
In Silico and In Vitro Methods in the Characterization of Beta-Carotene as Pharmaceutical Material via Acetylcholine Esterase Inhibitory Actions

 , , ,
, , ,
Abstract
:1. Introduction
2. Results
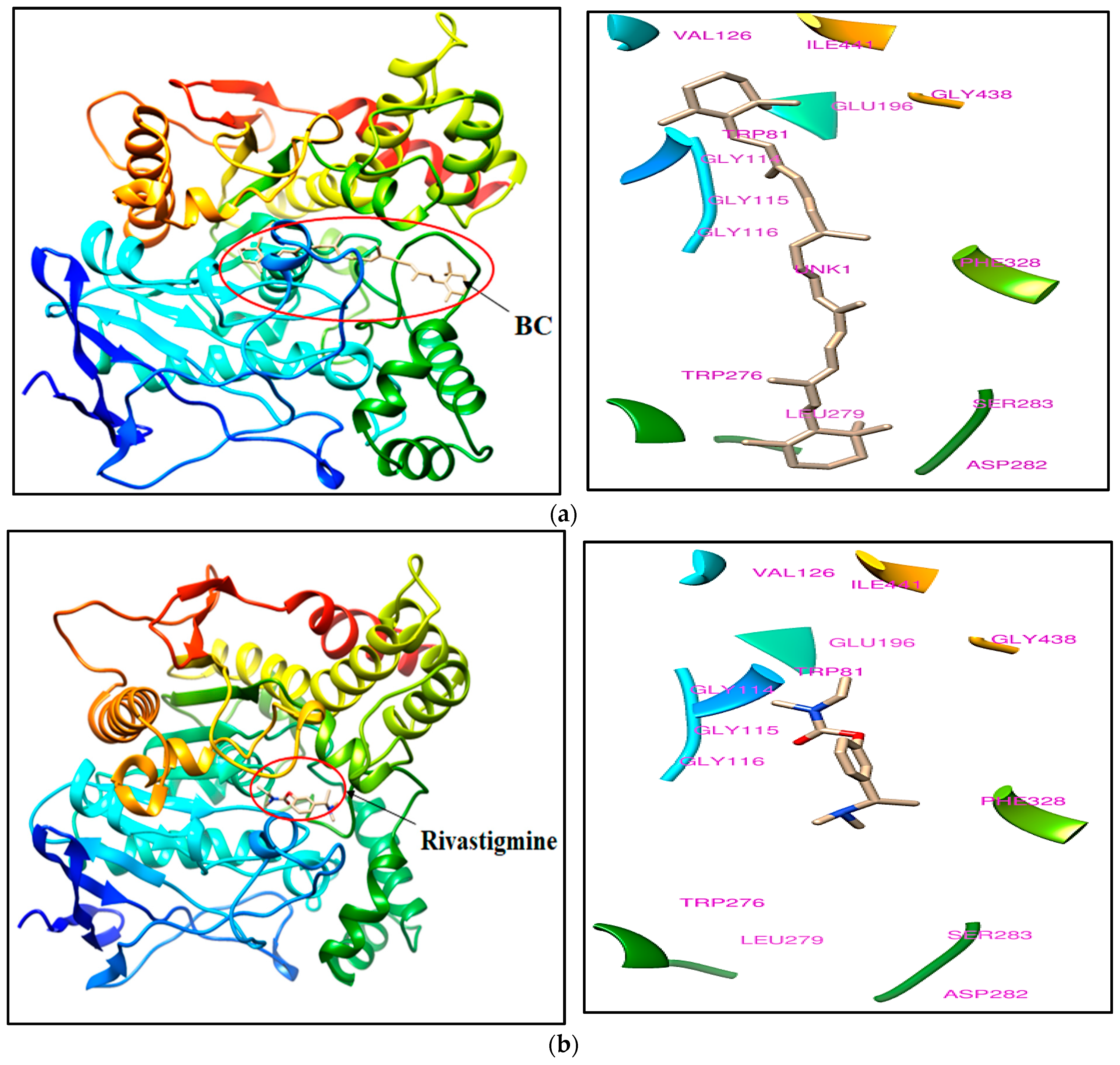
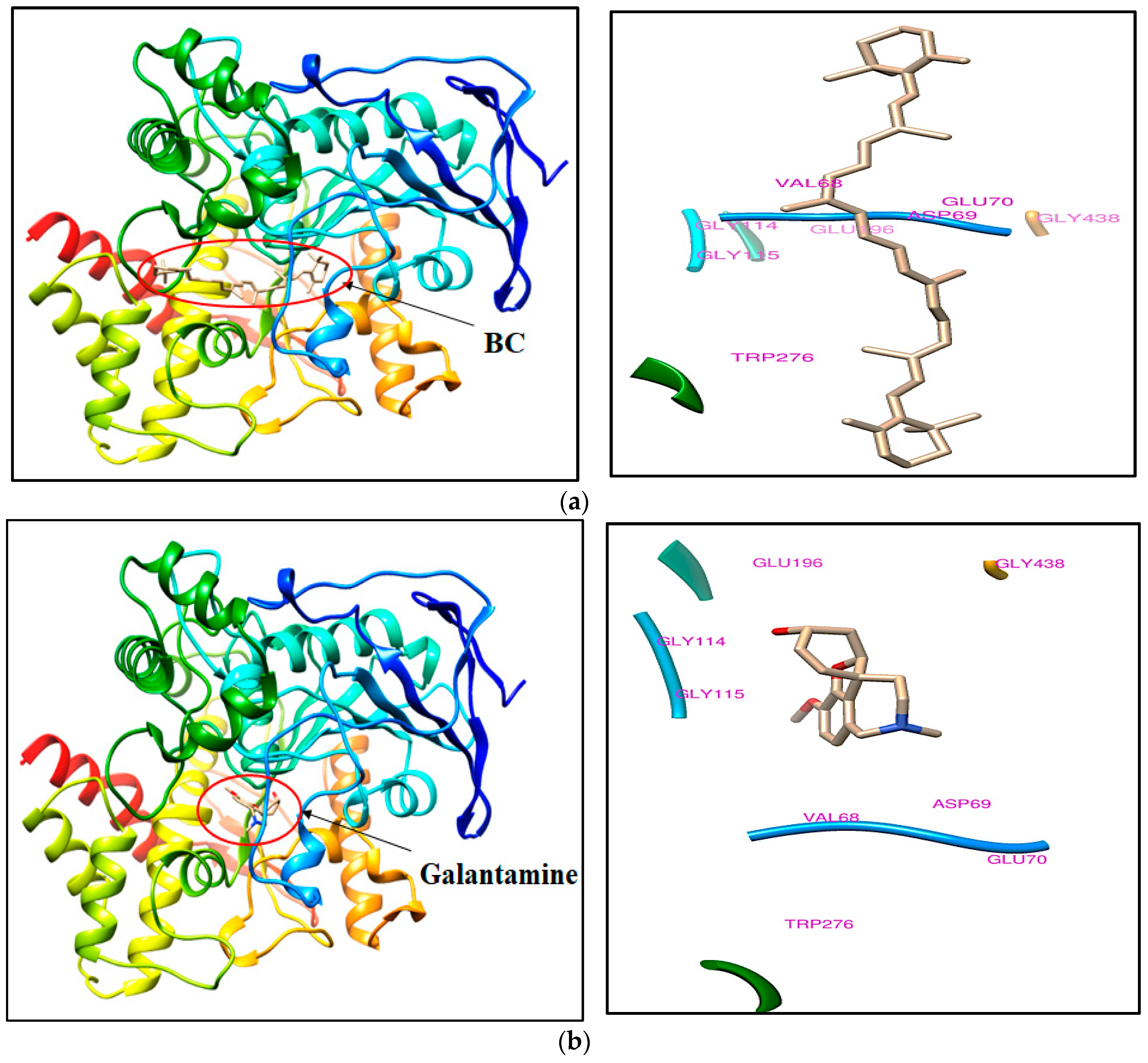
2.1. Ligand Binding Landscape Analysis of BC
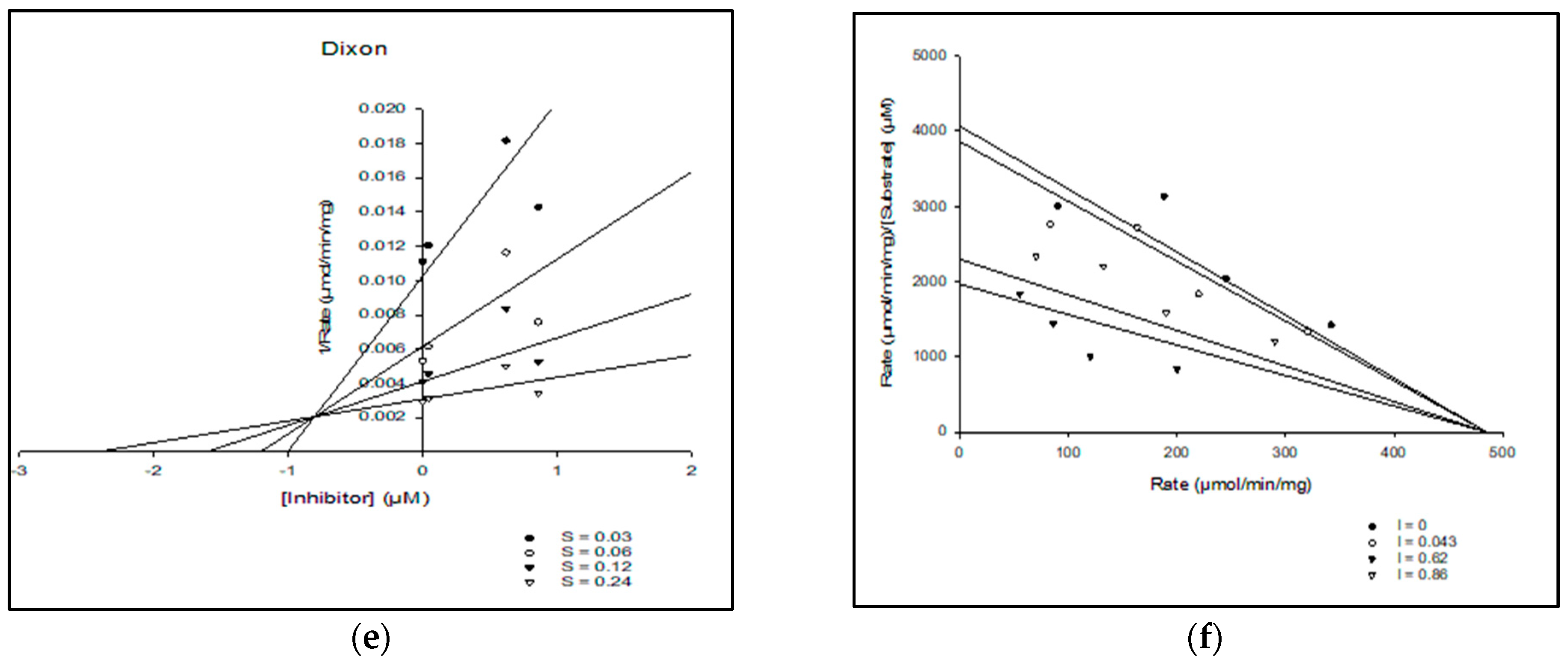
2.2. In Vitro Kinetic Assessment of Acetylcholine Esterase Inhibition
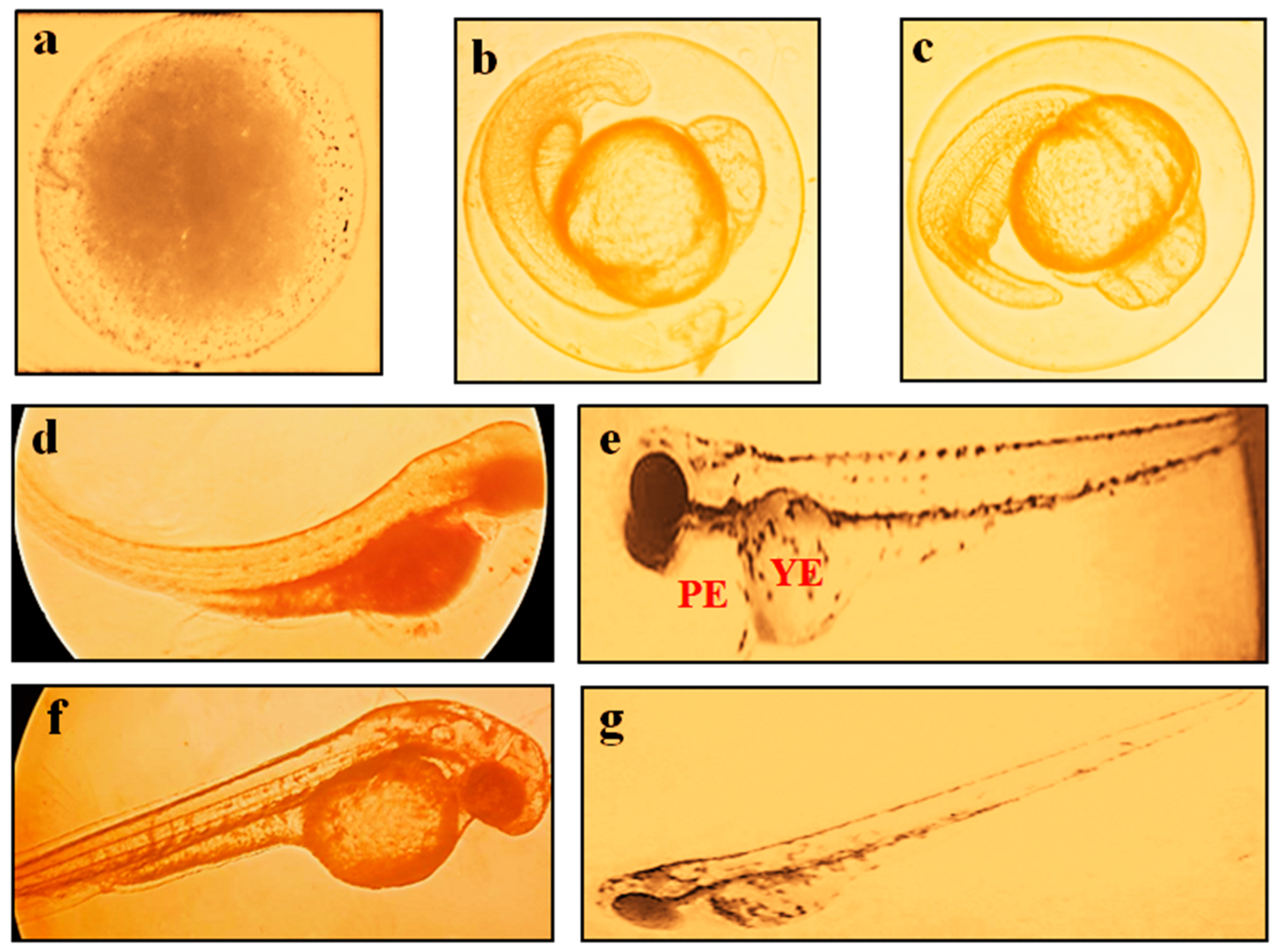
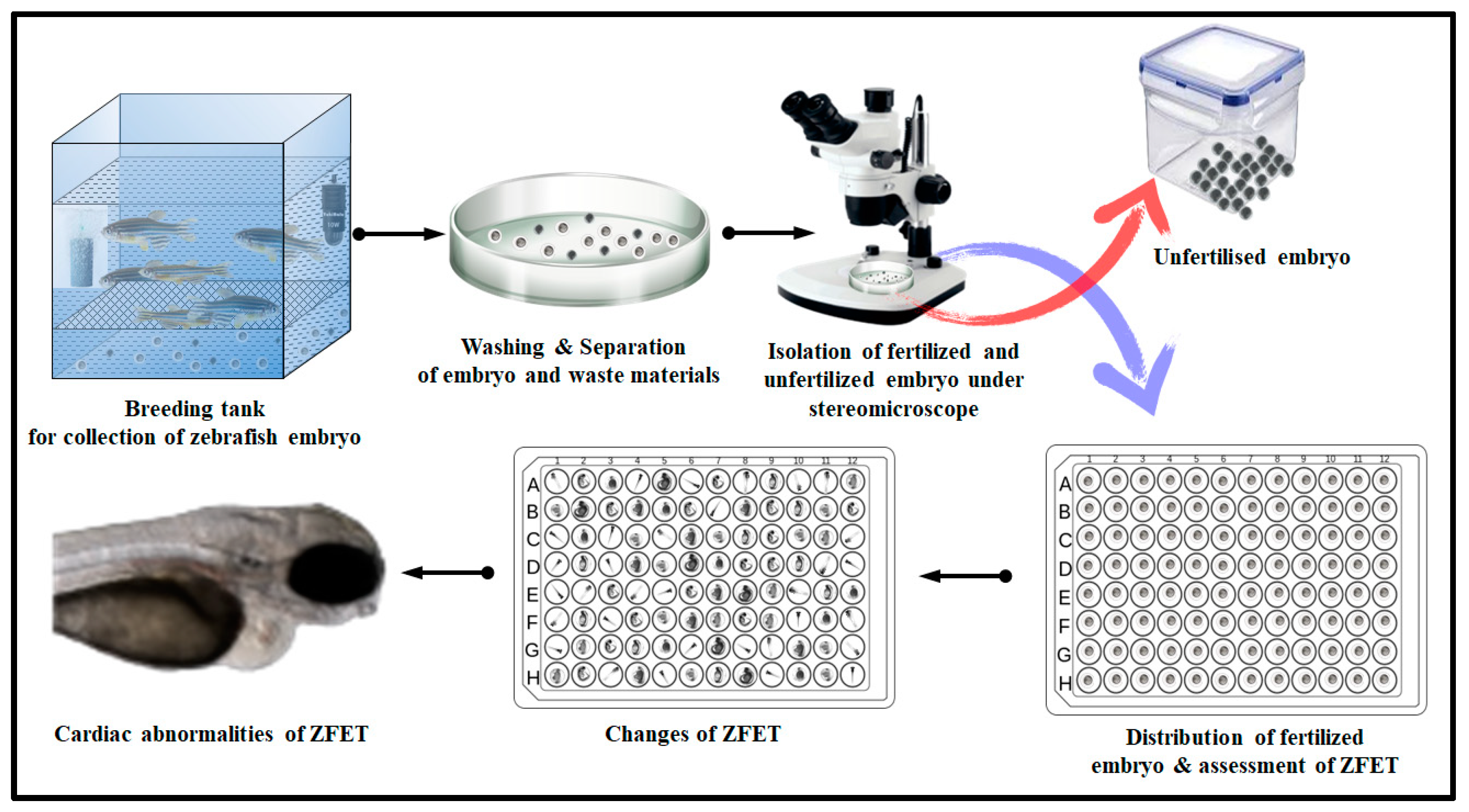
2.3. Effect of BC in ZFET
2.4. Effect of BC in AChE and AP Activity Levels in the Zebrafish Embryo
3. Discussion
4. Materials and Methods
4.1. In Silico Computational Studies to Characterize the Ligand Binding Mode
4.2. In Vitro Kinetic Studies to Determine the ACE Inhibition Mechanism of the Ligand
4.3. Study of BC in Zebrafish Embryo Toxicity Test (ZFET)
4.4. Estimation of AP Activity
4.5. Estimation of AChE Activity
4.6. Estimation of Tissue Total Proteins
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Dhanasekaran, S.; Perumal, P.; Palayan, M. In-Vitro Screening for Acetylcholinesterase Enzyme Inhibition Potential and Antioxidant Activity of Extracts of Ipomoea Aquatica Forsk: Therapeutic Lead for Alzheimer’s Disease. J. Appl. Pharm. Sci. 2015, 5, 012–016. [Google Scholar] [CrossRef]
- Balkis, A.; Tran, K.; Lee, Y.Z.; Ng, K. Screening Flavonoids for Inhibition of Acetylcholinesterase Identified Baicalein as the Most Potent Inhibitor. J. Agric. Sci. 2015, 7, 26. [Google Scholar] [CrossRef]
- Ferreira, J.; Santos, S.; Pereira, H. In vitro Screening for Acetylcholinesterase Inhibition and Antioxidant Activity of Quercus Suber Cork and Corkback Extracts. Evid. Based Complement. Altern. Med. 2020, 2020, 3825629. [Google Scholar] [CrossRef]
