
Benzoylaconitine Alleviates Progression of Psoriasis via Suppressing STAT3 Phosphorylation in Keratinocytes
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. BAC Restrains the Abnormal Proliferation but Not the Survival of HaCaT Cells
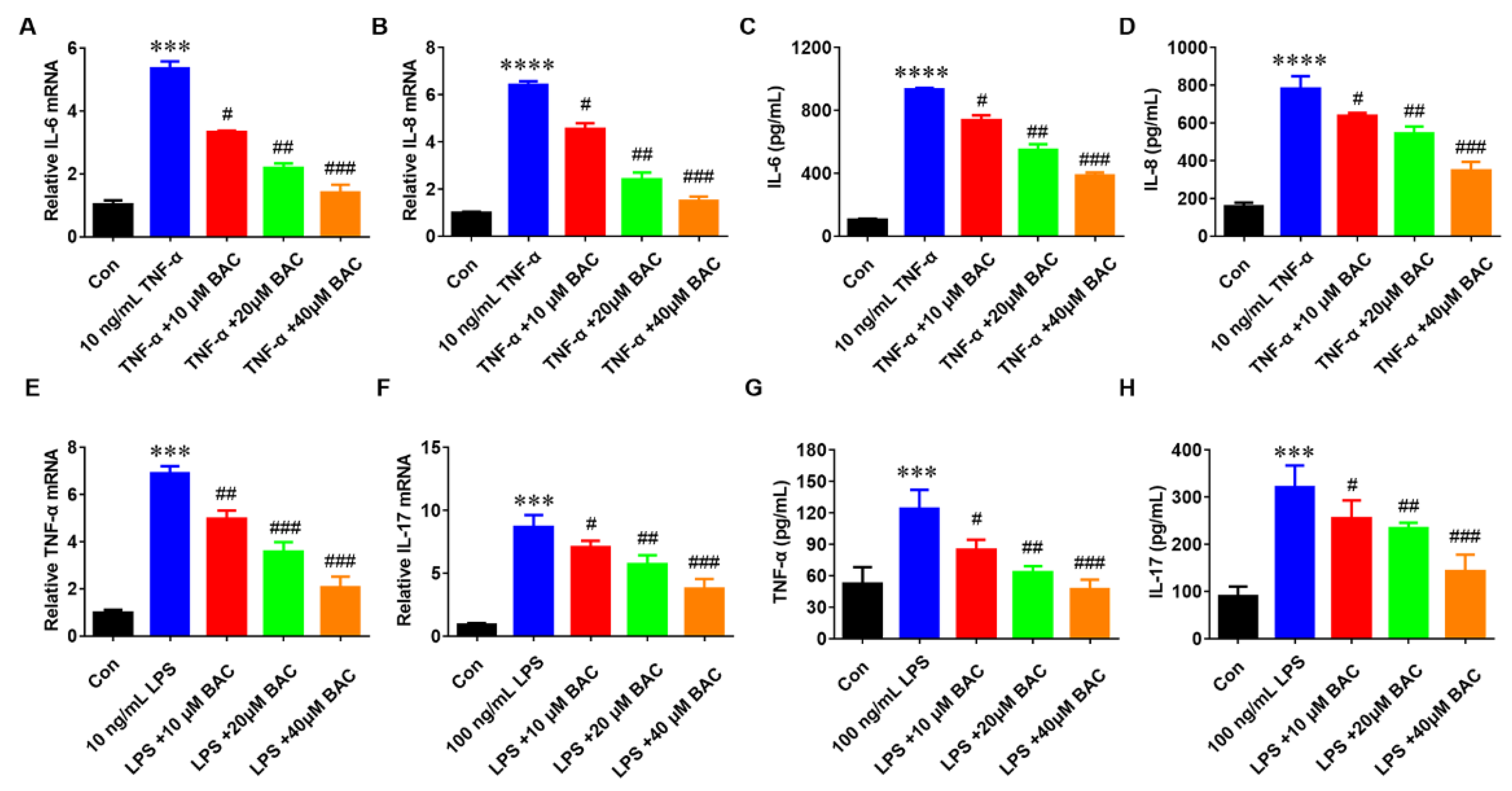
2.2. BAC Suppressed the Production of Inflammatory Cytokines in TNF-α/LPS-Induced HaCaT Cells
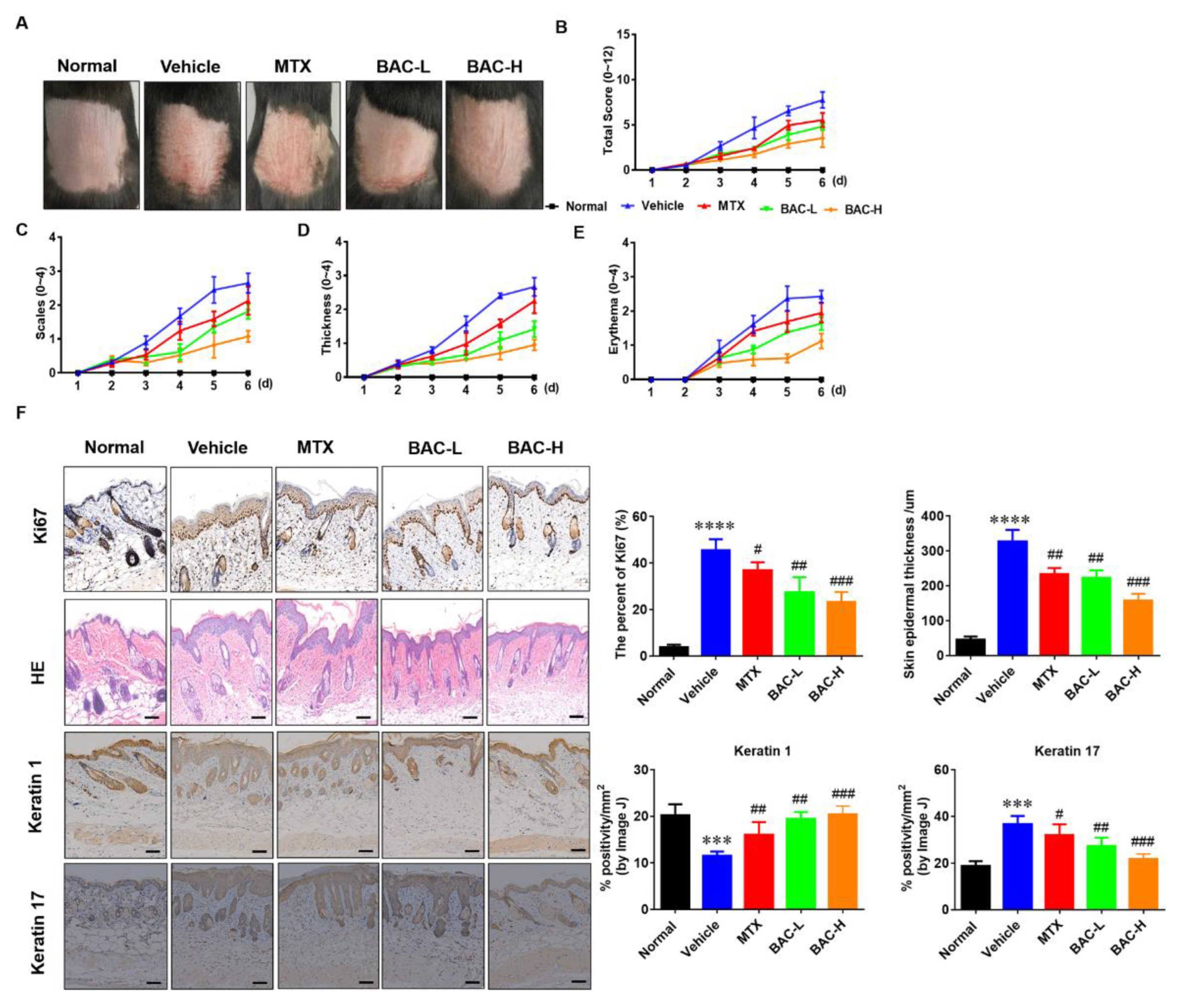
2.3. BAC Attenuated IMQ-Induced Murine Psoriasis-like Skin Inflammation
2.4. BAC Inhibited the Th17 Cells Accumulation in Model Mice
2.5. BAC Reduced the Release of Inflammatory Cytokines in IMQ-Induced Mouse Skin
2.6. Safety of BAC Administration
2.7. BAC-Regulated STAT3 Pathways in Keratinocytes
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Detection of Cell Viability
4.3. RNA Extraction and RT-qPCR
4.4. Measurement of Inflammatory Cytokines by ELISA
4.5. IMQ-Induced Psoriasis-like Skin Inflammation in Mice
4.6. Skin Cell Preparation and Flow Cytometry
4.7. Western Blotting
4.8. Skin Histopathological Analysis
4.9. Drug Safety Analyses
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Fabian, E.; Oesch, F.; Ott, K.; Landsiedel, R.; van Ravenzwaay, B. A protocol to determine dermal absorption of xenobiotica through human skin in vitro. Arch. Toxicol. 2017, 91, 1497–1511. [Google Scholar] [CrossRef] [PubMed]
- Helmick, C.G.; Lee-Han, H.; Hirsch, S.C.; Baird, T.L.; Bartlett, C.L. Prevalence of psoriasis among adults in the US 2003–2006 and 2009–2010 national health and nutrition examination surveys. Am. J. Prev. Med. 2014, 4, 37–45. [Google Scholar] [CrossRef] [PubMed]
- Benhadou, F.; Mintoff, D.; del Marmol, V. Psoriasis: Keratinocytes or immune cells-which is the trigger? Dermatology 2019, 235, 91–100. [Google Scholar] [CrossRef] [PubMed]
- Garzorz-Stark, N.; Eyerich, K. Psoriasis pathogenesis: Keratinocytes are back in the spotlight. J. Investig. Dermatol. 2019, 139, 995–996. [Google Scholar] [CrossRef]
