
Spice-Derived Phenolic Compounds: Potential for Skin Cancer Prevention and Therapy
 , ,
, ,
Abstract
:1. Introduction
2. Skin Cancers
3. Phenolic Compounds
4. Overview of Selected Spices
4.1. Allspice
4.2. Alpinia galanga
4.3. Black Cumin
4.4. Black Pepper
4.5. Cinnamon
4.6. Coriander
4.7. Fenugreek
4.8. Ginger
4.9. Oregano
4.10. Nutmeg
4.11. Red Chili
4.12. Rosemary
4.13. Saffron
4.14. Sichuan Pepper
4.15. Star Anise
4.16. Sumac
4.17. Thyme
4.18. Turmeric
5. Effect of Spices-Derived Phenolic Compounds against Melanoma and Non-Melanoma Skin Cancer
5.1. Phenolic Acids
5.2. Flavonoids
5.3. Other Phenolic Compounds
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shabir Ahmad, R.; Imran, M.; Kamran Khan, M.; Haseeb Ahmad, M.; Sajid Arshad, M.; Ateeq, H.; Abdul Rahim, M. Introductory Chapter: Herbs and Spices—An Overview. In Herbs and Spices—New Processing Technologies; Shabir Ahmad, R., Ed.; IntechOpen Limited: London, UK, 2021; Volume 1, p. 276. [Google Scholar]
- Bi, X.; Lim, J.; Henry, C.J. Spices in the management of diabetes mellitus. Food Chem. 2017, 217, 281–293. [Google Scholar] [PubMed]
- Tapsell, L.C.; Hemphill, I.; Cobiac, L.; Sullivan, D.R.; Fenech, M.; Patch, C.S.; Roodenrys, S.; Keogh, J.B.; Clifton, P.M.; Williams, P.G.; et al. Health benefits of herbs and spices: The past, the present, the future. Med. J. Aust. 2006, 185, S1–S24. [Google Scholar] [CrossRef]
- Essa, M.M.; Akbar, M.; Guillemin, G. The Benefits of Natural Products for Neurodegenerative Diseases; Essa, M.M., Akbar, M., Guillemin, G., Eds.; Springer: Cham, Switzerland, 2016; Volume 12, p. 496. [Google Scholar]
- Singh, N.; Yadav, S.S. A review on health benefits of phenolics derived from dietary spices. Curr. Res. Food Sci. 2022, 5, 1508–1523. [Google Scholar] [CrossRef] [PubMed]
- Avila-Galvez, M.A.; Gimenez-Bastida, J.A.; Espin, J.C.; Gonzalez-Sarrias, A. Dietary Phenolics against Breast Cancer. A Critical Evidence-Based Review and Future Perspectives. Int. J. Mol. Sci. 2020, 21, 5718. [Google Scholar] [CrossRef] [PubMed]
- Muller, A.G.; Sarker, S.D.; Saleem, I.Y.; Hutcheon, G.A. Delivery of natural phenolic compounds for the potential treatment of lung cancer. DARU J. Pharm. Sci. 2019, 27, 433–449. [Google Scholar] [CrossRef] [PubMed]
- De, S.; Paul, S.; Manna, A.; Majumder, C.; Pal, K.; Casarcia, N.; Mondal, A.; Banerjee, S.; Nelson, V.K.; Ghosh, S.; et al. Phenolic Phytochemicals for Prevention and Treatment of Colorectal Cancer: A Critical Evaluation of In Vivo Studies. Cancers 2023, 15, 993. [Google Scholar]
- Kountouri, A.M.; Kaliora, A.C.; Koumbi, L.; Andrikopoulos, N.K. In-vitro gastric cancer prevention by a polyphenol-rich extract from olives through induction of apoptosis. Eur. J. Cancer Prev. 2009, 18, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Spagnuolo, C.; Flores, G.; Russo, G.L.; del Castillo, M.L.R. A Phenolic Extract Obtained from Methyl Jasmonate-Treated Strawberries Enhances Apoptosis in a Human Cervical Cancer Cell Line. Nutr. Cancer 2016, 68, 1140–1150. [Google Scholar] [CrossRef] [PubMed]
- Ji, X.; Usman, A.; Razalli, N.H.; Sambanthamurthi, R.; Gupta, S.V. Oil palm phenolics (OPP) inhibit pancreatic cancer cell proliferation via suppression of NF-kappaB pathway. Anticancer Res. 2015, 35, 97–106. [Google Scholar]
- Leiter, U.; Keim, U.; Garbe, C. Epidemiology of Skin Cancer: Update 2019. Adv. Exp. Med. Biol. 2020, 1268, 123–139. [Google Scholar]
- O’Neill, C.H.; Scoggins, C.R. Melanoma. J. Surg. Oncol. 2019, 120, 873–881. [Google Scholar] [CrossRef]
- Garbe, C.; Keim, U.; Gandini, S.; Amaral, T.; Katalinic, A.; Hollezcek, B.; Martus, P.; Flatz, L.; Leiter, U.; Whiteman, D. Epidemiology of cutaneous melanoma and keratinocyte cancer in white populations 1943–2036. Eur. J. Cancer 2021, 152, 18–25. [Google Scholar] [CrossRef]
- Krynitz, B.; Olsson, H.; Lundh Rozell, B.; Lindelof, B.; Edgren, G.; Smedby, K.E. Risk of basal cell carcinoma in Swedish organ transplant recipients: A population-based study. Br. J. Dermatol. 2016, 174, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Peris, K.; Fargnoli, M.C.; Garbe, C.; Kaufmann, R.; Bastholt, L.; Seguin, N.B.; Bataille, V.; Marmol, V.D.; Dummer, R.; Harwood, C.A.; et al. Diagnosis and treatment of basal cell carcinoma: European consensus–based interdisciplinary guidelines. Eur. J. Cancer 2019, 118, 10–34. [Google Scholar] [CrossRef] [PubMed]
- Tanese, K. Diagnosis and Management of Basal Cell Carcinoma. Curr. Treat. Options Oncol. 2019, 20, 13. [Google Scholar] [CrossRef]
- Roewert-Huber, J.; Stockfleth, E.; Kerl, H. Pathology and pathobiology of actinic (solar) keratosis—An update. Br. J. Dermatol. 2007, 157 (Suppl. S2), 18–20. [Google Scholar] [CrossRef]
- Eisen, D.B.; Asgari, M.M.; Bennett, D.D.; Connolly, S.M.; Dellavalle, R.P.; Freeman, E.E.; Goldenberg, G.; Leffell, D.J.; Peschin, S.; Sligh, J.E.; et al. Guidelines of care for the management of actinic keratosis. J. Am. Acad. Dermatol. 2021, 85, e209–e233. [Google Scholar] [CrossRef] [PubMed]
- Kallini, J.R.; Hamed, N.; Khachemoune, A. Squamous cell carcinoma of the skin: Epidemiology, classification, management, and novel trends. Int. J. Dermatol. 2015, 54, 130–140. [Google Scholar] [CrossRef]
- Mogensen, M.; Jemec, G.B.E. Diagnosis of Nonmelanoma Skin Cancer/Keratinocyte Carcinoma: A Review of Diagnostic Accuracy of Nonmelanoma Skin Cancer Diagnostic Tests and Technologies. Dermatol. Surg. 2007, 33, 1158–1174. [Google Scholar] [CrossRef]
- Waldman, A.; Schmults, C. Cutaneous Squamous Cell Carcinoma. Hematol. Oncol. Clin. N. Am. 2019, 33, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Neale, H.; Michelon, M.; Jacob, S.; Pinkston, M.; Ukaegbu, R.; Zamor, W.; Morrison, E.; Deng, A.; Levin, N.A. Topical 5% 5-fluorouracil versus procedural modalities for squamous cell carcinoma in situ and superficial basal cell carcinoma: A retrospective cohort analysis. J. Am. Acad. Dermatol. 2022, 87, 423–425. [Google Scholar] [CrossRef]
- Dirschka, T.; Bierhoff, E.; Pflugfelder, A.; Garbe, C. Topical 3.0% diclofenac in 2.5% hyaluronic acid gel induces regression of cancerous transformation in actinic keratoses. J. Eur. Acad. Dermatol. Venereol. 2010, 24, 258–263. [Google Scholar] [CrossRef] [PubMed]
- Piquero-Casals, J.; Morgado-Carrasco, D.; Gilaberte, Y.; Del Rio, R.; Macaya-Pascual, A.; Granger, C.; López-Estebaranz, J.L. Management Pearls on the Treatment of Actinic Keratoses and Field Cancerization. Dermatol. Ther. 2020, 10, 903–915. [Google Scholar] [CrossRef]
- Garbe, C.; Amaral, T.; Peris, K.; Hauschild, A.; Arenberger, P.; Basset-Seguin, N.; Bastholt, L.; Bataille, V.; del Marmol, V.; Dréno, B.; et al. European consensus-based interdisciplinary guideline for melanoma. Part 1: Diagnostics: Update 2022. Eur. J. Cancer 2022, 170, 236–255. [Google Scholar] [CrossRef] [PubMed]
- Erdei, E.; Torres, S.M. A new understanding in the epidemiology of melanoma. Expert Rev. Anticancer Ther. 2010, 10, 1811–1823. [Google Scholar] [CrossRef] [PubMed]
- Rastrelli, M.; Tropea, S.; Rossi, C.R.; Alaibac, M. Melanoma: Epidemiology, risk factors, pathogenesis, diagnosis and classification. In Vivo 2014, 28, 1005–1011. [Google Scholar]
- Leonardi, G.C.; Falzone, L.; Salemi, R.; Zanghi, A.; Spandidos, D.A.; McCubrey, J.A.; Candido, S.; Libra, M. Cutaneous melanoma: From pathogenesis to therapy (Review). Int. J. Oncol. 2018, 52, 1071–1080. [Google Scholar] [CrossRef]
- Rashid, S.; Shaughnessy, M.; Tsao, H. Melanoma classification and management in the era of molecular medicine. Dermatol. Clin. 2023, 41, 49–63. [Google Scholar] [CrossRef] [PubMed]
- Gong, H.Z.; Zheng, H.Y.; Li, J. Amelanotic melanoma. Melanoma Res 2019, 29, 221–230. [Google Scholar] [CrossRef]
- Garbe, C.; Peris, K.; Hauschild, A.; Saiag, P.; Middleton, M.; Bastholt, L.; Grob, J.J.; Malvehy, J.; Newton-Bishop, J.; Stratigos, A.J.; et al. Diagnosis and treatment of melanoma. European consensus-based interdisciplinary guideline—Update 2016. Eur. J. Cancer 2016, 63, 201–217. [Google Scholar] [CrossRef] [PubMed]
- Dummer, R.; Hauschild, A.; Lindenblatt, N.; Pentheroudakis, G.; Keilholz, U. Cutaneous melanoma: ESMO Clinical Practice Guidelines for diagnosis, treatment and follow-up. Ann. Oncol. 2015, 26 (Suppl. S5), v126-32. [Google Scholar] [CrossRef]
- Swetter, S.M.; Thompson, J.A.; Albertini, M.R.; Barker, C.A.; Baumgartner, J.; Boland, G.; Chmielowski, B.; DiMaio, D.; Durham, A.; Fields, R.C.; et al. NCCN Guidelines(R) Insights: Melanoma: Cutaneous, Version 2.2021. J. Natl. Compr. Cancer Netw. 2021, 19, 364–376. [Google Scholar] [CrossRef]
- Alu’datt, M.H.; Rababah, T.; Alhamad, M.N.; Al-Mahasneh, M.A.; Almajwal, A.; Gammoh, S.; Ereifej, K.; Johargy, A.; Alli, I. A review of phenolic compounds in oil-bearing plants: Distribution, identification and occurrence of phenolic compounds. Food Chem. 2017, 218, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Robbins, R.J. Phenolic Acids in Foods: An Overview of Analytical Methodology. J. Agric. Food Chem. 2003, 51, 2866–2887. [Google Scholar] [CrossRef]
- Zhang, Y.; Cai, P.; Cheng, G.; Zhang, Y. A Brief Review of Phenolic Compounds Identified from Plants: Their Extraction, Analysis, and Biological Activity. Nat. Prod. Commun. 2022, 17, 1934578X211069721. [Google Scholar] [CrossRef]
- Rodríguez De Luna, S.L.; Ramírez-Garza, R.E.; Serna-Saldívar, S.O. Environmentally friendly methods for flavonoid extraction from plant material: Impact of their operating conditions on yield and antioxidant properties. Sci. World J. 2020, 2020, 6792069. [Google Scholar] [CrossRef]
- Dias, M.C.; Pinto, D.C.G.A.; Silva, A.M.S. Plant Flavonoids: Chemical Characteristics and Biological Activity. Molecules 2021, 26, 5377. [Google Scholar] [CrossRef]
- Serrano, J.; Puupponen-Pimiä, R.; Dauer, A.; Aura, A.-M.; Saura-Calixto, F. Tannins: Current knowledge of food sources, intake, bioavailability and biological effects. Mol. Nutr. Food Res. 2009, 53 (Suppl. S2), S310–S329. [Google Scholar] [CrossRef]
- Pecyna, P.; Wargula, J.; Murias, M.; Kucinska, M. More than Resveratrol: New Insights into Stilbene-Based Compounds. Biomolecules 2020, 10, 1111. [Google Scholar] [CrossRef] [PubMed]
- Stringlis, I.A.; De Jonge, R.; Pieterse, C.M.J. The Age of Coumarins in Plant–Microbe Interactions. Plant Cell Physiol. 2019, 60, 1405–1419. [Google Scholar] [CrossRef] [PubMed]
- Saleem, M.; Kim, H.J.; Ali, M.S.; Lee, Y.S. An update on bioactive plant lignans. Nat. Prod. Rep. 2005, 22, 696–716. [Google Scholar] [CrossRef]
- Ginwala, R.; Bhavsar, R.; Chigbu, D.G.I.; Jain, P.; Khan, Z.K. Potential Role of Flavonoids in Treating Chronic Inflammatory Diseases with a Special Focus on the Anti-Inflammatory Activity of Apigenin. Antioxidants 2019, 8, 35. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Guan, S.; Lu, J.; Chen, Z.; Huang, G.; Li, G.; Xiong, Y.; Zhang, S.; Yue, Z.; Deng, X. Suppressive effects of fisetin on mice T lymphocytes in vitro and in vivo. J. Surg. Res. 2013, 185, 399–409. [Google Scholar] [CrossRef]
- Kamisah, Y.; Jalil, J.; Yunos, N.M.; Zainalabidin, S. Cardioprotective Properties of Kaempferol: A Review. Plants 2023, 12, 2096. [Google Scholar] [CrossRef] [PubMed]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Dourado, N.S.; Souza, C.d.S.; de Almeida, M.M.A.; da Silva, A.B.; dos Santos, B.L.; Silva, V.D.A.; De Assis, A.M.; da Silva, J.S.; Souza, D.O.; Costa, M.d.F.D.; et al. Neuroimmunomodulatory and Neuroprotective Effects of the Flavonoid Apigenin in in vitro Models of Neuroinflammation Associated With Alzheimer’s Disease. Front. Aging Neurosci. 2020, 12, 119. [Google Scholar] [CrossRef]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef]
- Fernández, J.; Silván, B.; Entrialgo-Cadierno, R.; Villar, C.J.; Capasso, R.; Uranga, J.A.; Lombó, F.; Abalo, R. Antiproliferative and palliative activity of flavonoids in colorectal cancer. BioMedicine 2021, 143, 112241. [Google Scholar] [CrossRef]
- Abdullah, H.; Ismail, I.; Suppian, R.; Zakaria, N.M. Natural Gallic Acid and Methyl Gallate Induces Apoptosis in Hela Cells through Regulation of Intrinsic and Extrinsic Protein Expression. Int. J. Mol. Sci. 2023, 24, 8495. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.-C.; Liu, Y.-N.; La, X.-Q.; Li, S.-T.; Wen, L.-N.; Liu, T.; Li, H.-Q.; Li, A.-P.; Wu, H.; Wu, C.-X.; et al. The bound polyphenols of foxtail millet (Setaria italica) inner shell inhibit breast cancer by promoting lipid accumulation-induced autophagic death. Food Chem. Toxicol. 2023, 177, 113855. [Google Scholar] [CrossRef]
- Lin, W.; Tongyi, S. Role of Bax/Bcl-2 family members in green tea polyphenol induced necroptosis of p53-deficient Hep3B cells. Tumor Biol. 2014, 35, 8065–8075. [Google Scholar] [CrossRef]
- Michalkova, R.; Kello, M.; Kudlickova, Z.; Gazdova, M.; Mirossay, L.; Mojzisova, G.; Mojzis, J. Programmed Cell Death Alterations Mediated by Synthetic Indole Chalcone Resulted in Cell Cycle Arrest, DNA Damage, Apoptosis and Signaling Pathway Modulations in Breast Cancer Model. Pharmaceutics 2022, 14, 503. [Google Scholar] [CrossRef] [PubMed]
- Barreca, M.M.; Alessandro, R.; Corrado, C. Effects of Flavonoids on Cancer, Cardiovascular and Neurodegenerative Diseases: Role of NF-kappaB Signaling Pathway. Int. J. Mol. Sci. 2023, 24, 9236. [Google Scholar] [CrossRef]
- Russo, M.; Moccia, S.; Luongo, D.; Russo, G.L. Senolytic Flavonoids Enhance Type-I and Type-II Cell Death in Human Radioresistant Colon Cancer Cells through AMPK/MAPK Pathway. Cancers 2023, 15, 2660. [Google Scholar] [CrossRef] [PubMed]
- Mirossay, L.; Varinská, L.; Mojžiš, J. Antiangiogenic Effect of Flavonoids and Chalcones: An Update. Int. J. Mol. Sci. 2017, 19, 27. [Google Scholar] [CrossRef] [PubMed]
- Varinska, L.; van Wijhe, M.; Belleri, M.; Mitola, S.; Perjesi, P.; Presta, M.; Koolwijk, P.; Ivanova, L.; Mojzis, J. Anti-angiogenic activity of the flavonoid precursor 4-hydroxychalcone. Eur. J. Pharmacol. 2012, 691, 125–133. [Google Scholar] [CrossRef]
- Khater, M.; Greco, F.; Osborn, H.M.I. Antiangiogenic Activity of Flavonoids: A Systematic Review and Meta-Analysis. Molecules 2020, 25, 4712. [Google Scholar] [CrossRef]
- Zhang, L.; Lokeshwar, B.L. Medicinal Properties of the Jamaican Pepper Plant Pimenta dioica and Allspice. Curr. Drug Targets 2012, 13, 1900–1906. [Google Scholar] [CrossRef] [PubMed]
- Al-Rehaily, A.J.; Al-Said, M.S.; Al-Yahya, M.A.; Mossa, J.S.; Rafatullah, S. Ethnopharmacological Studies on Allspice (Pimenta dioica) in Laboratory Animals. Pharm. Biol. 2002, 40, 200–205. [Google Scholar] [CrossRef]
- Miyajima, Y.; Kikuzaki, H.; Hisamoto, M.; Nakatani, N. Antioxidative polyphenols from berries of Pimenta dioica. BioFactors 2004, 22, 301–303. [Google Scholar] [CrossRef]
- El Gizawy, H.A.; Boshra, S.A.; Mostafa, A.; Mahmoud, S.H.; Ismail, M.I.; Alsfouk, A.A.; Taher, A.T.; Al-Karmalawy, A.A. Pimenta dioica (L.) Merr. Bioactive Constituents Exert Anti-SARS-CoV-2 and Anti-Inflammatory Activities: Molecular Docking and Dynamics, In Vitro, and In Vivo Studies. Molecules 2021, 26, 5844. [Google Scholar] [CrossRef]
- Marzouk, M.S.; Moharram, F.A.; Mohamed, M.A.; Gamal-Eldeen, A.M.; Aboutabl, E.A. Anticancer and antioxidant tannins from Pimenta dioica leaves. Z. Naturforsch. C J. Biosci. 2007, 62, 526–536. [Google Scholar] [CrossRef]
- Milenković, A.; Stanojević, J.; Stojanović-Radić, Z.; Pejčić, M.; Cvetković, D.; Zvezdanović, J.; Stanojević, L. Chemical composition, antioxidative and antimicrobial activity of allspice (Pimenta dioica (L.) Merr.) essential oil and extract. Adv. Technol. 2020, 9, 27–36. [Google Scholar] [CrossRef]
- Rao, P.S.; Navinchandra, S.; Jayaveera, K.N. An important spice, Pimenta dioica (Linn.) Merill: A Review. Int. Curr. Pharm. J. 2012, 1, 221–225. [Google Scholar] [CrossRef]
- Rema, J.; Krishnamoorthy, B. Handbook of Herbs and Spices, 2nd ed.; Woodhead Publishing: Cambridge, UK, 2012; pp. 166–192. [Google Scholar]
- Mandal, D.; Sarkar, T.; Chakraborty, R. Critical Review on Nutritional, Bioactive, and Medicinal Potential of Spices and Herbs and Their Application in Food Fortification and Nanotechnology. Appl. Biochem. Biotechnol. 2023, 195, 1319–1513. [Google Scholar] [CrossRef]
- Chudiwal, A.K.; Jain, D.P.; Rahul Somani, R. Alpinia galanga Willd—An overview on phyto-pharmacological properties. Indian J. Nat. Prod. Res. 2010, 1, 143–149. [Google Scholar]
- Eram, S.; Mujahid, M.; Bagga, P.; Ansari, V.; Ahmad, M.; Kumar, A.; Ahsan, F.; Akhter, M. A Review on Phytopharmacological Activity of Alpinia Galanga. Int. J. Pharm. Pharm. Sci. 2019, 11, 6–11. [Google Scholar] [CrossRef]
