
Effects of the Novel LaPLa-Enriched Medium- and Long-Chain Triacylglycerols on Body Weight, Glycolipid Metabolism, and Gut Microbiota Composition in High Fat Diet-Fed C57BL/6J Mice

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
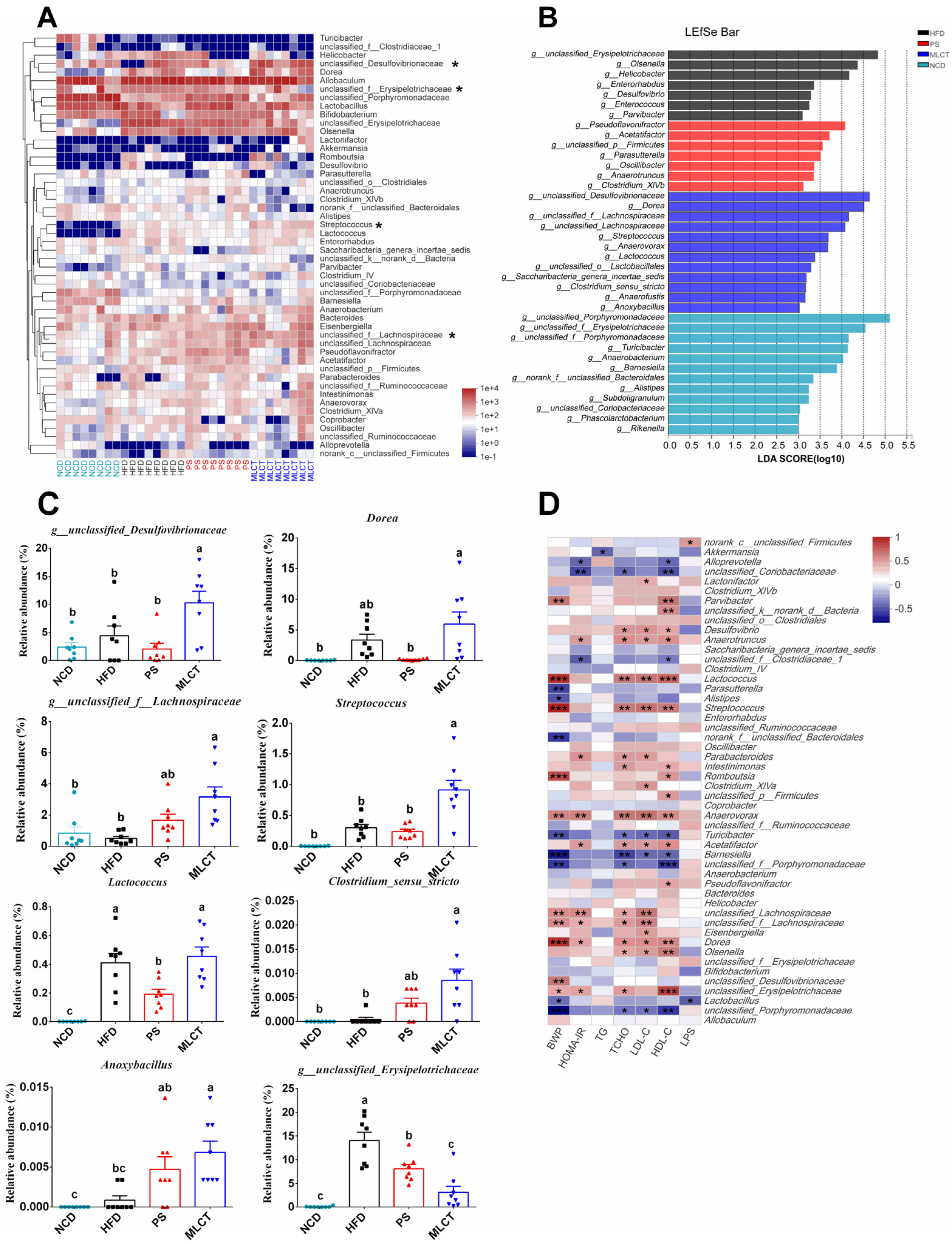
2. Results
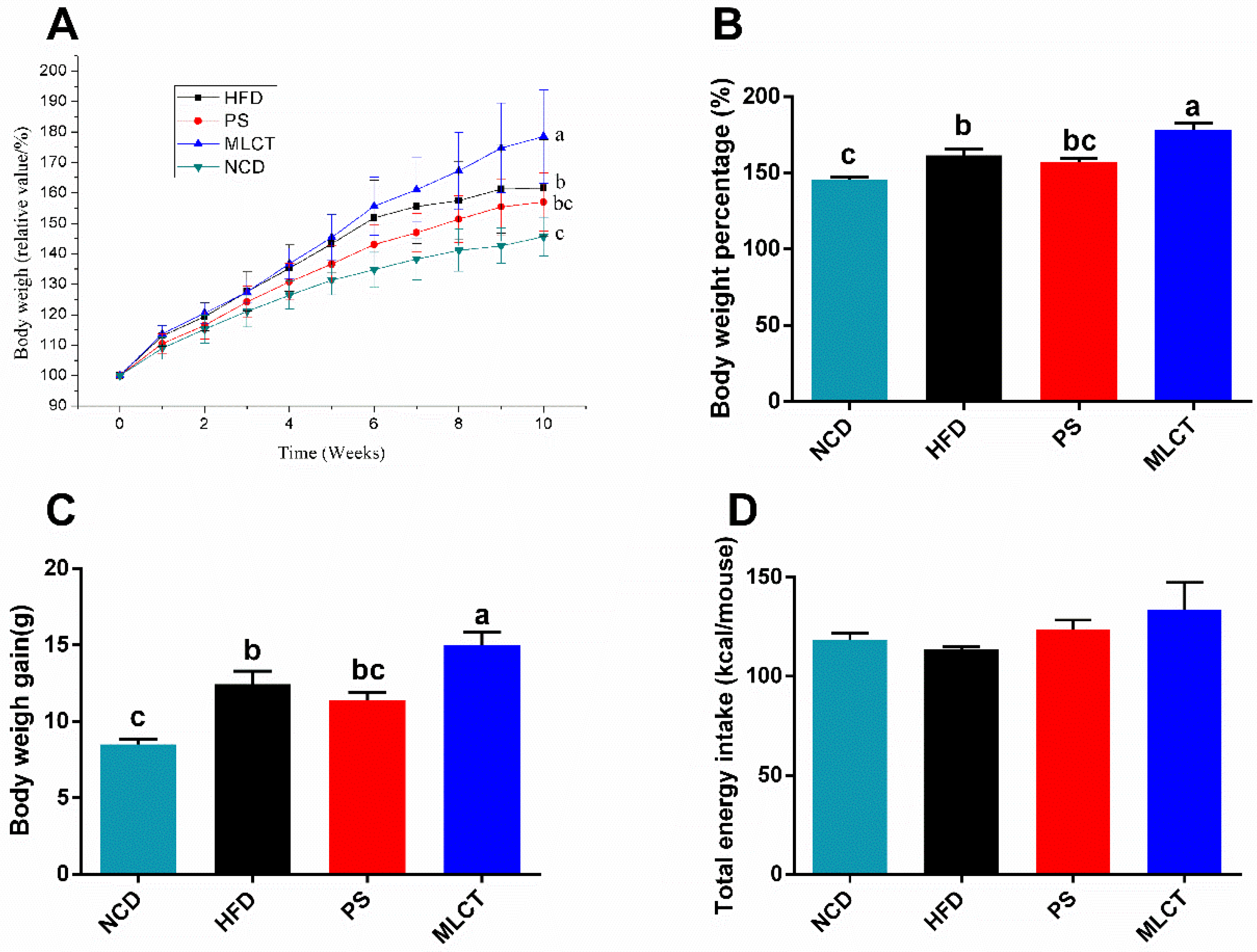
2.1. Body Weight Gain and Energy Intake
2.2. Organ Indexes and Morphological Changes
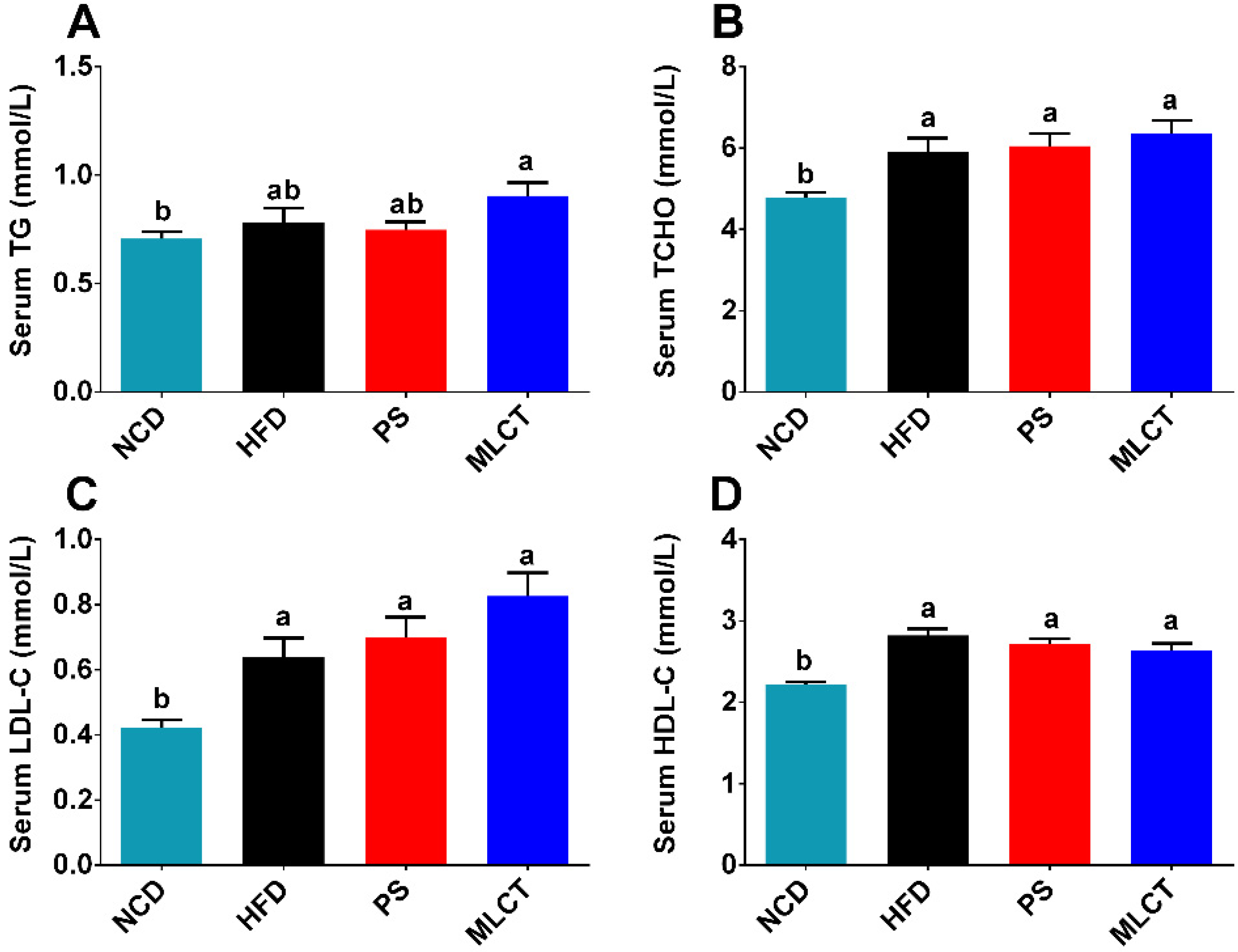
2.3. Serum Lipids
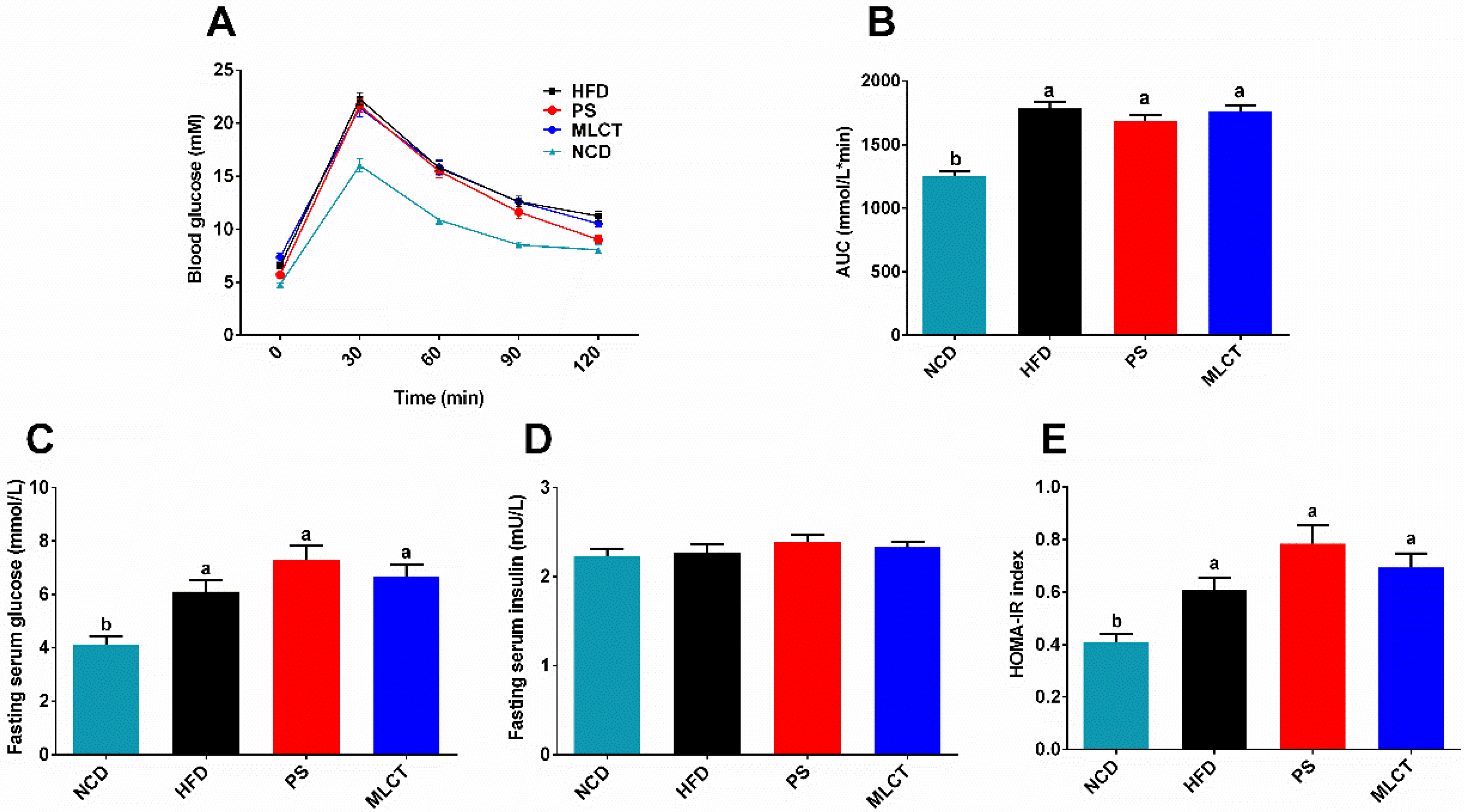
2.4. Glucose Homeostasis
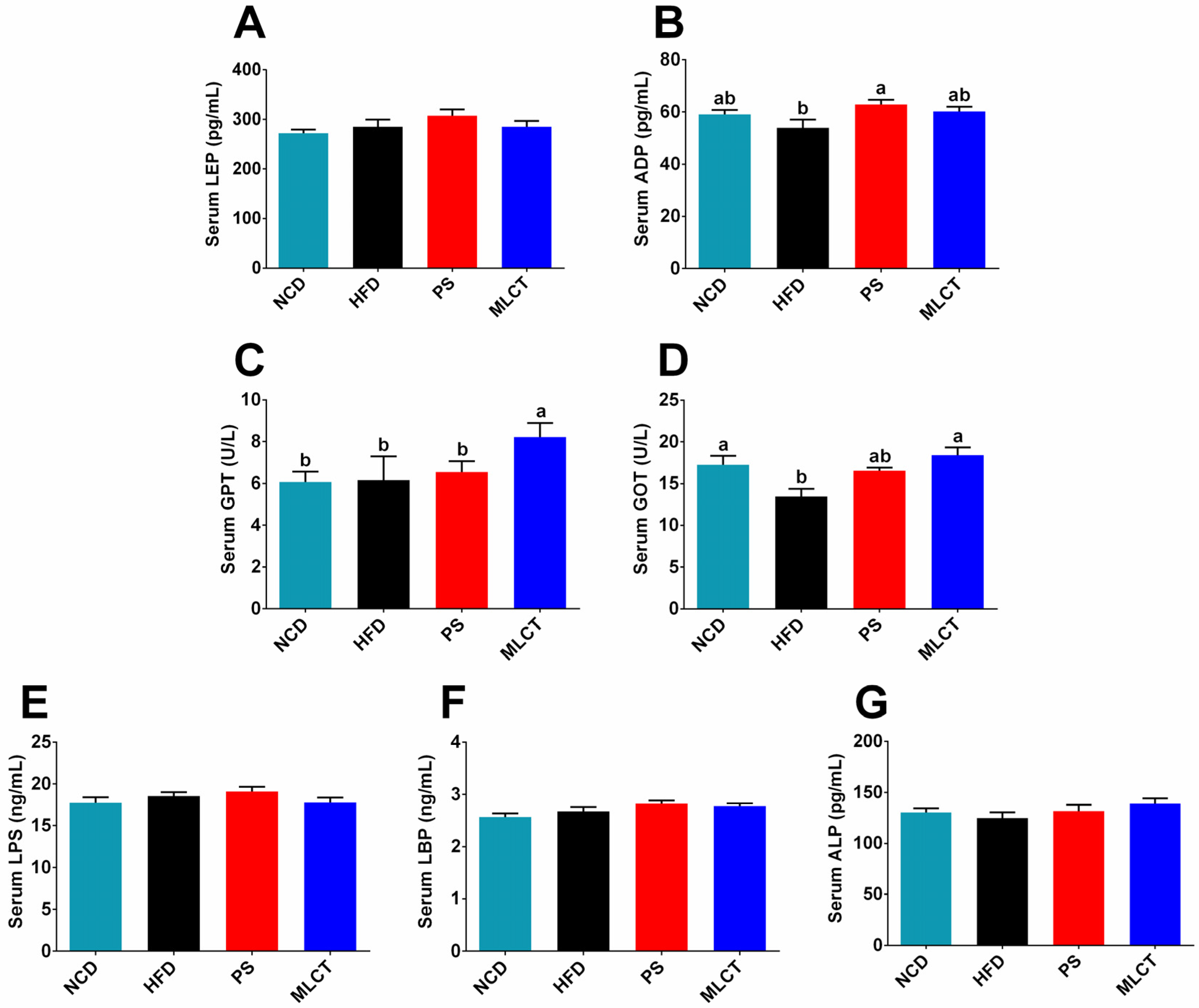
2.5. The Effect of MLCT on Hormones, Inflammatory Response, and Endotoxin in Mice
2.6. Organ Indexes and Morphological Changes
3. Discussion
4. Materials and Methods
4.1. Materials and Chemicals
4.2. Animal Experiments Design
4.3. Intraperitoneal Injection of Glucose Tolerance Test
4.4. Determination of Serum Biochemical Indexes
4.5. Morphological Analysis of Tissue Sections
4.6. 16S rRNA Analysis of the Gut Microbiota
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guo, Z.; Vikbjerg, A.F.; Xu, X. Enzymatic modification of phospholipids for functional applications and human nutrition. Biotechnol. Adv. 2005, 23, 203–259. [Google Scholar] [CrossRef]
- Ifeduba, E.A.; Akoh, C.C. Modification of Stearidonic Acid Soybean Oil by Immobilized Rhizomucor miehei Lipase to Incorporate Caprylic Acid. J. Am. Oil Chem. Soc. 2014, 91, 953–965. [Google Scholar] [CrossRef]
