

Dietary Supplementation of Aspirin Promotes Drosophila Defense against Viral Infection

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
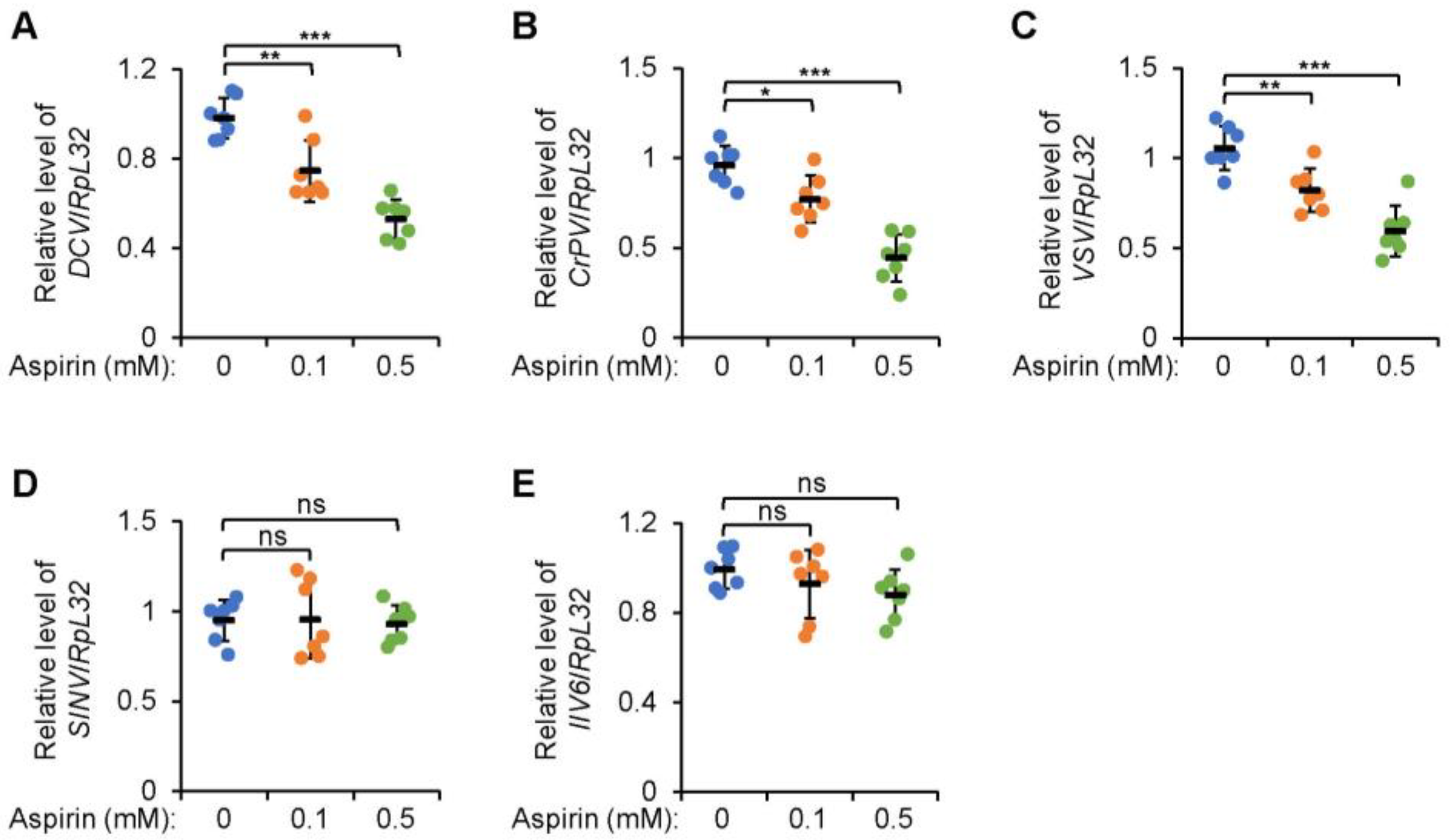
2.1. Additional Aspirin Leads to Decreased Viral Titers in Drosophila S2 Cells
2.2. Viral Proliferation Is Limited in Adult Flies with Dietary Supplementation of Aspirin
2.3. Dietary Supplementation of Aspirin Enhances Fly Survival after Viral Infection
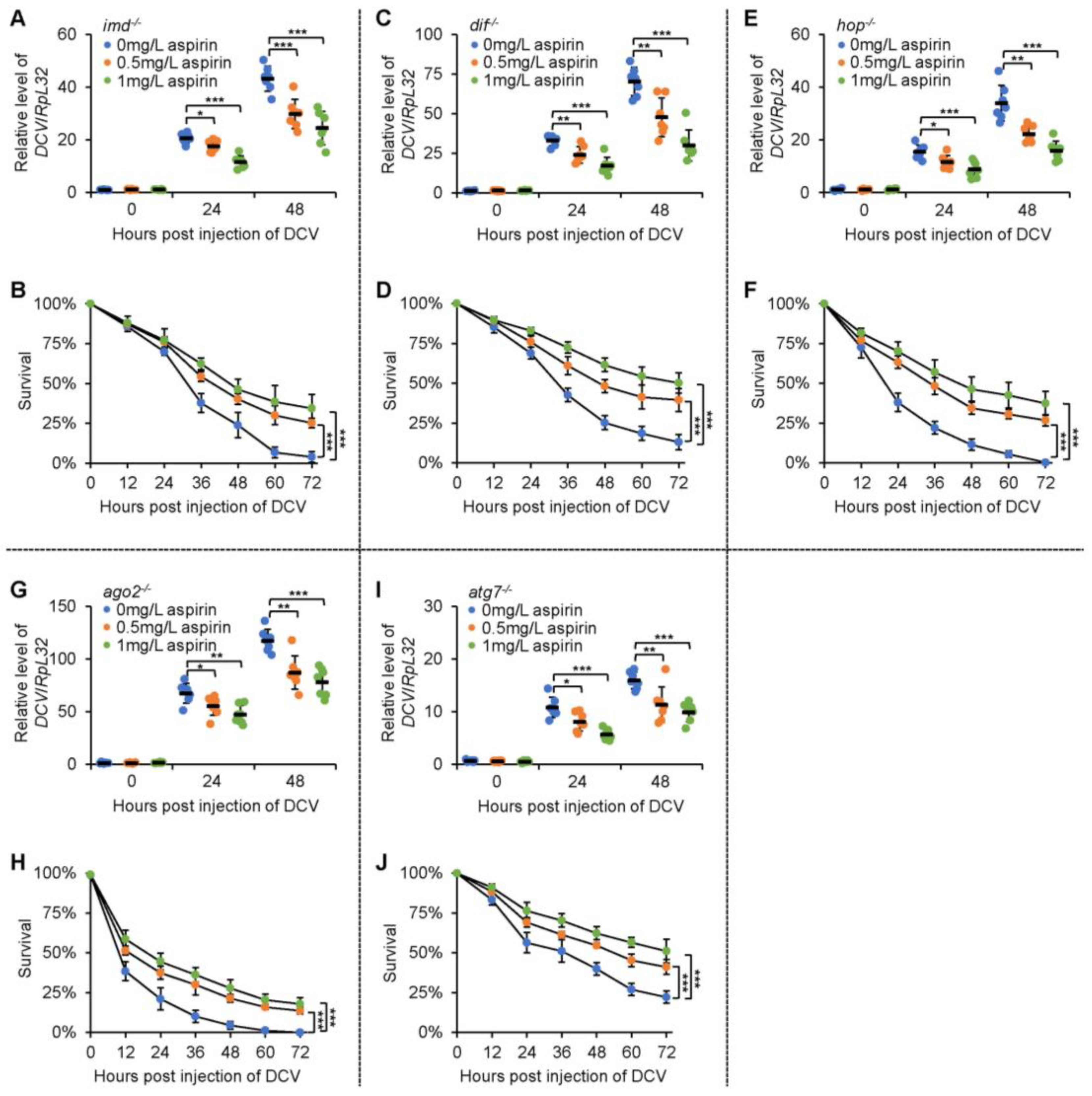
2.4. Aspirin Modulates Fly Survival and Viral Titers after Infection in an IMD-Independent Manner
2.5. Aspirin Relies on Sting and Relish to Regulate Fly Survival and Viral Replication after Infection
2.6. Additional Aspirin Increases the Expressions of STING-Induced Genes
3. Discussion
4. Significance and Limitations of this Study
5. Materials and Methods
5.1. Fly Strains and Husbandry
5.2. S2 Cell Manipulation
5.3. Viral Infection in Flies and Survival Rate Assays
5.4. RT-qPCR Assays
5.5. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Martinez, J.P.; Sasse, F.; Bronstrup, M.; Diez, J.; Meyerhans, A. Antiviral drug discovery: Broad-spectrum drugs from nature. Nat. Prod. Rep. 2015, 32, 29–48. [Google Scholar] [CrossRef]
- Andreolla, A.P.; Borges, A.A.; Bordignon, J.; Dos Santos, C.N.D. Mayaro virus: The state-of-the-art for antiviral drug development. Viruses 2022, 14, 1787. [Google Scholar] [CrossRef]
- Ma-Lauer, Y.; Lei, J.; Hilgenfeld, R.; von Brunn, A. Virus-host interactomes--antiviral drug discovery. Curr. Opin. Virol. 2012, 2, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Yoon, B.K.; Jeon, W.Y.; Sut, T.N.; Cho, N.J.; Jackman, J.A. Stopping membrane-enveloped viruses with nanotechnology strategies: Toward antiviral drug development and pandemic preparedness. ACS Nano 2021, 15, 125–148. [Google Scholar] [CrossRef]
- Park, S.; Tae, H.; Cho, N.J. Biophysical measurement strategies for antiviral drug development: Recent progress in virus-mimetic platforms down to the single particle level. Acc. Chem. Res. 2021, 54, 3204–3214. [Google Scholar] [CrossRef]
