

Zanubrutinib Ameliorates Cardiac Fibrosis and Inflammation Induced by Chronic Sympathetic Activation
 , ,
, ,
Abstract
:1. Introduction
2. Results
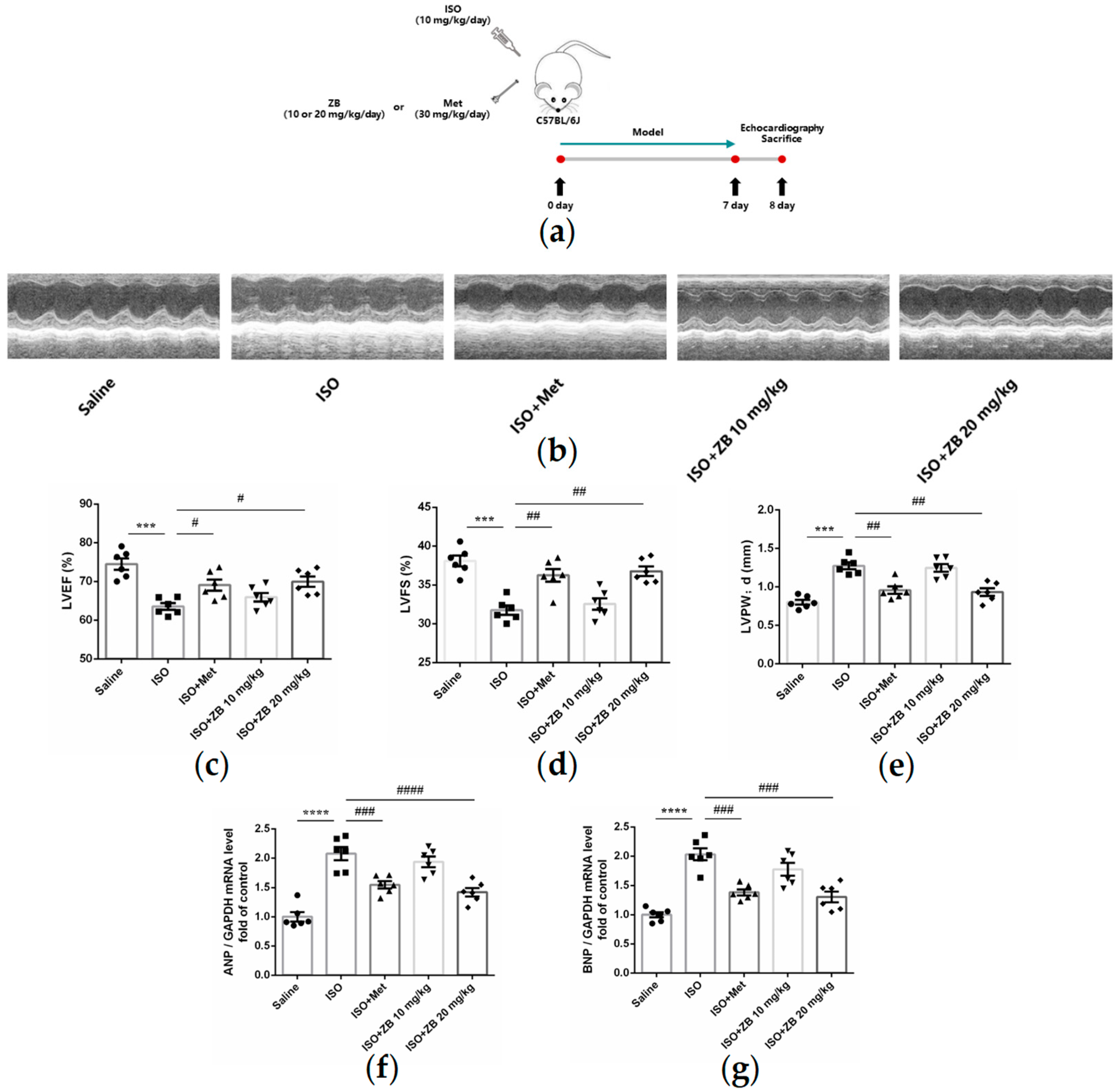
2.1. ZB Alleviated ISO-Induced Cardiac Dysfunction In Vivo
2.2. ZB Alleviated ISO-Induced Cardiac Fibrosis In Vivo
2.3. ZB Reduced ISO-Induced Cardiac Fibroblasts Activation In Vitro
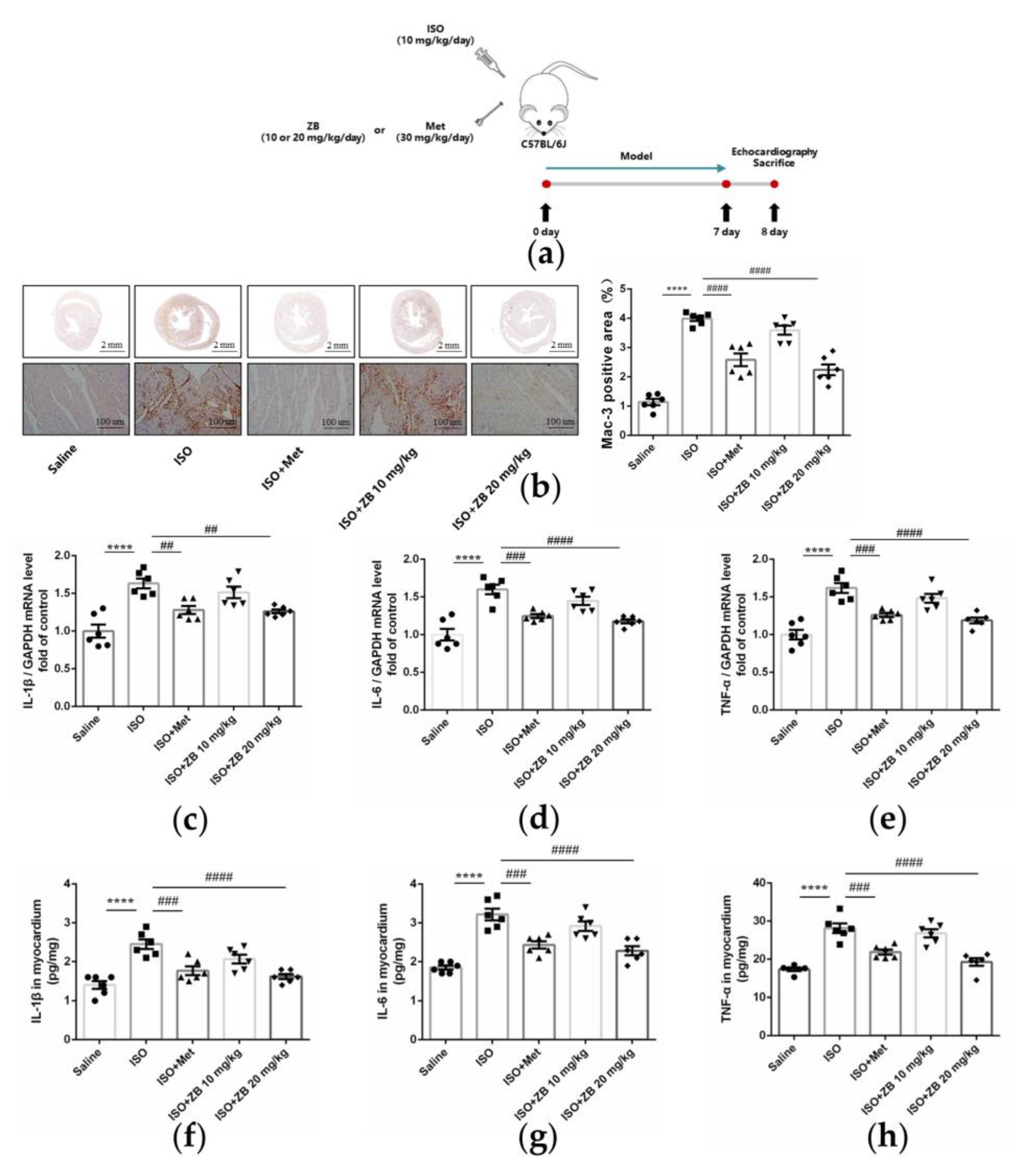
2.4. ZB Alleviated ISO-Induced Cardiac Inflammation In Vivo
2.5. ZB Reduced ISO-Induced Macrophage Pro-Inflammatory Production In Vitro
2.6. ZB Reduced ISO-Induced Cardiac Fibroblasts Activated by STAT3 and PI3K/Akt Signaling Pathways
2.7. ZB Reduced ISO-Induced Macrophage Pro-Inflammatory Production by PI3K/Akt and NF-κB Signaling Pathways
2.8. ZB Reduced ISO-Induced Cardiac Fibrosis and Inflammation by STAT3, NF-κB, and PI3K/Akt Signaling Pathways In Vivo
2.9. Working Model of the Effects of ZB on β-AR Stimulation-Induced Cardiac Fibrosis and Inflammation
3. Discussion
4. Materials and Methods
4.1. Animal Model and Treatments
4.2. Echocardiographic Measurements
4.3. Isolation of Cardiac Fibroblasts
4.4. Histological Analysis
4.5. Total RNA Extraction and Real-Time PCR Analysis
4.6. Cell Culture and Treatments
4.7. Western Blot Analysis
4.8. Immunofluorescence
4.9. Immunohistochemistry
4.10. Enzyme-Linked Immunosorbent Assay (ELISA)
4.11. Cell Counting Kit-8 Assay
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
References
- Travers, J.G.; Kamal, F.A.; Robbins, J.; Yutzey, K.E.; Blaxall, B.C. Cardiac Fibrosis: The Fibroblast Awakens. Circ. Res. 2016, 118, 1021–1040. [Google Scholar] [CrossRef] [PubMed]
- Schimmel, K.; Jung, M.; Foinquinos, A.; Jose, G.S.; Beaumont, J.; Bock, K.; Grote-Levi, L.; Xiao, K.; Bar, C.; Pfanne, A.; et al. Natural Compound Library Screening Identifies New Molecules for the Treatment of Cardiac Fibrosis and Diastolic Dysfunction. Circulation 2020, 141, 751–767. [Google Scholar] [CrossRef]
- Carter, J.R.; Goldstein, D.S. Sympathoneural and adrenomedullary responses to mental stress. Compr. Physiol. 2015, 5, 119–146. [Google Scholar] [CrossRef]