- Mu, H. The digestion of dietary triacylglycerols. Prog. Lipid Res. 2004, 43, 105–133. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Tang, T.K.; Phuah, E.T.; Karim, N.A.A.; Alitheen, N.B.M.; Tan, C.P.; Razak, I.S.A.; Lai, O.M. Structural difference of palm based Medium- and Long-Chain Triacylglycerol (MLCT) further reduces body fat accumulation in DIO C57BL/6J mice when consumed in low fat diet for a mid-term period. Food Res. Int. 2018, 103, 200–207. [Google Scholar] [CrossRef]
- Mu, H.; Porsgaard, T. The metabolism of structured triacylglycerols. Prog. Lipid Res. 2005, 44, 430–448. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Tang, T.K.; Chan, E.S.; Phuah, E.T.; Lai, O.M.; Tan, C.P.; Wang, Y.; Ab Karim, N.A.; Mat Dian, N.H.; Tan, J.S. Medium chain triglyceride and medium-and long chain triglyceride: Metabolism, production, health impacts and its applications—A review. Crit. Rev. Food Sci. Nutr. 2021, 65, 4169–4185. [Google Scholar] [CrossRef]
- Kelly, T.; Yang, W.; Chen, C.S.; Reynolds, K.; He, J. Global burden of obesity in 2005 and projections to 2030. Int. J. Obes. 2008, 32, 1431–1437. [Google Scholar] [CrossRef] [Green Version]
- St-Onge, M.P.; Mayrsohn, B.; O’Keeffe, M.; Kissileff, H.R.; Choudhury, A.R.; Laferrere, B. Impact of medium and long chain triglycerides consumption on appetite and food intake in overweight men. Eur. J. Clin. Nutr. 2014, 68, 1134–1140. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Tang, T.K.; Lai, O.M. Health benefits, enzymatic production, and application of medium- and long-chain triacylglycerol (MLCT) in food industries: A review. J. Food Sci. 2012, 77, R137–R144. [Google Scholar] [CrossRef]
