A.; Tufano, M.A.; Donnarumma, G.; Perez, J.J. Amygdalin Analogues Inhibit IFN-γ Signalling and Reduce the Inflammatory Response in Human Epidermal Keratinocytes. Inflammation 2013, 36, 1316–1326. [Google Scholar] [CrossRef]
- Yang, L.; Cohn, L.; Zhang, D.H.; Homer, R.; Ray, A.; Ray, P. Essential role of nuclear factor kappaB in the induction of eosinophilia in allergic airway inflammation. J. Exp. Med. 1998, 188, 1739–1750. [Google Scholar] [CrossRef] [Green Version]
- Roux, P.P.; Blenis, J. ERK and p38 MAPK-activated protein kinases: A family of protein kinases with diverse biological functions. Microbiol. Mol. Biol. Rev. 2004, 68, 320–344. [Google Scholar] [CrossRef]
- Sigala, I.; Zacharatos, P.; Toumpanakis, D.; Michailidou, T.; Noussia, O.; Theocharis, S.; Roussos, C.; Papapetropoulos, A.; Vassilakopoulos, T. MAPKs and NF-κB differentially regulate cytokine expression in the diaphragm in response to resistive breathing: The role of oxidative stress. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R1152–R1162. [Google Scholar] [CrossRef] [Green Version]
- Sung, Y.Y.; Kim, Y.S.; Kim, H.K. Illicium verum extract inhibits TNF-α- and IFN-γ-induced expression of chemokines and cytokines in human keratinocytes. J. Ethnopharmacol. 2012, 144, 182–189. [Google Scholar] [CrossRef]
- Shin, H.S.; See, H.J.; Jung, S.Y.; Choi, D.W.; Kwon, D.A.; Bae, M.J.; Sung, K.-S.; Shon, D.-H. Turmeric (Curcuma longa) attenuates food allergy symptoms by regulating type 1/type 2 helper T cells (Th1/Th2) balance in a mouse model of food allergy. J. Ethnopharmacol. 2015, 175, 21–29. [Google Scholar] [CrossRef]
- Kinney, S.R.M.; Carlson, L.; Ser-Dolansky, J.; Thompson, C.; Shah, S.; Gambrah, A.; Xing, W.; Schneider, S.S.; Mathias, C.B. Curcumin Ingestion Inhibits Mastocytosis and Suppresses Intestinal Anaphylaxis in a Murine Model of Food Allergy. PLoS ONE 2015, 10, e0132467. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.; Li, H.; Jia, J.; He, M. Anti-inflammatory effect of curcumin on mast cell-mediated allergic responses in ovalbumin-induced allergic rhinitis mouse. Cell. Immunol. 2015, 298, 88–95. [Google Scholar] [CrossRef]
- Gelders Géraldine Veerle, B.; Anke, V.D.P. Linking Neuroinflammation and Neurodegeneration in Parkinson’s Disease. J. Immunol. Res. 2018, 2018, 4784268. [Google Scholar]
- Fu, M.; Fu, S.; Ni, S.; Wang, D.; Hong, T. Inhibitory effects of bisdemethoxycurcumin on mast cell-mediated allergic diseases. Int. Immunopharmacol. 2018, 65, 182–189. [Google Scholar] [CrossRef]
- Wang, Y.; Zhang, P.; Zhang, J.; Hong, T. Bisdemethoxycurcumin attenuates OVA-induced food allergy by inhibiting the MAPK and NF-κB signaling pathways. Exp. Med. 2022, 23, 401. [Google Scholar] [CrossRef]
- Boothe, W.D.; Tarbox, J.A.; Tarbox, M.B. Atopic Dermatitis: Pathophysiology. Adv. Exp. Med. Biol. 2017, 1027, 21–37. [Google Scholar]
- Mehta, N.N.; Teague, H.L.; Swindell, W.R.; Baumer, Y.; Ward, N.L.; Xing, X.; Baugous, B.; Johnston, A.; Joshi, A.A.; Silverman, J.; et al. IFN-γ and TNF-α synergism may provide a link between psoriasis and inflammatory atherogenesis. Sci. Rep. 2017, 7, 13831. [Google Scholar] [CrossRef] [Green Version]