- Chouni, A.; Paul, S. A Review on Phytochemical and Pharmacological Potential of Alpinia galanga. Pharmacogn. J. 2018, 10, 9–15. [Google Scholar] [CrossRef]
- Namdeo, A.G.; Kale, V.M. Comparative pharmacognostic and phytochemical investigation of two Alpinia species from Zingiberaceae Family. World J. Pharm. Res. 2015, 4, 1417–1432. [Google Scholar]
- Tungmunnithum, D.; Tanaka, N.; Uehara, A.; Iwashina, T. Flavonoids Profile, Taxonomic Data, History of Cosmetic Uses, Anti-Oxidant and Anti-Aging Potential of Alpinia galanga (L.) Willd. Cosmetics 2020, 7, 89. [Google Scholar] [CrossRef]
- Ramanunny, A.K.; Wadhwa, S.; Gulati, M.; Vishwas, S.; Khursheed, R.; Paudel, K.R.; Gupta, S.; Porwal, O.; Alshahrani, S.M.; Jha, N.K.; et al. Journey of Alpinia galanga from kitchen spice to nutraceutical to folk medicine to nanomedicine. J. Ethnopharmacol. 2022, 291, 115144. [Google Scholar] [CrossRef]
- Zhou, Y.-Q.; Liu, H.; He, M.-X.; Wang, R.; Zeng, Q.-Q.; Wang, Y.; Ye, W.-C.; Zhang, Q.-W. Chapter 11—A Review of the Botany, Phytochemical, and Pharmacological Properties of Galangal. In Natural and Artificial Flavoring Agents and Food Dyes; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: San Diego, CA, USA, 2018; pp. 351–396. [Google Scholar]
- Nampoothiri, S.V.; Esakkidurai, T.; Pitchumani, K. Identification and Quantification of Phenolic Compounds in Alpinia galanga and Alpinia calcarata and its Relation to Free Radical Quenching Properties: A Comparative Study. J. Herbs Spices Med. Plants 2015, 21, 140–147. [Google Scholar] [CrossRef]
- Suja, S.; Chinnaswamy, P. Inhibition of in vitro cytotoxic effect evoked by Alpinia galanga and Alpinia officinarum on PC-3 cell line. Anc. Sci. Life 2008, 27, 33–40. [Google Scholar] [PubMed]
- Choi, J.-Y.; Lee, N.-K.; Wang, Y.-Y.; Hong, J.-P.; Son, S.R.; Gu, D.-H.; Jang, D.S.; Choi, J.-H. 1′-Acetoxyeugenol Acetate Isolated from Thai Ginger Induces Apoptosis in Human Ovarian Cancer Cells by ROS Production via NADPH Oxidase. Antioxidants 2022, 11, 293. [Google Scholar] [CrossRef] [PubMed]
- Suciati, A.; Maryati. Systematic Review: Anticancer Potential of Active Compounds from Galangal (Alpinia galanga). In Proceedings of the 4th International Conference Current Breakthrough in Pharmacy (ICB-Pharma 2022), Sukoharjo, Indonesia, 14–15 January 2022. [Google Scholar]
- Ali, B.H.; Blunden, G. Pharmacological and toxicological properties of Nigella sativa. Phytother. Res. 2003, 17, 299–305. [Google Scholar] [CrossRef]
- Kooti, W.; Hasanzadeh-Noohi, Z.; Sharafi-Ahvazi, N.; Asadi-Samani, M.; Ashtary-Larky, D. Phytochemistry, pharmacology, and therapeutic uses of black seed (Nigella sativa). Chin. J. Nat. Med. 2016, 14, 732–745. [Google Scholar] [CrossRef]
- Ismail, N.; Abd Ghafar, S.A.; Abu Bakar, M.Z. Chapter Fifteen—Antioxidant activity and phenolic content of black cumin seeds. In Biochemistry, Nutrition, and Therapeutics of Black Cumin Seed; Mariod, A.A., Ed.; Academic Press: San Diego, CA, USA, 2023; pp. 169–188. [Google Scholar]
- Srinivasan, K. Cumin (Cuminum cyminum) and black cumin (Nigella sativa) seeds: Traditional uses, chemical constituents, and nutraceutical effects. Food Qual. Saf. 2018, 2, 1–16. [Google Scholar] [CrossRef]
- Milenkovic, A.; Stanojević, L. Black pepper: Chemical composition and biological activities. Adv. Technol. 2021, 10, 40–50. [Google Scholar] [CrossRef]
- Al-Khayri, J.M.; Upadhya, V.; Pai, S.R.; Naik, P.M.; Al-Mssallem, M.Q.; Alessa, F.M. Comparative Quantification of the Phenolic Compounds, Piperine Content, and Total Polyphenols along with the Antioxidant Activities in the Piper trichostachyon and P. nigrum. Molecules 2022, 27, 5965. [Google Scholar] [CrossRef]
- Ashokkumar, K.; Murugan, M.; Dhanya, M.K.; Pandian, A.; Warkentin, T.D. Phytochemistry and therapeutic potential of black pepper [Piper nigrum (L.)] essential oil and piperine: A review. Clin. Phytosci. 2021, 7, 52. [Google Scholar] [CrossRef]
- Butt, M.S.; Pasha, I.; Sultan, M.T.; Randhawa, M.A.; Saeed, F.; Ahmed, W. Black Pepper and Health Claims: A Comprehensive Treatise. Crit. Rev. Food Sci. Nutr. 2013, 53, 875–886. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Qiu, B.; Sun, M.; Wang, Y.; Wei, M.; Gong, Y.; Yan, M. Preparation of Black pepper (Piper nigrum L.) essential oil nanoparticles and its antitumor activity on triple negative breast cancer in vitro. J. Food Biochem. 2022, 46, e14406. [Google Scholar] [CrossRef]
- Cardoso, L.P.; de Sousa, S.O.; Gusson-Zanetoni, J.P.; Silva, L.L.d.M.M.; Frigieri, B.M.; Henrique, T.; Tajara, E.H.; Oliani, S.M.; Rodrigues-Lisoni, F.C. Piperine Reduces Neoplastic Progression in Cervical Cancer Cells by Downregulating the Cyclooxygenase 2 Pathway. Pharmaceuticals 2023, 16, 103. [Google Scholar] [CrossRef] [PubMed]
- Manayi, A.; Nabavi, S.M.; Setzer, W.N.; Jafari, S. Piperine as a Potential Anti-cancer Agent: A Review on Preclinical Studies. Curr. Med. Chem. 2018, 25, 4918–4928. [Google Scholar] [CrossRef] [PubMed]
- Rao, P.V.; Gan, S.H. Cinnamon: A Multifaceted Medicinal Plant. Evid.-Based Complement. Altern. Med. 2014, 2014, 642942. [Google Scholar] [CrossRef]
- Sadeghi, S.; Davoodvandi, A.; Pourhanifeh, M.H.; Sharifi, N.; ArefNezhad, R.; Sahebnasagh, R.; Moghadam, S.A.; Sahebkar, A.; Mirzaei, H. Anti-cancer effects of cinnamon: Insights into its apoptosis effects. Eur. J. Med. Chem. 2019, 178, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Sahib, A.S. Anti-diabetic and antioxidant effect of cinnamon in poorly controlled type-2 diabetic Iraqi patients: A randomized, placebo-controlled clinical trial. J. Intercult. Ethnopharmacol. 2016, 5, 108–113. [Google Scholar] [CrossRef]
- Schink, A.; Naumoska, K.; Kitanovski, Z.; Kampf, C.J.; Fröhlich-Nowoisky, J.; Thines, E.; Pöschl, U.; Schuppan, D.; Lucas, K. Anti-inflammatory effects of cinnamon extract and identification of active compounds influencing the TLR2 and TLR4 signaling pathways. Food Funct. 2018, 9, 5950–5964. [Google Scholar] [CrossRef]
- Raeisi, M.; Tajik, H.; Yarahmadi, A.; Sanginabadi, S. Antimicrobial Effect of Cinnamon Essential Oil Against Escherichia Coli and Staphylococcus aureus. Health Scope 2015, 4, e21808. [Google Scholar] [CrossRef]
- Pagliari, S.; Forcella, M.; Lonati, E.; Sacco, G.; Romaniello, F.; Rovellini, P.; Fusi, P.; Palestini, P.; Campone, L.; Labra, M.; et al. Antioxidant and Anti-Inflammatory Effect of Cinnamon (Cinnamomum verum J. Presl) Bark Extract after In Vitro Digestion Simulation. Foods 2023, 12, 452. [Google Scholar] [CrossRef] [PubMed]
- Shahidi, F.; Hossain, A. Bioactives in spices, and spice oleoresins: Phytochemicals and their beneficial effects in food preservation and health promotion. J. Food Bioact. 2018, 3, 8–75. [Google Scholar] [CrossRef]
- Cardoso-Ugarte, G.A.; López-Malo, A.; Sosa-Morales, M.E. Chapter 38—Cinnamon (Cinnamomum zeylanicum) Essential Oils. In Essential Oils in Food Preservation, Flavor and Safety; Preedy, V.R., Ed.; Academic Press: San Diego, CA, USA, 2016; pp. 339–347. [Google Scholar]
- Vallverdú-Queralt, A.; Regueiro, J.; Martínez-Huélamo, M.; Alvarenga, J.F.R.; Leal, L.N.; Lamuela-Raventos, R.M. A comprehensive study on the phenolic profile of widely used culinary herbs and spices: Rosemary, thyme, oregano, cinnamon, cumin and bay. Food Chem. 2014, 154, 299–307. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.; Rao, A.S.; Nandal, A.; Kumar, S.; Yadav, S.S.; Ganaie, S.A.; Narasimhan, B. Phytochemical and pharmacological review of Cinnamomum verum J. Presl-a versatile spice used in food and nutrition. Food Chem. 2021, 338, 127773. [Google Scholar] [CrossRef]
- Baliga, M.S.; Mane, P.P.; Timothy Nallemgera, J.; Thilakchand, K.R.; Kalekhan, F. Chapter 5–Dietary Spices in the Prevention of Rheumatoid Arthritis: Past, Present, and Future. In Foods and Dietary Supplements in the Prevention and Treatment of Disease in Older Adults; Watson, R.R., Ed.; Academic Press: San Diego, CA, USA, 2015; pp. 41–49. [Google Scholar]
- Kumar, V.; Marković, T.; Mila, E.; Dey, A. Herbs: Composition and Dietary Importance. In Encyclopedia of Food and Health; Caballero, B., Finglas, P.M., Toldrá, F., Eds.; Elsevier: Kidlington, UK, 2015; Volume 3, pp. 332–337. [Google Scholar]
- Laribi, B.; Kouki, K.; M’Hamdi, M.; Bettaieb, T. Coriander (Coriandrum sativum L.) and its bioactive constituents. Fitoterapia 2015, 103, 9–26. [Google Scholar] [CrossRef] [PubMed]
- Msaada, K.; Ben Jemia, M.; Salem, N.; Bachrouch, O.; Sriti, J.; Tammar, S.; Bettaieb, I.; Jabri, I.; Kefi, S.; Limam, F.; et al. Antioxidant activity of methanolic extracts from three coriander (Coriandrum sativum L.) fruit varieties. Arab. J. Chem. 2017, 10, S3176–S3183. [Google Scholar] [CrossRef]
- Barros, L.; Dueñas, M.; Dias, M.I.; Sousa, M.J.; Santos-Buelga, C.; Ferreira, I.C. Phenolic profiles of in vivo and in vitro grown Coriandrum sativum L. Food Chem. 2012, 132, 841–848. [Google Scholar] [CrossRef]
- Oganesyan, E.T.; Nersesyan, Z.M.; Parkhomenko, A.Y. Chemical composition of the above-ground part of Coriandrum sativum. Pharm. Chem. J. 2007, 41, 149–153. [Google Scholar] [CrossRef]
- Nambiar, V.; Daniel, M.; Guin, P. Characterization of polyphenols from coriander leaves (Coriandrum sativum), red amaranthus (A paniculatus) and green amaranthus (A frumentaceus) using paper chromatography and their health implications. J. Herb Med. Toxicol. 2010, 4, 173–177. [Google Scholar]
- Mechchate, H.; de Oliveira, R.C.; Es-Safi, I.; Mourão, E.M.V.; Bouhrim, M.; Kyrylchuk, A.; Pontes, G.S.; Bousta, D.; Grafov, A. Antileukemic Activity and Molecular Docking Study of a Polyphenolic Extract from Coriander Seeds. Pharmaceuticals 2021, 14, 770. [Google Scholar] [CrossRef]
- Elmas, L.; Secme, M.; Mammadov, R.; Fahrioglu, U.; Dodurga, Y. The determination of the potential anticancer effects of Coriandrum sativum in PC-3 and LNCaP prostate cancer cell lines. J. Cell Biochem. 2019, 120, 3506–3513. [Google Scholar] [CrossRef]
- Eroğlu, C.; Seçme, M.; Bağcı, G.; Dodurga, Y. Assessment of the anticancer mechanism of ferulic acid via cell cycle and apoptotic pathways in human prostate cancer cell lines. Tumor Biol. 2015, 36, 9437–9446. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.J.; Pham, T.H.; Bak, Y.; Ryu, H.W.; Oh, S.R.; Yoon, D.Y. Orientin inhibits invasion by suppressing MMP-9 and IL-8 expression via the PKCalpha/ERK/AP-1/STAT3-mediated signaling pathways in TPA-treated MCF-7 breast cancer cells. Phytomedicine 2018, 50, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Nakamura, T.; Yasuzawa, T.; Ueshima, S. Effects of Coriandrum sativum on Migration and Invasion Abilities of Cancer Cells. J. Nutr. Sci. Vitaminol. 2020, 66, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.A.; Kumar, P. Fenugreek: A review on its nutraceutical properties and utilization in various food products. J. Saudi Soc. Agric. Sci. 2018, 17, 97–106. [Google Scholar] [CrossRef]
- Sun, W.; Shahrajabian, M.H.; Cheng, Q. Fenugreek Cultivation with Emphasis on Historical Aspects and its uses in Traditional Medicine and Modern Pharmaceutical Science. Mini-Rev. Med. Chem. 2021, 21, 724–730. [Google Scholar] [CrossRef]
- Visuvanathan, T.; Than, L.T.L.; Stanslas, J.; Chew, S.Y.; Vellasamy, S. Revisiting Trigonella foenum-graecum L.: Pharmacology and Therapeutic Potentialities. Plants 2022, 11, 1450. [Google Scholar] [CrossRef] [PubMed]
- Bahmani, M.; Shirzad, H.; Mirhosseini, M.; Mesripour, A.; Rafieian-Kopaei, M. A Review on Ethnobotanical and Therapeutic Uses of Fenugreek (Trigonella foenum-graceum L). J. Evid.-Based Complement. Altern. Med. 2015, 21, 53–62. [Google Scholar] [CrossRef]
- Nagulapalli Venkata, K.C.; Swaroop, A.; Bagchi, D.; Bishayee, A. A small plant with big benefits: Fenugreek (Trigonella foenum-graecum Linn.) for disease prevention and health promotion. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Skaltsa, H. Fenugreek—The Genus Trigonella, 1st ed.; CRC Press: London, UK, 2002; p. 226. [Google Scholar]
- Benayad, Z.; Gomez-Cordoves, C.; Es-Safi, N.E. Characterization of flavonoid glycosides from fenugreek (Trigonella foenum-graecum) crude seeds by HPLC-DAD-ESI/MS analysis. Int. J. Mol. Sci. 2014, 15, 20668–20685. [Google Scholar] [CrossRef]
- Niknam, R.; Kiani, H.; Mousavi, Z.E.; Mousavi, M. Extraction, Detection, and Characterization of Various Chemical Components of Trigonella foenum-graecum L. (Fenugreek) Known as a Valuable Seed in Agriculture. Fenugreek 2021, 189–217. [Google Scholar] [CrossRef]
- Shawky, E.; Sobhy, A.A.; Ghareeb, D.A.; Shams Eldin, S.M.; Selim, D.A. Comparative metabolomics analysis of bioactive constituents of the leaves of different Trigonella species: Correlation study to α-amylase and α-glycosidase inhibitory effects. Ind. Crops Prod. 2022, 182, 114947. [Google Scholar] [CrossRef]
- Al-Timimi, L.A.N. Antibacterial and Anticancer Activities of Fenugreek Seed Extract. Asian Pac. J. Cancer Prev. 2019, 20, 3771–3776. [Google Scholar] [CrossRef] [PubMed]
- Alsemari, A.; Alkhodairy, F.; Aldakan, A.; Al-Mohanna, M.; Bahoush, E.; Shinwari, Z.; Alaiya, A. The selective cytotoxic anti-cancer properties and proteomic analysis of Trigonella Foenum-Graecum. BMC Complement Altern. Med. 2014, 14, 114. [Google Scholar] [CrossRef] [PubMed]
- Ajmal, S.; Shafqat, M.; Ajmal, L.; Younas, H.; Tasadduq, R.; Mahmood, N. Evaluation of Anti-cancer and Anti-proliferative Activity of Medicinal Plant Extracts (Saffron, Green Tea, Clove, Fenugreek) on Toll Like Receptors Pathway. Nat. Prod. Sci. 2022, 28, 121–129. [Google Scholar] [CrossRef]
- A Alrumaihi, F.; A Khan, M.; Allemailem, K.S.; A Alsahli, M.; Almatroudi, A.; Younus, H.; A Alsuhaibani, S.; Algahtani, M.; Khan, A. Methanolic Fenugreek Seed Extract Induces p53-Dependent Mitotic Catastrophe in Breast Cancer Cells, Leading to Apoptosis. J. Inflamm. Res. 2021, 14, 1511–1535. [Google Scholar] [CrossRef]
- Singletary, K. Ginger: An Overview of Health Benefits. Nutr. Today 2010, 45, 171–183. [Google Scholar] [CrossRef]
- Khodaie, L.; Sadeghpoor, O. Ginger From Ancient Times to the New Outlook. Jundishapur J. Nat. Pharm. Prod. 2015, 10, e18402. [Google Scholar] [CrossRef]
- Kiyama, R. Nutritional implications of ginger: Chemistry, biological activities and signaling pathways. J. Nutr. Biochem. 2020, 86, 108486. [Google Scholar] [CrossRef]
- Semwal, R.B.; Semwal, D.K.; Combrinck, S.; Viljoen, A.M. Gingerols and shogaols: Important nutraceutical principles from ginger. Phytochemistry 2015, 117, 554–568. [Google Scholar] [CrossRef] [PubMed]
- Schadich, E.; Hlaváč, J.; Volná, T.; Varanasi, L.; Hajdúch, M.; Džubák, P. Effects of Ginger Phenylpropanoids and Quercetin on Nrf2-ARE Pathway in Human BJ Fibroblasts and HaCaT Keratinocytes. BioMed Res. Int. 2016, 2016, 2173275. [Google Scholar] [CrossRef]
- Ozkur, M.; Benlier, N.; Takan, I.; Vasileiou, C.; Georgakilas, A.G.; Pavlopoulou, A.; Cetin, Z.; Saygili, E.I. Ginger for Healthy Ageing: A Systematic Review on Current Evidence of Its Antioxidant, Anti-Inflammatory, and Anticancer Properties. Oxidative Med. Cell. Longev. 2022, 2022, 4748447. [Google Scholar] [CrossRef]
- Kundu, J.K.; Na, H.K.; Surh, Y.J. Ginger-derived phenolic substances with cancer preventive and therapeutic potential. Forum Nutr. 2009, 61, 182–192. [Google Scholar] [PubMed]
- Bae, W.Y.; Choi, J.S.; Kim, J.E.; Park, C.; Jeong, J.W. Zingerone suppresses angiogenesis via inhibition of matrix metalloproteinases during tumor development. Oncotarget 2016, 7, 47232–47241. [Google Scholar] [CrossRef]
- Gîrd, C.E.; Costea, T.; Mitran, V. Evaluation of cytotoxic activity and anticancer potential of indigenous Rosemary (Rosmarinus officinalis L.) and Oregano (Origanum vulgare L.) dry extracts on MG-63 bone osteosarcoma human cell line. Romanian J. Morphol. Embryol. 2021, 62, 525–535. [Google Scholar] [CrossRef]
- Bouyahya, A.; Chamkhi, I.; Benali, T.; Guaouguaou, F.-E.; Balahbib, A.; El Omari, N.; Taha, D.; Belmehdi, O.; Ghokhan, Z.; El Menyiy, N. Traditional use, phytochemistry, toxicology, and pharmacology of Origanum majorana L. J. Ethnopharmacol. 2021, 265, 113318. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, G.; Baydar, H.; Erbas, S. The influence of harvest time on essential oil composition, phenolic constituents and antioxidant properties of Turkish oregano (Origanum onites L.). J. Sci. Food Agric. 2010, 90, 205–209. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, B.; Marques, A.; Ramos, C.; Serrano, C.; Matos, O.; Neng, N.R.; Nogueira, J.M.F.; Saraiva, J.A.; Nunes, M.L. Chemical composition and bioactivity of different oregano (Origanum vulgare) extracts and essential oil. J. Sci. Food Agric. 2013, 93, 2707–2714. [Google Scholar] [CrossRef]