- Formiguera, X.; Canton, A. Obesity: Epidemiology and clinical aspects. Best Pract. Res. Clin. Gastroenterol. 2004, 18, 1125–1146. [Google Scholar] [CrossRef]
- Kondreddy, V.K.; Anikisetty, M.; Naidu, K.A. Medium-chain triglycerides and monounsaturated fatty acids potentiate the beneficial effects of fish oil on selected cardiovascular risk factors in rats. J. Nutr. Biochem. 2016, 28, 91–102. [Google Scholar] [CrossRef]
- Terada, S.; Sekine, S.; Aoyama, T. Dietary Intake of Medium- and Long-chain Triacylglycerols Prevents the Progression of Hyperglycemia in Diabetic ob/ob Mice. J. Oleo Sci. 2015, 64, 683–688. [Google Scholar] [CrossRef] [Green Version]
- Tinius, R.A.; Blankenship, M.M.; Furgal, K.E.; Cade, W.T.; Pearson, K.J.; Rowland, N.S.; Pearson, R.C.; Hoover, D.L.; Maples, J.M. Metabolic flexibility is impaired in women who are pregnant and overweight/obese and related to insulin resistance and inflammation. Metabolism 2020, 104, 154142. [Google Scholar] [CrossRef] [PubMed]
- McCarty, M.F.; DiNicolantonio, J.J. Lauric acid-rich medium-chain triglycerides can substitute for other oils in cooking applications and may have limited pathogenicity. Open Heart 2016, 3, e000467. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Wang, Y.; Jiang, Y.; Zhang, Z.; Sun, X.; Yu, L.L. Dietary Intake of Structured Lipids with Different Contents of Medium-Chain Fatty Acids on Obesity Prevention in C57BL/6J Mice. J. Food Sci. 2017, 82, 1968–1977. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, Y.; Wang, J.; Zhang, R.; Jing, H.; Yu, X.; Zhang, Y.; Xu, Q.; Zhang, J.; Zheng, Z.; et al. Medium- and long-chain triacylglycerols reduce body fat and blood triacylglycerols in hypertriacylglycerolemic, overweight but not obese, Chinese individuals. Lipids 2010, 45, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Li, C.; Feng, F.; Wei, W.; Zhang, H. Synthesis of medium and long-chain triacylglycerols by enzymatic acidolysis of algal oil and lauric acid. LWT 2021, 136, 110309. [Google Scholar] [CrossRef]
- Wu, G.H.; Zaniolo, O.; Schuster, H.; Schlotzer, E.; Pradelli, L. Structured triglycerides versus physical mixtures of medium- and long-chain triglycerides for parenteral nutrition in surgical or critically ill adult patients: Systematic review and meta-analysis. Clin. Nutr. 2017, 36, 150–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DebMandal, M.; Mandal, S. Coconut (Cocos nucifera L.: Arecaceae): In health promotion and disease prevention. Asian Pac. J. Trop. Med. 2011, 4, 241–247. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Wang, Y.; Jacoby, J.J.; Jiang, Y.; Zhang, Y.; Yu, L.L. Effects of Medium- and Long-Chain Triacylglycerols on Lipid Metabolism and Gut Microbiota Composition in C57BL/6J Mice. J. Agric. Food Chem. 2017, 65, 6599–6607. [Google Scholar] [CrossRef]
- Ye, Z.; Xu, Y.-J.; Liu, Y. Influences of dietary oils and fats, and the accompanied minor content of components on the gut microbiota and gut inflammation: A review. Trends Food Sci. Technol. 2021, 113, 255–276. [Google Scholar] [CrossRef]
- Jiang, Z.; Zhao, M.; Zhang, H.; Li, Y.; Liu, M.; Feng, F. Antimicrobial Emulsifier-Glycerol Monolaurate Induces Metabolic Syndrome, Gut Microbiota Dysbiosis, and Systemic Low-Grade Inflammation in Low-Fat Diet Fed Mice. Mol. Nutr. Food Res. 2018, 62, 1700547. [Google Scholar] [CrossRef] [PubMed]