- Sivaprakasam, K. Towards a Unifying Hypothesis of Alzheimers Disease: Cholinergic System Linked to Plaques, Tangles and Neuroinflammation. Curr. Med. Chem. 2006, 13, 2179–2188. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, S.A.; Varatharajan, R.; Muthuraman, A. Palm Oil Derived Tocotrienol-Rich Fraction Attenuates Vascular Dementia in Type 2 Diabetic Rats. Int. J. Mol. Sci. 2022, 23, 13531. [Google Scholar] [CrossRef] [PubMed]
- Giap, L.K.; Varatharajan, R.; Muthuraman, A. Therapeutic Investigations of Palm Oil Induced Beta-Carotene in Diabetic Vascular Dementia in Rat. Res. J. Pharm. Technol. 2023, 16, 566–572. [Google Scholar] [CrossRef]
- Muthuraman, A.; Thilagavathi, L.; Jabeen, S.; Ravishankar, S.B.; Ahmed, S.S.; George, T.; Rishitha, N.; Paramakrishnan, N. Curcumin Prevents Cigarette Smoke Extract Induced Cognitive Impairment. Front. Biosci. Elite 2019, 11, 109–120. [Google Scholar] [CrossRef]
- Rishitha, N.; Muthuraman, A. Preventative Effects of Alpha-Naphtho Flavone in Vascular Dementia. Front. Biosci. 2020, 12, 79–94. [Google Scholar] [CrossRef]
- Rishitha, N.; Muthuraman, A. Therapeutic Investigation of Alpha Napthoflavone in the Intracerebroventricular Injection of L-cysteine Induced Vascular Dementia in Rats. Alzheimer’s Dement. 2021, 17, e058344. [Google Scholar] [CrossRef]
- Dhamodharan, J.; Muthuraman, A.; Sekhar, G. Neuroanatomical Evaluation of the Therapeutic Potential of Rutin and Gefitinib in Beta-Amyloid Oligomer-Induced Alzheimer Disease in Swiss Albino Mice and to Probe the Neuroanatomical Parallels between Lissencephalic Alzheimer Mice Brain with Gyrencephali. In Journal of Anatomy; Wiley: Hoboken, NJ, USA, 2022; Volume 240, pp. 783–784. [Google Scholar]
- Lim, K.G.; Muthuraman, A. Ameliorative Processes of Beta-Carotene in Streptozotocin-Induced Diabetic Vascular Dementia in Rats. Processes 2022, 10, 1324. [Google Scholar] [CrossRef]
