
Germinated Rice Seeds Improved Resveratrol Production to Suppress Adipogenic and Inflammatory Molecules in 3T3-L1 Adipocytes

Abstract
:
1. Introduction
2. Results
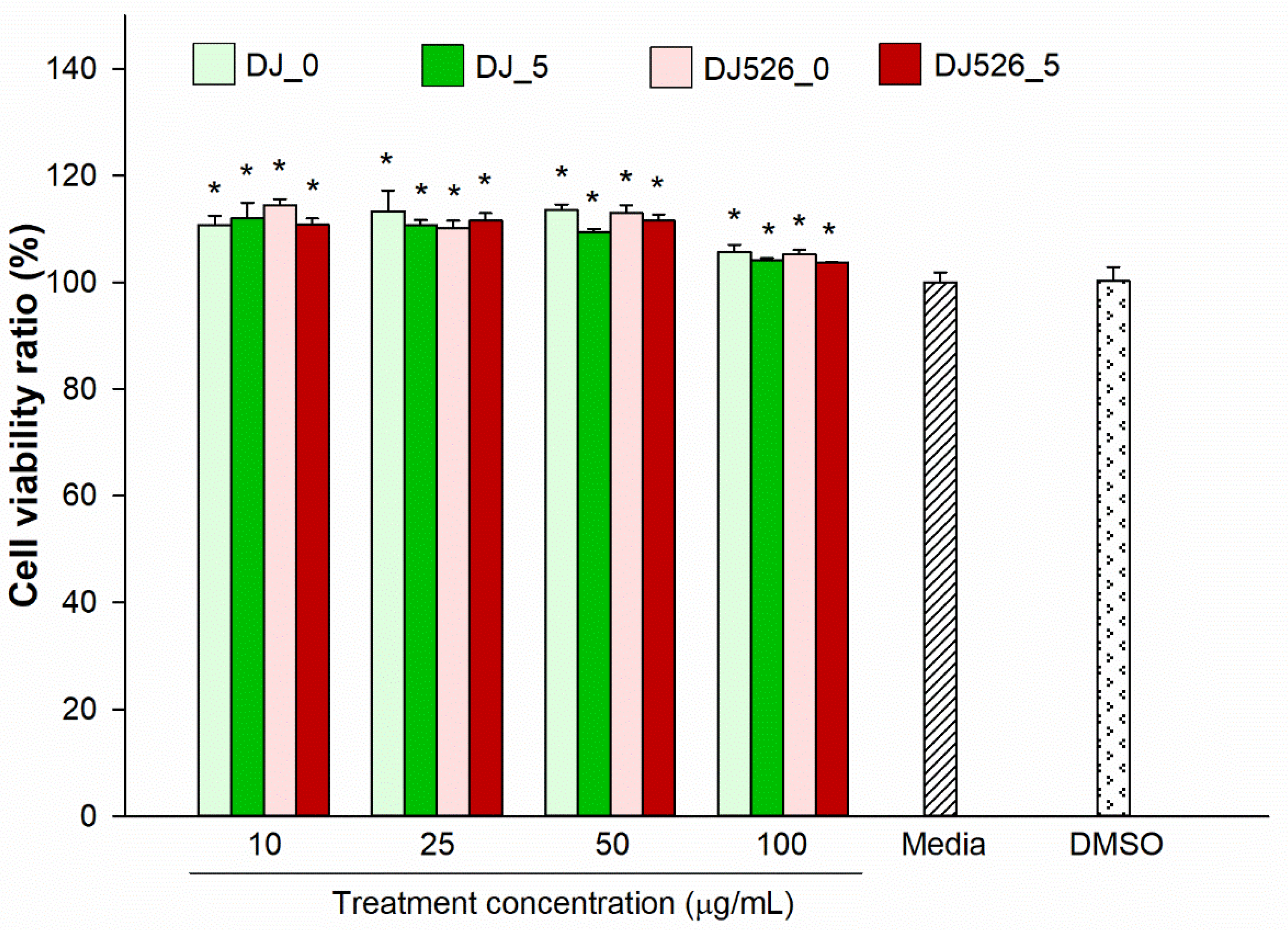
2.1. Effect of Germinated Rice Seed Extracts on Cell Viability
2.2. Effect of Germinated Rice Seed Extracts on Cell Differentiation
2.3. Effect of Germinated Rice Seed Extracts on Lipid Accumulation in Cells
2.4. Effect of Germinated Rice Seed Extracts on Adipogenic Gene Expression
2.5. Effect of Germinated Rice Seed Extracts on Expression of Adipogenic Proteins
2.6. Effect of Germinated Rice Seed Extracts on Viability of LPS-Stimulated Adipocytes
2.7. Effect of Germinated Rice Seed Extracts on Expression of Inflammatory Genes
2.8. Effect of Germinated Rice Seed Extracts on Expression of Inflammatory Proteins
3. Discussion
4. Materials and Methods
4.1. Treatments
4.2. 3T3-L1 Cell Culture
4.3. 3T3-L1 Cell Viability Assay
4.4. 3T3-L1 Cell Differentiation
4.5. Oil Red O Staining Assay
4.6. RNA Extraction and cDNA Synthesis
4.7. Real-Time PCR Analysis
4.8. Western Blot Analysis
4.9. Anti-Inflammatory Adipocyte Assays
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Kim, H.L.; Lee, S.K.; Min, D.E.; Choi, B.K.; Lee, D.R. Anti-obesity effects of a mixture of Atractylodes macrocephala and Amomum villosum extracts on 3T3-L1 adipocytes and high-fat diet-induced obesity in mice. Molecules 2022, 27, 906–920. [Google Scholar] [CrossRef] [PubMed]
- Jakab, J.; Miškić, B.; Mikšić, Š.; Juranić, B.; Ćosić, V.; Schwarz, D.; Včev, A. Adipogenesis as a potential anti-obesity target: A review of pharmacological treatment and natural products. Diabetes Metab. Syndr. Obes. Targets Ther. 2021, 14, 67–83. [Google Scholar] [CrossRef] [PubMed]
- Xia, M.; Liu, K.; Feng, J.; Zheng, Z.; Xie, X. Prevalence and risk factors of type 2 diabetes and prediabetes among middle-aged and elderly adults in China: A cross-sectional study. Diabetes Metab. Syndr. Obes. Targets Ther. 2021, 14, 1975–1985. [Google Scholar] [CrossRef] [PubMed]
- Li, N.; Chen, K.; Dong, H.; Yang, J.; Yoshizawa, M.; Kagami, H.; Li, X. Alliin inhibits adipocyte differentiation by downregulating Akt expression: Implications for metabolic disease. Exp. Ther. Med. 2021, 21, 563. [Google Scholar] [CrossRef]
- Shirasawa, T.; Ochiai, H.; Yoshimoto, T.; Nagahama, S.; Kobayashi, M.; Ohtsu, I.; Sunaga, Y.; Kokaze, A. Associations between normal weight central obesity and cardiovascular disease risk factors in Japanese middle-aged adults: A cross-sectional study. J. Health Popul. Nutr. 2019, 38, 46. [Google Scholar] [CrossRef] [Green Version]