- Ruparelia, N.; Chai, J.T.; Fisher, E.A.; Choudhury, R.P. Inflammatory processes in cardiovascular disease: A route to targeted therapies. Nat. Rev. Cardiol. 2017, 14, 133–144. [Google Scholar] [CrossRef]
- Fu, Y.; Xiao, H.; Zhang, Y. Beta-adrenoceptor signaling pathways mediate cardiac pathological remodeling. Front. Biosci. 2012, 4, 1625–1637. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Wu, J.M.; Hu, G.M.; Li, M.Z.; Cong, W.W.; Feng, Y.N.; Wang, S.X.; Li, Z.J.; Xu, M.; Dong, E.D.; et al. Membrane nanotubes facilitate the propagation of inflammatory injury in the heart upon overactivation of the beta-adrenergic receptor. Cell Death Dis. 2020, 11, 958. [Google Scholar] [CrossRef]
- Hauser, A.S.; Attwood, M.M.; Rask-Andersen, M.; Schioth, H.B.; Gloriam, D.E. Trends in GPCR drug discovery: New agents, targets and indications. Nat. Rev. Drug Discov. 2017, 16, 829–842. [Google Scholar] [CrossRef]
- Xiao, H.; Li, H.; Wang, J.J.; Zhang, J.S.; Shen, J.; An, X.B.; Zhang, C.C.; Wu, J.M.; Song, Y.; Wang, X.Y.; et al. IL-18 cleavage triggers cardiac inflammation and fibrosis upon beta-adrenergic insult. Eur. Heart J. 2018, 39, 60–69. [Google Scholar] [CrossRef] [Green Version]
- Tang, M.; Li, Z.; Zhang, C.; Lu, X.; Tu, B.; Cao, Z.; Li, Y.; Chen, Y.; Jiang, L.; Wang, H.; et al. SIRT7-mediated ATM deacetylation is essential for its deactivation and DNA damage repair. Sci. Adv. 2019, 5, eaav1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Berlo, J.H.; Maillet, M.; Molkentin, J.D. Signaling effectors underlying pathologic growth and remodeling of the heart. J. Clin. Investig. 2013, 123, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Chaulet, H.; Lin, F.; Guo, J.; Owens, W.A.; Michalicek, J.; Kesteven, S.H.; Guan, Z.; Prall, O.W.; Mearns, B.M.; Feneley, M.P.; et al. Sustained augmentation of cardiac alpha1A-adrenergic drive results in pathological remodeling with contractile dysfunction, progressive fibrosis and reactivation of matricellular protein genes. J. Mol. Cell. Cardiol. 2006, 40, 540–552. [Google Scholar] [CrossRef] [PubMed]
- Neys, S.F.H.; Rip, J.; Hendriks, R.W.; Corneth, O.B.J. Bruton’s Tyrosine Kinase Inhibition as an Emerging Therapy in Systemic Autoimmune Disease. Drugs 2021, 81, 1605–1626. [Google Scholar] [CrossRef]