- Latha, P.G.; Sindhu, P.G.; Suja, S.R.; Geetha, B.S.; Pushpangadan, P.; Rajasekharan, S. Pharmacology and chemistry of Myristica fragrans Houtt.—A review. J. Spices Aromat. Crops 2005, 14, 94–101. [Google Scholar]
- Kuete, V. Chapter 23—Myristica fragrans: A Review. In Medicinal Spices and Vegetables from Africa; Kuete, V., Ed.; Academic Press: San Diego, CA, USA, 2017; pp. 497–512. [Google Scholar]
- Zhang, W.K.; Tao, S.S.; Li, T.T.; Li, Y.S.; Li, X.J.; Tang, H.B.; Cong, R.H.; Ma, F.L.; Wan, C.J. Nutmeg oil alleviates chronic inflammatory pain through inhibition of COX-2 expression and substance P release in vivo. Food Nutr. Res. 2016, 60, 30849. [Google Scholar] [CrossRef]
- Hayfaa, A.A.; Sahar, A.M.; Awatif, M.A. Evaluation of analgesic activity and toxicity of alkaloids in Myristica fragrans seeds in mice. J. Pain Res. 2013, 6, 611–615. [Google Scholar] [CrossRef] [PubMed]
- Shafiei, Z.; Shuhairi, N.N.; Md Fazly Shah Yap, N.; Harry Sibungkil, C.A.; Latip, J. Antibacterial Activity of Myristica fragrans against Oral Pathogens. Evid. Based Complement. Altern. Med. 2012, 2012, 825362. [Google Scholar] [CrossRef]
- Narasimhan, B.; Dhake, A.S. Antibacterial principles from Myristica fragrans seeds. J. Med. Food. 2006, 9, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Morita, T.; Jinno, K.; Kawagishi, H.; Arimoto, Y.; Suganuma, H.; Inakuma, T.; Sugiyama, K. Hepatoprotective Effect of Myristicin from Nutmeg (Myristica fragrans) on Lipopolysaccharide/d-Galactosamine-Induced Liver Injury. J. Agric. Food Chem. 2003, 51, 1560–1565. [Google Scholar] [CrossRef]
- Rezende, D.; Oliveira, C.D.; Batista, L.R.; Ferreira, V.R.F.; Brandao, R.M.; Caetano, A.R.S.; Alves, M.V.P.; Cardoso, M.G. Bactericidal and antioxidant effects of essential oils from Satureja montana L., Myristica fragrans H. and Cymbopogon flexuosus. Lett. Appl. Microbiol. 2022, 74, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Bandyopadhyay, P.S.V.C. Estimation of phenolic acids in cinnamon, clove, cardamom, nutmeg and mace by high performance liquid chromatography. J. Spices Aromat. 1995, 4, 129–134. [Google Scholar]
- Odubanjo, V.O.; Olasehinde, T.A.; Oyeleye, S.I.; Oboh, G.; Boligon, A.A. Seed extracts from Myristica fragrans (Nutmeg) and Moringa oleifera (Drumstick tree) inhibits enzymes relevant to erectile dysfunction and metal-induced oxidative damage in rats’ penile tissues. J. Food Biochem. 2017, 42, e12452. [Google Scholar] [CrossRef]
- Kraft, K.H.; Brown, C.H.; Nabhan, G.P.; Luedeling, E.; Luna Ruiz Jde, J.; Coppens d’Eeckenbrugge, G.; Hijmans, R.J.; Gepts, P. Multiple lines of evidence for the origin of domesticated chili pepper, Capsicum annuum, in Mexico. Proc. Natl. Acad. Sci. USA 2014, 111, 6165–6170. [Google Scholar] [CrossRef]
- Reyes-Escogido Mde, L.; Gonzalez-Mondragon, E.G.; Vazquez-Tzompantzi, E. Chemical and pharmacological aspects of capsaicin. Molecules 2011, 16, 1253–1270. [Google Scholar] [CrossRef]
- Pugliese, A.; O’Callaghan, Y.; Tundis, R.; Galvin, K.; Menichini, F.; O’Brien, N.; Loizzo, M.R. In vitro investigation of the bioaccessibility of carotenoids from raw, frozen and boiled red chili peppers (Capsicum annuum). Eur. J. Nutr. 2014, 53, 501–510. [Google Scholar] [CrossRef]
- Antonio, A.S.; Wiedemann, L.S.M.; Veiga Junior, V.F. The genus Capsicum: A phytochemical review of bioactive secondary metabolites. RSC Adv. 2018, 8, 25767–25784. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Sun, X.; Battino, M.; Wei, X.; Shi, J.; Zhao, L.; Liu, S.; Xiao, J.; Shi, B.; Zou, X. A comparative overview on chili pepper (capsicum genus) and sichuan pepper (zanthoxylum genus): From pungent spices to pharma-foods. Trends Food Sci. 2021, 117, 148–162. [Google Scholar] [CrossRef]
- Ayob, O.; Hussain, P.R.; Suradkar, P.; Naqash, F.; Rather, S.A.; Joshi, S.; Ahmad Azad, Z.R.A. Evaluation of chemical composition and antioxidant activity of Himalayan Red chilli varieties. LWT 2021, 146, 111413. [Google Scholar] [CrossRef]
- Batiha, G.E.; Alqahtani, A.; Ojo, O.A.; Shaheen, H.M.; Wasef, L.; Elzeiny, M.; Ismail, M.; Shalaby, M.; Murata, T.; Zaragoza-Bastida, A.; et al. Biological Properties, Bioactive Constituents, and Pharmacokinetics of Some Capsicum spp. and Capsaicinoids. Int. J. Mol. Sci. 2020, 21, 5179. [Google Scholar] [CrossRef] [PubMed]
- Clark, R.; Lee, S.H. Anticancer Properties of Capsaicin Against Human Cancer. Anticancer Res. 2016, 36, 837–843. [Google Scholar] [PubMed]
- Heinrich, M.; Kufer, J.; Leonti, M.; Pardo-de-Santayana, M. Ethnobotany and ethnopharmacology--interdisciplinary links with the historical sciences. J. Ethnopharmacol. 2006, 107, 157–160. [Google Scholar] [CrossRef]
- de Oliveira, J.R.; Camargo, S.E.A.; de Oliveira, L.D. Rosmarinus officinalis L. (rosemary) as therapeutic and prophylactic agent. J. Biomed. Sci. 2019, 26, 5. [Google Scholar] [CrossRef] [PubMed]
- Hussain, A.I.; Anwar, F.; Chatha, S.A.; Jabbar, A.; Mahboob, S.; Nigam, P.S. Rosmarinus officinalis essential oil: Antiproliferative, antioxidant and antibacterial activities. Braz. J. Microbiol. 2010, 41, 1070–1078. [Google Scholar] [CrossRef] [PubMed]
- Hcini, K.; Abidi, M.; Quílez, M.; Jordán Maria, J.; Sadok, B. Total Phenolic Content and Polyphenolic Profile of Tunisian Rosemary Rosmarinus officinalis L.) Residues. In Natural Drugs from Plants; Hany, A.E.-S., Ed.; IntechOpen: Rijeka, Croatia, 2021. [Google Scholar]
- Moore, J.; Yousef, M.; Tsiani, E. Anticancer Effects of Rosemary (Rosmarinus officinalis L.) Extract and Rosemary Extract Polyphenols. Nutrients 2016, 8, 731. [Google Scholar] [CrossRef]
- Gonzalez-Vallinas, M.; Reglero, G.; Ramirez de Molina, A. Rosemary (Rosmarinus officinalis L.) Extract as a Potential Complementary Agent in Anticancer Therapy. Nutr. Cancer 2015, 67, 1221–1229. [Google Scholar] [CrossRef]
- Saxena, R.B. Botany, Taxonomy and Cytology of Crocus sativus series. Ayu 2010, 31, 374–381. [Google Scholar] [CrossRef]
- Melnyk, J.P.; Wang, S.; Marcone, M.F. Chemical and biological properties of the world’s most expensive spice: Saffron. Food Res. Int. 2010, 43, 1981–1989. [Google Scholar] [CrossRef]
- Xing, B.; Li, S.; Yang, J.; Lin, D.; Feng, Y.; Lu, J.; Shao, Q. Phytochemistry, pharmacology, and potential clinical applications of saffron: A review. J. Ethnopharmacol. 2021, 281, 114555. [Google Scholar] [CrossRef]
- Groppo, M.; Pirani, J.R. A new species of Zanthoxylum (Rutaceae) with a key to the species from Northeastern Brazil. Phytotaxa 2017, 314, 259. [Google Scholar] [CrossRef]
- Okagu, I.U.; Ndefo, J.C.; Aham, E.C.; Udenigwe, C.C. Zanthoxylum Species: A Comprehensive Review of Traditional Uses, Phytochemistry, Pharmacological and Nutraceutical Applications. Molecules 2021, 26, 4023. [Google Scholar] [CrossRef]
- Ekka, G.; Jadhav, S.; Quraishi, A. An Overview of Genus Zanthoxylum with Special Reference to Its Herbal Significance and Application. Herbs Spices 2020. [Google Scholar] [CrossRef]
- Tanoh, E.A.; Boue, G.B.; Nea, F.; Genva, M.; Wognin, E.L.; Ledoux, A.; Martin, H.; Tonzibo, Z.F.; Frederich, M.; Fauconnier, M.L. Seasonal Effect on the Chemical Composition, Insecticidal Properties and Other Biological Activities of Zanthoxylum leprieurii Guill. & Perr. Essential oils. Foods 2020, 9, 550. [Google Scholar] [PubMed]
- Ji, Y.; Li, S.; Ho, C.-T. Chemical composition, sensory properties and application of Sichuan pepper (Zanthoxylum genus). Food Sci. Hum. Wellness 2019, 8, 115–125. [Google Scholar] [CrossRef]
- Wang, G.W.; Hu, W.T.; Huang, B.K.; Qin, L.P. Illicium verum: A review on its botany, traditional use, chemistry and pharmacology. J. Ethnopharmacol. 2011, 136, 10–20. [Google Scholar] [CrossRef]
- Sharafan, M.; Jafernik, K.; Ekiert, H.; Kubica, P.; Kocjan, R.; Blicharska, E.; Szopa, A. Illicium verum (Star Anise) and Trans-Anethole as Valuable Raw Materials for Medicinal and Cosmetic Applications. Molecules 2022, 27, 650. [Google Scholar] [CrossRef]
- Ghosh, S.; Chisti, Y.; Banerjee, U.C. Production of shikimic acid. Biotechnol. Adv. 2012, 30, 1425–1431. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Wu, Z.; Xia, Y.; Tan, J.; Zhao, H.; Chen, S.; Li, Y.; Tang, H.; Wang, G.; Zhang, Y. Antiviral and Antioxidant Components from the Fruits of Illicium verum Hook.f. (Chinese Star Anise). J. Agric. Food Chem. 2022, 70, 3697–3707. [Google Scholar] [CrossRef] [PubMed]
- De, M.; De, A.K.; Sen, P.; Banerjee, A.B. Antimicrobial properties of star anise (Illicium verum Hook f). Phytother. Res. 2002, 16, 94–95. [Google Scholar] [CrossRef] [PubMed]
- Majali, I.S. Antioxidant and Anti-Inflammatory Activity of Star Anise (Illicium verum) in Murine Model. Biomed. Pharmacol. J. 2022, 15, 1097–1108. [Google Scholar] [CrossRef]
- Ibrahim, F.; Ibrahim, A.; EL Gohary, A.E.; Hussein, M.; Ahmed, K. Illicium verum Extracts Anti-Gastro Ulcerogenic Potential on Experimentally Rat Models. Int. J. PharmTech Res. 2016, 9, 65–80. [Google Scholar]
- Patra, J.K.; Das, G.; Bose, S.; Banerjee, S.; Vishnuprasad, C.N.; Del Pilar Rodriguez-Torres, M.; Shin, H.S. Star anise (Illicium verum): Chemical compounds, antiviral properties, and clinical relevance. Phytother. Res. 2020, 34, 1248–1267. [Google Scholar] [CrossRef] [PubMed]
- Asif, M.; Yehya, A.H.S.; Al-Mansoub, M.A.; Revadigar, V.; Ezzat, M.O.; Khadeer Ahamed, M.B.; Oon, C.E.; Murugaiyah, V.; Abdul Majid, A.S.; Abdul Majid, A.M.S. Anticancer attributes of Illicium verum essential oils against colon cancer. S. Afr. J. Bot. 2016, 103, 156–161. [Google Scholar] [CrossRef]
- Pahore, A.K.; Khan, S.; Karim, N. Anticancer effect of Illicium verum (star anise fruit) against human breast cancer MCF-7 cell line. Pak. J. Med. Sci. 2023, 39, 70–74. [Google Scholar] [CrossRef]
- Chen, C.H.; deGraffenried, L.A. Anethole suppressed cell survival and induced apoptosis in human breast cancer cells independent of estrogen receptor status. Phytomedicine 2012, 19, 763–767. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.; Shamim, S. Anisi Stellati Fructus, a Significant Traditional Chinese Medicine (TCM) Herb and Its Bioactivity against Gastric Cancer. Evid. Based Complement. Altern. Med. 2022, 2022, 4071489. [Google Scholar] [CrossRef] [PubMed]
- Najar, B.; Shortrede, J.E.; Pistelli, L.; Buhagiar, J. Chemical Composition and in Vitro Cytotoxic Screening of Sixteen Commercial Essential Oils on Five Cancer Cell Lines. Chem. Biodivers. 2020, 17, e1900478. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.; Im, M.; Ma, J.Y. Anisi stellati fructus extract attenuates the in vitro and in vivo metastatic and angiogenic potential of malignant cancer cells by downregulating proteolytic activity and pro-angiogenic factors. Int. J. Oncol. 2014, 45, 1937–1948. [Google Scholar] [CrossRef]
- Batiha, G.E.; Ogunyemi, O.M.; Shaheen, H.M.; Kutu, F.R.; Olaiya, C.O.; Sabatier, J.M.; De Waard, M. Rhus coriaria L. (Sumac), a Versatile and Resourceful Food Spice with Cornucopia of Polyphenols. Molecules 2022, 27, 5179. [Google Scholar] [CrossRef]
- Alsamri, H.; Athamneh, K.; Pintus, G.; Eid, A.H.; Iratni, R. Pharmacological and Antioxidant Activities of Rhus coriaria L. (Sumac). Antioxidants 2021, 10, 73. [Google Scholar] [CrossRef]
- Sakhr, K.; El Khatib, S. Physiochemical properties and medicinal, nutritional and industrial applications of Lebanese Sumac (Syrian Sumac-Rhus coriaria): A review. Heliyon 2020, 6, e03207. [Google Scholar] [CrossRef]
- Kubatka, P.; Kello, M.; Kajo, K.; Samec, M.; Liskova, A.; Jasek, K.; Koklesova, L.; Kuruc, T.; Adamkov, M.; Smejkal, K.; et al. Rhus coriaria L. (Sumac) Demonstrates Oncostatic Activity in the Therapeutic and Preventive Model of Breast Carcinoma. Int. J. Mol. Sci. 2020, 22, 183. [Google Scholar] [CrossRef] [PubMed]
- Athamneh, K.; Hasasna, H.E.; Samri, H.A.; Attoub, S.; Arafat, K.; Benhalilou, N.; Rashedi, A.A.; Dhaheri, Y.A.; AbuQamar, S.; Eid, A.; et al. Rhus coriaria increases protein ubiquitination, proteasomal degradation and triggers non-canonical Beclin-1-independent autophagy and apoptotic cell death in colon cancer cells. Sci. Rep. 2017, 7, 11633. [Google Scholar] [CrossRef] [PubMed]
- Gabr, S.A.; Alghadir, H.A. Potential anticancer activities of Rhus coriaria (sumac) extract against human cancer cell lines. Biosci. Rep. 2021, 41, BSR20204384. [Google Scholar] [CrossRef] [PubMed]
- Romeo, F.V.; Ballistreri, G.; Fabroni, S.; Pangallo, S.; Nicosia, M.G.; Schena, L.; Rapisarda, P. Chemical Characterization of Different Sumac and Pomegranate Extracts Effective against Botrytis cinerea Rots. Molecules 2015, 20, 11941–11958. [Google Scholar] [CrossRef]
- Bozan, B.; Koşar, M.; ZTunalier Öztürk, N.; Baser, K.H.C. Antioxidant and Free Radical Scavenging Activities of Rhus coriaria and Cinnamomum cassia Extracts. Acta. Aliment. 2003, 32, 53–61. [Google Scholar] [CrossRef]
- Abu-Reidah, I.M.; Ali-Shtayeh, M.S.; Jamous, R.M.; Arraez-Roman, D.; Segura-Carretero, A. HPLC-DAD-ESI-MS/MS screening of bioactive components from Rhus coriaria L. (Sumac) fruits. Food Chem. 2015, 166, 179–191. [Google Scholar] [CrossRef]
- Patil, S.M.; Ramu, R.; Shirahatti, P.S.; Shivamallu, C.; Amachawadi, R.G. A systematic review on ethnopharmacology, phytochemistry and pharmacological aspects of Thymus vulgaris Linn. Heliyon 2021, 7, e07054. [Google Scholar] [CrossRef]
- Hammoudi Halat, D.; Krayem, M.; Khaled, S.; Younes, S. A Focused Insight into Thyme: Biological, Chemical, and Therapeutic Properties of an Indigenous Mediterranean Herb. Nutrients 2022, 14, 2104. [Google Scholar] [CrossRef]
- Ahmad, A.; Saeed, M.; Ansari, I.A. Molecular insights on chemopreventive and anticancer potential of carvacrol: Implications from solid carcinomas. J. Food Biochem. 2021, 45, e14010. [Google Scholar] [CrossRef] [PubMed]
- Niksic, H.; Becic, F.; Koric, E.; Gusic, I.; Omeragic, E.; Muratovic, S.; Miladinovic, B.; Duric, K. Cytotoxicity screening of Thymus vulgaris L. essential oil in brine shrimp nauplii and cancer cell lines. Sci. Rep. 2021, 11, 13178. [Google Scholar] [CrossRef]
- Kowalczyk, A.; Przychodna, M.; Sopata, S.; Bodalska, A.; Fecka, I. Thymol and Thyme Essential Oil-New Insights into Selected Therapeutic Applications. Molecules 2020, 25, 4125. [Google Scholar] [CrossRef] [PubMed]
- Roby, M.H.H.; Sarhan, M.A.; Selim, K.A.-H.; Khalel, K.I. Evaluation of antioxidant activity, total phenols and phenolic compounds in thyme (Thymus vulgaris L.), sage (Salvia officinalis L.), and marjoram (Origanum majorana L.) extracts. Ind. Crops Prod. 2013, 43, 827–831. [Google Scholar] [CrossRef]
- Teixeira, B.; Marques, A.; Ramos, C.; Neng, N.R.; Nogueira, J.M.F.; Saraiva, J.A.; Nunes, M.L. Chemical composition and antibacterial and antioxidant properties of commercial essential oils. Ind. Crops Prod. 2013, 43, 587–595. [Google Scholar] [CrossRef]
- Kotha, R.R.; Luthria, D.L. Curcumin: Biological, Pharmaceutical, Nutraceutical, and Analytical Aspects. Molecules 2019, 24, 2930. [Google Scholar] [CrossRef] [PubMed]
- Meng, F.-C.; Zhou, Y.-Q.; Ren, D.; Wang, R.; Wang, C.; Lin, L.-G.; Zhang, X.-Q.; Ye, W.-C.; Zhang, Q.-W. Chapter 10—Turmeric: A Review of Its Chemical Composition, Quality Control, Bioactivity, and Pharmaceutical Application. In Natural and Artificial Flavoring Agents and Food Dyes; Grumezescu, A.M., Holban, A.M., Eds.; Academic Press: San Diego, CA, USA, 2018; pp. 299–350. [Google Scholar]
- Yang, Q.-Q.; Cheng, L.-Z.; Zhang, T.; Yaron, S.; Jiang, H.-X.; Sui, Z.-Q.; Corke, H. Phenolic profiles, antioxidant, and antiproliferative activities of turmeric (Curcuma longa). Ind. Crops Prod. 2020, 152, 112561. [Google Scholar] [CrossRef]
- Fan, Y.; Zhang, X.; Tong, Y.; Chen, S.; Liang, J. Curcumin against gastrointestinal cancer: A review of the pharmacological mechanisms underlying its antitumor activity. Front. Pharmacol. 2022, 13, 990475. [Google Scholar] [CrossRef] [PubMed]
- Shahrajabian, M.H.; Sun, W. The Golden Spice for Life: Turmeric with the Pharmacological Benefits of Curcuminoids Components, including Curcumin, Bisdemethoxycurcumin, and Demethoxycurcumin. Curr. Org. Synth. 2023. ahead of print. [Google Scholar]
- Tang, Y.; Cao, Y. Curcumin Inhibits the Growth and Metastasis of Melanoma via miR-222-3p/SOX10/Notch Axis. Dis. Markers 2022, 2022, 3129781. [Google Scholar] [CrossRef] [PubMed]