- Kong, L.; Liu, J.; Wang, J.; Luo, Q.; Zhang, H.; Liu, B.; Xu, F.; Pang, Q.; Liu, Y.; Dong, J. Icariin inhibits TNF-α/IFN-γ induced inflammatory response via inhibition of the substance P and p38-MAPK signaling pathway in human keratinocytes. Int. Immunopharmacol. 2015, 29, 401–407. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, A.B.; Chamian, F.; Masud, S.; Cardinale, I.; Abello, M.V.; Lowes, M.A.; Chen, F.; Magliocco, M.; Krueger, J.G. TNF inhibition rapidly down-regulates multiple proinflammatory pathways in psoriasis plaques. J. Immunol. 2005, 175, 2721–2729. [Google Scholar] [CrossRef] [PubMed]
- An, H.-J.; Kim, J.-Y.; Kim, W.-H.; Gwon, M.-G.; Gu, H.M.; Jeon, M.J.; Han, S.-M.; Pak, S.C.; Lee, C.-K.; Park, I.S.; et al. Therapeutic effects of bee venom and its major component, melittin, on atopic dermatitis in vivo and in vitro. Br. J. Pharmacol. 2018, 175, 4310–4324. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abramovits, W.; Bejarano, J.R.; Valdecantos, W.C. Role of Interleukin 1 in Atopic Dermatitis. Dermatol. Clin. 2013, 31, 437–444. [Google Scholar] [CrossRef]
- Qi, X.F.; Kim, D.H.; Yoon, Y.S.; Li, J.H.; Song, S.B.; Jin, D.; Huang, X.-Z.; Teng, Y.-C.; Lee, K.-J. The adenylyl cyclase-cAMP system suppresses TARC/CCL17 and MDC/CCL22 production through p38 MAPK and NF-κB in HaCaT keratinocytes. Mol. Immunol. 2009, 46, 1925–1934. [Google Scholar] [CrossRef]
- Yosipovitch, G.; Berger, T.; Fassett, M.S. Neuroimmune interactions in chronic itch of atopic dermatitis. J. Eur. Acad. Dermatol. Venereol. 2019, 34, 239–250. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.R.; Lee, H.; Kim, J.-H.; Hong, I.-H.; Hwang, D.H.; Rho, I.R.; Kim, G.S.; Kim, E.; Kang, C. Therapeutic Effect of Rumex japonicus Houtt. on DNCB-Induced Atopic Dermatitis-Like Skin Lesions in Balb/c Mice and Human Keratinocyte HaCaT Cells. Nutrients 2019, 11, 573. [Google Scholar] [CrossRef] [Green Version]
- Ku, J.M.; Hong, S.H.; Kim, H.I.; Seo, H.S.; Shin, Y.C.; Ko, S.G. Effects of Angelicae dahuricae Radix on 2, 4-Dinitrochlorobenzene-Induced Atopic Dermatitis-Like Skin Lesions in mice model. Bmc Complement. Altern. Med. 2017, 17, 98. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Lan, Y.; Wei, B.; Dai, C.; Gu, Y.; Ma, J.; Liu, X.; Umezawa, K. External application of NF-κB inhibitor DHMEQ suppresses development of atopic dermatitis-like lesions induced with DNCB/OX in BALB/c mice. Immunopharmacol. Immunotoxicol. 2017, 39, 157–164. [Google Scholar] [CrossRef]
- Mashiko, S.; Bouguermouh, S.; Rubio, M.; Baba, N.; Bissonnette, R.; Sarfati, M. Human mast cells are major IL-22 producers in patients with psoriasis and atopic dermatitis. J. Allergy Clin. Immunol. 2015, 136, 351–359. [Google Scholar] [CrossRef]
- Galli, S.J.; Tsai, M.; Piliponsky, A.M. The development of allergic inflammation. Nature 2008, 454, 445–454. [Google Scholar] [CrossRef] [Green Version]
- Bernard, F.X.; Morel, F.; Camus, M.; Pedretti, N.; Barrault, C.; Garnier, J.; Lecron, J.C. Keratinocytes under Fire of Proinflammatory Cytokines: Bona Fide Innate Immune Cells Involved in the Physiopathology of Chronic Atopic Dermatitis and Psoriasis. J. Allergy 2012, 2012, 718725. [Google Scholar] [CrossRef]
- Gauvreau, G.M.; Sehmi, R.; Ambrose, C.S.; Griffiths, J.M. Thymic stromal lymphopoietin: Its role and potential as a therapeutic target in asthma. Expert Opin. Ther. Targets 2020, 24, 777–792. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, B. Functions of thymic stromal lymphopoietin in immunity and disease. Immunol. Res. 2012, 52, 211–223. [Google Scholar] [CrossRef] [Green Version]
