


Ethanolic Extract Propolis-Loaded Niosomes Diminish Phospholipase B1, Biofilm Formation, and Intracellular Replication of Cryptococcus neoformans in Macrophages


 , ,
, ,  , and
, and 
Abstract
:1. Introduction
2. Results
2.1. Physicochemical Characterization of Nio-EEP
2.1.1. Particle Size, Polydispersity Index (PDI), Zeta Potential (ZP), Entrapment Efficiency (EE), Loading Capacity (LC), and Morphology
2.1.2. Chemical Composition
2.1.3. In Vitro Release Study
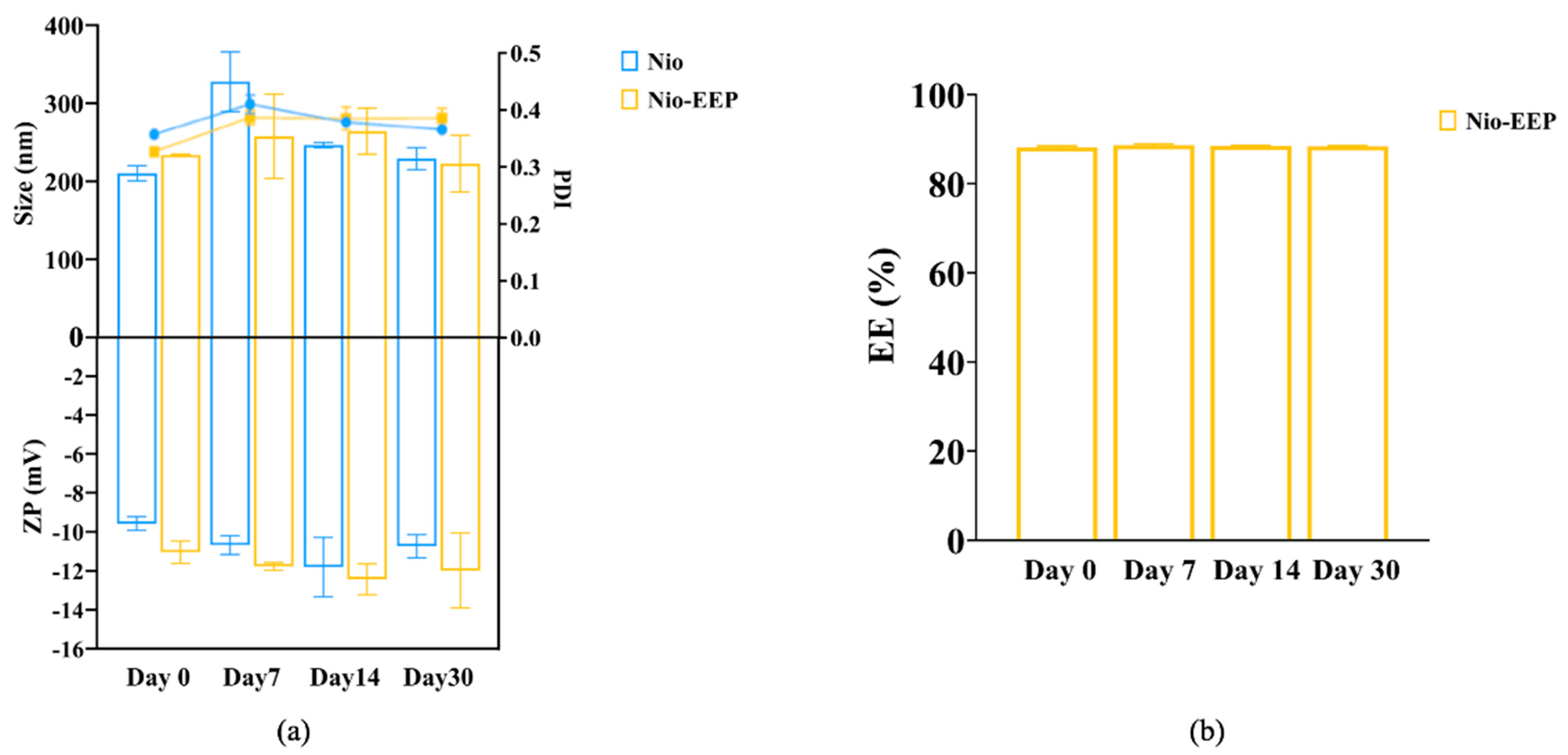
2.1.4. Stability Testing
2.2. In Vitro Biological Activity of Nio-EEP
2.2.1. Cytotoxicity Assay of Niosomes
2.2.2. Anti-Fungal Susceptibility Testing
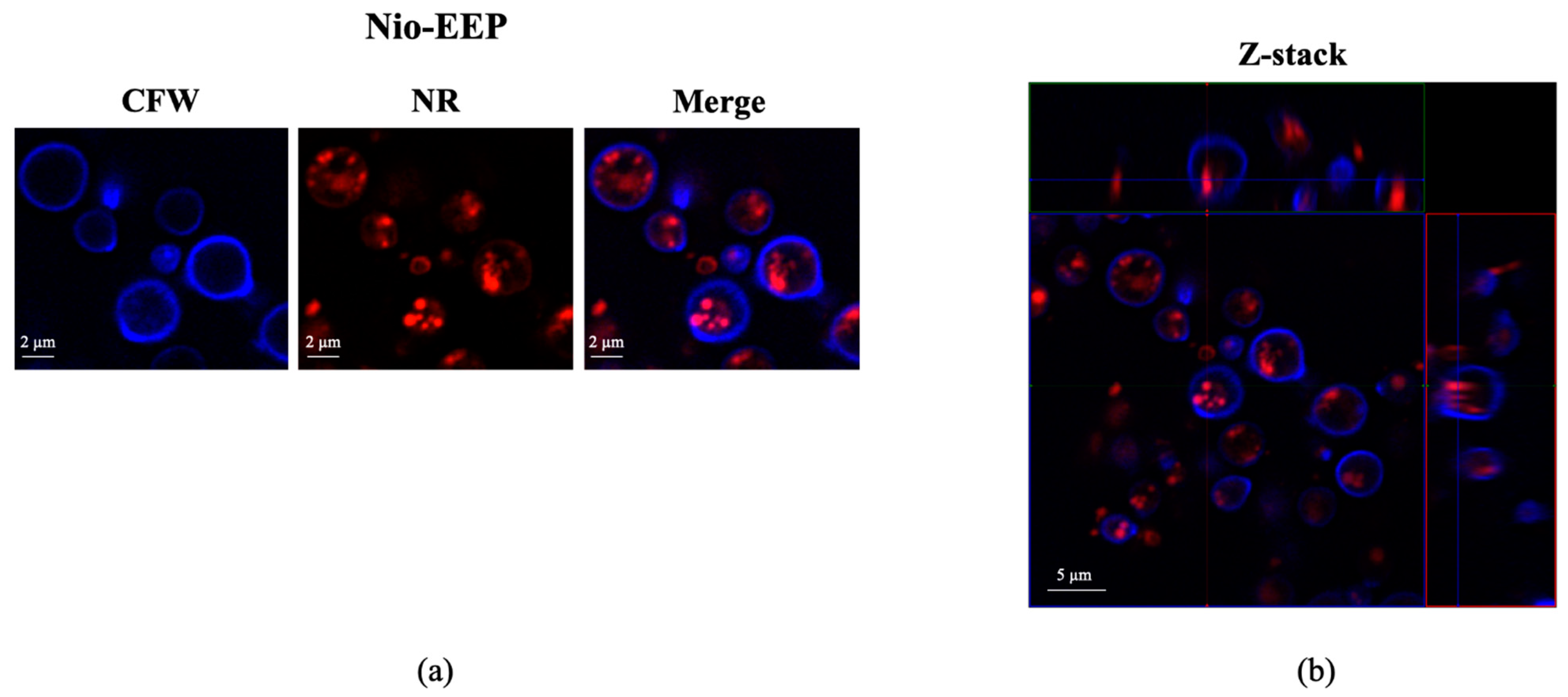
2.2.3. Localization of Nio-EEP
2.3. Anti-Virulence Factors of Nio-EEP
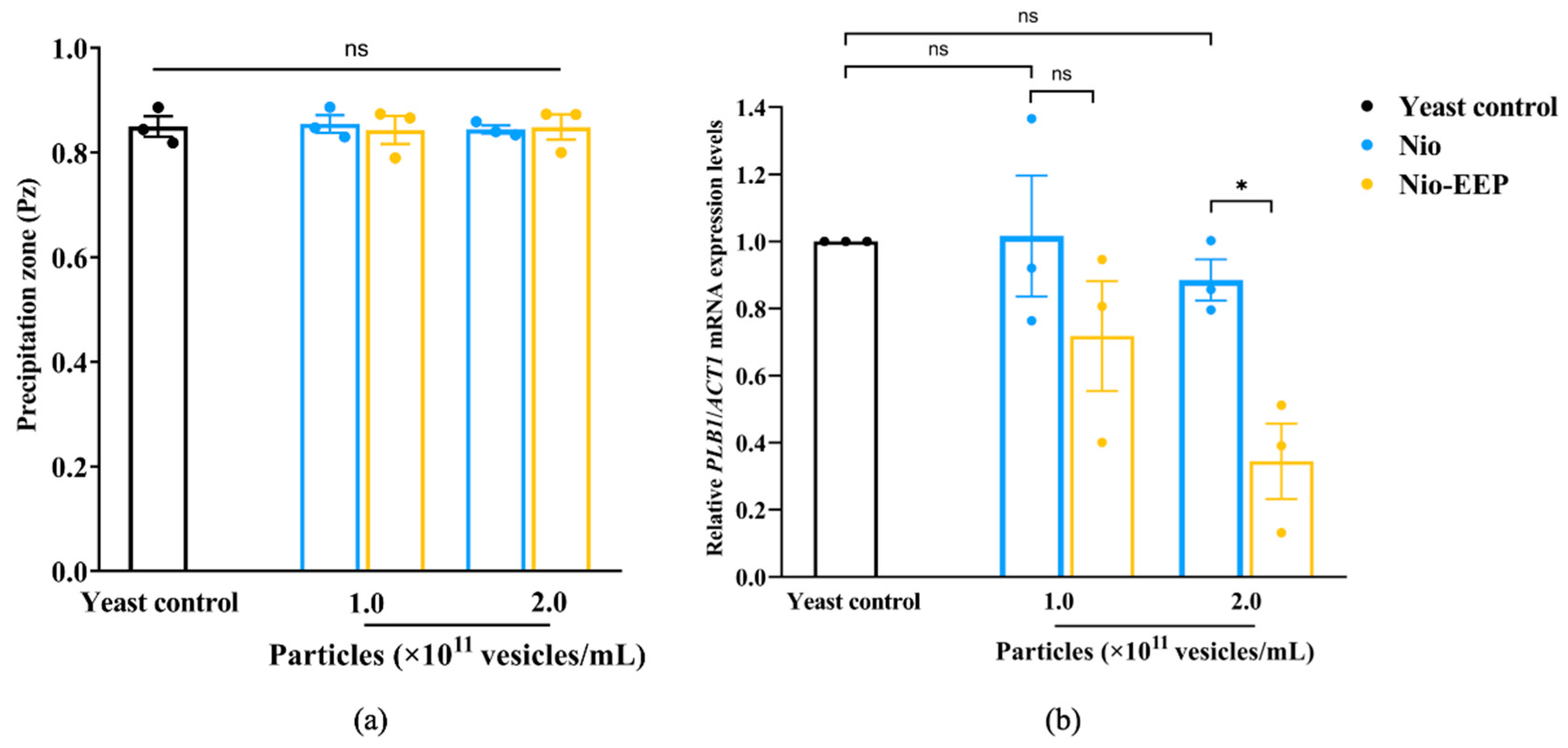
2.3.1. Phospholipase Production
2.3.2. Biofilm Formation
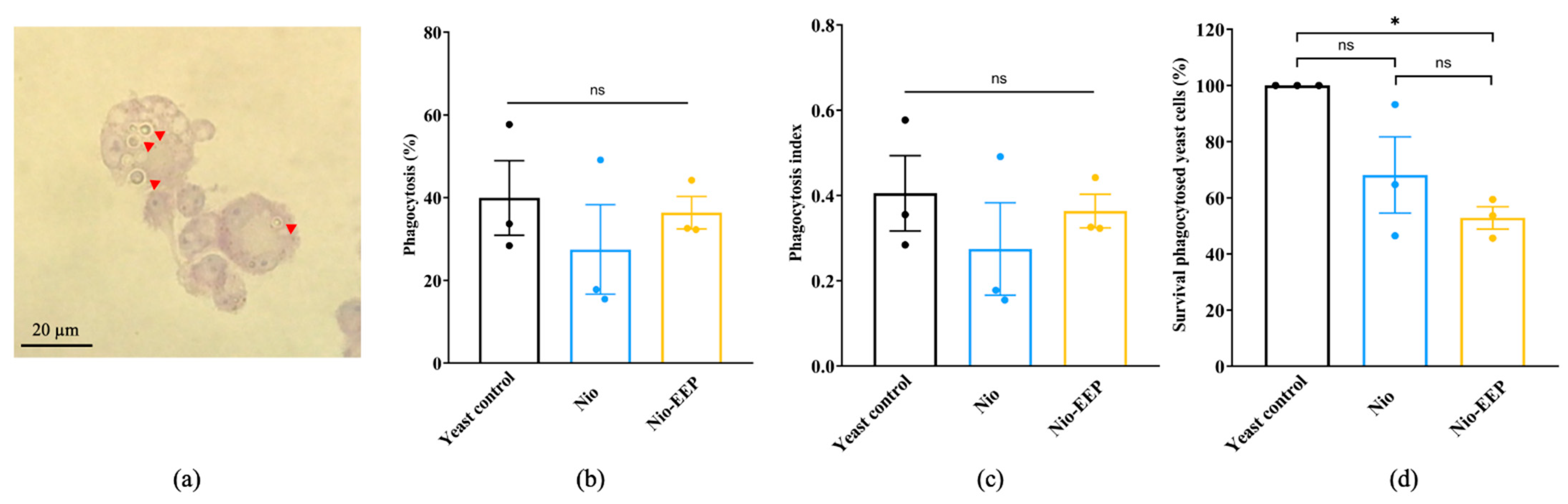
2.3.3. Nio-EEP-Induced Intracellular Killing
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Yeast and Cell Lines
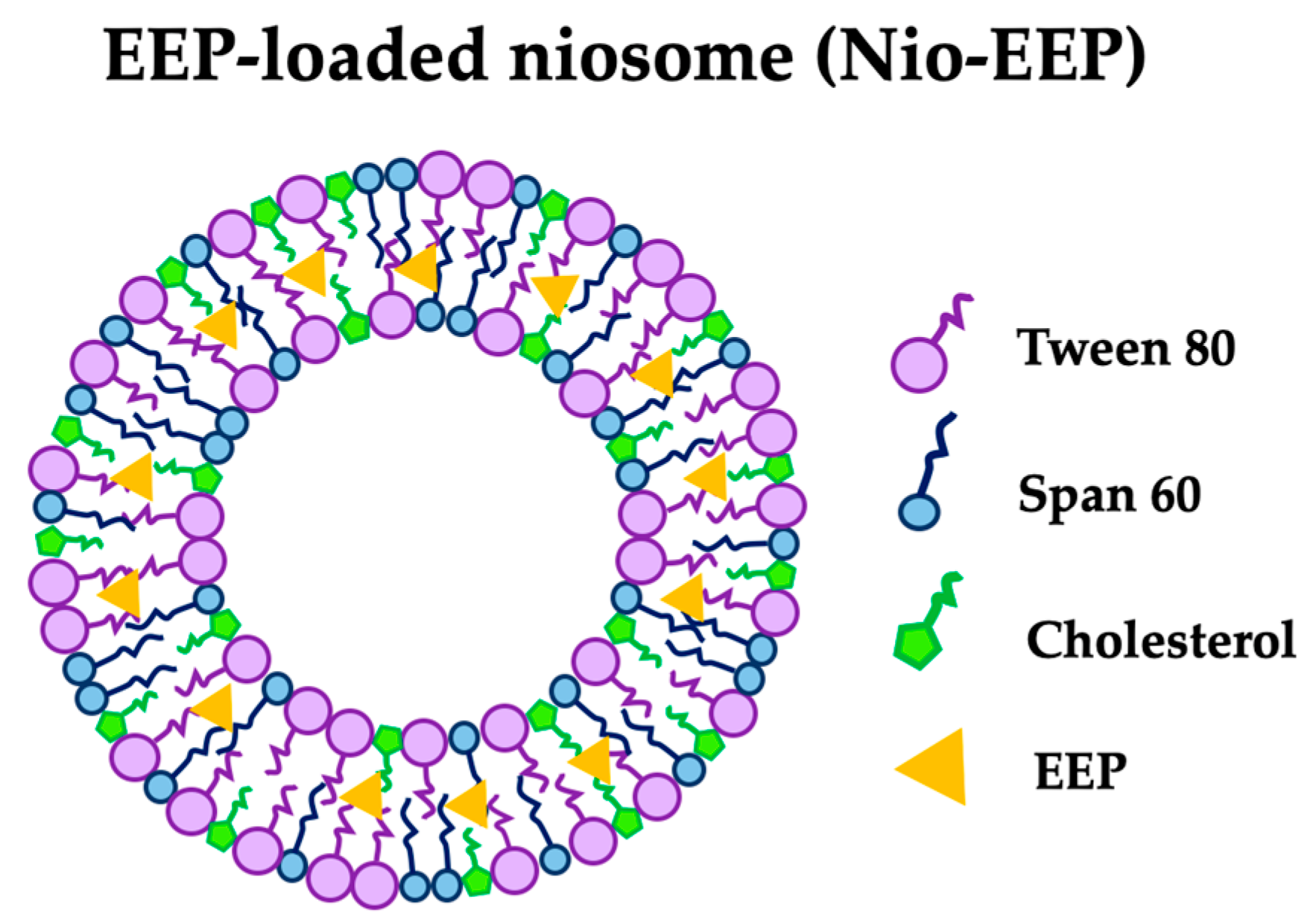
4.3. Formulation of Nio-EEP
4.4. Physicochemical Characterization of Niosomes
4.4.1. Particle Size, Polydispersity Index (PDI), Zeta Potential (ZP), Morphological Analysis, and Stability Testing
4.4.2. Nio-EEP Chemical Structure
4.4.3. Entrapment Efficiency (EE) and Loading Capacity (LC)
4.4.4. In Vitro Release Study
4.5. In Vitro Bioactivity of Nio-EEP
4.5.1. Cytotoxicity Assay
4.5.2. Anti-Fungal Susceptibility Testing
4.5.3. Yeast Cell Uptake of Nio-EEP
4.6. Effect of Nio-EEP on Virulence Factors of C. neoformans
4.6.1. Phospholipase Production
4.6.2. Biofilm Formation
4.6.3. Virulence-Related mRNA Expression
4.7. Phagocytosis Assay
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Setianingrum, F.; Rautemaa-Richardson, R.; Denning, D.W. Pulmonary cryptococcosis: A review of pathobiology and clinical aspects. Med. Mycol. 2019, 57, 133–150. [Google Scholar] [CrossRef]