- Michalek, I.M.; Loring, B.; John, S.M. A systematic review of worldwide epidemiology of psoriasis. J. Eur. Acad. Dermatol. 2017, 31, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Gowd, B.H.J.; Ahmed, M.G.; Hani, U.; Kesharwani, P.; Wahab, S.; Paul, K. Microneedles as a momentous platform for psoriasis therapy and diagnosis: A state-of-the-art review. Int. J. Pharm. 2023, 632, 122591. [Google Scholar] [CrossRef]
- Mortier, C.; Gracey, E.; Coudenys, J.; Manuello, T.; Decruy, T.; Maelegheer, M.; Stappers, F.; Gilis, E.; Gaublomme, D.; Van Hoorebeke, L.; et al. RORγt inhibition ameliorates IL-23 driven experimental psoriatic arthritis by predominantly modulating γδ-T cells. Rheumatology 2023, kead022. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Q.; Yu, J.; Zhou, H.; Wang, X.; Zhang, C.; Hu, J.; Hu, Y.; Zheng, H.; Zeng, F.; Yue, C.; et al. Intestinal dysbiosis exacerbates the pathogenesis of psoriasis-like phenotype through changes in fatty acid metabolism. Signal Transduct. Target. Ther. 2023, 8, 40. [Google Scholar] [CrossRef] [PubMed]
- Moos, S.; Mohebiany, A.N.; Waisman, A.; Kurschus, F.C. Imiquimod-induced psoriasis in mice depends on the IL-17 signaling of keratinocytes. J. Investig. Dermatol. 2019, 139, 1110–1117. [Google Scholar] [CrossRef]
- Alexander, H.; Nestle, F.O. Pathogenesis and immunotherapy in cutaneous psoriasis: What can rheumatologists learn? Curr. Opin. Rheumatol. 2017, 29, 71–78. [Google Scholar] [CrossRef]
- Cintra, G.A.D.; Pinto, L.A.; Calixto, G.M.F.; Soares, C.P.; Von Zuben, E.D.; Scarpa, M.V.; Gremiao, M.P.D.; Chorilli, M. Bioadhesive surfactant systems for methotrexate skin delivery. Molecules 2016, 21, 231. [Google Scholar] [CrossRef]
- Boehncke, W.H.; Schon, M.P. Psoriasis. Lancet 2015, 386, 983–994. [Google Scholar] [CrossRef] [PubMed]
- Mao, M.; Kuang, Y.; Chen, M.; Yan, K.; Lv, C.; Liu, P.; Lu, Y.; Chen, X.; Zhu, W.; Chen, W. The HLA-Cw*06 allele may predict the response to methotrexate (MTX) treatment in Chinese arthritis-free psoriasis patients. Arch. Dermatol. Res. 2022, 315, 1241–1247. [Google Scholar] [CrossRef]