- Rocha Silva, L.; Alves Nunes, J.; Zhan, P.; Łączkowski, K.Z.; Helena Cardoso, S.; Ferreira da Silva-Junior, E. Natural Coumarin Derivatives Targeting Melanoma. Curr. Med. Chem. 2023. ahead of print. [Google Scholar]
- Venza, I.; Visalli, M.; Oteri, R.; Beninati, C.; Teti, D.; Venza, M. Genistein reduces proliferation of EP3-expressing melanoma cells through inhibition of PGE2-induced IL-8 expression. Int. Immunopharmacol. 2018, 62, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Danciu, C.; Borcan, F.; Bojin, F.; Zupko, I.; Dehelean, C. Effect of the isoflavone genistein on tumor size, metastasis potential and melanization in a B16 mouse model of murine melanoma. Nat. Prod. Commun. 2013, 8, 343–346. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Fu, Y.; Malakhova, M.; Kurinov, I.; Zhu, F.; Yao, K.; Li, H.; Chen, H.; Li, W.; Lim, D.Y.; et al. Caffeic acid directly targets ERK1/2 to attenuate solar UV-induced skin carcinogenesis. Cancer Prev. Res. 2014, 7, 1056–1066. [Google Scholar] [CrossRef]
- Staniforth, V.; Chiu, L.T.; Yang, N.S. Caffeic acid suppresses UVB radiation-induced expression of interleukin-10 and activation of mitogen-activated protein kinases in mouse. Carcinogenesis 2006, 27, 1803–1811. [Google Scholar] [CrossRef]
- Agilan, B.; Rajendra Prasad, N.; Kanimozhi, G.; Karthikeyan, R.; Ganesan, M.; Mohana, S.; Velmurugan, D.; Ananthakrishnan, D. Caffeic Acid Inhibits Chronic UVB-Induced Cellular Proliferation Through JAK-STAT3 Signaling in Mouse Skin. Photochem. Photobiol. 2016, 92, 467–474. [Google Scholar] [CrossRef]
- Balupillai, A.; Prasad, R.N.; Ramasamy, K.; Muthusamy, G.; Shanmugham, M.; Govindasamy, K.; Gunaseelan, S. Caffeic Acid Inhibits UVB-induced Inflammation and Photocarcinogenesis through Activation of Peroxisome Proliferator-activated Receptor-gamma in Mouse Skin. Photochem. Photobiol. 2015, 91, 1458–1468. [Google Scholar] [CrossRef]
- Kang, N.J.; Lee, K.W.; Shin, B.J.; Jung, S.K.; Hwang, M.K.; Bode, A.M.; Heo, Y.S.; Lee, H.J.; Dong, Z. Caffeic acid, a phenolic phytochemical in coffee, directly inhibits Fyn kinase activity and UVB-induced COX-2 expression. Carcinogenesis 2009, 30, 321–330. [Google Scholar] [CrossRef]
- Balupillai, A.; Nagarajan, R.P.; Ramasamy, K.; Govindasamy, K.; Muthusamy, G. Caffeic acid prevents UVB radiation induced photocarcinogenesis through regulation of PTEN signaling in human dermal fibroblasts and mouse skin. Toxicol. Appl. Pharmacol. 2018, 352, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Pelinson, L.P.; Assmann, C.E.; Palma, T.V.; da Cruz, I.B.M.; Pillat, M.M.; Mânica, A.; Stefanello, N.; Weis, G.C.C.; de Oliveira Alves, A.; de Andrade, C.M.; et al. Antiproliferative and apoptotic effects of caffeic acid on SK-Mel-28 human melanoma cancer cells. Mol. Biol. Rep. 2019, 46, 2085–2092. [Google Scholar] [CrossRef] [PubMed]
- Kimsa-Dudek, M.; Synowiec-Wojtarowicz, A.; Krawczyk, A.; Kosowska, A.; Kimsa-Furdzik, M.; Francuz, T. The Apoptotic Effect of Caffeic or Chlorogenic Acid on the C32 Cells That Have Simultaneously Been Exposed to a Static Magnetic Field. Int. J. Mol. Sci. 2022, 23, 3859. [Google Scholar] [CrossRef]
- Kimsa-Dudek, M.; Synowiec-Wojtarowicz, A.; Krawczyk, A. Phenolic acids and a static magnetic field change the expression of transforming growth factor beta isoforms in amelanotic melanoma cells. Mol. Biol. Rep. 2023, 50, 4207–4216. [Google Scholar] [CrossRef] [PubMed]
- Kimsa-Dudek, M.; Krawczyk, A.; Synowiec-Wojtarowicz, A.; Dudek, S.; Pawlowska-Goral, K. The impact of the co-exposure of melanoma cells to chlorogenic acid and a moderate-strength static magnetic field. J. Food Biochem. 2020, 44, e13512. [Google Scholar] [CrossRef]
- Cui, K.; Zhang, L.; La, X.; Wu, H.; Yang, R.; Li, H.; Li, Z. Ferulic Acid and P-Coumaric Acid Synergistically Attenuate Non-Alcoholic Fatty Liver Disease through HDAC1/PPARG-Mediated Free Fatty Acid Uptake. Int. J. Mol. Sci. 2022, 23, 15297. [Google Scholar] [CrossRef]
- Ghaderi, S.; Gholipour, P.; Komaki, A.; Salehi, I.; Rashidi, K.; Esmaeil Khoshnam, S.; Rashno, M. p-Coumaric acid ameliorates cognitive and non-cognitive disturbances in a rat model of Alzheimer’s disease: The role of oxidative stress and inflammation. Int. Immunopharmacol. 2022, 112, 109295. [Google Scholar] [CrossRef]
- Moradi, M.; Farbood, Y.; Mard, S.A.; Dianat, M.; Goudarzi, G.; Khorsandi, L.; Seyedian, S.S. p-Coumaric acid has pure anti-inflammatory characteristics against hepatopathy caused by ischemia-reperfusion in the liver and dust exposure. Iran. J. Basic Med. Sci. 2023, 26, 164–175. [Google Scholar] [PubMed]
- Venkatesan, A.; Roy, A.; Kulandaivel, S.; Natesan, V.; Kim, S.J. p-Coumaric Acid Nanoparticles Ameliorate Diabetic Nephropathy via Regulating mRNA Expression of KIM-1 and GLUT-2 in Streptozotocin-Induced Diabetic Rats. Metabolites 2022, 12, 1166. [Google Scholar] [CrossRef]
- Tehami, W.; Nani, A.; Khan, N.A.; Hichami, A. New Insights into the Anticancer Effects of p-Coumaric Acid: Focus on Colorectal Cancer. Dose Response 2023, 21, 15593258221150704. [Google Scholar] [CrossRef]
- Saremi, S.; Kolahi, M.; Tabandeh, M.R.; Hashemitabar, M. Induction of apoptosis and suppression of Ras gene expression in MCF human breast cancer cells. J. Cancer Res. Ther. 2022, 18, 1052–1060. [Google Scholar]
- Hu, X.; Yang, Z.; Liu, W.; Pan, Z.; Zhang, X.; Li, M.; Liu, X.; Zheng, Q.; Li, D. The Anti-tumor Effects of p-Coumaric Acid on Melanoma A375 and B16 Cells. Front. Oncol. 2020, 10, 558414. [Google Scholar] [CrossRef] [PubMed]
- Gastaldello, G.H.; Cazeloto, A.C.V.; Ferreira, J.C.; Rodrigues, D.M.; Bastos, J.K.; Campo, V.L.; Zoccal, K.F.; Tefe-Silva, C. Green Propolis Compounds (Baccarin and p-Coumaric Acid) Show Beneficial Effects in Mice for Melanoma Induced by B16f10. Medicines 2021, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Ciazynska, M.; Olejniczak-Staruch, I.; Sobolewska-Sztychny, D.; Narbutt, J.; Skibinska, M.; Lesiak, A. Ultraviolet Radiation and Chronic Inflammation-Molecules and Mechanisms Involved in Skin Carcinogenesis: A Narrative Review. Life 2021, 11, 326. [Google Scholar] [CrossRef]
- Staniforth, V.; Huang, W.C.; Aravindaram, K.; Yang, N.S. Ferulic acid, a phenolic phytochemical, inhibits UVB-induced matrix metalloproteinases in mouse skin via posttranslational mechanisms. J. Nutr. Biochem. 2012, 23, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Ambothi, K.; Prasad, N.R.; Balupillai, A. Ferulic acid inhibits UVB-radiation induced photocarcinogenesis through modulating inflammatory and apoptotic signaling in Swiss albino mice. Food Chem. Toxicol. 2015, 82, 72–78. [Google Scholar] [CrossRef]
- Alias, L.M.; Manoharan, S.; Vellaichamy, L.; Balakrishnan, S.; Ramachandran, C.R. Protective effect of ferulic acid on 7,12-dimethylbenz[a]anthracene-induced skin carcinogenesis in Swiss albino mice. Exp. Toxicol. Pathol. 2009, 61, 205–214. [Google Scholar] [CrossRef]
- Wianowska, D.; Olszowy-Tomczyk, M. A Concise Profile of Gallic Acid-From Its Natural Sources through Biological Properties and Chemical Methods of Determination. Molecules 2023, 28, 1186. [Google Scholar] [CrossRef]
- Lo, C.; Lai, T.Y.; Yang, J.H.; Yang, J.S.; Ma, Y.S.; Weng, S.W.; Chen, Y.Y.; Lin, J.G.; Chung, J.G. Gallic acid induces apoptosis in A375.S2 human melanoma cells through caspase-dependent and -independent pathways. Int. J. Oncol. 2010, 37, 377–385. [Google Scholar]
- Lo, C.; Lai, T.Y.; Yang, J.S.; Yang, J.H.; Ma, Y.S.; Weng, S.W.; Lin, H.Y.; Chen, H.Y.; Lin, J.G.; Chung, J.G. Gallic acid inhibits the migration and invasion of A375.S2 human melanoma cells through the inhibition of matrix metalloproteinase-2 and Ras. Melanoma Res. 2011, 21, 267–273. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Lin, J.J.; Yang, Z.Y.; Tsai, C.C.; Hsu, J.L.; Wu, Y.J. Proteomic study reveals a co-occurrence of gallic acid-induced apoptosis and glycolysis in B16F10 melanoma cells. J. Agric. Food Chem. 2014, 62, 11672–11680. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, V.; Venkatesan, B.; Tumala, A.; Vellaichamy, E. Topical application of Gallic acid suppresses the 7,12-DMBA/Croton oil induced two-step skin carcinogenesis by modulating anti-oxidants and MMP-2/MMP-9 in Swiss albino mice. Food Chem. Toxicol. 2014, 66, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Tseng, T.H.; Hsu, J.D.; Lo, M.H.; Chu, C.Y.; Chou, F.P.; Huang, C.L.; Wang, C.J. Inhibitory effect of Hibiscus protocatechuic acid on tumor promotion in mouse skin. Cancer Lett. 1998, 126, 199–207. [Google Scholar] [CrossRef]
- Nakamura, Y.; Torikai, K.; Ohto, Y.; Murakami, A.; Tanaka, T.; Ohigashi, H. A simple phenolic antioxidant protocatechuic acid enhances tumor promotion and oxidative stress in female ICR mouse skin: Dose-and timing-dependent enhancement and involvement of bioactivation by tyrosinase. Carcinogenesis 2000, 21, 1899–1907. [Google Scholar] [CrossRef]
- Nakamura, Y.; Torikai, K.; Ohigashi, H. A catechol antioxidant protocatechuic acid potentiates inflammatory leukocyte-derived oxidative stress in mouse skin via a tyrosinase bioactivation pathway. Free Radic. Biol. Med. 2001, 30, 967–978. [Google Scholar] [CrossRef]
- Lin, H.H.; Chen, J.H.; Chou, F.P.; Wang, C.J. Protocatechuic acid inhibits cancer cell metastasis involving the down-regulation of Ras/Akt/NF-kappaB pathway and MMP-2 production by targeting RhoB activation. Br. J. Pharmacol. 2011, 162, 237–254. [Google Scholar] [CrossRef]
- da Silva, G.B.; Manica, D.; da Silva, A.P.; Marafon, F.; Moreno, M.; Bagatini, M.D. Rosmarinic acid decreases viability, inhibits migration and modulates expression of apoptosis-related CASP8/CASP3/NLRP3 genes in human metastatic melanoma cells. Chem. Biol. Interact. 2023, 375, 110427. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Chen, J.; Quan, J.; Xiang, D. Rosmarinic acid inhibits proliferation and migration, promotes apoptosis and enhances cisplatin sensitivity of melanoma cells through inhibiting ADAM17/EGFR/AKT/GSK3beta axis. Bioengineered 2021, 12, 3065–3076. [Google Scholar] [CrossRef] [PubMed]
- Olivares, A.; Alcaraz-Saura, M.; Achel, D.G.; Alcaraz, M. Effect of Rosmarinic Acid and Ionizing Radiation on Glutathione in Melanoma B16F10 Cells: A Translational Opportunity. Antioxidants 2020, 9, 1291. [Google Scholar] [CrossRef] [PubMed]
- Alcaraz, M.; Alcaraz-Saura, M.; Achel, D.G.; Olivares, A.; López-Morata, J.A.; Castillo, J. Radiosensitizing effect of rosmarinic acid in metastatic melanoma B16F10 cells. Anticancer Res. 2014, 34, 1913–1921. [Google Scholar] [PubMed]
- Sharmila, R.; Manoharan, S. Anti-tumor activity of rosmarinic acid in 7,12-dimethylbenz(a)anthracene (DMBA) induced skin carcinogenesis in Swiss albino mice. Indian J. Exp. Biol. 2012, 50, 187–194. [Google Scholar]
- Srinivasulu, C.; Ramgopal, M.; Ramanjaneyulu, G.; Anuradha, C.M.; Suresh Kumar, C. Syringic acid (SA)—A Review of Its Occurrence, Biosynthesis, Pharmacological and Industrial Importance. Biomed. Pharmacother. 2018, 108, 547–557. [Google Scholar] [CrossRef]
- Cikman, O.; Soylemez, O.; Ozkan, O.F.; Kiraz, H.A.; Sayar, I.; Ademoglu, S.; Taysi, S.; Karaayvaz, M. Antioxidant Activity of Syringic Acid Prevents Oxidative Stress in l-arginine-Induced Acute Pancreatitis: An Experimental Study on Rats. Int. Surg. 2015, 100, 891–896. [Google Scholar] [CrossRef]
- Mihanfar, A.; Darband, S.G.; Sadighparvar, S.; Kaviani, M.; Mirza-Aghazadeh-Attari, M.; Yousefi, B.; Majidinia, M. In vitro and in vivo anticancer effects of syringic acid on colorectal cancer: Possible mechanistic view. Chem. Biol. Interact. 2021, 337, 109337. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.J.; Lee, J.; Park, J.; Kim, Y.H.; Lee, N.H.; Kim, Y.E.; Song, K.M.; Chang, P.S.; Jeong, C.H.; Jung, S.K. Syringic acid prevents skin carcinogenesis via regulation of NoX and EGFR signaling. Biochem. Pharmacol. 2018, 154, 435–445. [Google Scholar] [CrossRef] [PubMed]
- Ghitu, A.; Schwiebs, A.; Radeke, H.H.; Avram, S.; Zupko, I.; Bor, A.; Pavel, I.Z.; Dehelean, C.A.; Oprean, C.; Bojin, F.; et al. A Comprehensive Assessment of Apigenin as an Antiproliferative, Proapoptotic, Antiangiogenic and Immunomodulatory Phytocompound. Nutrients 2019, 11, 858. [Google Scholar] [CrossRef]
- Madunic, J.; Madunic, I.V.; Gajski, G.; Popic, J.; Garaj-Vrhovac, V. Apigenin: A dietary flavonoid with diverse anticancer properties. Cancer Lett. 2018, 413, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Han, X.; Cheng, W.; Ni, J.; Zhang, Y.; Lin, J.; Song, Z. Apigenin inhibits proliferation and invasion, and induces apoptosis and cell cycle arrest in human melanoma cells. Oncol. Rep. 2017, 37, 2277–2285. [Google Scholar] [CrossRef]
- Cao, H.H.; Chu, J.H.; Kwan, H.Y.; Su, T.; Yu, H.; Cheng, C.Y.; Fu, X.Q.; Guo, H.; Li, T.; Tse, A.K.; et al. Inhibition of the STAT3 signaling pathway contributes to apigenin-mediated anti-metastatic effect in melanoma. Sci. Rep. 2016, 6, 21731. [Google Scholar] [CrossRef]
- Woo, J.S.; Choo, G.S.; Yoo, E.S.; Kim, S.H.; Lee, J.H.; Han, S.H.; Kim, H.J.; Jung, S.H.; Park, Y.S.; Kim, B.S.; et al. Apigenin induces apoptosis by regulating Akt and MAPK pathways in human melanoma cell A375SM. Mol. Med. Rep. 2020, 22, 4877–4889. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Zhang, Y.; Tian, K.; Chen, X.; Zhang, R.; Mu, X.; Wu, Y.; Wang, D.; Wang, S.; Liu, F.; et al. Apigenin suppresses PD-L1 expression in melanoma and host dendritic cells to elicit synergistic therapeutic effects. J. Exp. Clin. Cancer Res. 2018, 37, 261. [Google Scholar] [CrossRef] [PubMed]
- Mirzoeva, S.; Tong, X.; Bridgeman, B.B.; Plebanek, M.P.; Volpert, O.V. Apigenin Inhibits UVB-Induced Skin Carcinogenesis: The Role of Thrombospondin-1 as an Anti-Inflammatory Factor. Neoplasia 2018, 20, 930–942. [Google Scholar] [CrossRef] [PubMed]
- Bridgeman, B.B.; Wang, P.; Ye, B.; Pelling, J.C.; Volpert, O.V.; Tong, X. Inhibition of mTOR by apigenin in UVB-irradiated keratinocytes: A new implication of skin cancer prevention. Cell Signal 2016, 28, 460–468. [Google Scholar] [CrossRef]
- Byun, S.; Park, J.; Lee, E.; Lim, S.; Yu, J.G.; Lee, S.J.; Chen, H.; Dong, Z.; Lee, K.W.; Lee, H.J. Src kinase is a direct target of apigenin against UVB-induced skin inflammation. Carcinogenesis 2013, 34, 397–405. [Google Scholar] [CrossRef] [PubMed]
- Van Dross, R.T.; Hong, X.; Essengue, S.; Fischer, S.M.; Pelling, J.C. Modulation of UVB-induced and basal cyclooxygenase-2 (COX-2) expression by apigenin in mouse keratinocytes: Role of USF transcription factors. Mol. Carcinog. 2007, 46, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Caltagirone, S.; Rossi, C.; Poggi, A.; Ranelletti, F.O.; Natali, P.G.; Brunetti, M.; Aiello, F.B.; Piantelli, M. Flavonoids apigenin and quercetin inhibit melanoma growth and metastatic potential. Int. J. Cancer 2000, 87, 595–600. [Google Scholar] [CrossRef]
- Kiraly, A.J.; Soliman, E.; Jenkins, A.; Van Dross, R.T. Apigenin inhibits COX-2, PGE2, and EP1 and also initiates terminal differentiation in the epidermis of tumor bearing mice. Prostaglandins Leukot. Essent. Fatty Acids 2016, 104, 44–53. [Google Scholar] [CrossRef]
- Riaz, A.; Rasul, A.; Hussain, G.; Zahoor, M.K.; Jabeen, F.; Subhani, Z.; Younis, T.; Ali, M.; Sarfraz, I.; Selamoglu, Z. Astragalin: A Bioactive Phytochemical with Potential Therapeutic Activities. Adv. Pharmacol. Sci. 2018, 2018, 9794625. [Google Scholar] [CrossRef]
- You, O.H.; Shin, E.A.; Lee, H.; Kim, J.H.; Sim, D.Y.; Kim, J.H.; Kim, Y.; Khil, J.H.; Baek, N.I.; Kim, S.H. Apoptotic Effect of Astragalin in Melanoma Skin Cancers via Activation of Caspases and Inhibition of Sry-related HMg-Box Gene 10. Phytother. Res. 2017, 31, 1614–1620. [Google Scholar] [CrossRef]
- Rosenbaum, S.R.; Tiago, M.; Caksa, S.; Capparelli, C.; Purwin, T.J.; Kumar, G.; Glasheen, M.; Pomante, D.; Kotas, D.; Chervoneva, I.; et al. SOX10 requirement for melanoma tumor growth is due, in part, to immune-mediated effects. Cell Rep. 2021, 37, 110085. [Google Scholar] [CrossRef]
- Min, K.J.; Kwon, T.K. Anticancer effects and molecular mechanisms of epigallocatechin-3-gallate. Integr. Med. Res. 2014, 3, 16–24. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Ramasamy, V.; Moon, S.; Ruiz, C.; Muthugounder, S. Differential growth suppression of human melanoma cells by tea (Camellia sinensis) epicatechins (ECG, EGC and EGCG). Evid. Based Complement. Altern. Med. 2009, 6, 523–530. [Google Scholar] [CrossRef]
