
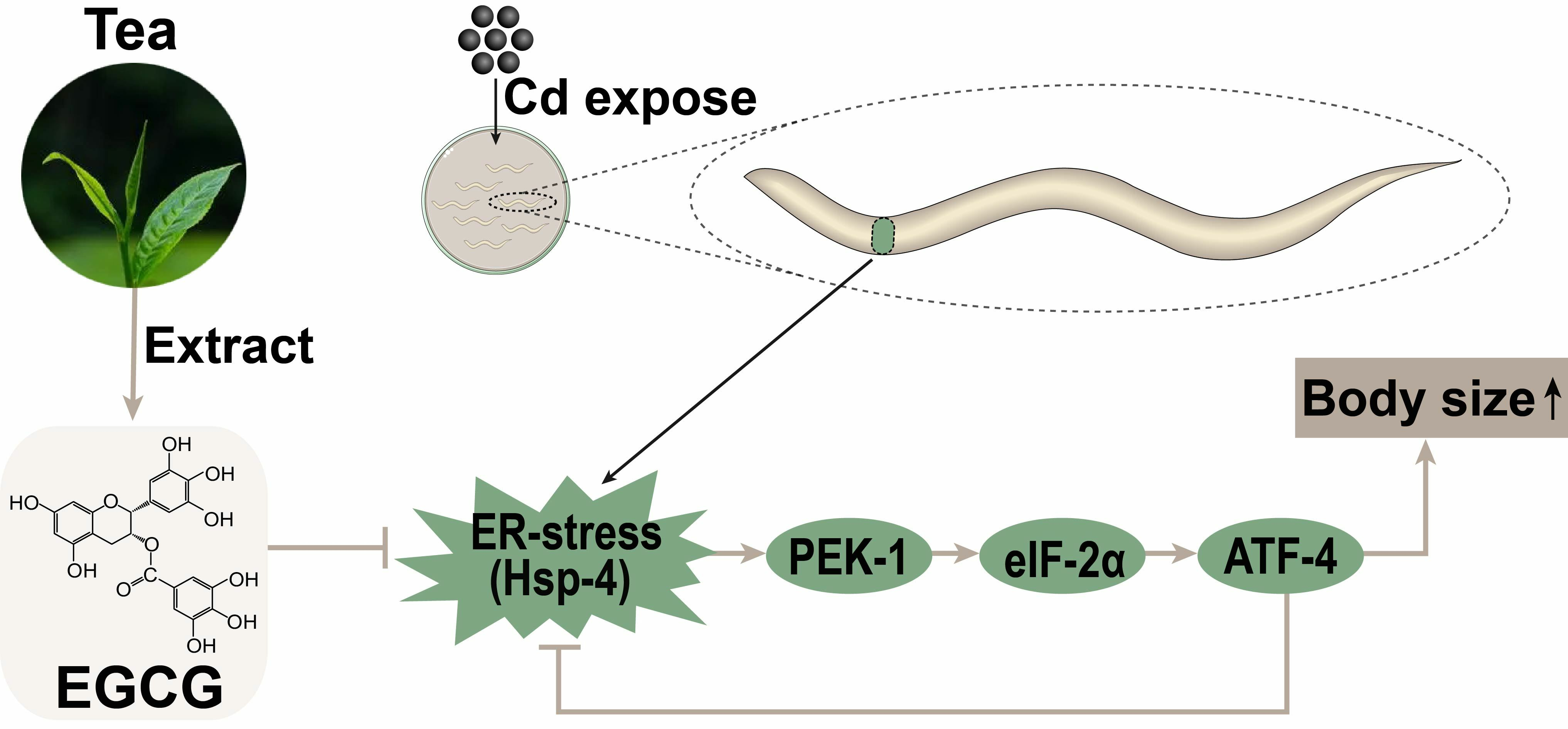
Epigallocatechin-3-Gallate Reduces Cd-Induced Developmental Toxicity of Bodysize in Caenorhabditis elegans via the PEK-1/eIF-2α/ATF-4 Pathway

Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Result
2.1. EGCG Alleviates Cd-Induced Body Size Developmental Toxicity in C. elegans
2.2. Detoxification of Body Size Developmental Toxicity by EGCG Is Not Related to Its Chelating Effect
2.3. EGCG Regulates ER Stress under Cd Exposure
2.4. EGCG Attenuates Cd-Induced Body Size Developmental Toxicity by Regulating ER Stress
2.5. EGCG Regulates ER Stress via the Pek-1/eif-2α/atf-4 Pathway
2.6. EGCG Reduces Cd-Induced Body Size Developmental Toxicity in Nematodes via the Pek-1/eif-2α/atf-4 Pathway
3. Discussion
4. Materials and Methods
4.1. C. elegans Strains and Maintenance
4.2. C. elegans Food
4.3. Drugs and Treatment
4.4. C. elegans Body Size Estimation
4.5. Analysis of C. elegans Development
4.6. Onset of Egg Laying
4.7. Lifespan Experiments
4.8. Pharyngeal Pumping Rate
4.9. UV–vis Spectra of EGCG
4.10. Free EGCG Content Assay
4.11. Accumulation and Efflux of Cd in Nematodes
4.12. RNAi
4.13. Determination of hsp-4p::GFP Expression Levels
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Genchi, G.; Sinicropi, M.S.; Lauria, G.; Carocci, A.; Catalano, A. The Effects of Cadmium Toxicity. Int. J. Environ. Res. Public Health 2020, 17, 3782. [Google Scholar] [CrossRef]
- Satarug, S.; Gobe, G.C.; Vesey, D.A. Multiple Targets of Toxicity in Environmental Exposure to Low-Dose Cadmium. Toxics 2022, 10, 472. [Google Scholar] [CrossRef]
- Souza-Arroyo, V.; Fabian, J.J.; Bucio-Ortiz, L.; Miranda-Labra, R.U.; Gomez-Quiroz, L.E.; Gutierrez-Ruiz, M.C. The mechanism of the cadmium-induced toxicity and cellular response in the liver. Toxicology 2022, 480, 153339. [Google Scholar] [CrossRef] [PubMed]
- Gundacker, C.; Hengstschlager, M. The role of the placenta in fetal exposure to heavy metals. Wien. Med. Wochenschr. (1946) 2012, 162, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Chandravanshi, L.; Shiv, K.; Kumar, S. Developmental toxicity of cadmium in infants and children: A review. Environ. Anal. Health Toxicol. 2021, 36, e2021003. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Chen, W.; Wang, D.; Jin, Y.; Chen, X.; Xu, Y. The effects of prenatal exposure to low-level cadmium, lead and selenium on birth outcomes. Chemosphere 2014, 108, 33–39. [Google Scholar] [CrossRef]
- Dursun, A.; Yurdakok, K.; Yalcin, S.S.; Tekinalp, G.; Aykut, O.; Orhan, G.; Morgil, G.K. Maternal risk factors associated with lead, mercury and cadmium levels in umbilical cord blood, breast milk and newborn hair. J. Matern. Fetal Neonatal Med. 2016, 29, 954–961. [Google Scholar] [CrossRef]