- Lim, K.G.; Varatharajan, R.; Muthuraman, A. The Attenuating Effect of Beta-Carotene on Streptozotocin Induced Diabetic Vascular Dementia Symptoms in Rats. Molecules 2022, 27, 4293. [Google Scholar] [CrossRef]
- Bilal, M.; Freed, S.; Ashraf, M.Z.; Zaka, S.M.; Khan, M.B. Activity of Acetylcholinesterase and Acid and Alkaline Phosphatases in Different Insecticide-Treated Helicoverpa Armigera (Hübner). Environ. Sci. Pollut. Res. Int. 2018, 25, 22903–22910. [Google Scholar] [CrossRef]
- Abou-Donia, M.B.; Abdo, K.M.; Timmons, P.R.; Proctor, J.E. Brain Acetylcholinesterase, Acid Phosphatase, and 2′,3′-cyclic Nucleotide-3′-phosphohydrolase and Plasma Butyrylcholinesterase Activities in Hens Treated with a Single Dermal Neurotoxic Dose of S,S,S-Tri-N-Butyl Phosphorotrithioate. Toxicol. Appl. Pharmacol. 1986, 82, 461–473. [Google Scholar] [CrossRef]
- Luo, X.; Lauwers, M.; Layer, P.G.; Wen, C. Non-Neuronal Role of Acetylcholinesterase in Bone Development and Degeneration. Front. Cell Dev. Biol. 2021, 8, 620543. [Google Scholar] [CrossRef]
- Wang, D.; Li, M.; Ma, J.; Wang, X.; Liu, J. Effects of Temperature on Cathepsin B, Cathepsin D and Acid Phosphatase during Embryo Development of the Hard Tick Haemaphysalis Longicornis. Exp. Appl. Acarol. 2023, 89, 105–115. [Google Scholar] [CrossRef]
- Finney, D.J. Probit Analysis, 3rd ed.; Cambridge University Press: Cambridge, UK, 1971. [Google Scholar]
- Behra, M.; Cousin, X.; Bertrand, C.; Vonesch, J.-L.; Biellmann, D.; Chatonnet, A.; Strähle, U. Acetylcholinesterase Is Required for Neuronal and Muscular Development in the Zebrafish Embryo. Nat. Neurosci. 2002, 5, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Alshami, A.; Varon, J. Acid Phosphatase. In StatPearls [Internet]; StatPearls Publishing: Tampa, FL, USA, 2021. [Google Scholar]