- Xanthopoulos, M.; Tapia, I.E. Obesity and common respiratory diseases in children. Paediatr. Respir. Rev. 2017, 23, 68–71. [Google Scholar] [CrossRef] [PubMed]
- Parikesit, D.; Mochtar, C.A.; Umbas, R.; Hamid, A.R.A.H. The impact of obesity towards prostate diseases. Prostate Int. 2016, 4, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Moseti, D.; Regassa, A.; Kim, W.-K. Molecular regulation of adipogenesis and potential anti-adipogenic bioactive molecules. Int. J. Mol. Sci. 2016, 17, 124. [Google Scholar] [CrossRef] [Green Version]
- Sarkar, P.; Thirumurugan, K. Modulatory functions of bioactive fruits, vegetables and spices in adipogenesis and angiogenesis. J. Funct. Foods 2019, 53, 318–336. [Google Scholar] [CrossRef]
- Lee, H.; Lee, J.-H.; Kim, D.; Hwang, D.; Lee, M.; Chung, H.; Kim, T.-J.; Kim, H.S. Micro-current stimulation can modulate the adipogenesis process by regulating the insulin signaling pathway in 3T3-L1 cells and ob/ob mice. Life 2023, 13, 404. [Google Scholar] [CrossRef]
- Oates, E.H.; Antoniewicz, M.R. Coordinated reprogramming of metabolism and cell function in adipocytes from proliferation to differentiation. Metab. Eng. 2022, 69, 221–230. [Google Scholar] [CrossRef]
- Han, J.-H.; Jang, K.-W.; Park, M.-H.; Myung, C.-S. Garcinia cambogia suppresses adipogenesis in 3T3-L1 cells by inhibiting p90RSK and Stat3 activation during mitotic clonal expansion. J. Cell. Physiol. 2021, 236, 1822–1839. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Cho, J.S.; Kim, J.T.; Moon, J.H.; Zhou, Y.; Lee, S.B.; Park, H.J.; Lee, H.J. Caudatin suppresses adipogenesis in 3T3-L1 adipocytes and reduces body weight gain in high-fat diet-fed mice through activation of hedgehog signaling. Phytomedicine 2021, 92, 153715. [Google Scholar] [CrossRef] [PubMed]
- Czech, M.P. Mechanisms of insulin resistance related to white, beige, and brown adipocytes. Mol. Metab. 2020, 34, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Kim, J.; Lee, H.D.; Cho, H.; Paje, L.A.; Shin, H.; Lee, S. Development of an analytical approach for the utilization of edible tree sprouts. Nat. Prod. Sci. 2022, 28, 27–32. [Google Scholar] [CrossRef]
- Ha, J.-H.; Jang, J.; Chung, S.-I.; Yoon, Y. AMPK and SREBP-1c mediate the anti-adipogenic effect of β-hydroxyisovalerylshikonin. Int. J. Mol. Med. 2016, 37, 816–824. [Google Scholar] [CrossRef] [Green Version]