- Doelling, R.; Mendelski, M.N.; Paul, R.J. Bacterial diet and weak cadmium stress affect the survivability of Caenorhabditis elegans and its resistance to severe stress. Heliyon 2019, 5, e01126. [Google Scholar] [CrossRef]
- Hirota, K.; Matsuoka, M. N-acetylcysteine restores the cadmium toxicity of Caenorhabditis elegans. BioMetals 2021, 34, 1207–1216. [Google Scholar] [CrossRef]
- Mu, Y.; Hu, X.; Yang, P.; Sun, L.; Gu, W.; Zhang, M. The effects of cadmium on the development of Drosophila and its transgenerational inheritance effects. Toxicology 2021, 462, 152931. [Google Scholar] [CrossRef]
- Saedi, S.; Shirazi, M.R.J.; Zamiri, M.J.; Totonchi, M.; Dadpasand, M.; Sedaghati, F. Impaired follicular development and endocrine disorders in female rats by prepubertal exposure to toxic doses of cadmium. Toxicol. Ind. Health 2020, 36, 63–75. [Google Scholar] [CrossRef]
- Yang, C.S.; Chen, T.; Ho, C.T. Redox and Other Biological Activities of Tea Catechins That May Affect Health: Mechanisms and Unresolved Issues. J. Agric. Food Chem. 2022, 70, 7887–7899. [Google Scholar] [CrossRef]
- Chen, L.; Mo, H.; Zhao, L.; Gao, W.; Wang, S.; Cromie, M.M.; Lu, C.; Wang, J.S.; Shen, C.L. Therapeutic properties of green tea against environmental insults. J. Nutr. Biochem. 2017, 40, 1–13. [Google Scholar] [CrossRef]
- Roychoudhury, S.; Agarwal, A.; Virk, G.; Cho, C.L. Potential role of green tea catechins in the management of oxidative stress-associated infertility. Reprod. Biomed. Online 2017, 34, 487–498. [Google Scholar] [CrossRef]
- Hou, R.-R.; Chen, J.-Z.; Chen, H.; Kang, X.-G.; Li, M.-G.; Wang, B.-R. Neuroprotective effects of (-)-epigallocatechin-3-gallate (EGCG) on paraquat-induced apoptosis in PC12 cells. Cell Biol. Int. 2008, 32, 22–30. [Google Scholar] [CrossRef]
- Tai, K.-K.; Truong, D.D. (-)-Epigallocatechin-3-gallate (EGCG), a green tea polyphenol, reduces dichlorodiphenyl-trichloroethane (DDT)-induced cell death in dopaminergic SHSY-5Y cells. Neurosci. Lett. 2010, 482, 183–187. [Google Scholar] [CrossRef] [PubMed]
- Hegde, S.; Poojary, K.K.; Rasquinha, R.; Crasta, D.N.; Gopalan, D.; Mutalik, S.; Siddiqui, S.; Adiga, S.K.; Kalthur, G. Epigallocatechin-3-gallate (EGCG) protects the oocytes from methyl parathion-induced cytoplasmic deformities by suppressing oxidative and endoplasmic reticulum stress. Pestic. Biochem. Physiol. 2020, 167, 104588. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Du, L.; Li, J.; Song, H. Epigallocatechin-3-gallate attenuates cadmium-induced chronic renal injury and fibrosis. Food Chem. Toxicol. 2016, 96, 70–78. [Google Scholar] [CrossRef] [PubMed]