- Fialho, E.; Silveira, A.B.; Masuda, H.; Silva-Neto, M.A.C. Oocyte Fertilization Triggers Acid Phosphatase Activity during Rhodnius Prolixus Embryogenesis. Insect Biochem. Mol. Biol. 2002, 32, 871–880. [Google Scholar] [CrossRef] [PubMed]
- Lopatina, E.V.; Gavrichenko, A.V.; Pasatetskaya, N.A. Involvement of Acetylcholine and Na+, K+-ATPase in the Regulation of Skeletal Muscle Growth in a Chicken Embryo. J. Evol. Biochem. Physiol. 2023, 59, 285–292. [Google Scholar] [CrossRef]
- Wu, K.; Cheng, T.; Zhai, Z.; Jiang, C.; Zhou, X. Alpha 7-nicotinic Cholinergic Regulation of Pericyte-containing Retinal Capillaries. Br. J. Pharmacol. 2023. [Google Scholar] [CrossRef]
- Samar, M.E.; Avila, R.E.; de Fabro, S.P. Acid Phosphatase Activity in the Chick Ovary during Embryonic Development. Basic Appl. Histochem. 1984, 28, 413–423. [Google Scholar]
- Abou-Donia, M.B. Role of Acid Phosphatase in Delayed Neurotoxicity Induced by Leptophos in Hens. Biochem. Pharmacol. 1978, 27, 2055–2058. [Google Scholar] [CrossRef] [PubMed]
- Yasbin, R.; Sawicki, J.; MacIntyre, R.J. A Developmental Study of Acid Phosphatase-1 in Drosophila Melanogaster. Dev. Biol. 1978, 63, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Haga, S.; Uji, S.; Suzuki, T. Evaluation of the Effects of Retinoids and Carotenoids on Egg Quality Using a Microinjection System. Aquaculture 2008, 282, 111–116. [Google Scholar] [CrossRef]
- Quadro, L.; Giordano, E.; Costabile, B.K.; Nargis, T.; Iqbal, J.; Kim, Y.; Wassef, L.; Hussain, M.M. Interplay between β-Carotene and Lipoprotein Metabolism at the Maternal-Fetal Barrier. Biochim. Biophys. Acta. Mol. Cell Biol. Lipids 2020, 1865, 158591. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, M.R. Vitamin A and Retinoids as Mitochondrial Toxicants. Oxidative Med. Cell. Longev. 2015, 2015, 140267. [Google Scholar] [CrossRef] [PubMed]