- Sabiiti, W.; May, R.C. Mechanisms of infection by the human fungal pathogen Cryptococcus neoformans. Future Microbiol. 2012, 7, 1297–1313. [Google Scholar] [CrossRef]
- Chen, S.C.; Wright, L.C.; Golding, J.C.; Sorrell, T.C. Purification and characterization of secretory phospholipase B, lysophospholipase and lysophospholipase/transacylase from a virulent strain of the pathogenic fungus Cryptococcus neoformans. Biochem. J. 2000, 347, 431–439. [Google Scholar] [CrossRef]
- Taylor-Smith, L.M. Cryptococcus-epithelial interactions. J. Fungi 2017, 3, 53. [Google Scholar] [CrossRef]
- Alvarez, M.; Casadevall, A. Phagosome extrusion and host-cell survival after Cryptococcus neoformans phagocytosis by macrophages. Curr. Biol. 2006, 16, 2161–2165. [Google Scholar] [CrossRef] [PubMed]
- Ma, H.; Croudace, J.E.; Lammas, D.A.; May, R.C. Expulsion of live pathogenic yeast by macrophages. Curr. Biol. 2006, 16, 2156–2160. [Google Scholar] [CrossRef] [PubMed]
- Lopes, W.; Vainstein, M.H.; De Sousa Araujo, G.R.; Frases, S.; Staats, C.C.; de Almeida, R.M.C.; Schrank, A.; Kmetzsch, L.; Vainstein, M.H. Geometrical distribution of Cryptococcus neoformans mediates flower-like biofilm development. Front. Microbiol. 2017, 8, 2534. [Google Scholar] [CrossRef] [PubMed]
- Martinez, L.R.; Casadevall, A. Biofilm formation by Cryptococcus neoformans. Microbiol. Spectr. 2015, 3, 3. [Google Scholar] [CrossRef]
- Martinez, L.R.; Casadevall, A. Cryptococcus neoformans cells in biofilms are less susceptible than planktonic cells to antimicrobial molecules produced by the innate immune system. Infect. Immun. 2006, 74, 6118–6123. [Google Scholar] [CrossRef] [PubMed]
- Perfect, J.R.; Dismukes, W.E.; Dromer, F.; Goldman, D.L.; Graybill, J.R.; Hamill, R.J.; Harrison, T.S.; Larsen, R.A.; Lortholary, O.; Nguyen, M.H.; et al. Clinical practice guidelines for the management of cryptococcal disease: 2010 update by the infectious diseases society of america. Clin. Infect. Dis. 2010, 50, 291–322. [Google Scholar] [CrossRef] [PubMed]