- An, Z.; Qi, Y.; Huang, D.; Gu, X.; Tian, Y.; Li, P.; Li, H.; Zhang, Y. EGCG inhibits Cd2+-induced apoptosis through scavenging ROS rather than chelating Cd2+ in HL-7702 cells. Toxicol. Mech. Methods 2014, 24, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Shen, K.; Ji, D. Natural compounds attenuate heavy metal-induced PC12 cell damage. J. Int. Med. Res. 2020, 48, 0300060520930847. [Google Scholar] [CrossRef]
- Bondad, S.E.C.; Kurasaki, M. Analysis of Cadmium, Epigallocatechin Gallate, and Vitamin C Co-exposure on PC12 Cellular Mechanisms. Biol. Trace Elem. Res. 2020, 198, 627–635. [Google Scholar] [CrossRef]
- Long, N.P.; Kang, J.S.; Kim, H.M. Caenorhabditis elegans: A model organism in the toxicity assessment of environmental pollutants. Environ. Sci. Pollut. Res. Int. 2023, 30, 39273–39287. [Google Scholar] [CrossRef]
- Xiao, X.; Zhang, X.; Zhang, C.; Li, J.; Zhao, Y.; Zhu, Y.; Zhang, J.; Zhou, X. Toxicity and multigenerational effects of bisphenol S exposure to Caenorhabditis elegans on developmental, biochemical, reproductive and oxidative stress. Toxicol. Res. 2019, 8, 630–640. [Google Scholar] [CrossRef]
- Langie, S.A.S.; Lara, J.; Mathers, J.C. Early determinants of the ageing trajectory. Best Pract. Res. Clin. Endocrinol. Metab. 2012, 26, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Xu, L.; Liang, Q.; Sun, Q.; Chen, C.; Zhang, Y.; Ding, Y.; Zhou, P. Metal chelator EGCG attenuates Fe(III)-induced conformational transition of alpha-synuclein and protects AS-PC12 cells against Fe(III)-induced death. J. Neurochem. 2017, 143, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Tamas, M.J.; Fauvet, B.; Christen, P.; Goloubinoff, P. Misfolding and aggregation of nascent proteins: A novel mode of toxic cadmium action in vivo. Curr. Genet. 2018, 64, 177–181. [Google Scholar] [CrossRef] [PubMed]
- Taylor, R.C.; Dillin, A. XBP-1 Is a Cell-Nonautonomous Regulator of Stress Resistance and Longevity. Cell 2013, 153, 1435–1447. [Google Scholar] [CrossRef] [PubMed]
- Frakes, A.E.; Dillin, A. The UPRER: Sensor and Coordinator of Organismal Homeostasis. Mol. Cell 2017, 66, 761–771. [Google Scholar] [CrossRef]
- Popham, J.D.; Webster, J.M. Cadmium toxicity in the free-living nematode, Caenorhabditis elegans. Environ. Res. 1979, 20, 183–191. [Google Scholar] [CrossRef]
- Duran-Izquierdo, M.; Taboada-Alquerque, M.; Sierra-Marquez, L.; Alvarez-Ortega, N.; Stashenko, E.; Olivero-Verbel, J. Hydroalcoholic extract of Haematoxylum brasiletto protects Caenorhabditis elegans from cadmium-induced toxicity. BMC Complement. Med. Ther. 2022, 22, 184. [Google Scholar] [CrossRef]