- Rial, S.; Karelis, A.; Bergeron, K.-F.; Mounier, C. Gut Microbiota and Metabolic Health: The Potential Beneficial Effects of a Medium Chain Triglyceride Diet in Obese Individuals. Nutrients 2016, 8, 281. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, Z.; Jin, G.; Yang, X.; Zhou, H. Regulating dyslipidemia effect of polysaccharides from Pleurotus ostreatus on fat-emulsion-induced hyperlipidemia rats. Int. J. Biol. Macromol. 2017, 101, 107–116. [Google Scholar] [CrossRef]
- Matsuo, T.; Takeuchi, H. Effects of structured medium-and long-chain triacylglycerols in diets with various levels of fat on body fat accumulation in rats. Br. J. Nutr. 2004, 91, 219–225. [Google Scholar] [CrossRef] [Green Version]
- Nowak, P.; Zaworska-Zakrzewska, A.; Frankiewicz, A.; Kasprowicz-Potocka, M. The effects and mechanisms of acids on the health of piglets and weaners—A review. Ann. Anim. Sci. 2021, 21, 433–455. [Google Scholar] [CrossRef]
- Mustafa, N.G. Biochemical Trails Associated with Different Doses of Alpha-Monolaurin in Chicks. Adv. Anim. Vet. Sci. 2018, 7, 187–192. [Google Scholar] [CrossRef] [Green Version]
- Kallio, H.; Nylund, M.; Bostrom, P.; Yang, B. Triacylglycerol regioisomers in human milk resolved with an algorithmic novel electrospray ionization tandem mass spectrometry method. Food Chem. 2017, 233, 351–360. [Google Scholar] [CrossRef]
- Saraswathi, V.; Kumar, N.; Gopal, T.; Bhatt, S.; Ai, W.; Ma, C.; Talmon, G.A.; Desouza, C.J.B. Lauric acid versus palmitic acid: Effects on adipose tissue inflammation, insulin resistance, and non-alcoholic fatty liver disease in obesity. Biology 2020, 9, 346. [Google Scholar] [CrossRef] [PubMed]
