
An Untargeted Metabolomics Approach to Study the Variation between Wild and Cultivated Soybeans
 , and
, and
Abstract
:
1. Introduction
2. Results and Discussion
2.1. Identification of Compounds
2.2. Comparison of Metabolites among Cultivars
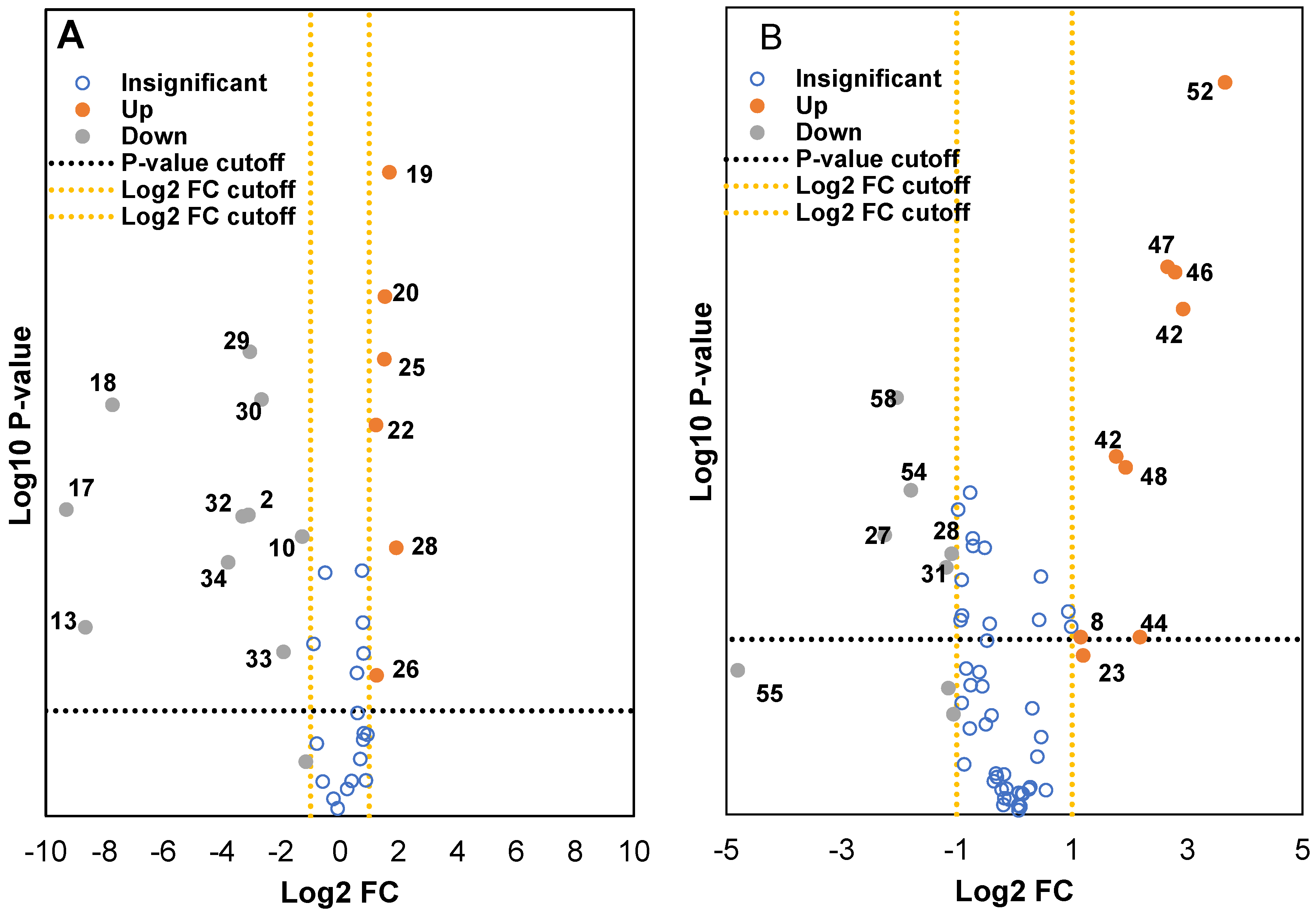
2.3. Classification of Wild and Cultivated Soybeans Using Principal Component Analysis (PCA) and Volcano Plots
3. Materials and Methods
3.1. Solvents and Materials
3.2. Samples
3.3. Extraction of Metabolites
3.4. Data Acquisition
3.5. Identification of Compounds
3.6. Data Processing
3.7. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Available online: https://ourworldindata.org/soy (accessed on 22 March 2023).