- Atallah, E.; Grove, J.I.; Crooks, C.; Burden-The, E.; Abhishek, A.; Moreea, S.; Jordan, K.M.; Ala, A.; Hutchinson, D.; Aspinall, R.J.; et al. Risk of liver fibrosis associated with long-term methotrexate therapy may be overestimated. J. Hepatol. 2023, 78, 989–997. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhao, G.; Han, K.; Sun, D.; Zhou, N.; Song, Z.; Liu, H.; Li, J.; Li, G. Applications of molecular imprinting technology in the study of traditional Chinese medicine. Molecules 2022, 28, 301. [Google Scholar] [CrossRef]
- Zhang, Q.Q.; Chen, F.H.; Wang, F.; Di, X.M.; Li, W.; Zhang, H. A novel modulator of the renin-angiotensin system, benzoylaconitine, attenuates hypertension by targeting ACE/ACE2 in enhancing vasodilation and alleviating vascular inflammation. Front. Pharmacol. 2022, 13, 841435. [Google Scholar] [CrossRef]
- Li, S.; Li, R.; Xu, Y.X.; Baak, J.P.A.; Gao, J.H.; Shu, J.Q.; Jing, L.J.; Meng, X.L.; Wen, C.B.; Gan, Y.X.; et al. Traditional Chinese medicine aconiti radix cocta improves rheumatoid arthritis via suppressing COX-1 and COX-2. Evid. Based Complement. Alternat. Med. 2021, 2021, 5523870. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.X.; Cheng, B.F.; Lian, J.J.; Guo, D.D.; Qin, J.W.; Zhang, Y.B.; Yang, H.J.; Wang, M.; Wang, L.; Feng, Z.W. Liquiritin, a flavone compound from licorice, inhibits IL-1 beta-induced inflammatory responses in SW982 human synovial cells. J. Funct. Foods 2017, 33, 142–148. [Google Scholar] [CrossRef]
- Zhou, C.; Gao, J.; Ji, H.; Li, W.; Xing, X.; Liu, D.; Guo, Q.; Zhou, L.; Jing, F. Benzoylaconine modulates LPS-induced responses through inhibition of Toll-like receptor-mediated NF-κB and MAPK signaling in RAW264.7 cells. Inflammation 2021, 44, 2018–2032. [Google Scholar] [CrossRef]
- Miroddi, M.; Navarra, M.; Calapai, F.; Mancari, F.; Giofre, S.V.; Gangemi, S.; Calapai, G. Review of clinical pharmacology of aloe vera L. in the treatment of psoriasis. Phytother. Res. 2015, 29, 648–655. [Google Scholar] [CrossRef]
- Martinez, G.J.; Nurieva, R.I.; Yang, X.O.; Dong, C. Regulation and function of proinflammatory TH17 cells. Ann. N. Y. Acad. Sci. 2008, 1143, 188–211. [Google Scholar] [CrossRef] [PubMed]
- Wilson, N.J.; Boniface, K.; Chan, J.R.; McKenzie, B.S.; Blumenschein, W.M.; Mattson, J.D.; Basham, B.; Smith, K.; Chen, T.; Morel, F.; et al. Development, cytokine profile and function of human interleukin 17-producing helper T cells. Nat. Immunol. 2007, 8, 950–957. [Google Scholar] [CrossRef] [PubMed]