- Samrani, L.M.M.; Pennings, J.L.A.; Hallmark, N.; Bars, R.; Tinwell, H.; Pallardy, M.; Piersma, A.H. Dynamic Regulation of Gene Expression and Morphogenesis in the Zebrafish Embryo Test after Exposure to All-Trans Retinoic Acid. Reprod. Toxicol. 2023, 115, 8–16. [Google Scholar] [CrossRef]
- Sandoval, C.; Mella, L.; Godoy, K.; Adeli, K.; Farías, J. β-Carotene Increases Activity of Cytochrome P450 2E1 during Ethanol Consumption. Antioxidants 2022, 11, 1033. [Google Scholar] [CrossRef]
- Chen, Y.; Wu, M.; Chen, F.; Wen, X.; Zhao, L.; Li, G.; Zhou, L. Potential Role of Inflammation in Relation to Dietary Sodium and β-Carotene with Non-Alcoholic Fatty Liver Disease: A Mediation Analysis. Nutr. Diabetes 2022, 12, 40. [Google Scholar] [CrossRef]
- Ni, R.; Leo, M.A.; Zhao, J.; Lieber, C.S. Toxicity of β-Carotene and Its Exacerbation by Acetaldehyde in HepG2 Cells. Alcohol Alcohol. 2001, 36, 281–285. [Google Scholar] [CrossRef]
- Pinos, I.; Coronel, J.; Albakri, A.; Blanco, A.; McQueen, P.; Molina, D.; Sim, J.; Fisher, E.A.; Amengual, J. β-Carotene Accelerates Resolution of Atherosclerosis by Promoting Regulatory T Cell Expansion in the Atherosclerotic Lesion. bioRxiv 2023. [Google Scholar] [CrossRef]
- Mignone, L.I.; Giovannucci, E.; Newcomb, P.A.; Titus-Ernstoff, L.; Trentham-Dietz, A.; Hampton, J.M.; Willett, W.C.; Egan, K.M. Dietary Carotenoids and the Risk of Invasive Breast Cancer. Int. J. Cancer 2009, 124, 2929–2937. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Gao, Y.; Zhi, X.; Ta, N.; Jiang, H.; Zheng, J. Association between Vitamin A, Retinol and Carotenoid Intake and Pancreatic Cancer Risk: Evidence from Epidemiologic Studies. Sci. Rep. 2016, 6, 38936. [Google Scholar] [CrossRef] [PubMed]