- do Prado, F.G.; Pagnoncelli, M.G.B.; de Melo Pereira, G.V.; Karp, S.G.; Soccol, C.R. Fermented soy products and their potential health benefits: A Review. Microorganisms 2022, 10, 1606. [Google Scholar] [CrossRef]
- Verma, A.; Chandrakar, M.R. Compound growth rate (CGR) of area, production & productivity of soybean in Chhattisgarh state. Pharma Innov. 2023, 12, 2774–2779. [Google Scholar]
- Available online: https://www.nass.usda.gov/Newsroom/2023/01-12-2023.php (accessed on 21 March 2023).
- Kim, I.S.; Kim, C.H.; Yang, W.S. Physiologically active molecules and functional properties of soybeans in human health—A current perspective. Int. J. Mol. Sci. 2021, 22, 4054. [Google Scholar] [CrossRef]
- Erb, M.; Kliebenstein, D.J. Plant secondary metabolites as defenses, regulators, and primary metabolites: The blurred functional trichotomy. Plant Physiol. 2020, 184, 39–52. [Google Scholar] [CrossRef]
- Fan, M.; Lian, W.; Li, T.; Fan, Y.; Rao, Z.; Li, Y.; Qian, H.; Zhang, H.; Wu, G.; Qi, X.; et al. Metabolomics approach reveals discriminatory metabolites associating with the blue pigments from Vaccinium bracteatum Thunb. Leaves at different growth stages. Ind. Crops Prod. 2020, 147, 112252. [Google Scholar] [CrossRef]
- Li, P.; Xiao, Z.; Sun, J.; Oyang, X.; Xie, X.; Li, Z.; Tian, X.; Li, J. Metabolic regulations in lettuce root under combined exposure to perfluorooctanoic acid and perfluorooctane sulfonate in hydroponic media. Sci. Total Environ. 2020, 726, 138382. [Google Scholar] [CrossRef]
- Huberty, M.; Choi, Y.; Heinen, R.; Bezemer, T.M. Aboveground plant metabolomic responses to plant-soil feedbacks and herbivory. J. Ecol. 2020, 108, 1703–1712. [Google Scholar] [CrossRef]
- Wu, L.; Gao, X.; Xia, F.; Joshi, J.; Borza, T.; Wang-Pruski, G. Biostimulant and fungicidal effects of phosphite assessed by GC-TOF-MS analysis of potato leaf metabolome. Physiol. Mol. Plant Pathol. 2019, 106, 49–56. [Google Scholar] [CrossRef]
- Wang, X.; Guo, R.; Li, M.; Liu, Y.; Zhao, M.; Fu, H.; Liu, X.; Wang, S.; Shi, L. Metabolomics reveals the drought-tolerance mechanism in wild soybean (Glycine soja). Acta Physiol. Plant 2019, 41, 161. [Google Scholar] [CrossRef]
- Valette, M.; Rey, M.; Gerin, F.; Comte, G.; Wisniewski-Dyé, F. A common metabolomic signature is observed upon inoculation of rice roots with various rhizobacteria. J. Integr. Plant Biol. 2020, 62, 228–246. [Google Scholar] [CrossRef]
- Clarke, J.D.; Alexander, D.C.; Ward, D.P.; Ryals, J.A.; Mitchell, M.W.; Wulff, J.E.; Guo, L. Assessment of genetically modified soybean in relation to natural variation in the soybean seed metabolome. Sci. Rep. 2013, 3, 3082. [Google Scholar] [CrossRef] [Green Version]