- de Chassey, B.; Meyniel-Schicklin, L.; Aublin-Gex, A.; Andre, P.; Lotteau, V. New horizons for antiviral drug discovery from virus-host protein interaction networks. Curr. Opin. Virol. 2012, 2, 606–613. [Google Scholar] [CrossRef]
- De Clercq, E. Antiviral drug discovery and development: Where chemistry meets with biomedicine. Antivir. Res. 2005, 67, 56–75. [Google Scholar] [CrossRef]
- Martins, N.; Imler, J.L.; Meignin, C. Discovery of novel targets for antivirals: Learning from flies. Curr. Opin. Virol. 2016, 20, 64–70. [Google Scholar] [CrossRef]
- Benoit, I.; Di Curzio, D.; Civetta, A.; Douville, R.N. Drosophila as a model for human viral neuroinfections. Cells 2022, 11, 2685. [Google Scholar] [CrossRef] [PubMed]
- Pandey, U.B.; Nichols, C.D. Human disease models in Drosophila melanogaster and the role of the fly in therapeutic drug discovery. Pharmacol. Rev. 2011, 63, 411–436. [Google Scholar] [CrossRef] [PubMed]
- Ugur, B.; Chen, K.; Bellen, H.J. Drosophila tools and assays for the study of human diseases. Dis. Model Mech. 2016, 9, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Adamson, A.L.; Chohan, K.; Swenson, J.; LaJeunesse, D. A Drosophila model for genetic analysis of influenza viral/host interactions. Genetics 2011, 189, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.; Hou, W.; Dakshanamurthy, S.; Tang, Q. Host targeted antiviral (HTA): Functional inhibitor compounds of scaffold protein RACK1 inhibit herpes simplex virus proliferation. Oncotarget 2019, 10, 3209–3226. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.G.; Huang, W.; Lee, H.; van de Leemput, J.; Kane, M.A.; Han, Z. Characterization of SARS-CoV-2 proteins reveals Orf6 pathogenicity, subcellular localization, host interactions and attenuation by Selinexor. Cell Biosci. 2021, 11, 1–12. [Google Scholar] [CrossRef]
- Menter, D.G.; Bresalier, R.S. An aspirin a day: New pharmacological developments and cancer chemoprevention. Annu. Rev. Pharmacol. Toxicol. 2023, 63, 165–186. [Google Scholar] [CrossRef]
