
The Study of Alanine Transaminase Activity in Tissues of Silkworm (Bombyx mori) via Direct Analysis in Real-Time (DART) Mass Spectrometry
 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Optimization of DART-MS/MS Experimental Parameters
2.1.1. Effect of Gas Temperature in DART Ionization Source
2.1.2. Effect of Grid Electrode Voltage
2.1.3. Effect of Sample Presentation Speed
2.2. The Characterization of Amino Acid via DART-MS/MS
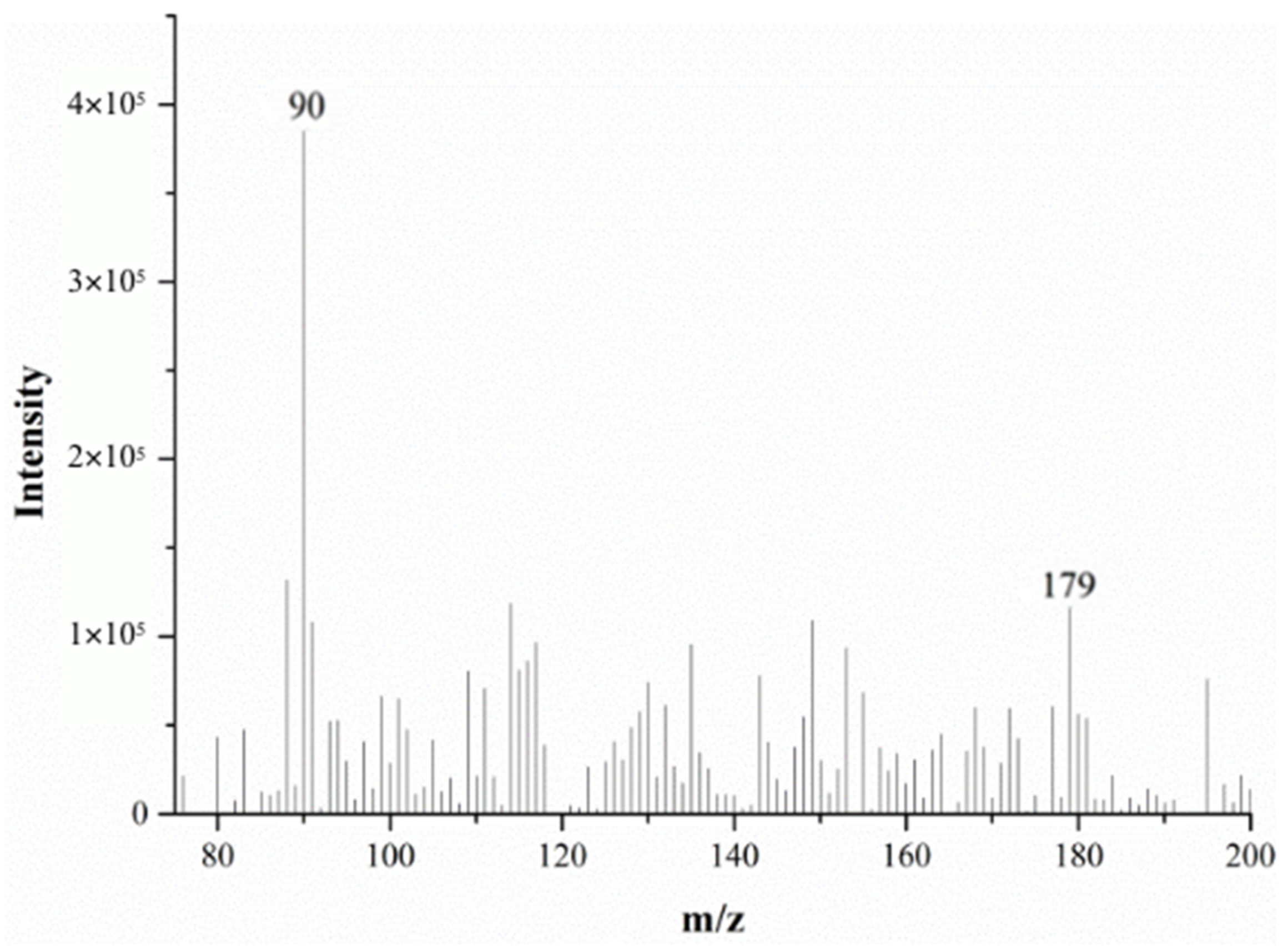
2.2.1. Alanine MS in Positive Ion Mode
2.2.2. Alanine MS in Negative Ion Mode
2.3. Determination of Alanine and ALT Activity in the Tissues of Silkworm by DART-MS
2.3.1. Calibration Curve
2.3.2. Precision and Spike Recovery Tests
2.4. The Evaluation of the ALT Activity in Silkworm Tissues
2.4.1. Quantifying ALT Activity in the Hemolymph of Silkworms
2.4.2. Determination of ALT Activity in Tissues of Bombyx mori L.: The Posterior Silk Gland, Midgut, Fat Body, Middle Silk Gland and Trachea
2.4.3. Limit of Detection
2.5. The Relation between the Variation in ALT Activity and the Production of Silk Protein
3. Materials and Methods
3.1. Chemicals and Reagents
3.2. Samples
3.2.1. Preparation of Tissues from Silkworm Larvae
3.2.2. Extraction of ALT
3.3. The Expression of ALT Activity Unit
3.4. Effect of Reaction Time on the Determination of Enzyme Activity
3.5. Instrumentation
3.5.1. Mass Spectrometer
3.5.2. DART SVP Ionization Source
3.6. Reitman-Frankel Method
3.7. Preparation of Samples in This Method
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Dai, M.L.; Feng, P.A.; Mao, T.T.; Gu, H.Y.; Bian, D.D.; Sun, H.N.; Li, F.C.; Wei, J.; Li, B. Study of compensatory growth based on different nutrition conditions of Bombyx mori. J. Asia Pac. Entomol. 2022, 25, 101948. [Google Scholar] [CrossRef]
- Zhou, Z.H.; Yang, H.J.; Chen, M.; Lou, C.F.; Zhang, Y.Z.; Chen, K.P.; Wang, Y.; Yu, M.L.; Yu, F.; Li, J.Y.; et al. Comparative proteomic analysis between the domesticated silkworm (Bombyx mori) reared on fresh mulberry leaves and on artificial diet. J. Proteome Res. 2008, 7, 5103–5111. [Google Scholar] [CrossRef]
- Li, B.; Hu, R.P.; Cheng, Z.; Cheng, J.; Xie, Y.; Gui, S.X.; Sun, Q.Q.; Sang, X.Z.; Gong, X.L.; Cui, Y.L.; et al. Titanium dioxide nanoparticles relieve biochemical dysfunctions of fifth-instar larvae of silkworms following exposure to phoxim insecticide. Chemosphere 2012, 89, 609–614. [Google Scholar] [CrossRef]
- Etebari, K.; Bizhannia, A.R.; Sorati, R.; Matindoost, L. Biochemical changes in haemolymph of silkworm larvae due to pyriproxyfen residue. Pestic. Biochem. Phys. 2007, 88, 14–19. [Google Scholar] [CrossRef]
- Zhang, L.L.; Bai, Y.L.; Shu, S.L.; Qian, D.W.; Zhen, O.Y.; Liu, L.; Duan, J.A. Simultaneous quantitation of nucleosides, nucleobases, amino acids, and alkaloids in mulberry leaf by ultra high performance liquid chromatography with triple quadrupole tandem mass spectrometry. J. Sep. Sci. 2014, 37, 1265–1275. [Google Scholar] [CrossRef]