- Iadnut, A.; Mamoon, K.; Thammasit, P.; Pawichai, S.; Tima, S.; Preechasuth, K.; Kaewkod, T.; Tragoolpua, Y.; Tragoolpua, K. In Vitro antifungal and antivirulence activities of biologically synthesized ethanolic extract of propolis-loaded PLGA nanoparticles against Candida albicans. Evid. Based Complement. Alternat Med. 2019, 2019, 3715481. [Google Scholar] [CrossRef]
- Gheflati, A.; Dehnavi, Z.; Ghannadzadeh, Y.A.; Khorasanchi, Z.; Raeisi-Dehkordi, H.; Ranjbar, G. The effects of propolis supplementation on metabolic parameters: A systematic review and meta-analysis of randomized controlled clinical trials. Avicenna J. Phytomed 2021, 11, 551–565. [Google Scholar] [CrossRef]
- Hallajzadeh, J.; Milajerdi, A.; Amirani, E.; Attari, V.E.; Maghsoudi, H.; Mirhashemi, S.M. Effects of propolis supplementation on glycemic status, lipid profiles, inflammation and oxidative stress, liver enzymes, and body weight: A systematic review and meta-analysis of randomized controlled clinical trials. J. Diabetes Metab. Disord. 2021, 20, 831–843. [Google Scholar] [CrossRef] [PubMed]
- Zulhendri, F.; Lesmana, R.; Tandean, S.; Christoper, A.; Chandrasekaran, K.; Irsyam, I.; Suwantika, A.A.; Abdulah, R.; Wathoni, N. Recent update on the anti-inflammatory activities of propolis. Molecules 2022, 27, 8473. [Google Scholar] [CrossRef]
- Corrêa, J.L.; Veiga, F.F.; Jarros, I.C.; Costa, M.I.; Castilho, P.F.; de Oliveira, K.M.P.; Rosseto, H.C.; Bruschi, M.L.; Svidzinski, T.I.E.; Negri, M. Propolis extract has bioactivity on the wall and cell membrane of Candida albicans. J. Ethnopharmacol. 2020, 256, 112791. [Google Scholar] [CrossRef]
- Fernandes, F.F.; Dias, A.L.; Ramos, C.L.; Ikegaki, M.; de Siqueira, A.M.; Franco, M.C. The “In Vitro” antifungal activity evaluation of propolis g12 ethanol extract on Cryptococcus neoformans. Rev. Inst. Med. Trop. Sao Paulo 2007, 49, 93–95. [Google Scholar] [CrossRef]
- Thammasit, P.; Iadnut, A.; Mamoon, K.; Khacha-Ananda, S.; Chupradit, K.; Tayapiwatana, C.; Kasinrerk, W.; Tragoolpua, Y.; Tragoolpua, K. A potential of propolis on major virulence factors of Cryptococcus neoformans. Microb. Pathog. 2018, 123, 296–303. [Google Scholar] [CrossRef]