- Sehanobish, E.; Asad, M.; Barbi, M.; Porcelli, S.A.; Jerschow, E. Aspirin actions in treatment of NSAID-exacerbated respiratory disease. Front. Immunol. 2021, 12, 695815. [Google Scholar] [CrossRef]
- Glatthaar-Saalmuller, B.; Mair, K.H.; Saalmuller, A. Antiviral activity of aspirin against RNA viruses of the respiratory tract-an in vitro study. Influenza Other Respir. Viruses 2017, 11, 85–92. [Google Scholar] [CrossRef]
- Pereira, C.F.; Paridaen, J.T.M.L.; Rutten, K.; Huigen, M.C.D.G.; van de Bovenkamp, M.; Middel, J.; Beerens, N.; Berkhout, B.; Schuurman, R.; Marnett, L.J.; et al. Aspirin-like molecules that inhibit human immunodeficiency virus 1 replication. Antivir. Res. 2003, 58, 253–263. [Google Scholar] [CrossRef]
- Rivas-Estilla, A.M.; Bryan-Marrugo, O.L.; Trujillo-Murillo, K.; Perez-Ibave, D.; Charles-Nino, C.; Pedroza-Roldan, C.; Rios-Ibarra, C.; Ramirez-Valles, E.; Ortiz-Lopez, R.; Islas-Carbajal, M.C.; et al. Cu/Zn superoxide dismutase (SOD1) induction is implicated in the antioxidative and antiviral activity of acetylsalicylic acid in HCV-expressing cells. Am. J Physiol. Gastrointest. Liver Physiol. 2012, 302, G1264–G1273. [Google Scholar] [CrossRef]
- Zhu, Y.; Cai, Q.; Zheng, X.; Liu, L.; Hua, Y.; Du, B.; Zhao, G.; Yu, J.; Zhuo, Z.; Xie, Z.; et al. Aspirin positively contributes to Drosophila intestinal homeostasis and delays aging through targeting Imd. Aging Dis. 2021, 12, 1821–1834. [Google Scholar] [CrossRef]
- Georgel, P.; Naitza, S.; Kappler, C.; Ferrandon, D.; Zachary, D.; Swimmer, C.; Kopczynski, C.; Duyk, G.; Reichhart, J.M.; Hoffmann, J.A. Drosophila immune deficiency (IMD) is a death domain protein that activates antibacterial defense and can promote apoptosis. Dev. Cell 2001, 1, 503–514. [Google Scholar] [CrossRef]
- Costa, A.; Jan, E.; Sarnow, P.; Schneider, D. The Imd pathway is involved in antiviral immune responses in Drosophila. PLoS ONE 2009, 4, e7436. [Google Scholar] [CrossRef] [PubMed]
