
TRPV3: Structure, Diseases and Modulators



Abstract
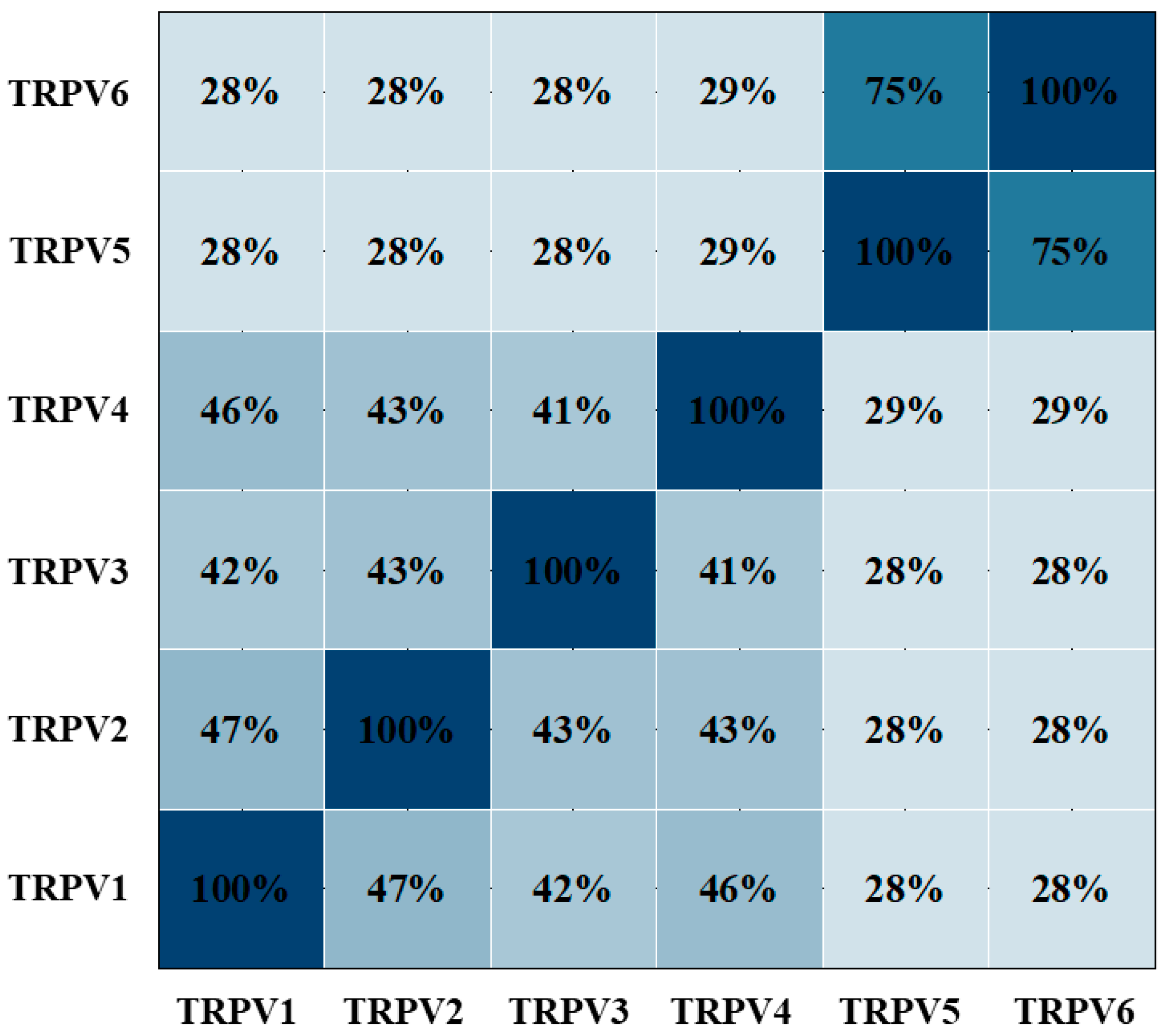
:1. Introduction
2. Diseases Related to TRPV3
2.1. Skin Diseases
2.2. Cancers
2.3. Cardiac Diseases
2.4. Others
3. Small-Molecule TRPV3 Modulators
3.1. TRPV3 Agonists
3.1.1. Natural Compounds
3.1.2. Synthetic Compounds
3.1.3. Endogenous Substances
3.2. TRPV3 Antagonists
3.2.1. Natural Compounds
3.2.2. Synthetic Compounds
3.2.3. Endogenous Substances

{kind=link}
{kind=link}
{kind=link}
| Entry | Name | Structure | Activate | Inhibit | EC50 for TRPV3 | Description/Use | Reference(s) |
|---|---|---|---|---|---|---|---|
| Natural compounds | |||||||
| 1 | Incensole acetate (IA) |  | TRPV3 | — | 16 µM | A Boswellia resin component can induce anti-anxiety-like and antidepressant behavior effects in wild-type (WT) mice. | [81] |
| 2 | Serratol |  | TRPV3 | — | 0.15 ± 0.01 μM | A TRPV3 agonist, isolated from Indian frankincense, had low retrofitting potential. Its affinity can be significantly improved by the acylation, esterification and oxidation of serratol. | [82] |
| 3 | Camphor |  | TRPV1 TRPV3 TRPM8 | TRPA1 | 6.03 ± 1.47 mM | Topical analgesia by desensitizing TRPV1; modulated the sensations of warmth in humans. | [78,80,86,131,132] |
| 4 | (+)-Borneol |  | TRPV3 TRPM8 | TRPA1 | 3.45 ± 0.13 mM | Borneol, a common Chinese medicine, had significant analgesic and anti-inflammatory effects by activating TRPM8. | [86,133,134] |
| 5 | Menthol |  | TRPV3 TRPM8 TRPA1 (low) | TRPA1 (high) | 1 mM | Menthol mainly activated the hot channel by TRPV3 and the cold channel by TRPM8, and it can induce an analgesic effect by the inhibition of TRPA1. | [135,136] |
| 6 | Carvacrol |  | TRPV3 TRPA1 | TRPM8 | 0.49 ± 0.07 mM | Carvacrol is rapidly desensitized by TRPA1. | [63,86,136] |
| 7 | Eugenol |  | TRPV1 TRPV3 TRPA1 | — | 2.3 mM | Carvol, isolated from oregano essential oil, is a powerful vasodilator by TRPV3 in the endothelium. | [63,137] |
| 8 | Citral |  | TRPV1 TRPV3 TRPM8 TRPA1 | TRPV1-4 TRPM8 TRPA1 | 926 µM | Citral, a bioactive ingredient in lemongrass, is used to treat abnormal pain, itching or other types of pain involving superficial sensory nerves and skin. | [85] |
| 9 | 6-tert-Butyl-m-cresol |  | TRPV3 | — | 0.37 ± 0.1 mM | A much more potent TRPV3 inhibitor than camphor is. | [86] |