- Thammasit, P.; Tharinjaroen, C.S.; Tragoolpua, Y.; Rickerts, V.; Georgieva, R.; Bäumler, H.; Tragoolpua, K. Targeted propolis-loaded poly (butyl) cyanoacrylate nanoparticles: An alternative drug delivery tool for the treatment of cryptococcal meningitis. Front. Pharmacol. 2021, 12, 723727. [Google Scholar] [CrossRef]
- Garcia-Verdugo, I.; Descamps, D.; Chignard, M.; Touqui, L.; Sallenave, J.M. Lung protease/anti-protease network and modulation of mucus production and surfactant activity. Biochimie 2010, 92, 1608–1617. [Google Scholar] [CrossRef] [PubMed]
- Pasarin, D.; Ghizdareanu, A.I.; Enascuta, C.E.; Matei, C.B.; Bilbie, C.; Paraschiv-Palada, L.; Veres, P.A. Coating materials to increase the stability of liposomes. Polymers 2023, 15, 782. [Google Scholar] [CrossRef] [PubMed]
- Terzano, C.; Allegra, L.; Alhaique, F.; Marianecci, C.; Carafa, M. Non-phospholipid vesicles for pulmonary glucocorticoid delivery. Eur. J. Pharm. Biopharm. 2005, 59, 57–62. [Google Scholar] [CrossRef]
- Milan, A.; Mioc, A.; Prodea, A.; Mioc, M.; Buzatu, R.; Ghiulai, R.; Racoviceanu, R.; Caruntu, F.; Şoica, C. The optimized delivery of triterpenes by liposomal nanoformulations: Overcoming the challenges. Int. J. Mol. Sci. 2022, 23, 1140. [Google Scholar] [CrossRef] [PubMed]
- Alsaadi, M.; Italia, J.L.; Mullen, A.B.; Ravi Kumar, M.N.; Candlish, A.A.; Williams, R.A.; Shaw, C.D.; Al Gawhari, F.; Coombs, G.H.; Wiese, M.; et al. The efficacy of aerosol treatment with non-ionic surfactant vesicles containing amphotericin B in rodent models of leishmaniasis and pulmonary aspergillosis infection. J. Control Release 2012, 160, 685–691. [Google Scholar] [CrossRef] [PubMed]
- Patel, J.; Ketkar, S.; Patil, S.; Fearnley, J.; Mahadik, K.R.; Paradkar, A.R. Potentiating antimicrobial efficacy of propolis through niosomal-based system for administration. Integr. Med. Res. 2015, 4, 94–101. [Google Scholar] [CrossRef]