- Girolomoni, G.; Strohal, R.; Puig, L.; Bachelez, H.; Barker, J.; Boehncke, W.H.; Prinz, J.C. The role of IL-23 and the IL-23/T(H)17 immune axis in the pathogenesis and treatment of psoriasis. J. Eur. Acad. Dermatol. 2017, 31, 1616–1626. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Wang, Z.; Liao, B.; Sun, Z.; Zhu, P. Anti-inflammatory and chondroprotective effects of platelet-derived growth factor-BB on osteoarthritis rat models. J. Gerontol. A Biol. Sci. Med. Sci. 2022, 78, 51–59. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, S.H.; Zuo, Y.G. Paradoxical phenomena of bullous pemphigoid induced and treated by identical biologics. Front. Immunol. 2023, 13, 1050373. [Google Scholar] [CrossRef]
- Perera, G.K.; Di Meglio, P.; Nestle, F.O. Psoriasis. Annu. Rev. Pathol. 2012, 7, 385–422. [Google Scholar] [CrossRef] [PubMed]
- Callon, S.; Brugel, M.; Botsen, D.; Royer, B.; Slimano, F.; Feliu, C.; Gozalo, C.; Konecki, C.; Devie, B.; Carlier, C.; et al. Renal impairment and abnormal liver function tests in pre-therapeutic phenotype-based DPD deficiency screening using uracilemia: A comprehensive population-based study in 1138 patients. Ther. Adv. Med. Oncol. 2023, 15, 17588359221148536. [Google Scholar] [CrossRef]
- Hirano, T.; Ishihara, K.; Hibi, M. Roles of STAT3 in mediating the cell growth, differentiation and survival signals relayed through the IL -6 family of cytokine receptors. Oncogene 2000, 19, 2548–2556. [Google Scholar] [CrossRef]
- Edson-Heredia, E.; Anderson, S.; Guo, J.Y.; Zhu, B.J.; Malatestinic, W.N.; Wine-Lee, L.; Swanson, E. Real-world claims analyses of comorbidity burden, treatment pattern, healthcare resource utilization, and costs in pediatric psoriasis. Adv. Ther. 2021, 38, 3948–3961. [Google Scholar] [CrossRef]
- Leng, H.; Pu, L.; Xu, L.J.; Shi, X.; Ji, J.; Chen, K. Effects of aloe polysaccharide, a polysaccharide extracted from Aloe vera, on TNF-alpha-induced HaCaT cell proliferation and the underlying mechanism in psoriasis. Mol. Med. Rep. 2018, 18, 3537–3543. [Google Scholar]