- Zambon, R.A.; Nandakumar, M.; Vakharia, V.N.; Wu, L.P. The Toll pathway is important for an antiviral response in Drosophila. Proc. Natl. Acad. Sci. USA 2005, 102, 7257–7262. [Google Scholar] [CrossRef] [PubMed]
- Dostert, C.; Jouanguy, E.; Irving, P.; Troxler, L.; Galianna-Arnoux, D.; Hetru, C.; Hoffmann, J.A.; Imler, J.L. The Jak-STAT signaling pathway is required but not sufficient for the antiviral response of drosophila. Nat. Immunol. 2005, 6, 946–953. [Google Scholar] [CrossRef]
- Deddouche, S.; Matt, N.; Budd, A.; Mueller, S.; Kemp, C.; Galiana-Arnoux, D.; Dostert, C.; Antoniewski, C.; Hoffmann, J.A.; Imler, J.L. The DExD/H-box helicase Dicer-2 mediates the induction of antiviral activity in drosophila. Nat. Immunol. 2008, 9, 1425–1432. [Google Scholar] [CrossRef]
- Lamiable, O.; Arnold, J.; Faria, I.; Olmo, R.P.; Bergami, F.; Meignin, C.; Hoffmann, J.A.; Marques, J.T.; Imler, J.L. Analysis of the contribution of hemocytes and autophagy to Drosophila antiviral immunity. J. Virol. 2016, 90, 5415–5426. [Google Scholar] [CrossRef]
- Goto, A.; Okado, K.; Martins, N.; Cai, H.; Barbier, V.; Lamiable, O.; Troxler, L.; Santiago, E.; Kuhn, L.; Paik, D.; et al. The kinase IKKbeta regulates a STING- and NF-kappaB-dependent antiviral response pathway in Drosophila. Immunity 2018, 49, 225–234. [Google Scholar] [CrossRef]
- Cai, H.; Holleufer, A.; Simonsen, B.; Schneider, J.; Lemoine, A.; Gad, H.H.; Huang, J.; Huang, J.; Chen, D.; Peng, T.; et al. 2′3′-cGAMP triggers a STING- and NF-kappaB-dependent broad antiviral response in Drosophila. Sci. Signal. 2020, 13, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Geiger, N.; Konig, E.M.; Oberwinkler, H.; Roll, V.; Diesendorf, V.; Fahr, S.; Obernolte, H.; Sewald, K.; Wronski, S.; Steinke, M.; et al. Acetylsalicylic acid and salicylic acid inhibit SARS-CoV-2 replication in precision-cut lung slices. Vaccines 2022, 10, 1619. [Google Scholar] [CrossRef]