- Li, L.-L.; Wu, Q.-Z.; Chen, J.-J.; Cui, Y.-H.; Zhu, C.-M.; Li, B.-Y.; Guo, Z.; Li, W.-W. Mechanical Insights into Thiol-Mediated Synergetic Biotransformation of Cadmium and Selenium in Nematodes. Environ. Sci. Technol. 2021, 55, 7531–7540. [Google Scholar] [CrossRef]
- Abib, R.T.; Peres, K.C.; Barbosa, A.M.; Peres, T.V.; Bernardes, A.; Zimmermann, L.M.; Quincozes-Santos, A.; Fiedler, H.D.; Leal, R.B.; Farina, M.; et al. Epigallocatechin-3-gallate protects rat brain mitochondria against cadmium-induced damage. Food Chem. Toxicol. 2011, 49, 2618–2623. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.-N.; Shen, S.-R.; Yin, J.-J. Effects of interactions of EGCG and Cd2+ on the growth of PC-3 cells and their mechanisms. Food Chem. Toxicol. 2007, 45, 244–249. [Google Scholar] [CrossRef]
- Alhafez, M.; Kheder, F.; Aljoubbeh, M. Synthesis, characterization and antioxidant activity of EGCG complexes with copper and zinc ions. J. Coord. Chem. 2019, 72, 2337–2350. [Google Scholar] [CrossRef]
- Zwolak, I. Epigallocatechin Gallate for Management of Heavy Metal-Induced Oxidative Stress: Mechanisms of Action, Efficacy, and Concerns. Int. J. Mol. Sci. 2021, 22, 4027. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Tang, Y.; Li, Z.; Hua, Q.; Wang, L.; Song, X.; Zou, B.; Ding, M.; Zhao, J.; Tang, C. Joint Toxicity of a Multi-Heavy Metal Mixture and Chemoprevention in Sprague Dawley Rats. Int. J. Environ. Res. Public Health 2020, 17, 1451. [Google Scholar] [CrossRef]
- Cui, Y.; Zhou, X.; Chen, L.; Tang, Z.; Mo, F.; Li, X.C.; Mao, H.; Wei, X.; Wang, C.; Wang, H. Crosstalk between Endoplasmic Reticulum Stress and Oxidative Stress in Heat Exposure-Induced Apoptosis Is Dependent on the ATF4-CHOP-CHAC1 Signal Pathway in IPEC-J2 Cells. J. Agric. Food Chem. 2021, 69, 15495–15511. [Google Scholar] [CrossRef]
- Kapulkin, W.J.; Hiester, B.G.; Link, C.D. Compensatory regulation among ER chaperones in C. elegans. FEBS Lett. 2005, 579, 3063–3068. [Google Scholar] [CrossRef]
- Zhou, L.; He, J.N.; Du, L.; Ho, B.M.; Ng, D.S.-C.; Chan, P.P.P.; Tham, C.C.; Pang, C.P.; Chu, W.K. Epigallocatechin-3-Gallate Protects Trabecular Meshwork Cells from Endoplasmic Reticulum Stress. Oxid. Med. Cell. Longev. 2022, 2022, 7435754. [Google Scholar] [CrossRef] [PubMed]
- Boonyong, C.; Vardhanabhuti, N.; Jianmongkol, S. Modulation of non-steroidal anti-inflammatory drug-induced, ER stress-mediated apoptosis in Caco-2 cells by different polyphenolic antioxidants: A mechanistic study. J. Pharm. Pharmacol. 2020, 72, 1574–1584. [Google Scholar] [CrossRef]
- Xiang, C.; Xiao, X.; Jiang, B.; Zhou, M.; Zhang, Y.; Li, H.; Hu, Z. Epigallocatechin-3-gallate protects from high glucose induced podocyte apoptosis via suppressing endoplasmic reticulum stress. Mol. Med. Rep. 2017, 16, 6142–6147. [Google Scholar] [CrossRef] [PubMed]