- Liu, Y.; Sun, H.; Li, C.; Pu, Z.; Wu, Z.; Xu, M.; Li, X.; Zhang, Y.; Li, H.; Dong, J.; et al. Comparative HPLC-MS/MS-based pharmacokinetic studies of multiple diterpenoid alkaloids following the administration of Zhenwu Tang and Radix Aconiti Lateralis Praeparata extracts to rats. Xenobiotica 2021, 51, 345–354. [Google Scholar] [CrossRef]
- Parisi, R.; Symmons, D.P.M.; Griffiths, C.E.M.; Ashcroft, D.M.; Management, I. Global Epidemiology of Psoriasis: A Systematic Review of Incidence and Prevalence. J. Investig. Dermatol. 2013, 133, 377–385. [Google Scholar] [CrossRef]
- Herman, A.; Herman, A.P. Topically used herbal products for the treatment of psoriasis-mechanism of action, Drug Delivery, Clinical Studies. Planta. Med. 2016, 82, 1447–1455. [Google Scholar] [CrossRef]
- Christmann, C.; Zenker, S.; Martens, L.; Hubner, J.; Loser, K.; Vogl, T.; Roth, J. Interleukin 17 promotes expression of alarmins S100A8 and S100A9 during the inflammatory response of keratinocytes. Front. Immunol. 2021, 11, 599947. [Google Scholar] [CrossRef] [PubMed]
- Shannon, E.; Noveck, R.; Sandoval, F.; Kamath, B. Thalidomide suppressed IL-1 beta while enhancing TNF-alpha and IL-10, when cells in whole blood were stimulated with lipopolysaccharide. Immunopharm. Immunot. 2008, 30, 447–457. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Xiao, Y.; Yin, M.Z.; Zhou, X.C.; Wu, L.S.; Chen, W.Q.; Luo, Y.; Kuang, Y.H.; Zhu, W. Polyethylene glycol ointment alleviates psoriasis-like inflammation through down-Regulating the Function of Th17 Cells and MDSCs. Front. Med. 2021, 7, 560579. [Google Scholar] [CrossRef]
- Kim, N.; Lee, S.; Kang, J.; Choi, Y.A.; Lee, B.; Kwon, T.K.; Jang, Y.H.; Kim, S.H. Hispidulin alleviates imiquimod-induced psoriasis-like skin inflammation by inhibiting splenic Th1/Th17 cell population and keratinocyte activation. Int. Immunopharmacol. 2020, 87, 106767. [Google Scholar] [CrossRef]
- Tang, K.W.; Lin, Z.C.; Wang, P.W.; Alalaiwe, A.; Tseng, C.H.; Fang, J.Y. Facile skin targeting of a thalidomide analog containing benzyl chloride moiety alleviates experimental psoriasis via the suppression of MAPK/NF-kappa B/AP-1 phosphorylation in keratinocytes. J. Dermatol. Sci. 2020, 99, 90–99. [Google Scholar] [CrossRef] [PubMed]