- Dobrovolsky, D.; Wang, E.S.; Morrow, S.; Leahy, C.; Faust, T.; Nowak, R.P.; Donovan, K.A.; Yang, G.; Li, Z.; Fischer, E.S.; et al. Bruton tyrosine kinase degradation as a therapeutic strategy for cancer. Blood 2019, 133, 952–961. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Nierman, P.; Kendall, E.K.; Cheung, J.; Gulrajani, M.; Herman, S.E.M.; Pleyer, C.; Ahn, I.E.; Stetler-Stevenson, M.; Yuan, C.M.; et al. Clinical and biological implications of target occupancy in CLL treated with the BTK inhibitor acalabrutinib. Blood 2020, 136, 93–105. [Google Scholar] [CrossRef]
- Roschewski, M.; Lionakis, M.S.; Sharman, J.P.; Roswarski, J.; Goy, A.; Monticelli, M.A.; Roshon, M.; Wrzesinski, S.H.; Desai, J.V.; Zarakas, M.A.; et al. Inhibition of Bruton tyrosine kinase in patients with severe COVID-19. Sci. Immunol. 2020, 5, eabd0110. [Google Scholar] [CrossRef]
- Holopainen, T.; Rasanen, M.; Anisimov, A.; Tuomainen, T.; Zheng, W.; Tvorogov, D.; Hulmi, J.J.; Andersson, L.C.; Cenni, B.; Tavi, P.; et al. Endothelial Bmx tyrosine kinase activity is essential for myocardial hypertrophy and remodeling. Proc. Natl. Acad. Sci. USA 2015, 112, 13063–13068. [Google Scholar] [CrossRef]
- Mitchell-Jordan, S.A.; Holopainen, T.; Ren, S.; Wang, S.; Warburton, S.; Zhang, M.J.; Alitalo, K.; Wang, Y.; Vondriska, T.M. Loss of Bmx nonreceptor tyrosine kinase prevents pressure overload-induced cardiac hypertrophy. Circ. Res. 2008, 103, 1359–1362. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Wang, Z.; Li, H.; Hu, Z.; Hong, H.; Sun, Y.; Ke, Y.; Du, X. ITK inhibition promotes long-term survival of cardiac allografts by regulating T cell PLCg phosphorylation. Am. J. Transl. Res. 2020, 12, 5762–5771. [Google Scholar]
- Wang, B.; Tan, Y.; Zhou, W.; Yang, J.; Jiang, Y.; Liu, X.; Zhan, Z. Loss of BTK ameliorates the pathological cardiac fibrosis and dysfunction. Matrix Biol. 2022, 112, 171–189. [Google Scholar] [CrossRef]
- Song, Y.; Zhou, K.; Zou, D.; Zhou, J.; Hu, J.; Yang, H.; Zhang, H.; Ji, J.; Xu, W.; Jin, J.; et al. Treatment of Patients with Relapsed or Refractory Mantle-Cell Lymphoma with Zanubrutinib, a Selective Inhibitor of Bruton’s Tyrosine Kinase. Clin. Cancer Res. 2020, 26, 4216–4224. [Google Scholar] [CrossRef]