- Nobili, V.; Bedogni, G.; Alisi, A.; Pietrobattista, A.; Rise, P.; Galli, C.; Agostoni, C. Docosahexaenoic acid supplementation decreases liver fat content in children with non-alcoholic fatty liver disease: Double-blind randomised controlled clinical trial. Arch. Dis. Child. 2011, 96, 350–353. [Google Scholar] [CrossRef] [Green Version]
- Scheele, C.; Nielsen, S. Metabolic regulation and the anti-obesity perspectives of human brown fat. Redox Biol. 2017, 12, 770–775. [Google Scholar] [CrossRef]
- Stolarczyk, E. Adipose tissue inflammation in obesity: A metabolic or immune response? Curr. Opin. Pharmacol. 2017, 37, 35–40. [Google Scholar] [CrossRef]
- Zygmunt, D.A.; Crowe, K.E.; Flanigan, K.M.; Martin, P.T. Comparison of Serum rAAV Serotype-Specific Antibodies in Patients with Duchenne Muscular Dystrophy, Becker Muscular Dystrophy, Inclusion Body Myositis, or GNE Myopathy. Hum. Gene 2017, 28, 737–746. [Google Scholar] [CrossRef]
- Pons, Z.; Guerrero, L.; Margalef, M.; Arola, L.; Arola-Arnal, A.; Muguerza, B. Effect of low molecular grape seed proanthocyanidins on blood pressure and lipid homeostasis in cafeteria diet-fed rats. J. Physiol. Biochem. 2014, 70, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Dehghan, A.; van Hoek, M.; Sijbrands, E.J.; Hofman, A.; Witteman, J.C. High serum uric acid as a novel risk factor for type 2 diabetes. Diabetes Care 2008, 31, 361–362. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.N.; Shen, J.R.; Xiong, C.Y.; Zhu, X.M.; Deng, Z.Y. Investigation of Lipid Metabolism by a New Structured Lipid with Medium- and Long-Chain Triacylglycerols from Cinnamomum camphora Seed Oil in Healthy C57BL/6J Mice. J. Agric. Food Chem. 2018, 66, 1990–1998. [Google Scholar] [CrossRef]
- Kim, Y.; Lim, J.H.; Kim, M.Y.; Kim, E.N.; Yoon, H.E.; Shin, S.J.; Choi, B.S.; Kim, Y.S.; Chang, Y.S.; Park, C.W. The Adiponectin Receptor Agonist AdipoRon Ameliorates Diabetic Nephropathy in a Model of Type 2 Diabetes. J. Am. Soc. Nephrol. 2018, 29, 1108–1127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geng, S.; Zhu, W.; Xie, C.; Li, X.; Wu, J.; Liang, Z.; Xie, W.; Zhu, J.; Huang, C.; Zhu, M.; et al. Medium-chain triglyceride ameliorates insulin resistance and inflammation in high fat diet-induced obese mice. Eur. J. Nutr. 2016, 55, 931–940. [Google Scholar] [CrossRef]
- Du, Y.X.; Chen, S.N.; Zhu, H.L.; Niu, X.; Li, J.; Fan, Y.W.; Deng, Z.Y. Consumption of Interesterified Medium- and Long-Chain Triacylglycerols Improves Lipid Metabolism and Reduces Inflammation in High-Fat Diet-Induced Obese Rats. J. Agric. Food Chem. 2020, 68, 8255–8262. [Google Scholar] [CrossRef] [PubMed]
- Zhong, H.; Deng, L.; Zhao, M.; Tang, J.; Liu, T.; Zhang, H.; Feng, F. Probiotic-fermented blueberry juice prevents obesity and hyperglycemia in high fat diet-fed mice in association with modulating the gut microbiota. Food Funct. 2020, 11, 9192–9207. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.-Y.; Hao, Y.-F.; Hao, Z.-X.; Liu, Q.; Zhang, L.; Jiang, C.-P.; Jiang, J.-G. Flavonoids from Rosa davurica Pall. fruits prevent high-fat diet-induced obesity and liver injury via modulation of the gut microbiota in mice. Food Funct. 2021, 12, 10097–10106. [Google Scholar] [CrossRef]