- Durocher, F.; Faure, R.; Labrie, Y.; Pelletier, L.; Bouchard, I.; Laframboise, R. A novel mutation in the EIF2AK3 gene with variable expressivity in two patients with Wolcott-Rallison syndrome. Clin. Genet. 2006, 70, 34–38. [Google Scholar] [CrossRef] [PubMed]
- Yuan, P.; Pan, L.Y.; Xiong, L.G.; Tong, J.W.; Li, J.; Huang, J.A.; Gong, Y.S.; Liu, Z.H. Black tea increases hypertonic stress resistance in C. elegans. Food Funct. 2018, 9, 3798–3806. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Song, B.; Jia, X.; Yin, H.; Li, R.; Liu, X.; Chen, J.; Zhang, J.; Wang, Z.; Zhong, S. Ceramides from Sea Red Rice Bran Improve Health Indicators and Increase Stress Resistance of Caenorhabditis elegans through Insulin/IGF-1 Signaling (IIS) Pathway and JNK-1. J. Agric. Food Chem. 2022, 70, 15080–15094. [Google Scholar] [CrossRef]
- Ke, J.P.; Yu, J.Y.; Gao, B.; Hu, F.L.; Xu, F.Q.; Yao, G.; Bao, G.H. Two new catechins from Zijuan green tea enhance the fitness and lifespan of Caenorhabditis elegans via insulin-like signaling pathways. Food Funct. 2022, 13, 9299–9310. [Google Scholar] [CrossRef]
- Yasuda, M.; Matsuda, C.; Ohshiro, A.; Inouye, K.; Tabata, M. Effects of metal ions (Cu2+, Fe2+ and Fe3+) on HPLC analysis of catechins. Food Chem. 2012, 133, 518–525. [Google Scholar] [CrossRef]
- Wu, X.; Al-Amin, M.; Zhao, C.; An, F.; Wang, Y.; Huang, Q.; Teng, H.; Song, H. Catechinic acid, a natural polyphenol compound, extends the lifespan of Caenorhabditis elegans via mitophagy pathways. Food Funct. 2020, 11, 5621–5634. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, S.; Chen, C.; Lu, Y. Epigallocatechin-3-Gallate Reduces Cd-Induced Developmental Toxicity of Bodysize in Caenorhabditis elegans via the PEK-1/eIF-2α/ATF-4 Pathway. Molecules 2023, 28, 6344. https://doi.org/10.3390/molecules28176344
Wang S, Chen C, Lu Y. Epigallocatechin-3-Gallate Reduces Cd-Induced Developmental Toxicity of Bodysize in Caenorhabditis elegans via the PEK-1/eIF-2α/ATF-4 Pathway. Molecules. 2023; 28(17):6344. https://doi.org/10.3390/molecules28176344
Chicago/Turabian StyleWang, Shuanghui, Chuhong Chen, and Yan Lu. 2023. "Epigallocatechin-3-Gallate Reduces Cd-Induced Developmental Toxicity of Bodysize in Caenorhabditis elegans via the PEK-1/eIF-2α/ATF-4 Pathway" Molecules 28, no. 17: 6344. https://doi.org/10.3390/molecules28176344
APA StyleWang, S., Chen, C., & Lu, Y. (2023). Epigallocatechin-3-Gallate Reduces Cd-Induced Developmental Toxicity of Bodysize in Caenorhabditis elegans via the PEK-1/eIF-2α/ATF-4 Pathway. Molecules, 28(17), 6344. https://doi.org/10.3390/molecules28176344