- Sano, S.; Chan, K.S.; Carbajal, S.; Clifford, J.; Peavey, M.; Kiguchi, K.; Itami, S.; Nickoloff, B.J.; DiGiovanni, J. Stat3 links activated keratinocytes and immunocytes required for development of psoriasis in a novel transgenic mouse model. Nat. Med. 2005, 11, 43–49. [Google Scholar] [CrossRef]
- Rasheed, Z.B.M.; Lee, Y.S.; Kim, S.H.; Rai, R.K.; Ruano, C.S.M.; Anucha, E.; Sullivan, M.H.F.; MacIntyre, D.A.; Bennett, P.R.; Sykes, L. Differential response of gestational tissues to TLR3 Viral Priming Prior to Exposure to Bacterial TLR2 and TLR2/6 Agonists. Front. Immunol. 2020, 11, 1–27. [Google Scholar] [CrossRef]
- Cai, Y.H.; Shen, X.Y.; Ding, C.L.; Qi, C.J.; Li, K.J.; Li, X.; Jala, V.R.; Zhang, H.G.; Wang, T.A.; Zheng, J.; et al. Pivotal role of dermal IL-17-producing gamma delta T cells in skin inflammation. Immunity 2011, 35, 649. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Guo, D.; Wang, Q.; Li, A.; Yin, S.; Li, S.; Li, Y.; Wang, B.; Guo, T.; Feng, S. Benzoylaconitine Alleviates Progression of Psoriasis via Suppressing STAT3 Phosphorylation in Keratinocytes. Molecules 2023, 28, 4473. https://doi.org/10.3390/molecules28114473
Li Y, Guo D, Wang Q, Li A, Yin S, Li S, Li Y, Wang B, Guo T, Feng S. Benzoylaconitine Alleviates Progression of Psoriasis via Suppressing STAT3 Phosphorylation in Keratinocytes. Molecules. 2023; 28(11):4473. https://doi.org/10.3390/molecules28114473
Chicago/Turabian StyleLi, Yuanbo, Dandan Guo, Qianqian Wang, Aifang Li, Sugai Yin, Shuxuan Li, Yalan Li, Baiyan Wang, Tao Guo, and Shuying Feng. 2023. "Benzoylaconitine Alleviates Progression of Psoriasis via Suppressing STAT3 Phosphorylation in Keratinocytes" Molecules 28, no. 11: 4473. https://doi.org/10.3390/molecules28114473
APA StyleLi, Y., Guo, D., Wang, Q., Li, A., Yin, S., Li, S., Li, Y., Wang, B., Guo, T., & Feng, S. (2023). Benzoylaconitine Alleviates Progression of Psoriasis via Suppressing STAT3 Phosphorylation in Keratinocytes. Molecules, 28(11), 4473. https://doi.org/10.3390/molecules28114473