- Hsieh, H.Y.; Li, L.H.; Hsu, R.Y.; Kao, W.F.; Huang, Y.C.; Hsu, C.C. Quantification of endogenous cholesterol in human serum on paper using direct analysis in Real Time Mass Spectrometry. Anal. Chem. 2017, 89, 6147–6153. [Google Scholar] [CrossRef]
- Peacock, P.M.; Zhang, W.J.; Trimpin, S. Advances in ionization for mass spectrometry. Anal. Chem. 2017, 89, 372–388. [Google Scholar] [CrossRef]
- Clendinen, C.S.; Monge, M.E.; Fernandez, F.M. Ambient mass spectrometry in metabolomics. Analyst 2017, 142, 3101–3117. [Google Scholar] [CrossRef]
- Liu, H.W.; Li, X.J.; Wang, X.; Li, N.; Bai, Y. Direct analysis in real time mass spectrometry: A powerful tool for fast analysis. Mass Spectrom. Lett. 2015, 6, 1–6. [Google Scholar] [CrossRef]
- Cody, R.B.; Laramee, J.A.; Durst, H.D. Versatile new ion source for the analysis of materials in open air under ambient conditions. Anal. Chem. 2005, 77, 2297–2302. [Google Scholar] [CrossRef] [PubMed]
- Knizner, K.T.; Guymon, J.P.; Garrard, K.P.; Bouvree, G.; Manni, J.; Hauschild, J.P.; Strupat, K. Next-generation infrared matrix-assisted laser desorption electrospray ionization source for mass spectrometry imaging and high-throughput screening. J. Am. Soc. Mass Spectrom. 2022, 33, 2070–2077. [Google Scholar] [CrossRef]
- Qi, W.S.; Zhang, L.; Guo, Y.L. Rapid identification of synthetic compounds in natural repellent products by direct analysis in real time mass spectrometry. Chin. Org. Chem. 2013, 33, 359–364. [Google Scholar] [CrossRef]
- Song, Y.Q.; Liao, J.; Zha, C.; Wang, B.; Liu, C.C. A novel approach to determine the tyrosine concentration in human plasma by DART-MS/MS. Anal. Methods 2015, 7, 1600–1605. [Google Scholar] [CrossRef]
- Maeno, K.; Shida, Y.; Shimada, H. Direct quantitative analysis of the natural moisturizing factor (NMF) in the stratum corneum by direct analysis in real time mass spectrometry (DART-MS). Anal. Methods 2017, 9, 4851–4857. [Google Scholar] [CrossRef]
- Gu, H.W.; Pan, Z.Z.; Xi, B.W.; Asiago, V.; Musselman, B.; Raftery, D. Principal component directed partial least squares analysis for combining nuclear magnetic resonance and mass spectrometry data in metabolomics: Application to the detection of breast cancer. Anal. Chim. Acta 2011, 686, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.S.; Guan, W.; Walker, L.D.; Mezencev, R.; Benigno, B.B.; Gray, A.; Fernandez, F.M.; Mcdonald, J.F. Rapid mass spectrometric metabolic profiling of blood sera detects ovarian cancer with high accuracy. Cancer Epidem. Biomar. 2010, 19, 2262–2271. [Google Scholar] [CrossRef]
- Cajka, T.; Danhelova, H.; Zachariasova, M.; Riddellova, K.; Hajslova, J. Application of direct analysis in real time ionization-mass spectrometry (DART-MS) in chicken meat metabolomics aiming at the retrospective control of feed fraud. Metabolomics 2013, 9, 545–557. [Google Scholar] [CrossRef]
- Thurman, E.M.; Ferrer, I.; Rosenblum, J.; Linden, K.; Ryan, J.N. Identification of polypropylene glycols and polyethylene glycol carboxylates in flowback and produced water from hydraulic fracturing. J. Hazard. 2017, 323, 11–17. [Google Scholar] [CrossRef]