- Hadrich, F.; Sayadi, S. Apigetrin inhibits adipogenesis in 3T3-L1 cells by downregulating PPARγ and CEBP-α. Lipids Health Dis. 2018, 17, 95. [Google Scholar] [CrossRef] [Green Version]
- Ceja-Galicia, Z.; Calderón-DuPont, D.; Daniel, A.; Chiu, L.M.; Díaz-Villaseñor, A. Leptin and adiponectin synthesis and secretion in mature 3T3-L1 adipocytes are differentially down-regulated by arsenic and palmitic acid exposure throughout different stages of adipogenesis. Life Sci. 2022, 291, 120262. [Google Scholar] [CrossRef]
- González-Arceo, M.; Gomez-Lopez, I.; Carr-Ugarte, H.; Eseberri, I.; González, M.; Cano, M.P.; Portillo, M.P.; Gómez-Zorita, S. Anti-Obesity Effects of Isorhamnetin and Isorhamnetin Conjugates. Int. J. Mol. Sci. 2023, 24, 299. [Google Scholar] [CrossRef]
- Kim, N.-Y.; Lim, C.-M.; Park, H.-M.; Kim, J.; Pham, T.-H.; Yang, Y.; Lee, H.P.; Hong, J.T.; Yoon, D.-Y. MMPP promotes adipogenesis and glucose uptake via binding to the PPARγ ligand binding domain in 3T3-L1 MBX cells. Front. Pharmacol. 2022, 13, 994584. [Google Scholar] [CrossRef]
- D’Esposito, V.; Ambrosio, M.R.; Liguoro, D.; Perruolo, G.; Lecce, M.; Cabaro, S.; Aprile, M.; Marino, A.; Pilone, V.; Forestieri, P.; et al. In severe obesity, subcutaneous adipose tissue cell-derived cytokines are early markers of impaired glucose tolerance and are modulated by quercetin. Int. J. Obes. 2021, 45, 1811–1820. [Google Scholar] [CrossRef]
- Luo, Y.; Lin, H. Inflammation initiates a vicious cycle between obesity and nonalcoholic fatty liver disease. Immun. Inflamm. Dis. 2021, 9, 59–73. [Google Scholar] [CrossRef] [PubMed]
- Boleti AP, D.A.; Cardoso PH, D.O.; Frihling BE, F.; e Silva, P.S.; de Moraes LF, R.; Migliolo, L. Adipose tissue, systematic inflammation, and neurodegenerative diseases. Neural Regen. Res. 2023, 18, 38–46. [Google Scholar] [CrossRef]
- Zatterale, F.; Longo, M.; Naderi, J.; Raciti, G.A.; Desiderio, A.; Miele, C.; Beguinot, F. Chronic adipose tissue inflammation linking obesity to insulin resistance and type 2 diabetes. Front. Physiol. 2019, 10, 1607. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Song, W.; Zushin, P.H.; Liu, B.; Jedrychowski, M.P.; Mina, A.I.; Deng, Z.; Cabarkapa, D.; Hall, J.A.; Palmer, C.J.; et al. Phosphorylation of Beta-3 adrenergic receptor at serine 247 by ERK MAP kinase drives lipolysis in obese adipocytes. Mol. Metab. 2018, 12, 25–38. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, H.; Takahashi-Muto, C.; Nagase, M.; Kassai, M.; Tanaka-Yachi, R.; Kiyose, C. Anti-inflammatory effects of extracts of Sweet Basil (Ocimum basilicum L.) on a co-culture of 3T3-L1 adipocytes and RAW264.7 macrophages. J. Oleo Sci. 2020, 69, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Harlan, L.; Mena, L.T.; Ramalingam, L.; Jayarathne, S.; Shen, C.-L.; Moustaid-Moussa, N. Mechanisms mediating anti-inflammatory effects of delta-tocotrienol and tart cherry anthocyanins in 3T3-L1 adipocytes. Nutrients 2020, 12, 3356. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-X.; Li, C.-X.; Kakar, M.U.; Khan, M.S.; Wu, P.-F.; Amir, R.M.; Dai, D.-F.; Naveed, M.; Li, Q.-Y.; Saeed, M.; et al. Resveratrol (RV): A pharmacological review and call for further research. Biomed. Pharmacother. 2021, 143, 112164. [Google Scholar] [CrossRef]
- Zhou, D.-D.; Luo, M.; Huang, S.-Y.; Saimaiti, A.; Shang, A.; Gan, R.-Y.; Li, H.-B. Effects and mechanisms of resveratrol on aging and age-related diseases. Oxid. Med. Cell. Longev. 2021, 2021, 9932218. [Google Scholar] [CrossRef]
- Gong, C.; Xia, H. Resveratrol suppresses melanoma growth by promoting autophagy through inhibiting the PI3K/AKT/mTOR signaling pathway. Exp. Ther. Med. 2020, 19, 1878–1886. [Google Scholar] [CrossRef]
- Toomer, O.T.; Vu, T.; Pereira, M.; Williams, K. Dietary supplementation with peanut skin polyphenolic extracts (PSPE) reduces hepatic lipid and glycogen stores in mice fed an atherogenic diet. J. Funct. Foods 2019, 55, 362–370. [Google Scholar] [CrossRef]
- Wang, N.; Luo, Z.; Jin, M.; Sheng, W.; Wang, H.T.; Long, X.; Wu, Y.; Hu, P.; Xu, H.; Zhang, X. Exploration of age-related mitochondrial dysfunction and the anti-aging effects of resveratrol in zebrafish retina. Aging 2019, 11, 3117–3137. [Google Scholar] [CrossRef] [PubMed]
- Meng, T.; Xiao, D.; Muhammed, A.; Deng, J.; Chen, L.; He, J. Anti-inflammatory action and mechanisms of resveratrol. Molecules 2021, 26, 229. [Google Scholar] [CrossRef]
- Chen, X.; Song, X.; Zhao, X.; Zhang, Y.; Wang, Y.; Jia, R.; Zou, Y.; Li, L.; Yin, Z. Insights into the anti-inflammatory and antiviral mechanisms of resveratrol. Mediat. Inflamm. 2022, 2022, 7138756. [Google Scholar] [CrossRef] [PubMed]
- Dyck, G.J.B.; Raj, P.; Zieroth, S.; Dyck, J.R.B.; Ezekowitz, J.A. The effects of resveratrol in patients with cardiovascular disease and heart failure: A narrative review. Int. J. Mol. Sci. 2019, 20, 904. [Google Scholar] [CrossRef] [Green Version]