- Sudha, T.; Salaheldin, T.A.; Darwish, N.H.; Mousa, S.A. Antitumor/anti-angiogenesis efficacy of epigallocatechin gallate nanoformulated with antioxidant in melanoma. Nanomedicine 2022, 17, 1039–1053. [Google Scholar] [CrossRef]
- Du, B.X.; Lin, P.; Lin, J. EGCG and ECG induce apoptosis and decrease autophagy via the AMPK/mTOR and PI3K/AKT/mTOR pathway in human melanoma cells. Chin. J. Nat. Med. 2022, 20, 290–300. [Google Scholar] [CrossRef] [PubMed]
- Ravindran Menon, D.; Li, Y.; Yamauchi, T.; Osborne, D.G.; Vaddi, P.K.; Wempe, M.F.; Zhai, Z.; Fujita, M. EGCG Inhibits Tumor Growth in Melanoma by Targeting JAK-STAT Signaling and Its Downstream PD-L1/PD-L2-PD1 Axis in Tumors and Enhancing Cytotoxic T-Cell Responses. Pharmaceuticals 2021, 14, 1081. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.Y.; Jiang, Y.W.; Kuo, C.L.; Way, T.D.; Chou, Y.C.; Chang, Y.S.; Chung, J.G. Chrysin inhibit human melanoma A375.S2 cell migration and invasion via affecting MAPK signaling and NF-kappaB signaling pathway in vitro. Environ. Toxicol. 2019, 34, 434–442. [Google Scholar] [CrossRef]
- Sassi, A.; Maatouk, M.; El Gueder, D.; Bzeouich, I.M.; Abdelkefi-Ben Hatira, S.; Jemni-Yacoub, S.; Ghedira, K.; Chekir-Ghedira, L. Chrysin, a natural and biologically active flavonoid suppresses tumor growth of mouse B16F10 melanoma cells: In vitro and In vivo study. Chem. Biol. Interact. 2018, 283, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Pichichero, E.; Cicconi, R.; Mattei, M.; Canini, A. Chrysin-induced apoptosis is mediated through p38 and Bax activation in B16-F1 and A375 melanoma cells. Int. J. Oncol. 2011, 38, 473–483. [Google Scholar]
- Alvarez, N.; Vicente, V.; Martínez, C. Synergistic effect of diosmin and interferon-alpha on metastatic pulmonary melanoma. Cancer Biother. Radiopharm. 2009, 24, 347–352. [Google Scholar] [PubMed]
- Buddhan, R.; Manoharan, S. Diosmin reduces cell viability of A431 skin cancer cells through apoptotic induction. J. Cancer Res. Ther. 2017, 13, 471–476. [Google Scholar]
- Martínez Conesa, C.; Vicente Ortega, V.; Yáñez Gascón, M.J.; Alcaraz Baños, M.; Canteras Jordana, M.; Benavente-García, O.; Castillo, J. Treatment of metastatic melanoma B16F10 by the flavonoids tangeretin, rutin, and diosmin. J. Agric. Food Chem. 2005, 53, 6791–6797. [Google Scholar] [CrossRef] [PubMed]
- Martinez, C.; Vicente, V.; Yanez, J.; Alcaraz, M.; Castells, M.T.; Canteras, M.; Benavente-Garcia, O.; Castillo, J. The effect of the flavonoid diosmin, grape seed extract and red wine on the pulmonary metastatic B16F10 melanoma. Histol. Histopathol. 2005, 20, 1121–1129. [Google Scholar] [PubMed]
- Zhang, W.; Tang, B.; Huang, Q.; Hua, Z. Galangin inhibits tumor growth and metastasis of B16F10 melanoma. J. Cell Biochem. 2013, 114, 152–161. [Google Scholar] [CrossRef]
- Benguedouar, L.; Lahouel, M.; Gangloff, S.C.; Durlach, A.; Grange, F.; Bernard, P.; Antonicelli, F. Ethanolic Extract of Algerian Propolis and Galangin Decreased Murine Melanoma Tumor Progression in Mice. Anticancer Agents Med. Chem. 2016, 16, 1172–1183. [Google Scholar] [CrossRef]
- Madduma Hewage, S.R.; Piao, M.J.; Kim, K.C.; Cha, J.W.; Han, X.; Choi, Y.H.; Chae, S.; Hyun, J.W. Galangin (3,5,7-trihydroxyflavone) shields human keratinocytes from ultraviolet B-induced oxidative stress. Biomol. Ther. 2015, 23, 165–173. [Google Scholar] [CrossRef] [PubMed]
- Madduma Hewage, S.R.K.; Piao, M.J.; Kang, K.A.; Ryu, Y.S.; Fernando, P.; Oh, M.C.; Park, J.E.; Shilnikova, K.; Moon, Y.J.; Shin, D.O.; et al. Galangin Activates the ERK/AKT-Driven Nrf2 Signaling Pathway to Increase the Level of Reduced Glutathione in Human Keratinocytes. Biomol. Ther. 2017, 25, 427–433. [Google Scholar] [CrossRef]
- Jaiswal, N.; Akhtar, J.; Singh, S.P.; Badruddeen Ahsan, F. An Overview on Genistein and its Various Formulations. Drug Res. 2019, 69, 305–313. [Google Scholar] [CrossRef]
- Hou, S. Genistein: Therapeutic and Preventive Effects, Mechanisms, and Clinical Application in Digestive Tract Tumor. Evid. Based Complement. Altern. Med. 2022, 2022, 5957378. [Google Scholar] [CrossRef] [PubMed]
- Bhat, S.S.; Prasad, S.K.; Shivamallu, C.; Prasad, K.S.; Syed, A.; Reddy, P.; Cull, C.A.; Amachawadi, R.G. Genistein: A Potent Anti-Breast Cancer Agent. Curr. Issues Mol. Biol. 2021, 43, 1502–1517. [Google Scholar] [CrossRef]
- Wang, H.Z.; Zhang, Y.; Xie, L.P.; Yu, X.Y.; Zhang, R.Q. Effects of genistein and daidzein on the cell growth, cell cycle, and differentiation of human and murine melanoma cells(1). J. Nutr. Biochem. 2002, 13, 421–426. [Google Scholar] [CrossRef]
- Darbon, J.M.; Penary, M.; Escalas, N.; Casagrande, F.; Goubin-Gramatica, F.; Baudouin, C.; Ducommun, B. Distinct Chk2 activation pathways are triggered by genistein and DNA-damaging agents in human melanoma cells. J Biol. Chem. 2000, 275, 15363–15369. [Google Scholar] [CrossRef]
- Cui, S.; Wang, J.; Wu, Q.; Qian, J.; Yang, C.; Bo, P. Genistein inhibits the growth and regulates the migration and invasion abilities of melanoma cells via the FAK/paxillin and MAPK pathways. Oncotarget 2017, 8, 21674–21691. [Google Scholar] [CrossRef] [PubMed]
- Heo, J.R.; Lee, G.A.; Kim, G.S.; Hwang, K.A.; Choi, K.C. Phytochemical-induced reactive oxygen species and endoplasmic reticulum stress-mediated apoptosis and differentiation in malignant melanoma cells. Phytomedicine 2018, 39, 100–110. [Google Scholar] [CrossRef]
- Tamura, S.; Bito, T.; Ichihashi, M.; Ueda, M. Genistein enhances the cisplatin-induced inhibition of cell growth and apoptosis in human malignant melanoma cells. Pigment. Cell Res. 2003, 16, 470–476. [Google Scholar] [CrossRef] [PubMed]
- Alhasan, S.A.; Ensley, J.F.; Sarkar, F.H. Genistein induced molecular changes in a squamous cell carcinoma of the head and neck cell line. Int. J. Oncol. 2000, 16, 333–341. [Google Scholar] [CrossRef] [PubMed]
- Record, I.R.; Broadbent, J.L.; King, R.A.; Dreosti, I.E.; Head, R.J.; Tonkin, A.L. Genistein inhibits growth of B16 melanoma cells in vivo and in vitro and promotes differentiation in vitro. Int. J. Cancer 1997, 72, 860–864. [Google Scholar] [CrossRef]
- Menon, L.G.; Kuttan, R.; Nair, M.G.; Chang, Y.C.; Kuttan, G. Effect of isoflavones genistein and daidzein in the inhibition of lung metastasis in mice induced by B16F-10 melanoma cells. Nutr. Cancer 1998, 30, 74–77. [Google Scholar] [CrossRef]
- Farina, H.G.; Pomies, M.; Alonso, D.F.; Gomez, D.E. Antitumor and antiangiogenic activity of soy isoflavone genistein in mouse models of melanoma and breast cancer. Oncol. Rep. 2006, 16, 885–891. [Google Scholar] [CrossRef]
- Wei, H.; Bowen, R.; Zhang, X.; Lebwohl, M. Isoflavone genistein inhibits the initiation and promotion of two-stage skin carcinogenesis in mice. Carcinogenesis 1998, 19, 1509–1514. [Google Scholar] [CrossRef]
- Wei, H.; Saladi, R.; Lu, Y.; Wang, Y.; Palep, S.R.; Moore, J.; Phelps, R.; Shyong, E.; Lebwohl, M.G. Isoflavone genistein: Photoprotection and clinical implications in dermatology. J. Nutr. 2003, 133 (Suppl. S1), 3811S–3819S. [Google Scholar] [CrossRef] [PubMed]
- Wei, H.; Zhang, X.; Wang, Y.; Lebwohl, M. Inhibition of ultraviolet light-induced oxidative events in the skin and internal organs of hairless mice by isoflavone genistein. Cancer Lett. 2002, 185, 21–29. [Google Scholar] [CrossRef]
- Valentova, K.; Vrba, J.; Bancirova, M.; Ulrichova, J.; Kren, V. Isoquercitrin: Pharmacology, toxicology, and metabolism. Food Chem. Toxicol. 2014, 68, 267–282. [Google Scholar] [CrossRef] [PubMed]
- Wullschleger, S.; Loewith, R.; Hall, M.N. TOR signaling in growth and metabolism. Cell 2006, 124, 471–484. [Google Scholar] [CrossRef] [PubMed]
- Won, Y.S.; Kim, J.H.; Lizardo, R.C.M.; Min, H.J.; Cho, H.D.; Hong, S.M.; Seo, K.I. The Flavonol Isoquercitrin Promotes Mitochondrial-Dependent Apoptosis in SK-Mel-2 Melanoma Cell via the PI3K/AKT/mTOR Pathway. Nutrients 2020, 12, 3683. [Google Scholar] [CrossRef]
- Gong, G.; Guan, Y.Y.; Zhang, Z.L.; Rahman, K.; Wang, S.J.; Zhou, S.; Luan, X.; Zhang, H. Isorhamnetin: A review of pharmacological effects. Biomed. Pharmacother. 2020, 128, 110301. [Google Scholar] [CrossRef]
- Duan, R.; Liang, X.; Chai, B.; Zhou, Y.; Du, H.; Suo, Y.; Chen, Z.; Li, Q.; Huang, X. Isorhamnetin Induces Melanoma Cell Apoptosis via the PI3K/Akt and NF-kappaB Pathways. Biomed. Res. Int. 2020, 2020, 1057943. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.E.; Lee, D.E.; Lee, K.W.; Son, J.E.; Seo, S.K.; Li, J.; Jung, S.K.; Heo, Y.S.; Mottamal, M.; Bode, A.M.; et al. Isorhamnetin suppresses skin cancer through direct inhibition of MEK1 and PI3-K. Cancer Prev. Res. 2011, 4, 582–591. [Google Scholar] [CrossRef]
- Li, J.; Mottamal, M.; Li, H.; Liu, K.; Zhu, F.; Cho, Y.Y.; Sosa, C.P.; Zhou, K.; Bowden, G.T.; Bode, A.M.; et al. Quercetin-3-methyl ether suppresses proliferation of mouse epidermal JB6 P+ cells by targeting ERKs. Carcinogenesis 2012, 33, 459–465. [Google Scholar] [CrossRef]
- Qiang, D.; Ci, C.; Liu, W.; Wang, J.; He, C.; Ji, B.; Shao, X. Inhibitory effect of kaempferol on mouse melanoma cell line B16 in vivo and in vitro. Adv. Dermatol. Allergol. 2021, 38, 498–504. [Google Scholar] [CrossRef]
- Yang, J.; Xiao, P.; Sun, J.; Guo, L. Anticancer effects of kaempferol in A375 human malignant melanoma cells are mediated via induction of apoptosis, cell cycle arrest, inhibition of cell migration and downregulation of m-TOR/PI3K/AKT pathway. J. BUON 2018, 23, 218–223. [Google Scholar]
- Zheng, X.; Pan, Y.; Yang, G.; Liu, Y.; Zou, J.; Zhao, H.; Yin, G.; Wu, Y.; Li, X.; Wei, Z.; et al. Kaempferol impairs aerobic glycolysis against melanoma metastasis via inhibiting the mitochondrial binding of HK2 and VDAC1. Eur. J. Pharmacol. 2022, 931, 175226. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.; Chen, H.; Liu, K.; Langfald, A.; Yang, G.; Zhang, Y.; Yu, D.H.; Kim, M.O.; Lee, M.-H.; Li, H.; et al. Kaempferol Targets RSK2 and MSK1 to Suppress UV Radiation-Induced Skin Cancer. Cancer Prev. Res. 2014, 7, 958–967. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Lee, K.W.; Jung, S.K.; Lee, E.J.; Heo, Y.S.; Bode, A.M.; Lubet, R.A.; Lee, H.J.; Dong, Z. Kaempferol inhibits UVB-induced COX-2 expression by suppressing Src kinase activity. Biochem. Pharmacol. 2010, 80, 2042–2049. [Google Scholar] [CrossRef] [PubMed]
- Schomberg, J.; Wang, Z.; Farhat, A.; Guo, K.L.; Xie, J.; Zhou, Z.; Liu, J.; Kovacs, B.; Liu-Smith, F. Luteolin inhibits melanoma growth in vitro and in vivo via regulating ECM and oncogenic pathways but not ROS. Biochem. Pharmacol. 2020, 177, 114025. [Google Scholar] [CrossRef]
- Yao, X.; Jiang, W.; Yu, D.; Yan, Z. Luteolin inhibits proliferation and induces apoptosis of human melanoma cells in vivo and in vitro by suppressing MMP-2 and MMP-9 through the PI3K/AKT pathway. Food Funct. 2019, 10, 703–712. [Google Scholar] [CrossRef]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef]
- Li, C.; Wang, Q.; Shen, S.; Wei, X.; Li, G. HIF-1α/VEGF signaling-mediated epithelial-mesenchymal transition and angiogenesis is critically involved in anti-metastasis effect of luteolin in melanoma cells. Phytother. Res. 2019, 33, 798–807. [Google Scholar] [CrossRef]
- Ruan, J.S.; Liu, Y.P.; Zhang, L.; Yan, L.G.; Fan, F.T.; Shen, C.S.; Wang, A.Y.; Zheng, S.Z.; Wang, S.M.; Lu, Y. Luteolin reduces the invasive potential of malignant melanoma cells by targeting beta3 integrin and the epithelial-mesenchymal transition. Acta Pharmacol. Sin. 2012, 33, 1325–1331. [Google Scholar] [CrossRef]
- Kim, J.K.; Kang, K.A.; Ryu, Y.S.; Piao, M.J.; Han, X.; Oh, M.C.; Boo, S.J.; Jeong, S.U.; Jeong, Y.J.; Chae, S.; et al. Induction of Endoplasmic Reticulum Stress via Reactive Oxygen Species Mediated by Luteolin in Melanoma Cells. Anticancer Res. 2016, 36, 2281–2289. [Google Scholar] [PubMed]
- Li, T.; Fu, X.; Liu, B.; Wang, X.; Li, J.; Zhu, P.; Niu, X.; Bai, J.; Liu, Y.; Lu, X.; et al. Luteolin binds Src, promotes STAT3 protein ubiquitination and exerts anti-melanoma effects in cell and mouse models. Biochem. Pharmacol. 2022, 200, 115044. [Google Scholar] [CrossRef] [PubMed]
- George, V.C.; Naveen Kumar, D.R.; Suresh, P.K.; Kumar, S.; Kumar, R.A. Comparative studies to evaluate relative in vitro potency of luteolin in inducing cell cycle arrest and apoptosis in HaCaT and A375 cells. Asian Pac. J. Cancer Prev. 2013, 14, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Neagu, M.; Constantin, C.; Caruntu, C.; Dumitru, C.; Surcel, M.; Zurac, S. Inflammation: A key process in skin tumorigenesis. Oncol. Lett. 2019, 17, 4068–4084. [Google Scholar] [CrossRef]
- Wolfle, U.; Esser, P.R.; Simon-Haarhaus, B.; Martin, S.F.; Lademann, J.; Schempp, C.M. UVB-induced DNA damage, generation of reactive oxygen species, and inflammation are effectively attenuated by the flavonoid luteolin in vitro and in vivo. Free Radic. Biol. Med. 2011, 50, 1081–1093. [Google Scholar] [CrossRef] [PubMed]
- Kang, N.J.; Jung, S.K.; Lee, K.W.; Lee, H.J. Myricetin is a potent chemopreventive phytochemical in skin carcinogenesis. Ann. N. Y. Acad. Sci. 2011, 1229, 124–132. [Google Scholar] [CrossRef]
- Afroze, N.; Pramodh, S.; Hussain, A.; Waleed, M.; Vakharia, K. A review on myricetin as a potential therapeutic candidate for cancer prevention. 3 Biotech. 2020, 10, 211. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.K.; Lee, K.W.; Byun, S.; Kang, N.J.; Lim, S.H.; Heo, Y.S.; Bode, A.M.; Bowden, G.T.; Lee, H.J.; Dong, Z. Myricetin suppresses UVB-induced skin cancer by targeting Fyn. Cancer Res. 2008, 68, 6021–6029. [Google Scholar] [CrossRef]
- Sun, W.; Tao, Y.; Yu, D.; Zhao, T.; Wu, L.; Yu, W.; Han, W. Myricetin exerts potent anticancer effects on human skin tumor cells. Trop. J. Pharm. 2018, 17, 1067–1072. [Google Scholar] [CrossRef]
- Lee, K.M.; Kang, N.J.; Han, J.H.; Lee, K.W.; Lee, H.J. Myricetin down-regulates phorbol ester-induced cyclooxygenase-2 expression in mouse epidermal cells by blocking activation of nuclear factor kappa B. J. Agric. Food Chem. 2007, 55, 9678–9684. [Google Scholar] [CrossRef]
- Lee, K.W.; Kang, N.J.; Rogozin, E.A.; Kim, H.G.; Cho, Y.Y.; Bode, A.M.; Lee, H.J.; Surh, Y.J.; Bowden, G.T.; Dong, Z. Myricetin is a novel natural inhibitor of neoplastic cell transformation and MEK1. Carcinogenesis 2007, 28, 1918–1927. [Google Scholar] [CrossRef]
- Kumamoto, T.; Fujii, M.; Hou, D.X. Myricetin directly targets JAK1 to inhibit cell transformation. Cancer Lett. 2009, 275, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Jung, S.K.; Lee, K.W.; Byun, S.; Lee, E.J.; Kim, J.E.; Bode, A.M.; Dong, Z.; Lee, H.J. Myricetin inhibits UVB-induced angiogenesis by regulating PI-3 kinase in vivo. Carcinogenesis 2010, 31, 911–917. [Google Scholar] [CrossRef]
- Choi, J.; Lee, D.H.; Jang, H.; Park, S.Y.; Seol, J.W. Naringenin exerts anticancer effects by inducing tumor cell death and inhibiting angiogenesis in malignant melanoma. Int. J. Med. Sci. 2020, 17, 3049–3057. [Google Scholar] [CrossRef] [PubMed]
- Pafumi, I.; Festa, M.; Papacci, F.; Lagostena, L.; Giunta, C.; Gutla, V.; Cornara, L.; Favia, A.; Palombi, F.; Gambale, F.; et al. Naringenin Impairs Two-Pore Channel 2 Activity And Inhibits VEGF-Induced Angiogenesis. Sci. Rep. 2017, 7, 5121. [Google Scholar] [CrossRef]
- Skelding, K.A.; Barry, D.L.; Theron, D.Z.; Lincz, L.F. Targeting the two-pore channel 2 in cancer progression and metastasis. Explor. Target Antitumor. Ther. 2022, 3, 62–89. [Google Scholar] [PubMed]
- Lentini, A.; Forni, C.; Provenzano, B.; Beninati, S. Enhancement of transglutaminase activity and polyamine depletion in B16-F10 melanoma cells by flavonoids naringenin and hesperitin correlate to reduction of the in vivo metastatic potential. Amino Acids 2007, 32, 95–100. [Google Scholar] [PubMed]
- Ahamad, M.S.; Siddiqui, S.; Jafri, A.; Ahmad, S.; Afzal, M.; Arshad, M. Induction of apoptosis and antiproliferative activity of naringenin in human epidermoid carcinoma cell through ROS generation and cell cycle arrest. PLoS ONE 2014, 9, e110003. [Google Scholar]
- Kumar, R.; Bhan Tiku, A. Naringenin Suppresses Chemically Induced Skin Cancer in Two-Stage Skin Carcinogenesis Mouse Model. Nutr. Cancer 2020, 72, 976–983. [Google Scholar] [PubMed]
- Martinez, R.M.; Pinho-Ribeiro, F.A.; Steffen, V.S.; Caviglione, C.V.; Vignoli, J.A.; Barbosa, D.S.; Baracat, M.M.; Georgetti, S.R.; Verri, W.A., Jr.; Casagrande, R. Naringenin Inhibits UVB Irradiation-Induced Inflammation and Oxidative Stress in the Skin of Hairless Mice. J. Nat. Prod. 2015, 78, 1647–1655. [Google Scholar] [CrossRef] [PubMed]