- Lim, K.J.C.; Tam, C.S. Zanubrutinib for the treatment of Waldenstrom Macroglobulinemia. Expert Rev. Hematol. 2020, 13, 1303–1310. [Google Scholar] [CrossRef]
- Feng, Y.; Zhang, Y.; Xiao, H. AMPK and cardiac remodelling. Sci. China Life Sci. 2018, 61, 14–23. [Google Scholar] [CrossRef]
- Wang, W.; Li, W.; Liu, K.; Niu, X.; Guan, K.; Jiang, Y.; Li, Z.; Dong, E. Src mediates beta-adrenergic receptor induced YAP tyrosine phosphorylation. Sci. China Life Sci. 2020, 63, 697–705. [Google Scholar] [CrossRef]
- Cao, N.; Wang, J.J.; Wu, J.M.; Xu, W.L.; Wang, R.; Chen, X.D.; Feng, Y.N.; Cong, W.W.; Zhang, Y.Y.; Xiao, H.; et al. Glibenclamide alleviates beta adrenergic receptor activation-induced cardiac inflammation. Acta Pharmacol. Sin. 2022, 43, 1243–1250. [Google Scholar] [CrossRef] [PubMed]
- Xin, J.Z.; Wu, J.M.; Hu, G.M.; Gu, H.J.; Feng, Y.N.; Wang, S.X.; Cong, W.W.; Li, M.Z.; Xu, W.L.; Song, Y.; et al. α1-AR overactivation induces cardiac inflammation through NLRP3 inflammasome activation. Acta Pharmacol. Sin. 2020, 41, 311–318. [Google Scholar] [CrossRef]
- Hoch, M.; Fischer, P.; Stapel, B.; Missol-Kolka, E.; Sekkali, B.; Scherr, M.; Favret, F.; Braun, T.; Eder, M.; Schuster-Gossler, K.; et al. Erythropoietin preserves the endothelial differentiation capacity of cardiac progenitor cells and reduces heart failure during anticancer therapies. Cell Stem Cell 2011, 9, 131–143. [Google Scholar] [CrossRef] [Green Version]
- Dakhlallah, D.; Wang, Y.; Bobo, T.A.; Ellis, E.; Mo, X.; Piper, M.G.; Eubank, T.D.; Marsh, C.B. Constitutive AKT Activity Predisposes Lung Fibrosis by Regulating Macrophage, Myofibroblast and Fibrocyte Recruitment and Changes in Autophagy. Adv. Biosci. Biotechnol. 2019, 10, 346–373. [Google Scholar] [CrossRef] [PubMed]
- Petro, J.B.; Rahman, S.M.; Ballard, D.W.; Khan, W.N. Bruton’s tyrosine kinase is required for activation of IkappaB kinase and nuclear factor κB in response to B cell receptor engagement. J. Exp. Med. 2000, 191, 1745–1754. [Google Scholar] [CrossRef] [PubMed]
- Stilgenbauer, S.; Eichhorst, B.; Schetelig, J.; Coutre, S.; Seymour, J.F.; Munir, T.; Puvvada, S.D.; Wendtner, C.M.; Roberts, A.W.; Jurczak, W.; et al. Venetoclax in relapsed or refractory chronic lymphocytic leukaemia with 17p deletion: A multicentre, open-label, phase 2 study. Lancet Oncol. 2016, 17, 768–778. [Google Scholar] [CrossRef]