- Chebrolu, K.K.; Fritschi, F.B.; Ye, S.; Krishnan, H.B.; Smith, J.R.; Gillman, J.D. Impact of heat stress during seed development on soybean seed metabolome. Metabolomics 2016, 12, 28. [Google Scholar] [CrossRef]
- Gupta, R.; Min, C.W.; Kim, S.W.; Wang, Y.; Agrawal, G.K.; Rakwal, R.; Kim, S.G.; Lee, B.W.; Ko, J.M.; Baek, I.Y.; et al. Comparative investigation of seed coats of brown- versus yellow-colored soybean seeds using an integrated proteomics and metabolomics approach. Proteomics 2015, 15, 1706–1716. [Google Scholar] [CrossRef]
- Bragagnolo, F.S.; Funari, C.S.; Ibáñez, E.; Cifuentes, A. metabolomics as a tool to study underused soy parts: In search of bioactive compounds. Foods 2021, 10, 1308. [Google Scholar] [CrossRef]
- Hoeck, J.; Fehr, W.; Murphy, P.; Welke, G. Influence of genotype and environment on isoflavone contents of soybean. Crop Sci. 2000, 40, 48–51. [Google Scholar] [CrossRef]
- Caldwell, C.R.; Britz, S.J.; Mirecki, R.M. Effect of temperature, elevated carbon dioxide, and drought during seed development on the isoflavone content of dwarf soybean [Glycine max (L.) Merrill] grown in controlled environments. J. Agric. Food Chem. 2005, 53, 1125–1129. [Google Scholar] [CrossRef]
- Joshi, T.; Qiuming, Y.; Levi, D.; Brechenmacher, L.; Valliyodan, B.; Stacey, G.; Nguyen, H.; Xu, D. SoyMetDB: The soybean metabolome database. In Proceedings of the 2010 IEEE International Conference on Bioinformatics and Biomedicine (BIBM), Hong Kong, China, 18–21 December 2010; pp. 203–208. [Google Scholar]
- John, K.M.M.; Savithiry, N.; Devanand, L. Metabolite changes in nine different soybean varieties grown under field and greenhouse conditions. Food Chem. 2016, 211, 347–355. [Google Scholar] [CrossRef] [Green Version]
- Kotha, R.R.; Natarajan, S.; Wang, D.; Luthria, D.L. Compositional analysis of non-polar and polar metabolites in 14 soybeans using spectroscopy and chromatography tools. Foods 2019, 8, 557. [Google Scholar] [CrossRef] [Green Version]
- Bi, W.; Zhao, G.; Zhou, Y. Metabolomics analysis of flavonoids in seeds and sprouts of two Chinese soybean cultivars. Sci. Rep. 2022, 12, 5541. [Google Scholar] [CrossRef]
- Mi, J.L.; Ill-Min, C.; Hunjung, K.; Mun, Y.J. High-resolution LC–ESI-TOF-mass spectrometry method for fast separation, identification, and quantification of 12 isoflavones in soybeans and soybean products. Food Chem. 2015, 176, 254–262. [Google Scholar]
- Yeon, H.L.; Bokyeong, K.; Seung-Ryul, H.; Kyun, K.; Jin, H.L. Rapid characterization of metabolites in soybean using ultra-high-performance liquid chromatography coupled with electrospray ionization quadrupole time-of-flight mass spectrometry (UPLC-ESI-Q-TOF-MS/MS) and screening for α-glucosidase inhibitory and antioxidant properties through different solvent systems. J. Food Drug Anal. 2018, 26, 277–291. [Google Scholar]