- Csepanyi, E.; Czompa, A.; Haines, D.; Lekli, I.; Bakondi, E.; Balla, G.; Tosaki, A.; Bak, I. Cardiovascular Effects of Low versus High-Dose Beta-Carotene in a Rat Model. Pharmacol. Res. 2015, 100, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Bartolucci, C.; Perola, E.; Pilger, C.; Fels, G.; Lamba, D. Three-Dimensional Structure of a Complex of Galanthamine (Nivalin®) with Acetylcholinesterase from Torpedo Californica: Implications for the Design of New Anti-Alzheimer Drugs. Proteins Struct. Funct. Genet. 2001, 42, 182–191. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera—A Visualization System for Exploratory Research and Analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Ellman, G.L.; Courtney, K.D.; Andres, V., Jr.; Featherstone, R.M. A New and Rapid Colorimetric Determination of Acetylcholinesterase Activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Di Giovanni, S.; Borloz, A.; Urbain, A.; Marston, A.; Hostettmann, K.; Carrupt, P.A.; Reist, M. In vitro Screening Assays to Identify Natural or Synthetic Acetylcholinesterase Inhibitors: Thin Layer Chromatography versus Microplate Methods. Eur. J. Pharm. Sci. 2008, 33, 109–119. [Google Scholar] [CrossRef]
- Adatto, I.; Lawrence, C.; Thompson, M.; Zon, L.I. A New System for the Rapid Collection of Large Numbers of Developmentally Staged Zebrafish Embryos. PLoS ONE 2011, 6, e21715. [Google Scholar] [CrossRef]
- Gonsar, N.; Schumann, A.C.; Buchard, J.N.; Liang, J.O. An Inexpensive, Efficient Method for Regular Egg Collection from Zebrafish in a Recirculating System. Zebrafish 2012, 9, 50–55. [Google Scholar] [CrossRef]
- Rawlings, J.M.; Belanger, S.E.; Connors, K.A.; Carr, G.J. Fish Embryo Tests and Acute Fish Toxicity Tests Are Interchangeable in the Application of the Threshold Approach. Environ. Toxicol. Chem. 2019, 38, 671–681. [Google Scholar] [CrossRef]