- Cole, S.W.; Sood, A.K. Molecular pathways: Beta-adrenergic signaling in cancer. Clin. Cancer Res. 2012, 18, 1201–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krum, H.; Abraham, W.T. Heart failure. Lancet 2009, 373, 941–955. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Liu, Y.; Hu, N.; Yu, D.; Zhou, C.; Shi, G.; Zhang, B.; Wei, M.; Liu, J.; Luo, L.; et al. Discovery of Zanubrutinib (BGB-3111), a Novel, Potent, and Selective Covalent Inhibitor of Bruton’s Tyrosine Kinase. J. Med. Chem. 2019, 62, 7923–7940. [Google Scholar] [CrossRef] [PubMed]
- Masso-Valles, D.; Jauset, T.; Serrano, E.; Sodir, N.M.; Pedersen, K.; Affara, N.I.; Whitfield, J.R.; Beaulieu, M.E.; Evan, G.I.; Elias, L.; et al. Ibrutinib exerts potent antifibrotic and antitumor activities in mouse models of pancreatic adenocarcinoma. Cancer Res. 2015, 75, 1675–1681. [Google Scholar] [CrossRef] [Green Version]
- Bataller, R.; Brenner, D.A. Liver fibrosis. J. Clin. Investig. 2005, 115, 209–218. [Google Scholar] [CrossRef]
- Homma, T.; Harada, H.; Koizumi, M. Diagnostic criteria for chronic pancreatitis by the Japan Pancreas Society. Pancreas 1997, 15, 14–15. [Google Scholar] [CrossRef]
- Gu, Y.; Huang, B.; Yang, Y.; Qi, M.; Lu, G.; Xia, D.; Li, H. Ibrutinib Exacerbates Bleomycin-Induced Pulmonary Fibrosis via Promoting Inflammation. Inflammation 2018, 41, 904–913. [Google Scholar] [CrossRef] [PubMed]
- Purvis, G.S.D.; Collino, M.; Aranda-Tavio, H.; Chiazza, F.; O’Riordan, C.E.; Zeboudj, L.; Mohammad, S.; Collotta, D.; Verta, R.; Guisot, N.E.S.; et al. Inhibition of Bruton’s TK regulates macrophage NF-κB and NLRP3 inflammasome activation in metabolic inflammation. Br. J. Pharmacol. 2020, 177, 4416–4432. [Google Scholar] [CrossRef] [PubMed]
- Alemasi, A.; Cao, N.; An, X.; Wu, J.; Gu, H.; Yu, H.; Song, Y.; Wang, H.; Zhang, Y.; Xiao, H.; et al. Exercise Attenuates Acute beta-Adrenergic Overactivation-Induced Cardiac Fibrosis by Modulating Cytokines. J. Cardiovasc. Transl. Res. 2019, 12, 528–538. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (3′-5′) |