- Sangboonruang, S.; Semakul, N.; Suriyaprom, S.; Kitidee, K.; Khantipongse, J.; Intorasoot, S.; Tharinjaroen, C.; Wattananandkul, U.; Butr-Indr, B.; Phunpae, P.; et al. Nano-delivery system of ethanolic extract of propolis targeting mycobacterium tuberculosis via aptamer-modified-niosomes. Nanomaterials 2023, 13, 269. [Google Scholar] [CrossRef] [PubMed]
- Mozafari, M.; Mazaheri, E.; Dormiani, K. Simple equations pertaining to the particle number and surface area of metallic, polymeric, lipidic and vesicular nanocarriers. Sci. Pharm. 2021, 89, 15. [Google Scholar] [CrossRef]
- Wenfei, L.; Ruixiang, Y.; Yang, L.; Zhanhai, Y.; Shumei, L. Preparation and characterization of reactive type dripping agent containing α-double bond. Asian J. Chem. 2014, 26, 8577–8580. [Google Scholar] [CrossRef]
- Rustandi, R.R. Polysorbate 80 and histidine quantitative analysis by nmr in the presence of virus-like particles. Electrophor. 2022, 43, 1408–1414. [Google Scholar] [CrossRef]
- Khatun, R.; Hunter, H.; Magcalas, W.; Sheng, Y.; Carpick, B.; Kirkitadze, M. Nuclear magnetic resonance (NMR) study for the detection and quantitation of cholesterol in HSV529 therapeutic vaccine candidate. Comput. Struct. Biotechnol. J. 2017, 15, 14–20. [Google Scholar] [CrossRef]
- Kasote, D.M.; Pawar, M.V.; Bhatia, R.S.; Nandre, V.S.; Gundu, S.S.; Jagtap, S.D.; Kulkarni, M.V. HPLC, NMR based chemical profiling and biological characterisation of indian propolis. Fitoterapia 2017, 122, 52–60. [Google Scholar] [CrossRef]
- Tran, C.T.N.; Brooks, P.R.; Bryen, T.J.; Williams, S.; Berry, J.; Tavian, F.; McKee, B.; Tran, T.D. Quality assessment and chemical diversity of australian propolis from Apis mellifera bees. Sci. Rep. 2022, 12, 13574. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO) Report. WHO Fungal Priority Pathogens List to Guide Research, Development and Public Health Action. Available online: https://www.who.int/publications/i/item/9789240060241 (accessed on 16 December 2022).