- Dede, E.B.; Kaglo, H.D. Aqua-Toxicological Effects of Water Soluble Fractions (WSF) of Diesel Fuel on O. Niloticus Fingerlings. J. Appl. Sci. Environ. Manag. 2001, 5. [Google Scholar] [CrossRef]
- Neil, M.W.; Horner, M.W. Studies on Acid Hydrolases in Adult and Foetal Tissues. Acid P-Nitrophenyl Phosphate Phosphohydrolases of Adult Guinea-Pig Liver. Biochem. J. 1964, 92, 217. [Google Scholar] [CrossRef] [PubMed]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein Measurement with the Folin Phenol Reagent. J. Biol. Chem. 1951, 193, 265–275. [Google Scholar] [CrossRef]
- Hedgpeth, B.M.; Redman, A.D.; Alyea, R.A.; Letinski, D.J.; Connelly, M.J.; Butler, J.D.; Zhou, H.; Lampi, M.A. Analysis of Sublethal Toxicity in Developing Zebrafish Embryos Exposed to a Range of Petroleum Substances. Environ. Toxicol. Chem. 2019, 38, 1302–1312. [Google Scholar] [CrossRef] [PubMed]
- Busquet, F.; Strecker, R.; Rawlings, J.M.; Belanger, S.E.; Braunbeck, T.; Carr, G.J.; Cenijn, P.; Fochtman, P.; Gourmelon, A.; Hübler, N. OECD Validation Study to Assess Intra-and Inter-Laboratory Reproducibility of the Zebrafish Embryo Toxicity Test for Acute Aquatic Toxicity Testing. Regul. Toxicol. Pharmacol. 2014, 69, 496–511. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Ligands | Docking Scores | Important Binding Site Residues |
|---|---|---|---|
| Acetylcholine Esterase (PDB ID:1GQR) | |||
| 1 | BC | 27.7 | Trp81, Gly114, Gly115, Gly116, Val126, Glu196, Trp276, Leu279, Asp282, Ser283, Phe328, Gly438 and Ile441 |
| 2 | Rivastigmine (Bound ligand) | −7.3 | |
| Acetylcholine Esterase (PDB ID:1QTI) | |||
| 1 | BC | 14.1 | Val68, Asp69, Glu70, Gly114, Gly115, Glu196, Trp276, and Gly438 |
| 2 | Galanthamine (Bound ligand) | −9.8 | |
| Compounds | Vmax (µM/min) | Km (mM) | AICc | R2 | Type of Inhibition |
|---|---|---|---|---|---|
| BC | 642.9 | 0.19 | 160.2 | 0.986 | Competitive type |
| Donepezil | 107.8 | 0.18 | 85.2 | 0.921 | Mixed type |