| 10 | Thymol |  | TRPV3 TRPM8 TRPA1 | — | 0.86 ± 0.07 mM | A TRP channel agonist derived from thyme (thymus vulgaris) and oregano (origanum vulgare). | [63,86,131] |
| 11 | Dihydrocarve-ol |  | TRPV3 | — | 2.57 ± 0.42 mM | A potent TRPV3 inhibitor that is used as food additives and artificial spices. | [86] |
| 12 | (-)-Carveol |  | TRPV3 | — | 3.03 ± 1.16 mM | A potent TRPV3 inhibitor that is used as food additives and artificial spices. | [86] |
| 13 | Tetrahydrocannabivarin (THCV) |  | TRPV2 TRPV3 TRPV4 | — | 3.8 ± 0.4 µM | A non-specific TRPV channel agonist that is used as an anti-tumor, nervous system protection, immunomodulatory, anti-inflammatory and anti-oxidant agent. | [92,138,139] |
| 14 | Cannabidiol (CBD) |  | TRPV1 TRPV2 TRPV3 TRPV4 TRPM8 TRPA1 | — | 3.7 ± 1.6 µM | Cannabidiol has anti-epileptic, immunomodulatory, analgesic, anti-oxidation, anti-convulsive, anti-anxiety and other effects. | [92,140] |
| Synthetic compounds | |||||||
| 15 | 2-Aminoethoxydiphenyl borate (2-APB) |  | TRPV1 TRPV2 TRPV3 | TRPC1-6 TRPM7 TRPM8 | 28 µM | 2-APB can activate TRPV1, TRPV2 and TRPV3 at higher concentrations, and it inhibits the IP3 receptor and TRP channels. | [96] |
| 16 | Diphenylborinic anhydride (DPBA) |  | TRPV1 TRPV2 TRPV3 TRPV4 | — | 85.1 µM | 2-APB structural analogue. | [115] |
| 17 | Drofenine |  | TRPV3 | — | 207 µM | 2-APB structural analogue that can improve TRPV3 selectivity. | [100] |
| Endogenous substances | |||||||
| 18 | FPP |  | — | 0.13 µM | The first endogenous TRPV3 activator. | [35] | |
| 19 | Nitric oxide | NO | — | NA | A pleiotropic cellular signaling molecule that can activate TRPV3 through cysteine nitrosation. | [102] | |
| Entry | Name | Structure | Activate | Inhibit | IC50 for TRPV3 | Description/Use | Reference(s) |
|---|---|---|---|---|---|---|---|
| Natural compounds | |||||||
| 1 | Citrusinine II |  | — | TRPV3 | 12.43 μM | Citrusinine II can selectively inhibit TRPV3 and reduce itchy behavior by interacting with Y564 in the TRPV3 helix. | [104] |
| 2 | Osthole |  | — | TRPV1 TRPV3 | 37.0 ± 1.9 μM | Cusson, isolated from Cnidium monnieri (L.), is used as an antipruritic herbal medicine. | [105,141] |
| 3 | Isochlorogenic acid A |  | — | TRPV3 | 2.7 ± 1.3 μM | A TRPV3 specific inhibitor that can significantly reverse ear swelling in dermatitis and chronic pruritus. | [107] |