- Kenneth, D.R.S.; Aedin, C. Dietary isoflavones: Biological effects and relevance to human health. J. Nutr. 1999, 129, 758S–767S. [Google Scholar]
- Sharma, S.; Kaur, M.; Goyal, R.; Gill, B.S. Physical characteristics and nutritional composition of some new soybean (Glycine max (L.) Merrill) genotypes. J. Food Sci. Technol. 2014, 51, 551–557. [Google Scholar] [CrossRef] [Green Version]
- Patil, G.; Mian, R.; Vuong, T.; Pantalone, V.; Song, Q.; Chen, P.; Shannon, G.J.; Carter, T.C.; Nguyen, H.T. Molecular mapping and genomics of soybean seed protein: A review and perspective for the future. Theor. Appl. Genet. 2017, 130, 1975–1991. [Google Scholar] [CrossRef] [Green Version]
- Chotekajorn, A.; Hashiguchi, T.; Hashiguchi, M.; Tanaka, H.; Akashi, R. Evaluation of seed amino acid content and its correlation network analysis in wild soybean (Glycine soja) germplasm in Japan. Plant Genet. Resour. 2021, 19, 35–43. [Google Scholar] [CrossRef]
- Hyeon, H.; Xu, J.L.; Kim, J.K.; Choi, Y. Comparative metabolic profiling of cultivated and wild black soybeans reveals distinct metabolic alterations associated with their domestication. Food Res. Int. 2020, 134, 109290. [Google Scholar] [CrossRef]
- Song, J.; Wu, G.; Li, T.; Liu, C.; Li, D. Changes in the sugars, amino acids and organic acids of post-harvest spermine-treated immature vegetable soybean (Glycine max L. Merr.) as determined by 1H NMR spectroscopy. Food Prod. Process. Nutr. 2020, 2, 7. [Google Scholar] [CrossRef]
- Zarkadas, C.G.; Gagnon, C.; Gleddie, S.; Khanizadeh, S.; Cober, E.R.; Guillemette, R.J.D. Assessment of the protein quality of fourteen soybean [Glycine max (L.) Merr.] cultivars using amino acid analysis and two-dimensional electrophoresis. Food Res. Int. 2007, 40, 129–146. [Google Scholar] [CrossRef]
- Ali, N.M.; Yeap, S.-K.; Yusof, H.M.; Beh, B.-K.; Ho, W.-Y.; Koh, S.-P.; Abdullah, M.P.; Alitheen, N.B.; Long, K. Comparison of free amino acids, antioxidants, soluble phenolic acids, cytotoxicity and immunomodulation of fermented mung bean and soybean. J. Sci. Food Agric. 2016, 96, 1648–1658. [Google Scholar] [CrossRef]
- Dajanta, K.; Apichartsrangkoon, A.; Chukeatirote, E.; Frazier, R.A. Free-amino acid profiles of thua nao, a Thai fermented soybean. Food Chem. 2011, 125, 342–347. [Google Scholar] [CrossRef]
- Kim, E.H.; Kim, S.H.; Chung, J.I.; Chi, H.Y.; Kim, J.A.; Chung, I.M. Analysis of phenolic compounds and isoflavones in soybean seeds (Glycine max (L.) Merill) and sprouts grown under different conditions. Eur. Food Res. Technol. 2006, 222, 201–208. [Google Scholar] [CrossRef]
- Naczk, M.; Shahidi, F. Phenolics in cereals, fruits and vegetables: Occurrence, extraction and analysis. J. Pharm. Biomed. Anal. 2006, 41, 1523–1542. [Google Scholar] [CrossRef]