- Martinez, R.M.; Pinho-Ribeiro, F.A.; Steffen, V.S.; Silva, T.C.; Caviglione, C.V.; Bottura, C.; Fonseca, M.J.; Vicentini, F.T.; Vignoli, J.A.; Baracat, M.M.; et al. Topical Formulation Containing Naringenin: Efficacy against Ultraviolet B Irradiation-Induced Skin Inflammation and Oxidative Stress in Mice. PLoS ONE 2016, 11, e0146296. [Google Scholar]
- Anand David, A.V.; Arulmoli, R.; Parasuraman, S. Overviews of Biological Importance of Quercetin: A Bioactive Flavonoid. Pharmacogn. Rev. 2016, 10, 84–89. [Google Scholar] [PubMed]
- Soll, F.; Ternent, C.; Berry, I.M.; Kumari, D.; Moore, T.C. Quercetin Inhibits Proliferation and Induces Apoptosis of B16 Melanoma Cells In Vitro. Assay Drug Dev. Technol. 2020, 18, 261–268. [Google Scholar] [CrossRef]
- Zhang, X.M.; Chen, J.; Xia, Y.G.; Xu, Q. Apoptosis of murine melanoma B16-BL6 cells induced by quercetin targeting mitochondria, inhibiting expression of PKC-alpha and translocating PKC-delta. Cancer Chemother. Pharmacol. 2005, 55, 251–262. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, Q.; Saiki, I. Quercetin inhibits the invasion and mobility of murine melanoma B16-BL6 cells through inducing apoptosis via decreasing Bcl-2 expression. Clin. Exp. Metastasis 2000, 18, 415–421. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.H.; Tse, A.K.; Kwan, H.Y.; Yu, H.; Cheng, C.Y.; Su, T.; Fong, W.F.; Yu, Z.L. Quercetin exerts anti-melanoma activities and inhibits STAT3 signaling. Biochem. Pharmacol. 2014, 87, 424–434. [Google Scholar] [CrossRef]
- Kim, S.H.; Yoo, E.S.; Woo, J.S.; Han, S.H.; Lee, J.H.; Jung, S.H.; Kim, H.J.; Jung, J.Y. Antitumor and apoptotic effects of quercetin on human melanoma cells involving JNK/P38 MAPK signaling activation. Eur. J. Pharmacol. 2019, 860, 172568. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.M.; Huang, S.P.; Xu, Q. Quercetin inhibits the invasion of murine melanoma B16-BL6 cells by decreasing pro-MMP-9 via the PKC pathway. Cancer Chemother. Pharmacol. 2004, 53, 82–88. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.S.; Tsai, P.H.; Kandaswami, C.C.; Cheng, C.H.; Ke, F.C.; Lee, P.P.; Hwang, J.J.; Lee, M.T. Effects of dietary flavonoids, luteolin, and quercetin on the reversal of epithelial-mesenchymal transition in A431 epidermal cancer cells. Cancer Sci. 2011, 102, 1829–1839. [Google Scholar] [CrossRef]
- Patel, D.H.; Sharma, N. Inhibitory effect of quercetin on epithelial to mesenchymal transition in SK-MEL-28 human melanoma cells defined by in vitro analysis on 3D collagen gels. OncoTargets Ther. 2016, 9, 6445–6459. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.J.; Hsu, W.H.; Lee, K.H.; Chen, K.C.; Lin, C.W.; Lee, Y.A.; Ko, T.P.; Lee, L.T.; Lee, M.T.; Chang, M.S.; et al. Dietary Flavonoids Luteolin and Quercetin Inhibit Migration and Invasion of Squamous Carcinoma through Reduction of Src/Stat3/S100A7 Signaling. Antioxidants 2019, 8, 557. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.H.; Cheng, C.Y.; Su, T.; Fu, X.Q.; Guo, H.; Li, T.; Tse, A.K.; Kwan, H.Y.; Yu, H.; Yu, Z.L. Quercetin inhibits HGF/c-Met signaling and HGF-stimulated melanoma cell migration and invasion. Mol. Cancer 2015, 14, 103. [Google Scholar]
- Huang, Y.T.; Hwang, J.J.; Lee, P.P.; Ke, F.C.; Huang, J.H.; Huang, C.J.; Kandaswami, C.; Middleton, E., Jr.; Lee, M.T. Effects of luteolin and quercetin, inhibitors of tyrosine kinase, on cell growth and metastasis-associated properties in A431 cells overexpressing epidermal growth factor receptor. Br. J. Pharmacol. 1999, 128, 999–1010. [Google Scholar]
- Rafiq, R.A.; Quadri, A.; Nazir, L.A.; Peerzada, K.; Ganai, B.A.; Tasduq, S.A. A Potent Inhibitor of Phosphoinositide 3-Kinase (PI3K) and Mitogen Activated Protein (MAP) Kinase Signalling, Quercetin (3, 3′, 4′, 5, 7-Pentahydroxyflavone) Promotes Cell Death in Ultraviolet (UV)-B-Irradiated B16F10 Melanoma Cells. PLoS ONE 2015, 10, e0131253. [Google Scholar]
- Peng, D.; Chen, L.; Sun, Y.; Sun, L.; Yin, Q.; Deng, S.; Niu, L.; Lou, F.; Wang, Z.; Xu, Z.; et al. Melanoma suppression by quercein is correlated with RIG-I and type I interferon signaling. Biomed. Pharmacother. 2020, 125, 109984. [Google Scholar] [PubMed]
- Jung, M.; Bu, S.Y.; Tak, K.H.; Park, J.E.; Kim, E. Anticarcinogenic effect of quercetin by inhibition of insulin-like growth factor (IGF)-1 signaling in mouse skin cancer. Nutr. Res. Pract. 2013, 7, 439–445. [Google Scholar] [CrossRef]
- Vicentini, F.T.; Simi, T.R.; Del Ciampo, J.O.; Wolga, N.O.; Pitol, D.L.; Iyomasa, M.M.; Bentley, M.V.; Fonseca, M.J. Quercetin in w/o microemulsion: In vitro and in vivo skin penetration and efficacy against UVB-induced skin damages evaluated in vivo. Eur. J. Pharm. Biopharm. 2008, 69, 948–957. [Google Scholar] [CrossRef]
- Liu, D.; Hu, H.; Lin, Z.; Chen, D.; Zhu, Y.; Hou, S.; Shi, X. Quercetin deformable liposome: Preparation and efficacy against ultraviolet B induced skin damages in vitro and in vivo. J. Photochem. Photobiol. B 2013, 127, 8–17. [Google Scholar] [CrossRef]
- Zhu, X.; Zeng, X.; Zhang, X.; Cao, W.; Wang, Y.; Chen, H.; Wang, T.; Tsai, H.I.; Zhang, R.; Chang, D.; et al. The effects of quercetin-loaded PLGA-TPGS nanoparticles on ultraviolet B-induced skin damages in vivo. Nanomedicine 2016, 12, 623–632. [Google Scholar] [CrossRef] [PubMed]
- Nan, W.; Ding, L.; Chen, H.; Khan, F.U.; Yu, L.; Sui, X.; Shi, X. Topical Use of Quercetin-Loaded Chitosan Nanoparticles Against Ultraviolet B Radiation. Front. Pharmacol. 2018, 9, 826. [Google Scholar] [CrossRef] [PubMed]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [PubMed]
- Pinzaru, I.; Chioibas, R.; Marcovici, I.; Coricovac, D.; Susan, R.; Predut, D.; Georgescu, D.; Dehelean, C. Rutin Exerts Cytotoxic and Senescence-Inducing Properties in Human Melanoma Cells. Toxics 2021, 9, 226. [Google Scholar] [CrossRef] [PubMed]
- Menon, L.G.; Kuttan, R.; Kuttan, G. Inhibition of lung metastasis in mice induced by B16F10 melanoma cells by polyphenolic compounds. Cancer Lett. 1995, 95, 221–225. [Google Scholar] [CrossRef]
- Choi, K.S.; Kundu, J.K.; Chun, K.S.; Na, H.K.; Surh, Y.J. Rutin inhibits UVB radiation-induced expression of COX-2 and iNOS in hairless mouse skin: P38 MAP kinase and JNK as potential targets. Arch. Biochem. Biophys. 2014, 559, 38–45. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Wang, K.S.; Qi, M.; Zhou, Y.J.; Zeng, G.Y.; Tao, J.; Zhou, J.D.; Zhang, J.L.; Chen, X.; Peng, C. Vitexin compound 1, a novel extraction from a Chinese herb, suppresses melanoma cell growth through DNA damage by increasing ROS levels. J. Exp. Clin. Cancer Res. 2018, 37, 269. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Zhou, L.; Guo, L. Vitexin inhibited the invasion, metastasis, and progression of human melanoma cells by targeting STAT3 signaling pathway. bioRxiv 2020. [Google Scholar] [CrossRef]
- Murakami, A.; Ohura, S.; Nakamura, Y.; Koshimizu, K.; Ohigashi, H. 1′-Acetoxychavicol acetate, a superoxide anion generation inhibitor, potently inhibits tumor promotion by 12-O-tetradecanoylphorbol-13-acetate in ICR mouse skin. Oncology 1996, 53, 386–391. [Google Scholar] [CrossRef]
- Batra, V.; Syed, Z.; Gill, J.N.; Coburn, M.A.; Adegboyega, P.; DiGiovanni, J.; Mathis, J.M.; Shi, R.; Clifford, J.L.; Kleiner-Hancock, H.E. Effects of the tropical ginger compound,1′-acetoxychavicol acetate, against tumor promotion in K5.Stat3C transgenic mice. J. Exp. Clin. Cancer Res. 2012, 31, 57. [Google Scholar] [CrossRef]
- Li, J.; Mao, Y.; Li, W. Anethole suppresses the growth of human skin cancer cells by targeting microRNA498/STAT4 axis. Trop. J. Pharm. 2022, 21, 2391–2396. [Google Scholar] [CrossRef]
- Wang, T.; Ma, L.; Li, W.; Ding, L.; Gao, H. MicroRNA-498 reduces the proliferation and invasion of colorectal cancer cells via targeting Bcl-2. FEBS Open Bio 2020, 10, 168–175. [Google Scholar] [CrossRef]
- Zhang, X.; Xu, X.; Ge, G.; Zang, X.; Shao, M.; Zou, S.; Zhang, Y.; Mao, Z.; Zhang, J.; Mao, F.; et al. miR-498 inhibits the growth and metastasis of liver cancer by targeting ZEB2. Oncol. Rep. 2019, 41, 1638–1648. [Google Scholar] [CrossRef]
- Lombardi, V.R.; Carrera, I.; Cacabelos, R. In Vitro Screening for Cytotoxic Activity of Herbal Extracts. Evid. Based Complement. Altern. Med. 2017, 2017, 2675631. [Google Scholar]
- Nahar, L.; Al-Groshi, A.; Kumar, A.; Sarker, S.D. Arbutin: Occurrence in Plants, and Its Potential as an Anticancer Agent. Molecules 2022, 27, 8786. [Google Scholar]
- Jiang, L.; Wang, D.; Zhang, Y.; Li, J.; Wu, Z.; Wang, Z.; Wang, D. Investigation of the pro-apoptotic effects of arbutin and its acetylated derivative on murine melanoma cells. Int. J. Mol. Med. 2018, 41, 1048–1054. [Google Scholar] [CrossRef]
- Nawarak, J.; Huang-Liu, R.; Kao, S.H.; Liao, H.H.; Sinchaikul, S.; Chen, S.T.; Cheng, S.L. Proteomics analysis of A375 human malignant melanoma cells in response to arbutin treatment. Biochim. Biophys. Acta 2009, 1794, 159–167. [Google Scholar] [CrossRef]
- Caterina, M.J.; Schumacher, M.A.; Tominaga, M.; Rosen, T.A.; Levine, J.D.; Julius, D. The capsaicin receptor: A heat-activated ion channel in the pain pathway. Nature 1997, 389, 816–824. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.Y.; Huang, H.H.; Li, Q.C.; Zhan, F.B.; Wang, L.B.; He, T.; Yang, C.H.; Wang, Y.; Zhang, Y.; Quan, Z.X. Capsaicin Reduces Cancer Stemness and Inhibits Metastasis by Downregulating SOX2 and EZH2 in Osteosarcoma. Am. J. Chin. Med. 2023, 51, 1041–1066. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.Y. Nitric oxide triggers apoptosis in A375 human melanoma cells treated with capsaicin and resveratrol. Mol. Med. Rep. 2012, 5, 585–591. [Google Scholar] [CrossRef]
- Shin, D.H.; Kim, O.H.; Jun, H.S.; Kang, M.K. Inhibitory effect of capsaicin on B16-F10 melanoma cell migration via the phosphatidylinositol 3-kinase/Akt/Rac1 signal pathway. Exp. Mol. Med. 2008, 40, 486–494. [Google Scholar] [CrossRef]
- Jun, H.S.; Park, T.; Lee, C.K.; Kang, M.K.; Park, M.S.; Kang, H.I.; Surh, Y.J.; Kim, O.H. Capsaicin induced apoptosis of B16-F10 melanoma cells through down-regulation of Bcl-2. Food Chem. Toxicol. 2007, 45, 708–715. [Google Scholar] [CrossRef]
- Chu, H.; Li, M.; Wang, X. Capsaicin induces apoptosis and autophagy in human melanoma cells. Oncol. Lett. 2019, 17, 4827–4834. [Google Scholar] [CrossRef] [PubMed]
- Islam, A.; Hsieh, P.F.; Liu, P.F.; Chou, J.C.; Liao, J.W.; Hsieh, M.K.; Chueh, P.J. Capsaicin exerts therapeutic effects by targeting tNOX-SIRT1 axis and augmenting ROS-dependent autophagy in melanoma cancer cells. Am. J. Cancer Res. 2021, 11, 4199–4219. [Google Scholar]
- Bahri, S.; Jameleddine, S.; Shlyonsky, V. Relevance of carnosic acid to the treatment of several health disorders: Molecular targets and mechanisms. Biomed. Pharmacother. 2016, 84, 569–582. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.I.; Lin, C.C.; Kuo, S.M.; Lai, J.C.; Wang, Y.Q.; You, H.L.; Hsu, M.L.; Chen, C.H.; Shiu, L.Y. Carnosic acid impedes cell growth and enhances anticancer effects of carmustine and lomustine in melanoma. Biosci. Rep. 2018, 38, BSR20180005. [Google Scholar] [CrossRef] [PubMed]
- Park, S.Y.; Song, H.; Sung, M.K.; Kang, Y.H.; Lee, K.W.; Park, J.H. Carnosic acid inhibits the epithelial-mesenchymal transition in B16F10 melanoma cells: A possible mechanism for the inhibition of cell migration. Int. J. Mol. Sci. 2014, 15, 12698–12713. [Google Scholar] [CrossRef] [PubMed]
- Alcaraz, M.; Olivares, A.; Andreu-Galvez, M.; Achel, D.G.; Mercado, A.M.; Alcaraz-Saura, M. Paradoxical Radiosensitizing Effect of Carnosic Acid on B16F10 Metastatic Melanoma Cells: A New Treatment Strategy. Antioxidants 2022, 11, 2166. [Google Scholar] [CrossRef]
- Arakawa, N.; Okubo, A.; Yasuhira, S.; Takahashi, K.; Amano, H.; Akasaka, T.; Masuda, T.; Shibazaki, M.; Maesawa, C. Carnosic acid, an inducer of NAD(P)H quinone oxidoreductase 1, enhances the cytotoxicity of beta-lapachone in melanoma cell lines. Oncol. Lett. 2018, 15, 2393–2400. [Google Scholar]
- Johnson, J.J. Carnosol: A promising anti-cancer and anti-inflammatory agent. Cancer Lett. 2011, 305, 1–7. [Google Scholar] [CrossRef]
- O’Neill, E.J.; Hartogh, D.J.D.; Azizi, K.; Tsiani, E. Anticancer Properties of Carnosol: A Summary of in Vitro and In Vivo Evidence. Antioxidants 2020, 9, 961. [Google Scholar] [CrossRef]
- Huang, S.C.; Ho, C.T.; Lin-Shiau, S.Y.; Lin, J.K. Carnosol inhibits the invasion of B16/F10 mouse melanoma cells by suppressing metalloproteinase-9 through down-regulating nuclear factor-kappa B and c-Jun. Biochem. Pharmacol. 2005, 69, 221–232. [Google Scholar] [CrossRef]
- Choi, S.M.; Kim, D.-H.; Chun, K.-S.; Choi, J.-S. Carnosol induces apoptotic cell death through ROS-dependent inactivation of STAT3 in human melanoma G361 cells. Appl. Biol. Chem. 2019, 62, 55. [Google Scholar] [CrossRef]
- Tong, L.; Wu, S. The Mechanisms of Carnosol in Chemoprevention of Ultraviolet B-Light-Induced Non-Melanoma Skin Cancer Formation. Sci. Rep. 2018, 8, 3574. [Google Scholar] [CrossRef]
- Huang, M.T.; Ho, C.T.; Wang, Z.Y.; Ferraro, T.; Lou, Y.R.; Stauber, K.; Ma, W.; Georgiadis, C.; Laskin, J.D.; Conney, A.H. Inhibition of skin tumorigenesis by rosemary and its constituents carnosol and ursolic acid. Cancer Res. 1994, 54, 701–708. [Google Scholar] [PubMed]
- Yeo, I.J.; Park, J.H.; Jang, J.S.; Lee, D.Y.; Park, J.E.; Choi, Y.E.; Joo, J.H.; Song, J.K.; Jeon, H.O.; Hong, J.T. Inhibitory effect of Carnosol on UVB-induced inflammation via inhibition of STAT3. Arch. Pharm. Res. 2019, 42, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Maczka, W.; Twardawska, M.; Grabarczyk, M.; Winska, K. Carvacrol-A Natural Phenolic Compound with Antimicrobial Properties. Antibiotics 2023, 12, 824. [Google Scholar] [CrossRef] [PubMed]
- Joshi, S.; Kundu, S.; Priya, V.V.; Kulhari, U.; Mugale, M.N.; Sahu, B.D. Anti-inflammatory activity of carvacrol protects the heart from lipopolysaccharide-induced cardiac dysfunction by inhibiting pyroptosis via NLRP3/Caspase1/Gasdermin D signaling axis. Life Sci. 2023, 324, 121743. [Google Scholar] [CrossRef]
- Cerrah, S.; Ozcicek, F.; Gundogdu, B.; Cicek, B.; Coban, T.A.; Suleyman, B.; Altuner, D.; Bulut, S.; Suleyman, H. Carvacrol prevents acrylamide-induced oxidative and inflammatory liver damage and dysfunction in rats. Front. Pharmacol. 2023, 14, 1161448. [Google Scholar] [CrossRef]
- Singh, J.; Luqman, S.; Meena, A. Carvacrol as a Prospective Regulator of Cancer Targets/Signalling Pathways. Curr. Mol. Pharmacol. 2023, 16, 542–558. [Google Scholar] [CrossRef] [PubMed]
- Govindaraju, S.; Arulselvi, P.I. Characterization of Coleus aromaticus essential oil and its major constituent carvacrol for in vitro antidiabetic and antiproliferative activities. J. Herbs Spices Med. 2018, 24, 37–51. [Google Scholar] [CrossRef]
- Osanloo, M.; Alipanah, H.; Farjam, M.; Taheri, A.; Zarenezhad, E. Anticancer Activity of Chitosan Nanoparticles Containing Satureja khuzistanica Essential Oil, and Carvacrol against Human Melanoma and Breast Cancer. Russ. J. Bioorg. 2023, 49, 594–601. [Google Scholar] [CrossRef]
- Nanni, V.; Di Marco, G.; Sacchetti, G.; Canini, A.; Gismondi, A. Oregano Phytocomplex Induces Programmed Cell Death in Melanoma Lines via Mitochondria and DNA Damage. Foods 2020, 9, 1486. [Google Scholar] [CrossRef]
- de Oliveira, I.C.V.; Galvao-Moreira, L.V.; Vilela, J.L.; Duarte-Silva, M.; Aguiar-da-Silva, L.D.; Pereira, C.A.A.; Pereira, D.M.S.; Pinheiro, A.; Lima-Neto, L.G.; Fernandes, E.S.; et al. Cinnamaldehyde modulates host immunoinflammatory responses in rat ligature-induced periodontitis and peripheral blood mononuclear cell models. Int. Immunopharmacol. 2023, 115, 109669. [Google Scholar] [CrossRef]
- Qureshi, K.A.; Mohammed, S.A.A.; Khan, O.; Ali, H.M.; El-Readi, M.Z.; Mohammed, H.A. Cinnamaldehyde-Based Self-Nanoemulsion (CA-SNEDDS) Accelerates Wound Healing and Exerts Antimicrobial, Antioxidant, and Anti-Inflammatory Effects in Rats’ Skin Burn Model. Molecules 2022, 27, 5225. [Google Scholar] [CrossRef]
- Zhang, G.; Li, T.; Liu, J.; Wu, X.; Yi, H. Cinnamaldehyde-Contained Polymers and Their Biomedical Applications. Polymers 2023, 15, 1517. [Google Scholar] [CrossRef] [PubMed]
- Cabello, C.M.; Bair, W.B.; Lamore, S.D.; Ley, S.; Bause, A.S.; Azimian, S.; Wondrak, G.T. The cinnamon-derived Michael acceptor cinnamic aldehyde impairs melanoma cell proliferation, invasiveness, and tumor growth. Free Radic. Biol. Med. 2009, 46, 220–231. [Google Scholar] [CrossRef]