- Song, C.; Zhu, C.; Wu, Q.; Qi, J.; Gao, Y.; Zhang, Z.; Gaur, U.; Yang, D.; Fan, X.; Yang, M. Metabolome analysis of effect of aspirin on Drosophila lifespan extension. Exp. Gerontol. 2017, 95, 54–62. [Google Scholar] [CrossRef]
- Cai, Q.; Ji, S.; Li, M.; Zheng, S.; Zhou, X.; Huo, H.; Deng, S.; Zhu, J.; Li, D.; Xie, Z. Theaflavin-regulated Imd condensates control Drosophila intestinal homeostasis and aging. iScience 2021, 24, 102150. [Google Scholar] [CrossRef] [PubMed]
- Kounatidis, I.; Ligoxygakis, P. Drosophila as a model system to unravel the layers of innate immunity to infection. Open Biol. 2012, 2, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Cai, Q.; Guo, H.; Fang, R.; Hua, Y.; Zhu, Y.; Zheng, X.; Yan, J.; Wang, J.; Hu, Y.; Zhang, C.; et al. A Toll-dependent Bre1/Rad6-cact feedback loop in controlling host innate immune response. Cell Rep. 2022, 41, 111795. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, F.; Qadeer, A.; Xie, Y.; Jin, Y.; Li, Q.; Xiao, Y.; She, K.; Zheng, X.; Li, J.; Ji, S.; et al. Dietary Supplementation of Aspirin Promotes Drosophila Defense against Viral Infection. Molecules 2023, 28, 5300. https://doi.org/10.3390/molecules28145300
Kong F, Qadeer A, Xie Y, Jin Y, Li Q, Xiao Y, She K, Zheng X, Li J, Ji S, et al. Dietary Supplementation of Aspirin Promotes Drosophila Defense against Viral Infection. Molecules. 2023; 28(14):5300. https://doi.org/10.3390/molecules28145300
Chicago/Turabian StyleKong, Fanrui, Abdul Qadeer, Yali Xie, Yiheng Jin, Qingyang Li, Yihua Xiao, Kan She, Xianrui Zheng, Jiashu Li, Shanming Ji, and et al. 2023. "Dietary Supplementation of Aspirin Promotes Drosophila Defense against Viral Infection" Molecules 28, no. 14: 5300. https://doi.org/10.3390/molecules28145300
APA StyleKong, F., Qadeer, A., Xie, Y., Jin, Y., Li, Q., Xiao, Y., She, K., Zheng, X., Li, J., Ji, S., & Zhu, Y. (2023). Dietary Supplementation of Aspirin Promotes Drosophila Defense against Viral Infection. Molecules, 28(14), 5300. https://doi.org/10.3390/molecules28145300