- Bunea, C.I.; Pop, N.; Babeş, A.C.; Matea, C.; Dulf, F.V.; Bunea, A. Carotenoids, total polyphenols and antioxidant activity of grapes (Vitis vinifera) cultivated in organic and conventional systems. Chem. Cent. J. 2012, 6, 66–74. [Google Scholar] [CrossRef] [Green Version]
- Boeing, J.S.; Barizão, E.O.; e Silva, B.C.; Montanher, P.F.; de Cinque Almeida, V.; Visentainer, J.V. Evaluation of solvent effect on the extraction of phenolic compounds and antioxidant capacities from the berries: Application of principal component analysis. Chem. Cent. J. 2014, 8, 48. [Google Scholar] [CrossRef]
- Zhao, H.; Dong, J.; Lu, J.; Chen, J.; Li, Y.; Shan, L.; Lin, Y.; Fan, W.; Gu, G. Effects of extraction solvent mixtures on antioxidant activity evaluation and their extraction capacity and selectivity for free phenolic compounds in barley (Hordeum vulgare L.). J. Agric. Food Chem. 2006, 54, 7277–7286. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Identification | Formula | tR (min) | Calc. Mass m/z [M] | Observed Mass m/z [M ± H]±1 | (±) Fragment Ions |
|---|---|---|---|---|---|---|
| 1 | Choline | C5H13NO | 0.93 | 103.0997 | 104.1069 | 60.0806 |
| 2 | Iditol | C6H14O6 | 0.94 | 182.0790 | 181.0717 | |
| 3 | Malic acid | C4H6O5 | 0.97 | 134.0215 | 133.0142 | |
| 4 | Citric acid | C6H8O7 | 0.98 | 192.0270 | 191.0197 | |
| 5 | Furoic acid | C5H4O3 | 0.99 | 112.0161 | 111.0088 | 67.0187 |
| 6 | Catechin | C15H14O6 | 1.00 | 290.0791 | 289.0718 | 245.0812, 203.0707, 187.0398, 137.0240, 109.0291 |
| 7 | Glutamic acid | C5H9NO4 | 1.04 | 147.0531 | 146.0458 | 102.0556, 84.0451 |
| 8 | Gluconic acid | C6H12O7 | 1.05 | 196.0583 | 195.0510 | 129.0189, 75.0085, 59.0137 |
| 9 | Aspartic acid | C4H7NO4 | 1.06 | 133.0375 | 132.0302 | 115.0032, 88.0401 |
| 10 | Histidine | C6H9N3O2 | 1.09 | 155.0695 | 156.0768 | 110.0703, 93.0439 |
| 11 | Malic acid | C4H6O5 | 1.20 | 134.0215 | 133.0142 | 115.0033, 71.0136 |
| 12 | Isoleucine | C6H13NO2 | 1.27 | 131.0946 | 132.1019 | 86.0957, 69.0693 |
| 13 | Cynaroside | C21H20O11 | 1.31 | 448.1006 | 449.1079 | 68.997 |
| 14 | UDP-N-acetylglucosamine * | C17H27N3O17P2 | 1.34 | 607.0818 | 606.0745 | 384.9831, 158.9254, 96.9692 |
| 15 | Tryptophan | C11H12N2O2 | 1.36 | 204.0898 | 205.0971 | 188.0692, 146.0589, 118.0642 |
| 16 | Phenylalanine | C9H11NO2 | 1.38 | 165.0789 | 166.0863 | 120.0803, 103.0538, 91.0539 |
| 17 | Catechin * | C15H14O6 | 1.47 | 290.0791 | 289.0718 | 187.0398, 137.0240, 109.0291 |
| 18 | Catechin analog * | C15H14O6 | 4.41 | 290.0789 | 289.0717 | 245.0812, 203.0707, 137.0240, 109.0291 |
| 19 | 3,5-Dihydroxy-2-(4-hydroxyphenyl)-4-oxo-3,4-dihydro-2H-chromen-7-yl hexopyranoside | C21H22O11 | 4.57 | 450.1162 | 449.1089 | 287.0554, 259.0604, 125.0240, 57.0345 |
| 20 | Apigetrin | C21H20O10 | 4.91 | 432.1056 | 431.0983 | 269.0446, 117.0339, 89.0240 |