- Zhou, L.; Lu, Y.; Yang, G.; Wu, J. Research on tumorigenicity of cinnamaldehyde in melanoma cell lines and its mechanism. Tumour. Biol. 2014, 35, 5717–5722. [Google Scholar] [CrossRef]
- Patra, K.; Jana, S.; Sarkar, A.; Mandal, D.P.; Bhattacharjee, S. The inhibition of hypoxia-induced angiogenesis and metastasis by cinnamaldehyde is mediated by decreasing HIF-1alpha protein synthesis via PI3K/Akt pathway. Biofactors 2019, 45, 401–415. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.P.; Li, Y.Q.; Lv, Y.T.; Wang, J.M. Effect of curcumin on the proliferation, apoptosis, migration, and invasion of human melanoma A375 cells. Genet. Mol. Res. 2015, 14, 1056–1067. [Google Scholar] [CrossRef] [PubMed]
- Marin, Y.E.; Wall, B.A.; Wang, S.; Namkoong, J.; Martino, J.J.; Suh, J.; Lee, H.J.; Rabson, A.B.; Yang, C.S.; Chen, S.; et al. Curcumin downregulates the constitutive activity of NF-kappaB and induces apoptosis in novel mouse melanoma cells. Melanoma Res. 2007, 17, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Siwak, D.R.; Shishodia, S.; Aggarwal, B.B.; Kurzrock, R. Curcumin-induced antiproliferative and proapoptotic effects in melanoma cells are associated with suppression of IkappaB kinase and nuclear factor kappaB activity and are independent of the B-Raf/mitogen-activated/extracellular signal-regulated protein kinase pathway and the Akt pathway. Cancer 2005, 104, 879–890. [Google Scholar] [PubMed]
- Zheng, M.; Ekmekcioglu, S.; Walch, E.T.; Tang, C.H.; Grimm, E.A. Inhibition of nuclear factor-kappaB and nitric oxide by curcumin induces G2/M cell cycle arrest and apoptosis in human melanoma cells. Melanoma Res. 2004, 14, 165–171. [Google Scholar] [CrossRef]
- Jiang, A.J.; Jiang, G.; Li, L.T.; Zheng, J.N. Curcumin induces apoptosis through mitochondrial pathway and caspases activation in human melanoma cells. Mol. Biol. Rep. 2015, 42, 267–275. [Google Scholar] [CrossRef]
- Bush, J.A.; Cheung, K.J., Jr.; Li, G. Curcumin induces apoptosis in human melanoma cells through a Fas receptor/caspase-8 pathway independent of p53. Exp. Cell Res 2001, 271, 305–314. [Google Scholar] [CrossRef]
- Liao, W.; Xiang, W.; Wang, F.F.; Wang, R.; Ding, Y. Curcumin inhibited growth of human melanoma A375 cells via inciting oxidative stress. Biomed. Pharmacother. 2017, 95, 1177–1186. [Google Scholar] [CrossRef]
- Kocyigit, A.; Guler, E.M. Curcumin induce DNA damage and apoptosis through generation of reactive oxygen species and reducing mitochondrial membrane potential in melanoma cancer cells. Cell Mol. Biol. 2017, 63, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Bakhshi, J.; Weinstein, L.; Poksay, K.S.; Nishinaga, B.; Bredesen, D.E.; Rao, R.V. Coupling endoplasmic reticulum stress to the cell death program in mouse melanoma cells: Effect of curcumin. Apoptosis 2008, 13, 904–914. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.; Han, X.; Zheng, S.; Li, Z.; Sha, Y.; Ni, J.; Sun, Z.; Qiao, S.; Song, Z. Curcumin induces autophagy, inhibits proliferation and invasion by downregulating AKT/mTOR signaling pathway in human melanoma cells. Oncol. Rep. 2016, 35, 1065–1074. [Google Scholar] [CrossRef]
- Chiu, Y.J.; Yang, J.S.; Tsai, F.J.; Chiu, H.Y.; Juan, Y.N.; Lo, Y.H.; Chiang, J.H. Curcumin suppresses cell proliferation and triggers apoptosis in vemurafenib-resistant melanoma cells by downregulating the EGFR signaling pathway. Environ. Toxicol. 2022, 37, 868–879. [Google Scholar] [CrossRef]
- Yu, T.; Ji, J.; Guo, Y.L. MST1 activation by curcumin mediates JNK activation, Foxo3a nuclear translocation and apoptosis in melanoma cells. Biochem. Biophys. Res. Commun. 2013, 441, 53–58. [Google Scholar] [CrossRef]
- Shimada, K.; Ushijima, K.; Suzuki, C.; Horiguchi, M.; Ando, H.; Akita, T.; Shimamura, M.; Fujii, J.; Yamashita, C.; Fujimura, A. Pulmonary administration of curcumin inhibits B16F10 melanoma lung metastasis and invasion in mice. Cancer Chemother. Pharmacol. 2018, 82, 265–273. [Google Scholar] [CrossRef]
- Sonavane, K.; Phillips, J.; Ekshyyan, O.; Moore-Medlin, T.; Roberts Gill, J.; Rong, X.; Lakshmaiah, R.R.; Abreo, F.; Boudreaux, D.; Clifford, J.L.; et al. Topical curcumin-based cream is equivalent to dietary curcumin in a skin cancer model. J. Skin Cancer 2012, 2012, 147863. [Google Scholar] [CrossRef]
- Phillips, J.; Moore-Medlin, T.; Sonavane, K.; Ekshyyan, O.; McLarty, J.; Nathan, C.A. Curcumin inhibits UV radiation-induced skin cancer in SKH-1 mice. Otolaryngol. Head Neck Surg. 2013, 148, 797–803. [Google Scholar] [CrossRef] [PubMed]
- Tsai, K.D.; Lin, J.C.; Yang, S.M.; Tseng, M.J.; Hsu, J.D.; Lee, Y.J.; Cherng, J.M. Curcumin Protects against UVB-Induced Skin Cancers in SKH-1 Hairless Mouse: Analysis of Early Molecular Markers in Carcinogenesis. Evid. Based Complement. Altern. Med. 2012, 2012, 593952. [Google Scholar] [CrossRef]
- Jensen, J.D.; Dunn, J.H.; Luo, Y.; Liu, W.; Fujita, M.; Dellavalle, R.P. Ellagic acid inhibits melanoma growth in vitro. Dermatol. Rep. 2011, 3, e36. [Google Scholar] [CrossRef]
- Wang, F.; Chen, J.; Xiang, D.; Lian, X.; Wu, C.; Quan, J. Ellagic acid inhibits cell proliferation, migration, and invasion in melanoma via EGFR pathway. Am. J. Transl. Res. 2020, 12, 2295–2304. [Google Scholar]
- Hseu, Y.C.; Chou, C.W.; Senthil Kumar, K.J.; Fu, K.T.; Wang, H.M.; Hsu, L.S.; Kuo, Y.H.; Wu, C.R.; Chen, S.C.; Yang, H.L. Ellagic acid protects human keratinocyte (HaCaT) cells against UVA-induced oxidative stress and apoptosis through the upregulation of the HO-1 and Nrf-2 antioxidant genes. Food Chem. Toxicol. 2012, 50, 1245–1255. [Google Scholar] [CrossRef]
- Lembo, S.; Balato, A.; Di Caprio, R.; Cirillo, T.; Giannini, V.; Gasparri, F.; Monfrecola, G. The modulatory effect of ellagic acid and rosmarinic acid on ultraviolet-B-induced cytokine/chemokine gene expression in skin keratinocyte (HaCaT) cells. Biomed. Res. Int. 2014, 2014, 346793. [Google Scholar] [CrossRef] [PubMed]
- Islam, S.U.; Ahmed, M.B.; Ahsan, H.; Islam, M.; Shehzad, A.; Sonn, J.K.; Lee, Y.S. An Update on the Role of Dietary Phytochemicals in Human Skin Cancer: New Insights into Molecular Mechanisms. Antioxidants 2020, 9, 916. [Google Scholar] [CrossRef]
- Zari, A.T.; Zari, T.A.; Hakeem, K.R. Anticancer Properties of Eugenol: A Review. Molecules 2021, 26, 7407. [Google Scholar] [CrossRef]
- Padhy, I.; Paul, P.; Sharma, T.; Banerjee, S.; Mondal, A. Molecular Mechanisms of Action of Eugenol in Cancer: Recent Trends and Advancement. Life 2022, 12, 1795. [Google Scholar] [CrossRef] [PubMed]
- Jaganathan, S.K.; Supriyanto, E. Antiproliferative and molecular mechanism of eugenol-induced apoptosis in cancer cells. Molecules 2012, 17, 6290–6304. [Google Scholar] [CrossRef]
- Kim, G.-C.; Choe, D.S.; Im, J.-S.; Jeong, H.; Kim, I.; Lee, M.-H.; Park, B.-S. Caspases-dependent Apoptosis in Human Melanoma Cell by Eugenol. Anat. Cell Biol. 2006, 39, 245–253. [Google Scholar]
- Ghosh, R.; Nadiminty, N.; Fitzpatrick, J.E.; Alworth, W.L.; Slaga, T.J.; Kumar, A.P. Eugenol causes melanoma growth suppression through inhibition of E2F1 transcriptional activity. J. Biol. Chem. 2005, 280, 5812–5819. [Google Scholar] [CrossRef] [PubMed]
- Ra Choi, B.-B.; Shin, S.-H.; Kim, U.-K.; Hong, J.-W.; Kim, G.-C. S Phase Cell Cycle Arrest and Apoptosis is Induced by Eugenol in G361 Human Melanoma Cells. Int. J. Oral Biol. 2011, 36, 129–134. [Google Scholar]
- Júnior, P.L.; Câmara, D.A.; Costa, A.S.; Ruiz, J.L.; Levy, D.; Azevedo, R.A.; Pasqualoto, K.F.; de Oliveira, C.F.; de Melo, T.C.; Pessoa, N.D.; et al. Apoptotic effect of eugenol envolves G2/M phase abrogation accompanied by mitochondrial damage and clastogenic effect on cancer cell in vitro. Phytomedicine 2016, 23, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Valizadeh, A.; Khaleghi, A.A.; Alipanah, H.; Zarenezhad, E.; Osanloo, M. Anticarcinogenic Effect of Chitosan Nanoparticles Containing Syzygium aromaticum Essential Oil or Eugenol Toward Breast and Skin Cancer Cell Lines. BioNanoScience 2021, 11, 678–686. [Google Scholar] [CrossRef]
- Pal, D.; Banerjee, S.; Mukherjee, S.; Roy, A.; Panda, C.K.; Das, S. Eugenol restricts DMBA croton oil induced skin carcinogenesis in mice: Downregulation of c-Myc and H-ras, and activation of p53 dependent apoptotic pathway. J. Dermatol. Sci. 2010, 59, 31–39. [Google Scholar] [CrossRef]
- Sukumaran, K.; Unnikrishnan, M.C.; Kuttan, R. Inhibition of tumour promotion in mice by eugenol. Indian J. Physiol. Pharmacol. 1994, 38, 306–308. [Google Scholar]
- Kaur, G.; Athar, M.; Alam, M.S. Eugenol precludes cutaneous chemical carcinogenesis in mouse by preventing oxidative stress and inflammation and by inducing apoptosis. Mol. Carcinog. 2010, 49, 290–301. [Google Scholar] [CrossRef]
- Kalmes, M.; Hennen, J.; Blömeke, B. Eugenol and isoeugenol as antiproliferative agents in skin cancer cells. Toxicol. Lett. 2009, 189, S100. [Google Scholar] [CrossRef]
- Chrubasik, S.; Pittler, M.H.; Roufogalis, B.D. Zingiberis rhizoma: A comprehensive review on the ginger effect and efficacy profiles. Phytomedicine 2005, 12, 684–701. [Google Scholar] [CrossRef] [PubMed]
- Ganjikunta, V.S.; Maddula, R.R.; Bhasha, S.; Sahukari, R.; Kondeti Ramudu, S.; Chenji, V.; Kesireddy, S.R.; Zheng, Z.; Korivi, M. Cardioprotective Effects of 6-Gingerol against Alcohol-Induced ROS-Mediated Tissue Injury and Apoptosis in Rats. Molecules 2022, 27, 8606. [Google Scholar] [CrossRef]
- de Lima, R.M.T.; Dos Reis, A.C.; de Menezes, A.P.M.; Santos, J.V.O.; Filho, J.; Ferreira, J.R.O.; de Alencar, M.; da Mata, A.; Khan, I.N.; Islam, A.; et al. Protective and therapeutic potential of ginger (Zingiber officinale) extract and [6]-gingerol in cancer: A comprehensive review. Phytother. Res. 2018, 32, 1885–1907. [Google Scholar] [CrossRef] [PubMed]
- Aloliqi, A.A. Therapeutic Potential of 6-Gingerol in Prevention of Colon Cancer Induced by Azoxymethane through the Modulation of Antioxidant Potential and Inflammation. Curr. Issues Mol. Biol. 2022, 44, 6218–6228. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.C.; Min, J.K.; Kim, T.Y.; Lee, S.J.; Yang, H.O.; Han, S.; Kim, Y.M.; Kwon, Y.G. [6]-Gingerol, a pungent ingredient of ginger, inhibits angiogenesis in vitro and in vivo. Biochem. Biophys. Res. Commun. 2005, 335, 300–308. [Google Scholar] [CrossRef]
- Nigam, N.; Bhui, K.; Prasad, S.; George, J.; Shukla, Y. [6]-Gingerol induces reactive oxygen species regulated mitochondrial cell death pathway in human epidermoid carcinoma A431 cells. Chem. Biol. Interact. 2009, 181, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.O.; Kundu, J.K.; Shin, Y.K.; Park, J.H.; Cho, M.H.; Kim, T.Y.; Surh, Y.J. [6]-Gingerol inhibits COX-2 expression by blocking the activation of p38 MAP kinase and NF-kappaB in phorbol ester-stimulated mouse skin. Oncogene 2005, 24, 2558–2567. [Google Scholar] [CrossRef]
- Kim, J.K.; Kim, Y.; Na, K.M.; Surh, Y.J.; Kim, T.Y. [6]-Gingerol prevents UVB-induced ROS production and COX-2 expression in vitro and in vivo. Free Radic. Res. 2007, 41, 603–614. [Google Scholar] [CrossRef]
- Ju, S.A.; Park, S.M.; Lee, Y.S.; Bae, J.H.; Yu, R.; An, W.G.; Suh, J.H.; Kim, B.S. Administration of 6-gingerol greatly enhances the number of tumor-infiltrating lymphocytes in murine tumors. Int. J. Cancer 2012, 130, 2618–2628. [Google Scholar] [CrossRef] [PubMed]
- Park, K.K.; Chun, K.S.; Lee, J.M.; Lee, S.S.; Surh, Y.J. Inhibitory effects of [6]-gingerol, a major pungent principle of ginger, on phorbol ester-induced inflammation, epidermal ornithine decarboxylase activity and skin tumor promotion in ICR mice. Cancer Lett. 1998, 129, 139–144. [Google Scholar] [CrossRef]
- Bischoff-Kont, I.; Furst, R. Benefits of Ginger and Its Constituent 6-Shogaol in Inhibiting Inflammatory Processes. Pharmaceuticals 2021, 14, 571. [Google Scholar] [CrossRef]
- Wu, H.; Hsieh, M.C.; Lo, C.Y.; Liu, C.B.; Sang, S.; Ho, C.T.; Pan, M.H. 6-Shogaol is more effective than 6-gingerol and curcumin in inhibiting 12-O-tetradecanoylphorbol 13-acetate-induced tumor promotion in mice. Mol. Nutr. Food Res. 2010, 54, 1296–1306. [Google Scholar] [CrossRef]
- Chen, F.; Tang, Y.; Sun, Y.; Veeraraghavan, V.P.; Mohan, S.K.; Cui, C. 6-shogaol, a active constiuents of ginger prevents UVB radiation mediated inflammation and oxidative stress through modulating NrF2 signaling in human epidermal keratinocytes (HaCaT cells). J. Photochem. Photobiol. B 2019, 197, 111518. [Google Scholar] [CrossRef]
- Krause, S.T.; Liao, P.; Crocoll, C.; Boachon, B.; Forster, C.; Leidecker, F.; Wiese, N.; Zhao, D.; Wood, J.C.; Buell, C.R.; et al. The biosynthesis of thymol, carvacrol, and thymohydroquinone in Lamiaceae proceeds via cytochrome P450s and a short-chain dehydrogenase. Proc. Natl. Acad. Sci. USA 2021, 118, e2110092118. [Google Scholar] [CrossRef] [PubMed]
- Rathod, N.B.; Kulawik, P.; Ozogul, F.; Regenstein, J.M.; Ozogul, Y. Biological activity of plant-based carvacrol and thymol and their impact on human health and food quality. Trends Food Sci. 2021, 116, 733–748. [Google Scholar] [CrossRef]
- Satooka, H.; Kubo, I. Effects of thymol on B16-F10 melanoma cells. J. Agric. Food Chem. 2012, 60, 2746–2752. [Google Scholar] [CrossRef]
- Calo, R.; Visone, C.M.; Marabini, L. Thymol and Thymus Vulgaris L. activity against UVA- and UVB-induced damage in NCTC 2544 cell line. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2015, 791, 30–37. [Google Scholar] [CrossRef]
- Mapelli, M.; Calò, R.; Marabini, L. Thymol and Thymus vulgaris extract protects human keratinocyte cell line (HaCaT) from UVA and UVB damage. Oxid Antioxid. Med. Sci. 2016, 5, 39. [Google Scholar] [CrossRef]
- Cornaghi, L.; Arnaboldi, F.; Calo, R.; Landoni, F.; Baruffaldi Preis, W.F.; Marabini, L.; Donetti, E. Effects of UV Rays and Thymol/Thymus vulgaris L. Extract in an ex vivo Human Skin Model: Morphological and Genotoxicological Assessment. Cells Tissues Organs 2016, 201, 180–192. [Google Scholar] [CrossRef] [PubMed]



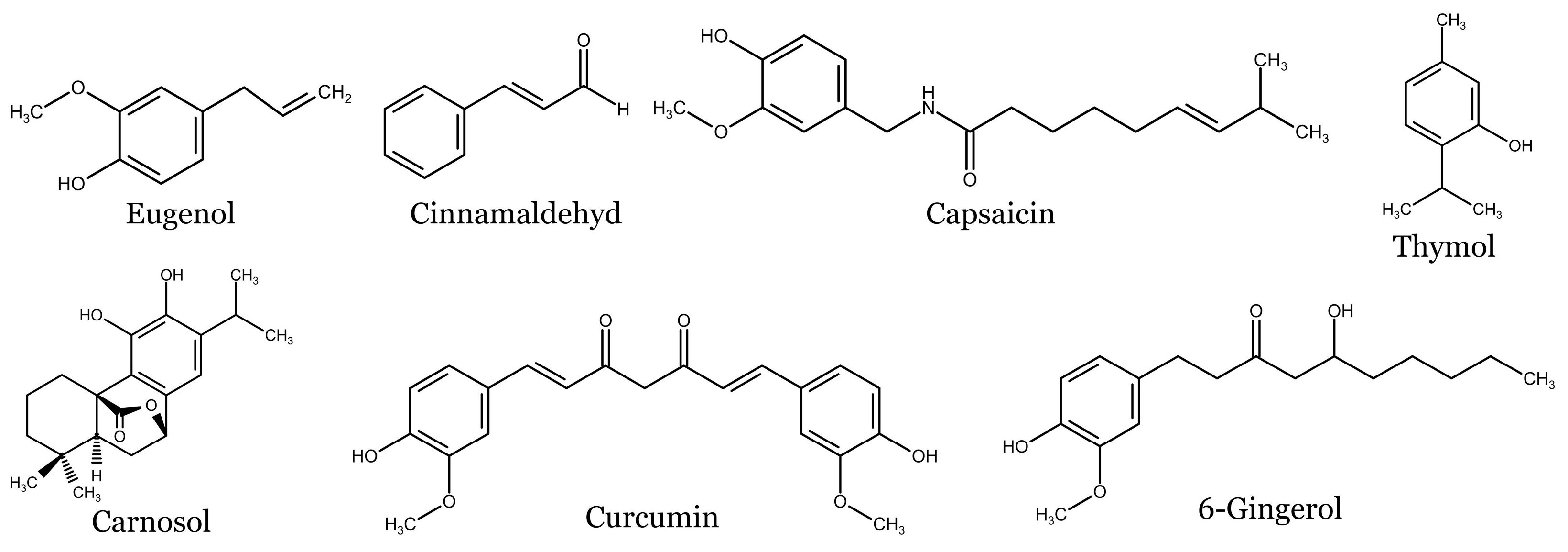

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spice | Phenolic Compounds | References |
|---|---|---|
| Allspice | Flavonoids: quercetin, quercitrin, kaempferol, and catechin, naringenin Phenolic acids: syringic acid, caffeic acid, coumaric acid, and cinnamic acid Phenylpropanoids: eugenol, methyl eugenol, isoeugenol, and chavicol Tannins: vascalaginone and grandininol | [65,66,67,68] |
| Alpinia galanga | Chalcones: galanganones A–C Flavonoids: galangin, galangin-3-methylether, and kaempferol, quercetin, isorhamnetin, apigenin, kumatakenin, and pinocembrin Lignans: galanganal and galanganol A–C Phenolic acids: ferulic acid and gallic acid Phenylpropanoids: 1′-acetoxychavicol acetate, and methyleugenol Tannins: ellagic acid | [72,73,74] |
| Black cumin | Flavonoids: kaempferol, kaempferol-3-glucoside, quercetin, quercitrin, and diosmin Monoterpenes: p-cymene Phenolic acids: caftaric acid, p-hydroxybenzoic acid, syringic acid, protocatechuic acid, and chlorogenic acid Phenylpropanoids: thymohydroquinone, thymol, carvacrol, and t-anethole | [81,82,83] |
| Black pepper | Alkaloids: piperine Terpenes: β-caryophyllene, limonene, β-pinene, α-pinene, sabinene, camphene, linalool, and terpinene-4-ol Phenolic acids: hydroxybenzoic acid, gallic acid, caffeic acid, and hydroxycinnamic acids Flavonoids: quercetin, catechin, epicatechin, myricetin kaempferol, isoquercetin, and isorhamnetin | [84,85,86] |
| Cinnamon | Phenylpropanoids: eugenol Phenolic acid: caffeic acid, chlorogenic acid, gallic, rosmarinic acid, p-coumaric acid, protocatechuic acid, p-hydroxybenzoic acid, and trans-vanillic acid Flavonoids: rutin, apigenin, catechin, and epicatechin Other compounds: cinnamaldehyde, cinnamyl alcohol, cinnamyl acetate, terpenes, and terpenoids | [97,98,99,100] |
| Coriander | Phenolic acids: caffeic acid, chlorogenic acid, ferulic acid, gallic acid, o-coumaric acid, trans-hydroxycinnamic acid, p-coumaric acid, rosmarinic acid, salicylic acid, trans-cinnamic acid, and vanillic acid Flavonoids: rutin, luteolin quercetin, kaempferol, naringin, apigenin, and diosmin Other compounds: esculin, esculetin, catechin, orientin, and maleic acid | [103,104,105,106,107] |
| Fenugreek | Flavonoids: quercetin, luteolin, vitexin, isovitexin, kaempferol, tricin, and naringenin Phenolic acid: p-coumaric acid, caffeic acid, and chlorogenic acid | [118,119,120,121] |
| Ginger | Flavonoids: quercetin Phenolic compounds: gingerols (6-gingerol, 8-gingerol, and 10-gingerol), shogalols (6-shogaol, 8-shogaol, and 10-shogaol), gingerenone-A, zingerone, 8-paradol, and 6-dehydrogingerdione | [128,129,130,131,132,133] |