- Bonifácio, B.V.; Silva, P.B.; Ramos, M.A.; Negri, K.M.; Bauab, T.M.; Chorilli, M. Nanotechnology-based drug delivery systems and herbal medicines: A review. Int. J. Nanomed. 2014, 9, 1–15. [Google Scholar] [CrossRef]
- Hidalgo, A.; Garcia-Mouton, C.; Autilio, C.; Carravilla, P.; Orellana, G.; Islam, M.N.; Bhattacharya, J.; Bhattacharya, S.; Cruz, A.; Pérez-Gil, J. Pulmonary surfactant and drug delivery: Vehiculization, release and targeting of surfactant/tacrolimus formulations. J. Control Release 2021, 329, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Junyaprasert, V.B.; Singhsa, P.; Suksiriworapong, J.; Chantasart, D. Physicochemical properties and skin permeation of Span 60/Tween 60 niosomes of ellagic acid. Int. J. Pharm. 2012, 423, 303–311. [Google Scholar] [CrossRef]
- Nematollahi, M.H.; Pardakhty, A.; Torkzadeh-Mahani, M.; Mehrabani, M.; Asadikaram, G. Changes in physical and chemical properties of niosome membrane induced by cholesterol: A promising approach for niosome bilayer intervention. RSC Adv. 2017, 7, 49463–49472. [Google Scholar] [CrossRef]
- Vyas, S.P.; Singh, R.P.; Jain, S.; Mishra, V.; Mahor, S.; Singh, P.; Gupta, P.N.; Rawat, A.; Dubey, P. Non-ionic surfactant based vesicles (niosomes) for non-invasive topical genetic immunization against hepatitis B. Int. J. Pharm. 2005, 296, 80–86. [Google Scholar] [CrossRef] [PubMed]
- Soliman, M.S.; Abd-Allah, F.I.; Hussain, T.; Saeed, N.M.; El-Sawy, H.S. Date seed oil loaded niosomes: Development, optimization and anti-inflammatory effect evaluation on rats. Drug Dev. Ind. Pharm. 2018, 44, 1185–1197. [Google Scholar] [CrossRef]
- Chinembiri, T.N.; Gerber, M.; du Plessis, L.H.; du Preez, J.L.; Hamman, J.H.; du Plessis, J. Topical delivery of Withania somnifera crude extracts in niosomes and solid lipid nanoparticles. Pharmacogn. Mag. 2017, 13, 663–671. [Google Scholar] [CrossRef]
- Ilhan-Ayisigi, E.; Ulucan, F.; Saygili, E.; Saglam-Metiner, P.; Gulce-Iz, S.; Yesil-Celiktas, O. Nano-vesicular formulation of propolis and cytotoxic effects in a 3D spheroid model of lung cancer. J. Sci. Food Agric. 2020, 100, 3525–3535. [Google Scholar] [CrossRef]
- Sangboonruang, S.; Semakul, N.; Obeid, M.A.; Ruano, M.; Kitidee, K.; Anukool, U.; Pringproa, K.; Chantawannakul, P.; Ferro, V.A.; Tragoolpua, Y.; et al. Potentiality of melittin-loaded niosomal vesicles against vancomycin-intermediate Staphylococcus aureus and staphylococcal skin infection. Int. J. Nanomed. 2021, 16, 7639–7661. [Google Scholar] [CrossRef]
- Gupta, A.; Aggarwal, G.; Singla, S.; Arora, R. Transfersomes: A novel vesicular carrier for enhanced transdermal delivery of sertraline: Development, characterization, and performance evaluation. Sci. Pharm. 2012, 80, 1061–1080. [Google Scholar] [CrossRef] [PubMed]
- Opatha, S.A.T.; Titapiwatanakun, V.; Chutoprapat, R. Transfersomes: A promising nanoencapsulation technique for transdermal drug delivery. Pharmaceutics 2020, 12, 855. [Google Scholar] [CrossRef] [PubMed]
- Obeid, M.A.; Gany, S.A.S.; Gray, A.I.; Young, L.; Igoli, J.O.; Ferro, V.A. Niosome-encapsulated balanocarpol: Compound isolation, characterisation, and cytotoxicity evaluation against human breast and ovarian cancer cell lines. Nanotechnology 2020, 31, 195101. [Google Scholar] [CrossRef] [PubMed]
- Un, R.; Barlas, F.; Yavuz, M.; Ag Seleci, D.; Seleci, M.; Gümüş, Z.; Guler Celik, E.; Demir, B.; Can, M.; Coşkunol, H.; et al. Phyto-niosomes: In vitro assessment of the novel nanovesicles containing marigold extract. Int. J. Polymer Mater. 2015, 64, 927–937. [Google Scholar] [CrossRef]
- Joseph, E.; Singhvi, G. Chapter 4—Multifunctional nanocrystals for cancer therapy: A potential nanocarrier. In Nanomaterials for Drug Delivery and Therapy, 1st ed.; Grumezescu, A.M., Ed.; William Andrew: Norwich, CT, USA; Elsevier: Amsterdam, The Netherlands, 2019; pp. 91–116. ISBN 978-0-12-816505-8. [Google Scholar]
- Chen, S.; Hanning, S.; Falconer, J.; Locke, M.; Wen, J. Recent advances in non-ionic surfactant vesicles (niosomes): Fabrication, characterization, pharmaceutical and cosmetic applications. Eur. J. Pharm. Biopharm. 2019, 144, 18–39. [Google Scholar] [CrossRef]
- Hao, Y.; Zhao, F.; Li, N.; Yang, Y.; Li, K. Studies on a high encapsulation of colchicine by a niosome system. Int. J. Pharm. 2002, 244, 73–80. [Google Scholar] [CrossRef]
- Hichmah, H.N. Effect of surfactant concentration on the entrapment efficiency niosomes aqueous extract of cassava leaves (Manihot esculenta Crantz). Asian J. Pharm. 2019, 13, 276–281. [Google Scholar] [CrossRef]
- Shirsand, S.; Para, M.; Nagendrakumar, D.; Kanani, K.; Keerthy, D. Formulation and evaluation of ketoconazole niosomal gel drug delivery system. Int. J. Pharm. Investig. 2012, 2, 201–207. [Google Scholar] [CrossRef]