| 21 | Daidzin | C21H20O9 | 5.82 | 416.1106 | 417.1179 | 227.0684, 199.0738, 137.0223 |
| 22 | Daidzein | C15H10O4 | 6.26 | 254.0577 | 255.0650 | 225.0552, 208.0527, 113.0295, 91.0187, 65.0033 |
| 23 | Apigetrin analog | C21H20O10 | 6.47 | 432.1054 | 477.1035 | 269.0446, 117.0339, 89.0240 |
| 24 | Glycitein | C16H12O5 | 6.64 | 284.0684 | 283.0612 | 268.0368, 240.0421 |
| 25 | Genistein | C15H10O5 | 7.15 | 270.0527 | 269.0454 | 225.0547, 181.0654 |
| 26 | Daidzein analog | C15H10O4 | 8.28 | 254.0579 | 253.0506 | 209.0598, 133.0292, 91.0186, 65.0031 |
| 27 | Naringenin | C15H12O5 | 9.26 | 272.0685 | 271.0612 | 151.0028, 119.0498, 93.0341, 65.0032 |
| 28 | Genistein analog | C15H10O5 | 9.53 | 270.0528 | 269.0455 | 225.0547, 181.0654 |
| 29 | Corchorifatty acid F | C18H32O5 | 9.61 | 328.2251 | 327.2178 | 229.1441, 211.13303, 171.1020, 85.0291, 57.0345 |
| 30 | (15Z)-9,12,13-Trihydroxy-15-octadecenoic acid * | C18H34O5 | 10.08 | 330.2407 | 329.2334 | 171.1020, 139.1123, 99.0812 |
| 31 | Soyasaponin I | C48H78O18 | 10.76 | 942.5182 | 941.5105 | 615.3881, 457.3671, 205.0709, 143.0345, 113.0241 |
| 32 | (±)9-HpODE * | C18H32O4 | 12.12 | 312.2301 | 311.2228 | 275.2005, 183.0115, 79.9569 |
| 33 | (+/−)9,10-dihydroxy-12Z-octadecenoic acid * | C18H34O4 | 13.00 | 314.2458 | 313.2385 | 277.2168, 201.1125, 171.1022 |
| 34 | 13(S)-HOTrE * | C18H30O3 | 13.95 | 294.2195 | 293.2123 | 195.1390, 95.9597, 79.9570 |
| No. | Identification | Formula | tR (min) | Calc. Mass m/z [M] | Observed m/z [M ± H]±1 | (±) Fragment Ions |
|---|---|---|---|---|---|---|
| 1 | Arginine | C6H14N4O2 | 0.83 | 174.1116 | 173.1044 | 156.0778, 131.0822, 114.0557 |
| 2 | Histidine | C6H9N3O2 | 0.85 | 155.0695 | 156.0768 | 110.0703, 93.0439 |
| 3 | Glutamic acid | C5H9NO4 | 0.93 | 147.0531 | 146.0458 | ND |
| 4 | Glucose | C6H12O6 | 0.93 | 180.0634 | 179.0561 | 101.024289.0242, 71.01369, 59.0138 |
| 5 | N-acetylornithine | C7H14N2O3 | 0.94 | 174.1004 | 173.0931 | 131.0822, 85.0769 |
| 6 | D-Ribose | C5H10O5 | 0.95 | 150.0528 | 149.0455 | 131.0346, 89.0243, 71.0137, 59.0138 |
| 7 | Malic acid | C4H6O5 | 0.97 | 134.0215 | 133.0142 | 115.0034, 89.0242, 72.9929, 71.0137 |
| 8 | Glutamine * | C5H10N2O3 | 0.99 | 146.0692 | 147.0765 | 127.0509, 109.0404, 84.0452 |
| 9 | Uridine | C9H12N2O6 | 1.00 | 244.0695 | 243.0623 | 200.0562, 152.0351, 110.0245 |
| 10 | Citric acid | C6H8O7 | 1.00 | 192.0269 | 191.0196 | 173.0090, 111.0085, 87.0087 |
| 11 | Arginine | C6H14N4O2 | 1.00 | 174.1117 | 175.1189 | 156.0778, 131.0822, 114.0557 |
| 12 | Furoic acid | C5H4O3 | 1.01 | 112.0161 | 111.0088 | 67.0189 |
| 13 | Pantothenic acid | C9H17NO5 | 1.01 | 219.1106 | 218.1033 | 146.0821, 88.0402 |
| 14 | Threonine | C4H9NO3 | 1.02 | 119.0582 | 120.0655 | 116.0696, 70.0645 |