- Companys, J.; Gosalbes, M.J.; Pla-Pagà, L.; Calderón-Pérez, L.; Llauradó, E.; Pedret, A.; Valls, R.M.; Jiménez-Hernández, N.; Sandoval-Ramirez, B.A.; Del Bas, J.M. Gut microbiota profile and its association with clinical variables and dietary intake in overweight/obese and lean subjects: A cross-sectional study. Nutrients 2021, 13, 2032. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, J.; Wang, Q.; Li, C.; Yang, M.; Hussain, M.; Zhang, J.; Feng, F.; Zhong, H. Effects of the Novel LaPLa-Enriched Medium- and Long-Chain Triacylglycerols on Body Weight, Glycolipid Metabolism, and Gut Microbiota Composition in High Fat Diet-Fed C57BL/6J Mice. Molecules 2023, 28, 722. https://doi.org/10.3390/molecules28020722
Shi J, Wang Q, Li C, Yang M, Hussain M, Zhang J, Feng F, Zhong H. Effects of the Novel LaPLa-Enriched Medium- and Long-Chain Triacylglycerols on Body Weight, Glycolipid Metabolism, and Gut Microbiota Composition in High Fat Diet-Fed C57BL/6J Mice. Molecules. 2023; 28(2):722. https://doi.org/10.3390/molecules28020722
Chicago/Turabian StyleShi, Jinyuan, Qianqian Wang, Chuang Li, Mengyu Yang, Muhammad Hussain, Junhui Zhang, Fengqin Feng, and Hao Zhong. 2023. "Effects of the Novel LaPLa-Enriched Medium- and Long-Chain Triacylglycerols on Body Weight, Glycolipid Metabolism, and Gut Microbiota Composition in High Fat Diet-Fed C57BL/6J Mice" Molecules 28, no. 2: 722. https://doi.org/10.3390/molecules28020722