- Fu, Y.; Kao, W.J. Drug release kinetics and transport mechanisms of non-degradable and degradable polymeric delivery systems. Expert. Opin. Drug Deliv. 2010, 7, 429–444. [Google Scholar] [CrossRef]
- Sukhanova, A.; Bozrova, S.; Sokolov, P.; Berestovoy, M.; Karaulov, A.; Nabiev, I. Dependence of nanoparticle toxicity on their physical and chemical properties. Nanoscale Res. Lett. 2018, 13, 44. [Google Scholar] [CrossRef]
- Kyselova, Z. Toxicological aspects of the use of phenolic compounds in disease prevention. Interdiscip. Toxicol. 2011, 4, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; He, P.Y.; Zhang, Y.; Li, N. Natural products targeting the mitochondria in cancers. Molecules 2020, 26, 92. [Google Scholar] [CrossRef]
- Rejman, J.; Oberle, V.; Zuhorn, I.S.; Hoekstra, D. Size-dependent internalization of particles via the pathways of clathrin- and caveolae-mediated endocytosis. Biochem. J. 2004, 377, 159–169. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo Borgognoni, C.; Kim, J.H.; Zucolotto, V.; Fuchs, H.; Riehemann, K. Human macrophage responses to metal-oxide nanoparticles: A review. Artif. Cells Nanomed. Biotechnol. 2018, 46, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Ganendren, R.; Carter, E.; Sorrell, T.; Widmer, F.; Wright, L. Phospholipase B activity enhances adhesion of Cryptococcus neoformans to a human lung epithelial cell line. Microbes Infect. 2006, 8, 1006–1015. [Google Scholar] [CrossRef] [PubMed]
- Cox, G.M.; McDade, H.C.; Chen, S.C.; Tucker, S.C.; Gottfredsson, M.; Wright, L.C.; Sorrell, T.C.; Leidich, S.D.; Casadevall, A.; Ghannoum, M.A.; et al. Extracellular phospholipase activity is a virulence factor for Cryptococcus neoformans. Mol. Microbiol. 2001, 39, 166–175. [Google Scholar] [CrossRef]
- Ghannoum, M.A. Potential role of phospholipases in virulence and fungal pathogenesis. Clin. Microbiol. Rev. 2000, 13, 122–143. [Google Scholar] [CrossRef]
- Kumari, P.; Arora, N.; Chatrath, A.; Gangwar, R.; Pruthi, V.; Poluri, K.M.; Prasad, R. Delineating the biofilm inhibition mechanisms of phenolic and aldehydic terpenes against Cryptococcus neoformans. ACS Omega 2019, 4, 17634–17648. [Google Scholar] [CrossRef]
- Wills, E.A.; Roberts, I.S.; Del Poeta, M.; Rivera, J.; Casadevall, A.; Cox, G.M.; Perfect, J.R. Identification and characterization of the Cryptococcus neoformans phosphomannose isomerase-encoding gene, Man1, and its impact on pathogenicity. Mol. Microbiol. 2001, 40, 610–620. [Google Scholar] [CrossRef]
- Bar-Peled, M.; Griffith, C.L.; Doering, T.L. Functional cloning and characterization of a UDP- glucuronic acid decarboxylase: The pathogenic fungus Cryptococcus neoformans elucidates UDP-xylose synthesis. Proc. Natl. Acad. Sci. USA 2001, 98, 12003–12009. [Google Scholar] [CrossRef]
- Bar-Peled, M.; Griffith, C.L.; Ory, J.J.; Doering, T.L. Biosynthesis of UDP-GlcA, a key metabolite for capsular polysaccharide synthesis in the pathogenic fungus Cryptococcus neoformans. Biochem. J. 2004, 381, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Griffith, C.L.; Klutts, J.S.; Zhang, L.; Levery, S.B.; Doering, T.L. UDP-glucose dehydrogenase plays multiple roles in the biology of the pathogenic fungus Cryptococcus neoformans. J. Biol. Chem. 2004, 279, 51669–51676. [Google Scholar] [CrossRef] [PubMed]
- Ozdal, T.; Capanoglu, E.; Altay, F. A review on protein–phenolic interactions and associated changes. Food Res. Int. 2013, 51, 954–970. [Google Scholar] [CrossRef]
- Casadevall, A.; Pirofski, L.A. A new synthesis for antibody-mediated immunity. Nat. Immunol. 2011, 13, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Mamoon, K.; Thammasit, P.; Iadnut, A.; Kitidee, K.; Anukool, U.; Tragoolpua, Y.; Tragoolpua, K. Unveiling the properties of thai stingless bee propolis via diminishing cell wall-associated cryptococcal melanin and enhancing the fungicidal activity of macrophages. Antibiotics 2020, 9, 420. [Google Scholar] [CrossRef] [PubMed]
- Zaragoza, O. Basic principles of the virulence of cryptococcus. Virulence 2019, 10, 490–501. [Google Scholar] [CrossRef]
- Chayakulkeeree, M.; Johnston, S.A.; Oei, J.B.; Lev, S.; Williamson, P.R.; Wilson, C.F.; Zuo, X.; Leal, A.L.; Vainstein, M.H.; Meyer, W.; et al. Sec14 is a specific requirement for secretion of phospholipase B1 and pathogenicity of Cryptococcus neoformans. Mol. Microbiol. 2011, 80, 1088–1101. [Google Scholar] [CrossRef]
- Shukr, M.H. Novel In Situ gelling ocular inserts for voriconazole-loaded niosomes: Design, In Vitro characterisation and In Vivo evaluation of the ocular irritation and drug pharmacokinetics. J. Microencapsul. 2016, 33, 71–79. [Google Scholar] [CrossRef]
- Cao, L.B.; Zeng, S.; Zhao, W. Highly stable pegylated poly(lactic-co-glycolic acid) (PLGA) nanoparticles for the effective delivery of docetaxel in prostate cancers. Nanoscale Res. Lett. 2016, 11, 305. [Google Scholar] [CrossRef]