| 15 | Asparagine | C4H8N2O3 | 1.03 | 132.0535 | 133.0608 | 116.0333, 87.0546, 74.0231 |
| 16 | Proline | C5H9NO2 | 1.03 | 115.0633 | 116.0706 | 70.0645 |
| 17 | Trans-Aconitic acid | C6H6O6 | 1.03 | 174.0164 | 173.0092 | 129.0191, 111.0085, 85.02936 |
| 18 | Cytosine | C4H5N3O | 1.04 | 111.0432 | 112.0505 | 95.0232, 69.0442 |
| 19 | Glutamic acid | C5H9NO4 | 1.04 | 147.0532 | 148.0605 | 128.0348, 102.0556, 84.0451 |
| 20 | Lysine | C6H14N2O2 | 1.04 | 146.1055 | 147.1128 | 130.0852, 84.0801, 56.0491 |
| 21 | Valine | C5H11NO2 | 1.08 | 117.0789 | 118.0862 | 100.0748, 72.0802, 55.0538 |
| 22 | Adenine | C5H5N5 | 1.11 | 135.0545 | 136.0617 | 119.0343 |
| 23 | Guanine * | C5H5N5O | 1.11 | 151.0494 | 152.0567 | 135.0294, 110.0341 |
| 24 | Succinic acid | C4H6O4 | 1.13 | 118.0266 | 117.0193 | 73.0293 |
| 25 | N-Acetylornithine | C7H14N2O3 | 1.21 | 174.1005 | 175.1077 | 131.0822, 85.0769 |
| 26 | Guanosine | C10H13N5O5 | 1.21 | 283.0917 | 282.0844 | 150.0419, 108.0202 |
| 27 | Hypoxanthine * | C5H4N4O | 1.21 | 136.0385 | 137.0458 | 119.0342, 110.0340, 94.0392 |
| 28 | Tyrosine * | C9H11NO3 | 1.22 | 181.0739 | 180.0666 | 163.0396, 119.0498, 72.0088 |
| 29 | Methionine | C5H11NO2S | 1.25 | 149.0511 | 150.0583 | 133.0307, 104.0520, 61.0102 |
| 30 | Isoleucine | C6H13NO2 | 1.30 | 131.0946 | 132.1018 | 86.0957, 69.0693 |
| 31 | Glutaric acid * | C5H8O4 | 1.32 | 132.0422 | 131.0349 | 113.0240, 87.0449, 69.0343 |
| 32 | Leucylproline | C11H20N2O3 | 1.40 | 228.1473 | 229.1546 | 116.0696, 86.0957, 70.0645 |
| 33 | Trans-3-Indoleacrylic acid | C11H9NO2 | 1.80 | 187.0632 | 188.0705 | 170.0586, 146.0588, 118.0641 |
| 34 | Tryptophan | C11H12N2O2 | 1.80 | 204.0898 | 205.0971 | 188.0692, 146.0589, 118.0642 |
| 35 | Glycidic acid | C10H10O3 | 2.17 | 178.0629 | 177.0557 | 133.0656, 71.01366 |
| 36 | 12-O-β-D-Glucopyranosyloxyjasmonic acid | C18H28O9 | 4.00 | 388.1732 | 387.1659 | 207.1023, 101.0242, 89.0242, 59.0138 |
| 37 | Sinensin | C21H22O11 | 4.26 | 450.1162 | 449.1089 | 287.0555, 259.0605, 125.02401 |
| 38 | Dihydrophaseic acid | C15H22O5 | 4.63 | 282.1468 | 281.1395 | 171.1175, 123.0813, 87.00853 |
| 39 | 12-O-β-D-Glucopyranosyloxyjasmonic acid | C18H28O9 | 4.95 | 388.1733 | 387.166 | 207.1023, 101.0242, 89.0242, 59.0138 |
| 40 | Daidzin * | C21H20O9 | 5.30 | 416.1108 | 417.1181 | 227.0684, 199.0738, 137.0223 |
| 41 | Glycitein | C16H12O5 | 5.33 | 284.0684 | 283.0611 | 268.0371, 240.0422 |
| 42 | Hdroxycaproic acid * | C6H12O3 | 5.39 | 132.0786 | 131.0714 | 113.0606, 85.0656, 57.0345 |
| 43 | N-Acetyl-l-phenylalanine | C11H13NO3 | 5.72 | 207.0895 | 206.0823 | 164.0713, 91.0551, 70.0296 |
| 44 | Pheyllactic acid * | C9H10O3 | 5.99 | 166.0629 | 165.0557 | 147.0449, 119.0501, 72.9929 |