| Oregano | Flavonoids: rutin, naringin, hesperetin, naringenin, apigenin, luteolin, acacetin, and vitexin Phenolic acids: caffeinic acid and rosmarinic acid Other compounds: carvacrol and thymol | [136,137] |
| Nutmeg | Phenolic acids: protocatechuic acid, caffeic acid, vanillic acid, p-coumaric acid, ferulic acid, and sinapic acid Flavonoids: catechin, epicatechin, rutin, quercitrin, isoquercitrin, quercetin, and kaempferol Other phenolic compounds: ellagic acid, myristicin, and elemicin | [146,147] |
| Red chili | Alkaloids: capsaicinoids (capsaicin, dihydrocapsaicin, nordihydrocapsaicin, homodihydrocapsaicin, and homocapsaicin) Flavonoids: quercetin, luteolin, kaempferol, myricetin, apigenin, naringenin, catechin, and epigallocatechin Phenolic acids: gallic acid, protocatechuic acid, vanillic acid, hydroxyl cinnamic acids, caffeic acid, ferulic acid, chlorogenic acid, and cinnamic acid | [149,150,151,152,153] |
| Rosemary | Phenolic acids: salvianic acid, caffeic acid, rosmarinic acid, and salvianic acid A Flavonoids: luteolin, luteolin−7-O-rutinoxide, luteolin-7-glucoronide, hesperidin, apigenin, cirsimaritin, genkwanin, and salvigenin Terpenes: rosmadial, 7-methylrosmanol, carnosol, carnosic acid, and 12-methylcarnosic acid | [158,159] |
| Saffron | Alkaloids: pyridin-3-ylmethanol, harman, and tribulusterine Phenolic acids: protocatechuic acid, 4-hydroxybenzoic acid, vanillic acid benzoic acid, and p-coumaric acid Flavonoids: kaempferol, kaempferide, kaempferol-3-O-sophoroside-7-O-glucoside, kaempferol-3,7,4′-triglucoside, kaempferol 7-O-β-D-glucopyranoside, isorhamnetin-3,4′-diglucoside, isorhamnetin-3-O-glucoside, astragalin, sophoraflavonoloside, and helichrysoside | [164] |
| Sichuan pepper | Polyphenols: isovitexin, vitexin, hyperoside, isoquercitrin, rutin, foeniculin, trifolin, quercitrin, astragalin, and afzelin | [169] |
| Star anise | Phenylpropanoids: cis- and trans-anethole, estragole, anisylacetone, ρ-anisaldehyde, and foeniculin | [177] |
| Sumac | Tannins: galloylhexose, O-galloylmorbergenin, and O-galloyl arbutin Flavonoids: quercetin, myrecetin, apigenin, kaempferol, and quercetin 3-glucoside Phenolic acids: gallic acid, ellagic acid, and protocatechuic acid Anthocyanins: delphidin-3-glucoside, cyanidin 3-(2″-galloyl)galactoside, and cyanidin-3-glucoside | [193] |
| Thyme | Monoterpenes and terpenoids: thymol and carvacrol Phenolic acids: caffeic acid, quinic acid, p-coumaric acid, ferulic acid, cinnamic acid, and rosmarinic acid Flavonoids: apigenin, naringnin, and luteolin-7-o-rutinose | [199] |
| Turmeric | Curcuminoids: curcumin, demethoxycurcumin, and bisdemethoxycurcumin Phenolic acids: gallic acid, protocatechuic acid, chlorogenic acid, cinnamic acid, and ferulic acid Flavonoids: rutin, myricetin, genistein, quercetin, catechin, and epicatechin | [202] |
| Phenolic Acids | Mechanism of Action | References |
|---|---|---|
| Caffeic acid | Inhibition of proliferation and colony formation, cell cycle arrest, induction of apoptosis, reduction of cell viability ↓ Erk1/2 signaling pathway, phospho- p90RSK, phospho-c-Myc, phospho-Elk1, COX-2, AP-1, Nf-кB, Fyn, DNA damage, ROS generation, PI3K, Akt, phospho-JNK, phospho-p38, ↑ PTEN, expression of caspase -1, -3, -8 | [209,213,214,215] |
| Chlorogenic acid | Induction of apoptosis, inhibition of viability of tumor cells ↓ TGFβ, SOD1, SOD2, GPX1 ↑ activity of caspase -3 | [216,217,218] |
| p-Coumaric acid | Inhibition of cell viability, proliferation and colony formation, S phase cell cycle arrest, induction of apoptosis, modulation of TME ↓ cyclins A and E, Cdk2, Bcl-2 ↑ cleaved caspase -3, -9, Apaf-1, cytochrome c release, Bax | [225,226] |
| Ferulic acid | Induction of apoptosis, reduction of colony formation, modulation of angiogenesis and inflammation, reduction of UVB-induced DNA damage ↓ MMP-2, MMP-9, iNOS, IL-6, TNF-α, VEGF, Bcl-2 ↑ Bax | [229] |
| Gallic acid | Induction of apoptosis, modulation of glycolysis, inhibition of migration and invasion, S phase cell cycle arrest, ROS generation ↓ MMP, MMP-2, Ras, Erk1/2, PI3K, p38-MAPK, Akt, Bcl-2, GADD153, Bcl-2, Bcl-xL, Mcl-1, ↑ activity capsase -3, -8, -9, Bid, cytochrome c release, GRP78, Fas, FasL, cleaved PARP, AIF, Bad, Bax, VDAC-1, SOD, G-3-P DHase, glucokinase, enolase, aldolase, ATPase | [232,233,234] |
| Protocatechuic acid | Modulation of inflammation and oxidative stress, inhibition of migration, invasion and metastasis ↓ MMP-2, Ras, phospho-Akt, nuclear NF-кB, PI3K, phospho-Akt, RhoA, Cdc42, Rac1 ↑ RhoB, TIMP-2 | [236,237,238,239] |
| Rosmarinic acid | Decrease of tumor cells viability, induction of apoptosis, inhibition of proliferation, invasion and migration ↓ expression of NLRP3 inflammasome, ADAM17, EGF, phospsho-AKT, phospho-GSK3β, MMP-2, MMP-9, Bcl-2 ↑ caspase -3, -8, Bax | [240,241,242,243] |
| Syringic acid | Inhibition of UVB-induced carcinogenesis ↓ COX-2, MMP-1, MMP-13, prostaglandin E2, phospho-EGFR, phospho-Erk1/2, phospho-JNK1/2, phospho-p38 MAPK, phospho-MEK1/2, phospho-MKK4/7, phospho-MKK3/6, phospho-B-Raf, phospho-Akt, phospho-Src | [248] |
| Flavonoids | Mechanism of Action | References |
|---|---|---|
| Apigenin | Reduction of viability, migration, invasion, angiogenesis and inflammation, induction of apoptosis, S and G2/M phase cell cycle arrest, reduction of Ki-67 positive cells ↓ phospho-Erk1/2, phospho-Akt, Akt, JNK, phospho-mTOR, phospho-STAT3, MMP-2, MMP-9, VEGF, Twist1, PD-L1, phospho-Src, phospho-JAK2, N-cadherin, fibronectin, Bcl-2, IL-6, MCP-1, IL-12, IL-10, COX2, Cdk2, phospho-p70S6K, phospho-p90RSK, phospho-MSK, phospho-4E-BP1, phospho-AMPK, phospho-CREB, USF-2, phospho-MEK1/2, phospho-MKK4, phospho-MKK3/6, phospho-EGFR, phospho-c-Raf ↑ E-cadherin, keratin-8, Bax, cleaved PARP, cleaved caspase -9, p53, LC3-II | [251,252,253,254,255,256,257,258] |
| Astragalin | Induction of DNA fragmentation and apoptosis, decrease of viability and G2/M cell cycle arrest, modulation of immune response ↓ cyclin D1, Mcl-1, SOX10, procaspase -9 ↑ Bax, cleaved caspase -3, cleaved PARP | [262,263] |
| Catechin | Inhibition of cell proliferation, induction of apoptosis and cell cycle arrest, inhibition of autophagy, decrease of Ki-67 positive cells ↓ Bcl-2, Beclin-1, LC3-I/II, phospho-AMPK, PD-L1/PD-L2 expression, phospsho-STAT3, STAT, IRF1, Sirt3 ↑ caspase -3 activation, phospho-PI3K, phospho-Akt, phospho-mTOR | [267,268] |
| Chrysin | Inhibited of cell proliferation, migration and invasion, G2/M cell cycle arrest and induction of apoptosis, modulation of immune response ↓ MMP-2, MMP-9, VEGF, PI3K, phospho-Akt, PKC, phospho-FAK, RhoA, phospho-c-Jun, N-cadherin, GRB2, Ras, NF-кB, phospho-Erk1/2, cytosolic Bax ↑ E-cadherin, caspase-3 activation, PBG-D, phospho-p38, mitochondial Bax | [269,270,271] |
| Diosmin | Induction of apoptosis, ROS generation and DNA fragmentation, inhibition of invasion, migration and cell growth ↓ Bcl-2, MMP-2, MMP-9 ↑ p53, caspase -3, -9 | [273,274,275] |
| Galangin | Inhibition of colony formation, cell motility, adhesion and migration, reduction of Ki-67 positive cells, induction of apoptosis and autophagy ↓ phospho-FAK, FAK, phospho-Akt, phospho-Erk1/2 ↑ GCLC, GSS, phopsho-Nrf2, Nrf2 | [276,277,279] |
| Genistein | Inhibition of cell proliferation, growth, viability and migration, S and G2/M phase cell cycle arrest, ROS generation and induction of ER stress-mediated apoptosis ↓ Cdk1, phospho-FAK, phospho-paxilin, phospho-p38, phospho-Erk1/2, phosho-JNK, tensin-2, vinculin, α-actinin, cyclin E and B, Cdc25C, Bcl-2, Bcl-xL ↑ activation of Chk2, p21, p27, p53, APAF | [207,283,284,285,286,287,288,289] |
| Isoquercitrin | Inhibition of viability and clonogenicity, DNA fragmentation, G1/S cell cycle arrest, induction of apoptosis ↓ Bcl-2, procaspase -8, -9, phospho-PI3K, phospho-Akt, phospho-mTOR ↑ Bax, AIF, Endo G, cleaved PARP | [297] |
| Isorhamnetin | Induction of apoptosis, inhibition of cell proliferation, growth, migration and colony formation, G2/M cell cycle arrest ↓ phospho-Akt, nuclear NF-кB, COX-2, phospho-Erk1/2, Bcl-2, COX-2, phosphpo-MEK1/2, phospho-Erk1/2, phospho-p90RSK ↑ activation of caspase -3, Bax | [299,300] |
| Kaempferol | Inhibition of cell proliferation, growth, migration and invasion, induction of apoptosis, G2/M cell cycle arrest ↓ phospho-PI3K, phospho-Akt, phospho-mTOR, activity of hexokinase, VDAC1, Akt/GSK-3β pathway, COX-2, phospho-Erk1/2, phospho-p38, phospho-JNK, Src activity | [303,304,305,306] |
| Luteolin | Suppression of cell proliferation, migration and invasion, induction of DNA damage and apoptosis, ER stress and ROS generation, G0/G1 and G2/M cell cycle arrest ↓ phospho-Akt, phospho-PI3K, MMP-2, MMP-9, HIF-1α, VEGF-A, VEGFR-2, MMP-2, MMP-9, N-cadherin, vimentin, integrin β3, STAT3, ZEB1, phospho-FAK, phospho-Src, Src ↑ TIMP-1, TIMP-2, E-cadherin, PERK, eIF-2α, ATF6, CHOP, cleaved caspase -12 | [308,310,311,312,313,314] |
| Myricetin | Inhibition of cell proliferation, colony formation, invasion, migration, inflammation and angiogenesis, induction of apoptosis and ROS generation ↓ COX-2, Fin kinase activity, Bcl-2, MMP, NF-кB, phospho-Erk1/2, phospho-38, phospho-JNK, phospho-p90RSK, phospho-MSK, phospho-MEK, MEK1, phospho- Raf, phospho-JAK1, AP-1, c-Fos, STAT3, ↑ Bax, p53, GADD45, caspase -3, -8, -9 | [319,320,321,322,323,324] |
| Naringenin | Induction of apoptosis, DNA fragmentation, ROS generation and mitochondrial damage, suppression of proliferation, migration, metastasis, tube formation and angiogenesis ↓ phospho-Erk1/2, phospho-JNK, Tie2, TPC2 activity, ↑ caspase -3 activation, cleavage of PARP, transglutaminase activity | [325,326,328,329] |
| Quercetin | Reduction of cell viability, proliferation, colony formation, migration and invasion, induction of DNA fragmentation and ROS generation, cell cycle arrest, inhibition of EMT ↓ MMP, Bcl-2, PKC-α, STAT3 expression, phosphorylation and nuclear localization, phospho-Src, phospho-JAK2, Mcl-1, Bcl-xL, MMP-2, MMP-9, VEGF, PKC, N-cadherin, vimentin, fibronectin, Twist, Snail, S100A7, phospho-Met, FAS, phospho-Fak, Fak, Gab1, phospho-Pak1, Nrf-2, catalase, B-Raf, phospho-MEK, phospho-Erk1/2, phospho-Akt, PI3K ↑ phospho-JNK, phospho-p38, E-cadherin, VCAM1, ICAM1, cleaved PARP, Bax, cleaved caspase -3, -8, Bim, AIF, RIG-I, STAT1, IRF7 | [334,336,337,338,339,340,341,342,343,345,346] |
| Rutin | Decrease of cell viability, nuclear fragmentation, induction of apoptosis ↓ COX-2, iNOS, phospho-STAT3, AP-1, phospho-JNK ↑ β-galactosidase, p38 MAPK | [353,354,355] |
| Vitexin | Induction of apoptosis, DNA damage, ROS generation and oxidative stress G2/M cell phase cell cycle arrest, inhibition of colony formation ↓ Bcl-2, Cdk1, Cdk6, cyclins A2 and E2, MMP-2, MMP-9, vimentin, Slug, Twist, phospho-Src, phospho-JAK1/2, Chk2 ↑ Bax, cleaved PARP, phospho-ATM, phospho-ATR, GADD45, p21, PUMA, phospho-Chk2, p53, γ-H2AX | [356,357] |
| Other Phenolic Compounds | Mechanism of Action | References |
|---|---|---|
| 1′-acetoxychavicol acetate | Inhibition of ROS production and lipid peroxidation ↓ NF-кB | [358,359] |
| trans-Anethole | Inhibition of cell proliferation, colony formation, induction of apoptosis, modulation of MiR-498/STAT4 axis ↑ miR-498 | [360,363] |
| Arbutin | Induction of mitochondrial dysfunction ↓ MMP, Bcl-2, Bxl-xL, vimentin, HSP90, α-enolase, Inosine-5′-monophosphate dehydrogenase 2, Peroxiredoxin-1 ↑ p53, VDAC-1, 14-3-3G | [365,366] |
| Capsaicin | Inhibition of cell growth, proliferation, migration, induction of intrinsic and extrinsic apoptosis and autophagy and DNA damage, ROS and RNS generation ↓ phospho-PI3K, phospho-Akt, phospho-mTOR, Rac1, Bcl-2, tNOX, MDM2, procaspase -3, -8, -9, SOX2, EZH2, Sirt1 ↑ p53 activation, Bax, DR4, Fas, PARP cleavage, caspase -3 activation, ATG5, ATG7, Beclin-1, LC3-I/II, iNOS, cytochrome c release, | [368,369,370,371,372,373] |
| Carnosic acid | Inhibition of growth, proliferation, migration, adhesion and colony formation, G0/G1 cell cycle arrest and induction of apoptosis ↓ p27, MMP-2, MMP-9, TIMP-1, uPA, VCAM-1, N-cadherin, vimentin, Snail, Slug, phospho-Src, phospho-Fak, phospho-Akt ↑ p21, E-cadherin, TIMP-2 | [375,376,377] |
| Carnosol | Inhibition of cell growth, proliferation, invasiveness and colony formation, induction of ROS production, DNA damage and apoptosis ↓ MMP-2, MMP-9, phospho-Erk1/2, phospho-p38, phospho-JNK, phospho-Akt, NF-кB and c-Jun nuclear translocation, phospho-Src, phospho-STAT3, cyclin D1, D2, D3, survivin, phospho-γH2AX, phospho-Chk1, iNOS, COX-2, TNF-α, IL-1β, TIMP-2, Bcl-2, Bcl-xL, MDM2, ↑ p53, cleaved caspase -3, -7, -9, cleaved PARP, Bax | [381,382,385] |
| Carvacrol | Suppression of cell proliferation, growth, induction of apoptosis and necroptosis, G2/M cell cycle arrest, ROS production, DNA fragmentation and mitochondrial damage ↓ Bcl-2, TOMM20, procaspase -3, ↑ cleavage of PARP, Bax, cytochrome c, p53, γH2AX | [390,392] |
| Cinnamaldehyde | Inhibition of tumor cell proliferation, migration, invasion and angiogenesis, G1 phase cell cycle arrest, induction of ROS production and apoptosis ↓ HIF-α, VEGF, PI3K, phospho-Akt, phospho-mTOR, MMP-2, MMP-9, IL-8, phospho-Flk1, CD31, vimentin, Zeb1, Twist ↑ caspase -3 activation, HO-1, p21, | [396,397,398] |
| Curcumin | Inhibition of cell survival, viability, proliferation, migration, invasion, induction of ROS generation, DNA damage, oxidative stress, ER stress, apoptosis and autophagy, G2/M cell cycle arrest ↓ MMP-2, MMP-9, phospho-JAK2, phospho-STAT3, activity of NF-кB, phospho-Erk1/2, iNOS, phospho-IκBα, p38, COX-2, PGE2, NO, cyclin D1, Bcl-2, Mcl-1, MMP, DNA-PKcs, XIAP, phospho-Akt, phospho-mTOR, phospho-P70S6K, phospho-S6, phospho-4EBP1, phospho-Rb, G6PD, GSH ↑ Bax, activation of caspases -3, -7, -8, -9, -12, TIMP2, phospho-p53, p53, p21, p27, Chk2, γH2AX, cleaved PARP, Fas, phospho-JNK, HLJ1, GRP94, GRP78, phospho-EIF2α, GADD153, cytochrome c release, HIF-1α, Bim1, Foxo3 nuclear translocation, MST1 | [399,400,401,402,403,404,405,406,407,408,410,411,412,414] |
| Ellagic acid | Induction of ROS generation and apoptosis, suppression of proliferation, migration and invasion, G1 phase cell cycle arrest ↓ NF-кB, phospho-EGFR, β-catenin, Keap1, vimentin, IL-1, IL-8, TNF-α, ↑ HO-1, SOD, Nrf2, IL-10, E-cadherin, | [415,416,417,418] |
| Eugenol | Inhibition of tumor cell proliferation and colony formation, induction of DNA fragmentation, apoptosis, G1 and G2/M phase cell cycle arrest ↓ MMP, cyclin A, D3, E, Cdk2, Cdk4, cdc2, E2F1, c-Myc, H-Ras, Bcl-2, ODC activity, COX-2, iNOS, IL-6, TNF-α, PGE2, NF-κB/p65 translocation ↑ activation of caspase -3, -6, -7 and lamin A, PARP and DFF45 cleavage, AIF and cytochrome c release, Bax, p53, p21, | [423,424,425,426,428,430,431] |
| 6-gingerol | G1 cell cycle arrest, ROS generation and induction of apoptosis, modulation of immune response, inhibition of angiogenesis ↓ MMP, Bcl-2, COX-2, NF-κB activation, cyclin D1, E, A, phospho-p38, phospho-IκBα, phospho-ATF2, LPO ↑ Bax, caspase -3, -9, cytochrome c, Apaf-1, | [436,437,438,439,440] |
| 6-shogaol | Inhibition of cell survival, proliferation, induction ↓ iNOS, COX-2, NF-κB nuclear translocation, phospho-IκBα, phospho-Erk1/2, phospho-p38 MAPK, phospho-PI3K, phospho-Akt, phospho-JNK1/2, IL-6, IL-10, TNF-α, HO-1 ↑ Nrf2 | [443,444] |
| Thymol | Reduction of UVA and UVB-induced DNA damage and oxidative stress | [447,448] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baloghová, J.; Michalková, R.; Baranová, Z.; Mojžišová, G.; Fedáková, Z.; Mojžiš, J. Spice-Derived Phenolic Compounds: Potential for Skin Cancer Prevention and Therapy. Molecules 2023, 28, 6251. https://doi.org/10.3390/molecules28176251
Baloghová J, Michalková R, Baranová Z, Mojžišová G, Fedáková Z, Mojžiš J. Spice-Derived Phenolic Compounds: Potential for Skin Cancer Prevention and Therapy. Molecules. 2023; 28(17):6251. https://doi.org/10.3390/molecules28176251
Chicago/Turabian StyleBaloghová, Janette, Radka Michalková, Zuzana Baranová, Gabriela Mojžišová, Zuzana Fedáková, and Ján Mojžiš. 2023. "Spice-Derived Phenolic Compounds: Potential for Skin Cancer Prevention and Therapy" Molecules 28, no. 17: 6251. https://doi.org/10.3390/molecules28176251
APA StyleBaloghová, J., Michalková, R., Baranová, Z., Mojžišová, G., Fedáková, Z., & Mojžiš, J. (2023). Spice-Derived Phenolic Compounds: Potential for Skin Cancer Prevention and Therapy. Molecules, 28(17), 6251. https://doi.org/10.3390/molecules28176251