- Berretta, M.; Bignucolo, A.; Di Francia, R.; Comello, F.; Facchini, G.; Ceccarelli, M.; Iaffaioli, R.V.; Quagliariello, V.; Maurea, N. Resveratrol in cancer patients: From bench to bedside. Int. J. Mol. Sci. 2020, 21, 2945. [Google Scholar] [CrossRef] [Green Version]
- Fu, X.; Li, M.; Tang, C.; Huang, Z.; Najafi, M. Targeting of cancer cell death mechanisms by resveratrol: A review. Apoptosis 2021, 26, 561–573. [Google Scholar] [CrossRef]
- Ren, B.; Kwah, M.X.-Y.; Liu, C.; Ma, Z.; Shanmugam, M.K.; Ding, L.; Xiang, X.; Ho, P.C.-L.; Wang, L.; Ong, P.S.; et al. Resveratrol for cancer therapy: Challenges and future perspectives. Cancer Lett. 2021, 515, 63–72. [Google Scholar] [CrossRef]
- Baek, S.-H.; Shin, W.-C.; Ryu, H.-S.; Lee, D.-W.; Moon, E.; Seo, C.-S.; Hwang, E.; Lee, H.-S.; Ahn, M.-H.; Jeon, Y.; et al. Creation of resveratrol-enriched rice for the treatment of metabolic syndrome and related diseases. PLoS ONE 2013, 8, e57930. [Google Scholar] [CrossRef]
- Monmai, C.; Kim, J.-S.; Baek, S.-H. Use of germination to enhance resveratrol content and its anti-inflammatory activity in lipopolysaccharide-stimulated RAW264.7 cells. Molecules 2023, 28, 4898. [Google Scholar] [CrossRef]
- Choi, S.-I.; Kwon, H.-Y.; Han, X.; Men, X.; Choi, Y.-E.; Jang, G.-W.; Park, K.-T.; Han, J.; Lee, O.-H. Environmental obesogens (bisphenols, phthalates and parabens) and their impacts on adipogenic transcription factors in the absence of dexamethasone in 3T3-L1 cells. J. Steroid Biochem. Mol. Biol. 2021, 214, 105994. [Google Scholar] [CrossRef]
- Park, T.J.; Park, A.; Kim, J.; Kim, J.Y.; Han, B.S.; Oh, K.J.; Lee, E.W.; Lee, S.C.; Bae, K.H.; Kim, W.K. Myonectin inhibits adipogenesis in 3T3-L1 preadipocytes by regulating p38 MAPK pathway. BMB Rep. 2021, 54, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Jemai, R.; Drira, R.; Makni, M.; Fetoui, H.; Sakamoto, K. Colocynth (Citrullus colocynthis) seed extracts attenuate adipogenesis by down-regulating PPARγ/ SREBP-1c and C/EBPα in 3T3-L1 cells. Food Biosci. 2020, 33, 100491. [Google Scholar] [CrossRef]
- Kurek, J.M.; Zielińska-Wasielica, J.; Kowalska, K.; Krejpcio, Z.; Olejnik, A. Modulating effects of steviol and steviol glycosides on adipogenesis, lipogenesis, glucose uptake and insulin resistance in 3T3-L1 adipocyte model. J. Funct. Foods 2022, 94, 105141. [Google Scholar] [CrossRef]
- Rayalam, S.; Yang, J.-Y.; Ambati, S.; Della-Fera, M.A.; Baile, C.A. Resveratrol induces apoptosis and inhibits adipogenesis in 3T3-L1 adipocytes. Phytother. Res. 2008, 22, 1367–1371. [Google Scholar] [CrossRef]