- Haunschmidt, M.; Klampfl, C.W.; Buchberger, W.; Hertsens, R. Rapid identification of stabilisers in polypropylene using time-of-flight mass spectrometry and DART as ion source. Analyst 2010, 135, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhao, P.Y.; Zhang, F.Z.; Li, Y.J.; Pan, C.P. Direct analysis in real time mass spectrometry for the rapid identification of four highly hazardous pesticides in agrochemicals. Rapid Commun. Mass Spectrom. 2012, 26, 1859–1867. [Google Scholar] [CrossRef]
- Cody, R.B. Observation of molecular ions and analysis of nonpolar compounds with the direct analysis in real time ion source. Anal. Chem. 2009, 81, 1101–1107. [Google Scholar] [CrossRef]
- Li, Y.; Meng, L.; Li, Z.Z.; Wang, Y.; Wang, X.; Liu, H.; Xiong, C.Q.; Nie, Z.X. Hand-powered ionization methods for the mass spectrometric detection of small molecules. Int. J. Mass Spectrom. 2021, 470, 116716. [Google Scholar] [CrossRef]
- Farre, M.; Pico, Y.; Barcelo, D. Direct peel monitoring of xenobiotics in fruit by direct analysis in real time coupled to a linear quadrupole ion trap-orbitrap mass spectrometer. Anal. Chem. 2013, 85, 2638–2644. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Zhang, D.Y.; Liu, Z.Y.; Zhang, Y.; Liu, L.; Li, L.; Liu, C.C.; Wu, G.H. Rapid determination of 1-deoxynojirimycin in Morus alba L. leaves by direct analysis in real time (DART) mass spectrometry. J. Pharmaceut. Biomed. 2015, 114, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Brand, M.; Ranish, J.A. Proteomic/transcriptomic analysis of erythropoiesis. Curr. Opin. Hematol. 2021, 128, 150–157. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.H.; You, Z.Y.; Feng, M.; Che, J.Q.; Zhang, Y.Y.; Qian, Q.J.; Komatsu, S.; Zhong, B.X. Analyses of the molecular mechanisms associated with silk production in silkworm by iTRAQ-based proteomics and RNA-sequencing-based transcriptomics. J. Proteome Res. 2016, 15, 15–28. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Measurement (μmol) | Spiked (μmol) | Spiked Measurement (μmol) | Recovery (%) |
|---|---|---|---|---|
| 0.16 | 0.356 ± 0.011 | 97.61% | ||
| Posterior | 0.20 | 0.22 | 0.432 ± 0.019 | 105.37% |
| silk gland | 0.26 | 0.491 ± 0.022 | 111.92% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, G.; Jiang, L.; Guo, J.; Li, W.; Ma, L.; Tang, B.; Liu, C.C. The Study of Alanine Transaminase Activity in Tissues of Silkworm (Bombyx mori) via Direct Analysis in Real-Time (DART) Mass Spectrometry. Molecules 2023, 28, 4131. https://doi.org/10.3390/molecules28104131
Wu G, Jiang L, Guo J, Li W, Ma L, Tang B, Liu CC. The Study of Alanine Transaminase Activity in Tissues of Silkworm (Bombyx mori) via Direct Analysis in Real-Time (DART) Mass Spectrometry. Molecules. 2023; 28(10):4131. https://doi.org/10.3390/molecules28104131
Chicago/Turabian StyleWu, Guohua, Lei Jiang, Jianjun Guo, Wushuang Li, Lin Ma, Bozhi Tang, and Charles C. Liu. 2023. "The Study of Alanine Transaminase Activity in Tissues of Silkworm (Bombyx mori) via Direct Analysis in Real-Time (DART) Mass Spectrometry" Molecules 28, no. 10: 4131. https://doi.org/10.3390/molecules28104131