| 45 | 2-(acetylamino)-3-(1H-indol-3-yl)propanoic acid | C13H14N2O3 | 6.30 | 246.1004 | 245.0931 | 116.0349, 98.0244, 74.0245, 58.0297 |
| 46 | Apigetrin * | C21H20O10 | 6.34 | 432.1056 | 431.0983 | 268.0368, 239.0335, 59.0137 |
| 47 | Genistin * | C21H20O10 | 6.47 | 432.1057 | 433.1129 | 271.0579, 215.0685, 153.0170 |
| 48 | Daidzein * | C15H10O4 | 6.48 | 254.0578 | 253.0505 | 208.0527, 113.0295, 91.0187, 65.0033 |
| 49 | Astragalin | C21H20O11 | 6.51 | 448.1005 | 447.0932 | 284.0320, 227.0345, 65.0032 |
| 50 | Glycitein analog | C16H12O5 | 6.56 | 284.0684 | 283.0611 | 268.0371, 240.0422 |
| 51 | Octyl glucoside | C14H28O6 | 6.72 | 292.1886 | 291.1813 | 85.0292, 59.0137 |
| 52 | 7-Hydroxy-2-(4-hydroxyphenyl)-4-oxo-3,4-dihydro-2H-chromen-5-yl β-d-glucopyranoside * | C21H22O10 | 7.04 | 434.1213 | 433.1139 | 271.0607, 243.0667, 151.0034, 93.0343 |
| 53 | Azelaic acid | C9H16O4 | 7.32 | 188.1048 | 187.0976 | 125.0969, 97.0656 |
| 54 | Galangin * | C15H10O5 | 7.49 | 270.0527 | 271.0599 | 215.0686, 153.0169, 115.0532 |
| 55 | Phloretin * | C15H14O5 | 7.77 | 274.0841 | 273.0768 | 167.0346, 123.0447, 93.0343 |
| 56 | Abscisic acid | C15H20O4 | 8.01 | 264.1362 | 263.1288 | 219.1386, 204.2252, 136.0526 |
| 57 | Daidzein analog | C15H10O4 | 8.18 | 254.0578 | 253.0505 | 225.0552, 132.0214, 91.0187, 65.0033 |
| 58 | 5,7-dihydroxy-3-(4-methoxyphenyl)-4H-chromen-4-one * | C16H12O5 | 8.56 | 284.0685 | 285.0757 | 229.0839, 197.0579, 118.0402 |
| 59 | Genistein | C15H10O5 | 9.43 | 270.0526 | 269.0454 | 225.0557, 181.0654 |
| 60 | (15Z)-9,12,13-trihydroxy-15-octadecenoic acid | C18H34O5 | 10.01 | 330.2405 | 329.2332 | 211.1335, 171.1023, 139.1125, 99.0813 |
| 61 | Tetradecanedioic acid | C14H26O4 | 10.63 | 258.1831 | 257.1758 | 239.1644 |
| 62 | Soyasaponin I | C48H78O18 | 10.77 | 942.5182 | 941.5107 | 733.4519, 457.3673, 257.0659, 101.0242 |
| 63 | (±)9-HpODE | C18H32O4 | 11.34 | 312.2301 | 311.2228 | 171.1023, 139.1125, 113.0968 |
| 64 | Thapsic acid | C16H30O4 | 11.75 | 286.2144 | 285.2071 | 267.2065, 59.0137 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tareq, F.S.; Kotha, R.R.; Natarajan, S.; Sun, J.; Luthria, D.L. An Untargeted Metabolomics Approach to Study the Variation between Wild and Cultivated Soybeans. Molecules 2023, 28, 5507. https://doi.org/10.3390/molecules28145507
Tareq FS, Kotha RR, Natarajan S, Sun J, Luthria DL. An Untargeted Metabolomics Approach to Study the Variation between Wild and Cultivated Soybeans. Molecules. 2023; 28(14):5507. https://doi.org/10.3390/molecules28145507
Chicago/Turabian StyleTareq, Fakir Shahidullah, Raghavendhar R. Kotha, Savithiry Natarajan, Jianghao Sun, and Devanand L. Luthria. 2023. "An Untargeted Metabolomics Approach to Study the Variation between Wild and Cultivated Soybeans" Molecules 28, no. 14: 5507. https://doi.org/10.3390/molecules28145507