- Park, I.S.; Han, Y.; Jo, H.; Lee, K.W.; Song, Y.S. Piceatannol is superior to resveratrol at suppressing adipogenesis in human visceral adipose-derived stem cells. Plants 2021, 10, 366. [Google Scholar] [CrossRef]
- Chen, S.; Li, Z.; Li, W.; Shan, Z.; Zhu, W. Resveratrol inhibits cell differentiation in 3T3-L1 adipocytes via activation of AMPK. Can. J. Physiol. Pharmacol. 2011, 89, 793–799. [Google Scholar]
- Bost, F.; Aouadi, M.; Caron, L.; Binétruy, B. The role of MAPKs in adipocyte differentiation and obesity. Biochimie 2005, 87, 51–56. [Google Scholar] [CrossRef] [Green Version]
- Sale, E.M.; Atkinson, P.G.; Sale, G.J. Requirement of MAP kinase for differentiation of fibroblasts to adipocytes, for insulin activation of p90 S6 kinase and for insulin or serum stimulation of DNA synthesis. EMBO J. 1995, 14, 674–684. [Google Scholar] [CrossRef]
- Prusty, D.; Park, B.H.; Davis, K.E.; Farmer, S.R. Activation of MEK/ERK signaling promotes adipogenesis by enhancing peroxisome proliferator-activated receptor gamma (PPARgamma) and C/EBPalpha gene expression during the differentiation of 3T3-L1 preadipocytes. J. Biol. Chem. 2002, 277, 46226–46232. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.H.; Karadeniz, F.; Lee, J.I.; Seo, Y.; Kong, C.S. Artemisia princeps inhibits adipogenic differentiation of 3T3-L1 pre-adipocytes via downregulation of PPARγ and MAPK pathways. Prev. Nutr. Food Sci. 2019, 24, 299–307. [Google Scholar] [CrossRef]
- Tseng, Y.-H.; Butte, A.J.; Kokkotou, E.; Yechoor, V.K.; Taniguchi, C.M.; Kriauciunas, K.M.; Cypess, A.M.; Niinobe, M.; Yoshikawa, K.; Patti, M.E.; et al. Prediction of preadipocyte differentiation by gene expression reveals role of insulin receptor substrates and necdin. Nat. Cell Biol. 2005, 7, 601–611. [Google Scholar] [CrossRef] [PubMed]
- Chung, Y.C.; Hyun, C.-G. Inhibitory effects of pinostilbene on adipogenesis in 3T3-L1 adipocytes: A study of possible mechanisms. Int. J. Mol. Sci. 2021, 22, 13446. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.L.; Ha, A.W.; Kim, W.K. Effect of saccharin on inflammation in 3T3-L1 adipocytes and the related mechanism. Nutr. Res. Pract. 2020, 14, 109–116. [Google Scholar] [CrossRef]
- Ding, S.; Jiang, J.; Wang, Z.; Zhang, G.; Yin, J.; Wang, X.; Wang, S.; Yu, Z. Resveratrol reduces the inflammatory response in adipose tissue and improves adipose insulin signaling in high-fat diet-fed mice. PeerJ 2018, 6, e5173. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Monmai, C.; Kim, J.-S.; Baek, S.-H. Transgenic rice seed extracts exert immunomodulatory effects by modulating immune-related biomarkers in RAW264.7 macrophage cells. Nutrients 2022, 14, 4143. [Google Scholar] [CrossRef]