| Groups | AChE Activity (μmol of AChI/min/mg of Protein) | AP Activity (mM) |
|---|---|---|
| Naïve (E3 medium) | 0.21 ± 0.03 | 0.9 ± 0.02 |
| BC (137.5) | 0.23 ± 0.06 | 1.3 ± 0.01 |
| BC (275) | 0.25 ± 0.02 | 1.1 ± 0.03 |
| BC (550) | 0.20 ± 0.04 | 1.2 ± 0.02 |
| BC (1100) | 0.29 ± 0.08 | 1.4 ± 0.04 |
| BC (2200) | 0.52 ± 0.13 a | 1.8 ± 0.08 a |
| DMSO (10%) | 0.43 ± 0.08 a | 2.4 ± 0.12 a |
| Donepezil (5 ng/mL) | 0.13 ± 0.07 b | 1.1 ± 0.03 b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muthuraman, A.; Ramesh, M.; Mustaffa, F.; Nadeem, A.; Nishat, S.; Paramakrishnan, N.; Lim, K.G. In Silico and In Vitro Methods in the Characterization of Beta-Carotene as Pharmaceutical Material via Acetylcholine Esterase Inhibitory Actions. Molecules 2023, 28, 4358. https://doi.org/10.3390/molecules28114358
Muthuraman A, Ramesh M, Mustaffa F, Nadeem A, Nishat S, Paramakrishnan N, Lim KG. In Silico and In Vitro Methods in the Characterization of Beta-Carotene as Pharmaceutical Material via Acetylcholine Esterase Inhibitory Actions. Molecules. 2023; 28(11):4358. https://doi.org/10.3390/molecules28114358
Chicago/Turabian StyleMuthuraman, Arunachalam, Muthusamy Ramesh, Fazlina Mustaffa, Ahmed Nadeem, Shamama Nishat, Nallupillai Paramakrishnan, and Khian Giap Lim. 2023. "In Silico and In Vitro Methods in the Characterization of Beta-Carotene as Pharmaceutical Material via Acetylcholine Esterase Inhibitory Actions" Molecules 28, no. 11: 4358. https://doi.org/10.3390/molecules28114358
APA StyleMuthuraman, A., Ramesh, M., Mustaffa, F., Nadeem, A., Nishat, S., Paramakrishnan, N., & Lim, K. G. (2023). In Silico and In Vitro Methods in the Characterization of Beta-Carotene as Pharmaceutical Material via Acetylcholine Esterase Inhibitory Actions. Molecules, 28(11), 4358. https://doi.org/10.3390/molecules28114358