- Son, Y.J.; McConville, J.T. Development of a standardized dissolution test method for inhaled pharmaceutical formulations. Int. J. Pharm. 2009, 382, 15–22. [Google Scholar] [CrossRef]
- Zajac, M.; Dreano, E.; Edwards, A.; Planelles, G.; Sermet-Gaudelus, I. Airway surface liquid pH regulation in airway epithelium current understandings and gaps in knowledge. Int. J. Mol. Sci. 2021, 22, 3384. [Google Scholar] [CrossRef] [PubMed]
- Honarvari, B.; Karimifard, S.; Akhtari, N.; Mehrarya, M.; Moghaddam, Z.S.; Ansari, M.J.; Jalil, A.T.; Matencio, A.; Trotta, F.; Yeganeh, F.E.; et al. Folate-targeted curcumin-loaded niosomes for site-specific delivery in breast cancer treatment: In silico and In Vitro study. Molecules 2022, 27, 4634. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.C.; Muller, M.; Zhou, J.Z.; Wright, L.C.; Sorrell, T.C. Phospholipase activity in Cryptococcus neoformans: A new virulence factor? J. Infect. Dis. 1997, 175, 414–420. [Google Scholar] [CrossRef]
- Benaducci, T.; Sardi, J.; Lourencetti, N.; Scorzoni, L.; Gullo, L.F.; Rossi, S.; Derissi, J.; Prata, M.; Almeida, A.; Mendes, G.M.J. Virulence of Cryptococcus sp. biofilms In Vitro and In Vivo using Galleria mellonella as an alternative model. Front. Microbiol. 2016, 7, 290. [Google Scholar] [CrossRef] [PubMed]
- Fonseca, F.L.; Guimarães, A.J.; Kmetzsch, L.; Dutra, F.F.; Silva, F.D.; Taborda, C.P.; Araujo, G.D.S.; Frases, S.; Staats, C.C.; Bozza, M.T.; et al. Binding of the wheat germ lectin to Cryptococcus neoformans chitooligomers affects multiple mechanisms required for fungal pathogenesis. Fungal Genet. Biol. 2013, 60, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Latouche, G.N.; Sorrell, T.C.; Meyer, W. Isolation and characterisation of the phospholipase B gene of Cryptococcus neoformans var. gattii. FEMS Yeast Res. 2002, 2, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Nicola, A.M.; Casadevall, A. In Vitro measurement of phagocytosis and killing of Cryptococcus neoformans by macrophages. Methods Mol. Biol. 2012, 844, 189–197. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formulations | Nio | Nio-EEP | ||||||
|---|---|---|---|---|---|---|---|---|
| Size (nm) | PDI | ZP (mV) | Size (nm) | PDI | ZP (mV) | EE (%) | LC (%) | |
| F1 | 255.53 ± 25.36 | 0.37 ± 0.06 | −9.38 ± 1.58 | 268.53 ± 10.89 | 0.32 ± 0.01 | −10.54 ± 1.37 | 88.13 ± 0.01 | 82.95 * ± 0.01 |
| F2 | 108.30 ± 6.53 | 0.34 ± 0.03 | −8.62 ± 1.41 | 152.15 ± 34.00 | 0.35 ± 0.05 | −10.05 ± 0.20 | 88.45 ± 0.00 | 45.76 # ± 0.00 |
| F3 | 253.59 ± 20.49 | 0.32 ± 0.04 | −10.51 ± 0.88 | 168.07 ± 23.91 | 0.32 ± 0.05 | −9.92 ± 0.30 | 86.75 ± 0.05 | 50.49 ± 0.03 |
| Formulations | SP60: TW80: CHOL (mM Ratio) | SP60 (mg) | TW80 (mg) | CHOL (mg) | EEP (mg/mL) |
|---|---|---|---|---|---|
| F1 | 1:1:1 | 4.3 | 13.1 | 3.8 | 2 |
| F2 | 2:2:1 | 8.6 | 26.2 | 3.8 | 2 |
| F3 | 1:2:1 | 4.3 | 26.2 | 3.8 | 2 |
| Primers | Primer Sequences (5′-3′) | References |
|---|---|---|
| PLB1 | TGATGAATGAGAGCACGGAAGC | [78] |
| CTCAGACCAGCCCAGTAGCT | ||
| MAN1 | GGCCTACGCTGAATTATGGA | This study |
| GTAAAGAGCCGTCCTTGCAG | ||
| UGD1 | GAGGAGGCTTGTGCTAATGC | This study |
| GACGACCTTGAAACCGATGT | ||
| UXS1 | AGCTGCATTTTACTCATCCCT | This study |
| TCCTTGATGTAGGCGGGAGA | ||
| ACT1 | CCTTGCTCCTTCTTCTAT | [67] |
| CTCGTCGTATTCGCTCTT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kietrungruang, K.; Sookkree, S.; Sangboonruang, S.; Semakul, N.; Poomanee, W.; Kitidee, K.; Tragoolpua, Y.; Tragoolpua, K. Ethanolic Extract Propolis-Loaded Niosomes Diminish Phospholipase B1, Biofilm Formation, and Intracellular Replication of Cryptococcus neoformans in Macrophages. Molecules 2023, 28, 6224. https://doi.org/10.3390/molecules28176224
Kietrungruang K, Sookkree S, Sangboonruang S, Semakul N, Poomanee W, Kitidee K, Tragoolpua Y, Tragoolpua K. Ethanolic Extract Propolis-Loaded Niosomes Diminish Phospholipase B1, Biofilm Formation, and Intracellular Replication of Cryptococcus neoformans in Macrophages. Molecules. 2023; 28(17):6224. https://doi.org/10.3390/molecules28176224
Chicago/Turabian StyleKietrungruang, Kritapat, Sanonthinee Sookkree, Sirikwan Sangboonruang, Natthawat Semakul, Worrapan Poomanee, Kuntida Kitidee, Yingmanee Tragoolpua, and Khajornsak Tragoolpua. 2023. "Ethanolic Extract Propolis-Loaded Niosomes Diminish Phospholipase B1, Biofilm Formation, and Intracellular Replication of Cryptococcus neoformans in Macrophages" Molecules 28, no. 17: 6224. https://doi.org/10.3390/molecules28176224
APA StyleKietrungruang, K., Sookkree, S., Sangboonruang, S., Semakul, N., Poomanee, W., Kitidee, K., Tragoolpua, Y., & Tragoolpua, K. (2023). Ethanolic Extract Propolis-Loaded Niosomes Diminish Phospholipase B1, Biofilm Formation, and Intracellular Replication of Cryptococcus neoformans in Macrophages. Molecules, 28(17), 6224. https://doi.org/10.3390/molecules28176224