|---|---|---|
| Mouse ANP | CTTCCAGGCCATATTGGAG | GGGGGCATGACCTCATCTT |
| Mouse BNP | ACAAGATAGACCGGATCGGA | AGCCAGGAGGTCTTCCTACA |
| Mouse CTGF | CCAACTATGATTAGAGCCAACTG | AGGCACAGGTCTTGATGAAC |
| Mouse Fibronectin | TCGGATACTTCAGCGTCAGGA | TCGGATACTTCAGCGTCAGGA |
| Mouse α-SMA | GTCCCAGACATCAGGGAGTAA | GTCCCAGACATCAGGGAGTAA |
| Mouse Collagen-I | ATGTGGACCCCTCCTGATAGT | ATGTGGACCCCTCCTGATAGT |
| Mouse Collagen-III | TGGTCCTCAGGGTGTAAAGG | GTCCAGCATCACCTTTTGGT |
| Mouse IL-1β | GAAATGCCACCTTTTGACAGTG | TGGATGCTCTCATCAGGACAG |
| Mouse IL-6 | CTGCAAGAGACTTCCATCCAG | AGTGGTATAGACAGGTCTGTTGG |
| Mouse TNF-α | CAGGCGGTGCCTATGTCTC | CGATCACCCCGAAGTTCAGTAG |
| Mouse GAPDH | AGGTCGGTGTGAACGGATTTG | GGGGTCGTTGATGGCAACA |
| Antibody | Company | Item No. |
|---|---|---|
| p-BTK | Affinity | AF0841 |
| BTK | Affinity | DF6472 |
| p-NF-κB | Affinity | AF2006 |
| NF-κB | Affinity | AF0874 |
| p-PI3K | Affinity | AF3241 |
| PI3K | Affinity | AF6241 |
| p-Akt | Affinity | AF0016 |
| Akt | Affinity | AF6261 |
| p-STAT3 | Affinity | AF0016 |
| STAT3 | Affinity | AF6294 |
| Fibronectin | Proteintech | 15613-1-AP |
| α-SMA | Affinity | AF1032 |
| Collagen-I | Cell Signaling Technology | 72026 |
| Mac-3 | BD Biosciences | 550292 |
| β-Tubulin | Affinity | AF7011 |
| GAPDH | Affinity | AF7021 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, W.; Zhu, S.; Liu, J.; Liu, Z.; Zhou, H.; Zhang, Q.; Yang, Y.; Chen, L.; Guo, X.; Zhang, T.; et al. Zanubrutinib Ameliorates Cardiac Fibrosis and Inflammation Induced by Chronic Sympathetic Activation. Molecules 2023, 28, 6035. https://doi.org/10.3390/molecules28166035
Li W, Zhu S, Liu J, Liu Z, Zhou H, Zhang Q, Yang Y, Chen L, Guo X, Zhang T, et al. Zanubrutinib Ameliorates Cardiac Fibrosis and Inflammation Induced by Chronic Sympathetic Activation. Molecules. 2023; 28(16):6035. https://doi.org/10.3390/molecules28166035
Chicago/Turabian StyleLi, Wenqi, Shuwen Zhu, Jing Liu, Zhigang Liu, Honggang Zhou, Qianyi Zhang, Yue Yang, Li Chen, Xiaowei Guo, Tiantian Zhang, and et al. 2023. "Zanubrutinib Ameliorates Cardiac Fibrosis and Inflammation Induced by Chronic Sympathetic Activation" Molecules 28, no. 16: 6035. https://doi.org/10.3390/molecules28166035