- Soumelis, V.; Reche, P.A.; Kanzler, H.; Yuan, W.; Edward, G.; Homey, B.; Gilliet, M.; Ho, S.; Antonenko, S.; Lauerma, A.; et al. Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP. Nat. Immunol. 2002, 3, 673–680. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.H.; Shin, T.Y. Anti-inflammatory effect of leaves of Eriobotrya japonica correlating with attenuation of p38 MAPK, ERK, and NF-kappaB activation in mast cells. Toxicol. Vitr. 2009, 23, 1215–1219. [Google Scholar] [CrossRef]
- Yang, C.-C.; Hung, Y.-L.; Ko, W.-C.; Tsai, Y.-J.; Chang, J.-F.; Liang, C.-W.; Chang, D.-C.; Hung, C.-F. Effect of Neferine on DNCB-Induced Atopic Dermatitis in HaCaT Cells and BALB/c Mice. Int. J. Mol. Sci. 2021, 22, 8237. [Google Scholar] [CrossRef]
- Bai, X.Y.; Liu, P.; Chai, Y.W.; Wang, Y.; Ren, S.H.; Li, Y.Y.; Zhou, H. Artesunate attenuates 2,4-dinitrochlorobenzene-induced atopic dermatitis by down-regulating Th17 cell responses in BALB/c mice. Eur. J. Pharmacol. 2020, 874, 173020. [Google Scholar] [CrossRef]
- Meng, Y.; Liu, Z.; Zhai, C.; Di, T.; Zhang, L.; Zhang, L.; Xie, X.; Lin, Y.; Wang, N.; Zhao, J.; et al. Paeonol inhibits the development of 1-chloro-2,4-dinitrobenzene-induced atopic dermatitis via mast and T cells in BALB/c mice. Mol. Med. Rep. 2019, 19, 3217–3229. [Google Scholar] [CrossRef] [Green Version]
- Matsuoka, H.; Maki, N.; Yoshida, S.; Arai, M.; Wang, J.; Oikawa, Y.; Ikeda, T.; Hirota, N.; Nakagawa, H.; Ishii, A. A mouse model of the atopic eczema/dermatitis syndrome by repeated application of a crude extract of house-dust mite Dermatophagoides farinae. Allergy 2003, 58, 139–145. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Forward | Reverse |
|---|---|---|
| GAPDH | CTG CTC CTC CTG TTC GAC AGT | CCG TTG ACT CCG ACC TTC AC |
| mIL-4 | TAC CAG GAG CCA TAT CCA CGG ATG | TGT GGT GTT CTT CGT TGC TGT GAG |
| mIFN-γ | GAG CCT AGA GAC TAT CAC ACC G | TAC CAG AGG GTG TAG TTA GCG G |
| mIL-1β | TGG ACC TTC CAG GAT GAG GAC A | GTT CAT CTC GGA GCC TGT AGT G |
| mIL-6 | AGT TGC CTT CTT GGG ACT GA | TCC ACG ATT TCC CAG AGA AC |
| mTSLP | AGC TTG TCT CCT GAA AAT CGA G | AGG TTT GAT TCA GGC AGA TGT T |
| hIL-1β | CTC TCA CCT CTC CTA CTC ACT | ATC AGA ATG TGG GAG CGA AT |
| hIL-6 | CGA GCC CAC CGG GAA CGA AA | GGA CCG AAG GCG CTT GTG GAG |
| hTARC | GTC TTG AAG CCT CCT CAC CC | GGA TCT CCC TCA CTG TGG CT |
| hMDC | GTT GTC CTC GTC CTC CTT GC | GGA GTC TGA GGT CCA GTA GAA GTG |
| hRANTES | CGC TGT CAT CCT CAT TGC TA | GCA CTT GCC ACT GGT GTA GA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Zhang, P.; Zhang, J.; Hong, T. Inhibitory Effect of Bisdemethoxycurcumin on DNCB-Induced Atopic Dermatitis in Mice. Molecules 2023, 28, 293. https://doi.org/10.3390/molecules28010293
Wang Y, Zhang P, Zhang J, Hong T. Inhibitory Effect of Bisdemethoxycurcumin on DNCB-Induced Atopic Dermatitis in Mice. Molecules. 2023; 28(1):293. https://doi.org/10.3390/molecules28010293
Chicago/Turabian StyleWang, Yanjie, Ping Zhang, Jingyu Zhang, and Tie Hong. 2023. "Inhibitory Effect of Bisdemethoxycurcumin on DNCB-Induced Atopic Dermatitis in Mice" Molecules 28, no. 1: 293. https://doi.org/10.3390/molecules28010293
APA StyleWang, Y., Zhang, P., Zhang, J., & Hong, T. (2023). Inhibitory Effect of Bisdemethoxycurcumin on DNCB-Induced Atopic Dermatitis in Mice. Molecules, 28(1), 293. https://doi.org/10.3390/molecules28010293