- Yamamoto, K.; Yamashita, M.; Inagawa, H.; Kohchi, C.; Soma, G.I. Anti-inflammatory and Insulin Signaling Phenotype Induced by Repeated Lipopolysaccharide Stimulation in 3T3-L1 Adipocytes. Anticancer Res. 2022, 42, 3983–3991. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Nucleotide Sequences | Accession No. |
|---|---|---|
| Adiponectin | FP: 5′–AAA GGA GAG CCT GGA GAA GC–3′ RP: 5′–GTA GAG TCC CGG AAT GTT GC–3′ | NM_009605.5 |
| C/EBPα | FP: 5′–TTA CAA CAG GCC AGG TTT CC–3′ RP: 5′–AAC TCC AGT CCC TCT GGG AT–3′ | NM_001287514.1 |
| FAS | FP: 5′–CTC TGA TCA GTG GCC TCC TC–3′ RP: 5′–TGC TGC AGT TTG GTC TGA AC–3′ | AF127033.1 |
| PPARγ | FP: 5′–CCC TGG CAA AGC ATT TGT AT–3′ RP: 5′–GAA ACT GGC ACC CTT GAA AA–3′ | AB644275.1 |
| SREBP-1 | FP: 5′–AGC TCA AAG ACC TGG TGG TG–3′ RP: 5′–TCA TGC CCT CCA TAG ACA CA–3′ | BC056922.1 |
| β-actin | FP: 5′–CCA CAG CTG AGA GGG AAA TC–3′ RP: 5′–AAG GAA GGC TGG AAA AGA GC–3′ | NM_007393.5 |
| Target Gene | Sequence (5′–3′) | Accession No. |
|---|---|---|
| COX-2 | FP: 5′–AGA AGG AAA TGG CTG CAG AA–3′ RP: 5′–GCT CGG CTT CCA GTA TTGAG–3′ | NM_011198.4 |
| TNF-α | FP: 5′–ATG AGC ACA GAA AGC ATG ATC–3′ RP: 5′–TAC AGGCTT GTC ACT CGA ATT–3′ | D84199.2 |
| IL-1β | FP: 5′–GGG CCT CAA AGG AAA GAA TC–3′ RP: 5′–TAC CAG TTG GGGAAC TCT GC–3′ | NM_008361.4 |
| IL-6 | FP: 5′–AGT TGC CTT CTT GGG ACT GA–3′ RP: 5′–CAG AAT TGC CAT TGC ACA AC–3′ | NM_031168.2 |
| β-actin | FP: 5′–CCA CAG CTG AGA GGG AAA TC–3′ RP: 5′–AAG GAA GGC TGG AAA AGA GC–3′ | NM_007393.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monmai, C.; Kim, J.-S.; Baek, S.-H. Germinated Rice Seeds Improved Resveratrol Production to Suppress Adipogenic and Inflammatory Molecules in 3T3-L1 Adipocytes. Molecules 2023, 28, 5750. https://doi.org/10.3390/molecules28155750
Monmai C, Kim J-S, Baek S-H. Germinated Rice Seeds Improved Resveratrol Production to Suppress Adipogenic and Inflammatory Molecules in 3T3-L1 Adipocytes. Molecules. 2023; 28(15):5750. https://doi.org/10.3390/molecules28155750
Chicago/Turabian StyleMonmai, Chaiwat, Jin-Suk Kim, and So-Hyeon Baek. 2023. "Germinated Rice Seeds Improved Resveratrol Production to Suppress Adipogenic and Inflammatory Molecules in 3T3-L1 Adipocytes" Molecules 28, no. 15: 5750. https://doi.org/10.3390/molecules28155750