| 4 | Isochlorogenic acid B |  | — | TRPV3 | 0.9 ± 0.3 μM | A TRPV3 specific inhibitor that can significantly reverse ear swelling in dermatitis and chronic pruritus. | [107] |
| 5 | Forsythoside B |  | — | TRPV3 | 6.7 ± 0.7 μM | A TRPV3 inhibitor that can significantly reduce acute pruritus. | [108] |
| 6 | Monanchomycalin B |  | — | TRPV1 TRPV2 TRPV3 | 3.25 μM | A TRPV inhibitor that is isolated from the marine sponge Monanchora pulchra. | [112] |
| 7 | Verbascoside |  | — | TRPV3 | 14.1 ± 3.3 μM | A TRPV3 inhibitor that can effectively relieve atopic dermatitis when applied topically. | [109] |
| 8 | Pulchranin A |  | — | TRPV1 TRPV3 TRPA1 | 71.8 ± 9.4 μM | A moderately potent TRPV1 inhibitor and a minimally potent TRPV3 and TRPA1 inhibitor that is isolated from marine sponge Monanchora pulchra. | [110,111] |
| 9 | Pulchranin B |  | — | TRPV1 TRPV3 TRPA1 | 117.9 ± 11.8 μM | A moderately potent TRPV1 inhibitor and a minimally potent TRPV3 and TRPA1 inhibitor that is isolated from marine sponge Monanchora pulchra. | [110] |
| 10 | Pulchranin C |  | — | TRPV1 TRPV3 TRPA1 | >200 μM | A moderate potent TRPV1 inhibitor and a minimally potent TRPV3 and TRPA1 inhibitor that is isolated from marine sponge Monanchora pulchra. | [110] |
| Synthetic compounds | |||||||
| 11 | Ruthenium Red |  | — | TRP(S) | NA | Broad-spectrum, non-selective, cationic channel blockers. | [113] |
| 12 | 2,2-diphenyltetrahydro-furan (DPTHF) |  | — | TRPV1 TRPV2 TRPV3 | 6–10 μM | 2-APB structural analogue. | [115] |
| 13 | PC5 |  | — | TRPV3 | 2.63 ± 0.28 μM | A TRPV3 inhibitor that is obtained by virtual protein structure screening and lead compound structure optimization. | [116] |
| 14 | 7C |  | — | TRPV3 | 1.05 μM | A TRPV3 inhibitor that is obtained by the bioelectron isoarrangement principle. | [117] |
| 15 | Bupivacaine |  | — | TRPV3 | 0.17 ± 0.04 mM | A TRPV3 inhibitor that can be used as local anesthetics by extracellular interactions of their charged forms with the TRPV3 channel pore. | [118] |
| 16 | Mepivacaine |  | — | TRPV3 | 1.4 ± 0.3 mM | A TRPV3 inhibitor that can be used as local anesthetics by extracellular interactions of their charged forms with the TRPV3 channel pore. | [118] |
| 17 | Lidocaine |  | — | TRPV3 | 2.5 ± 0.5 mM | A TRPV3 inhibitor that can be used as local anesthetics by extracellular interactions of their charged forms with the TRPV3 channel pore. | [118] |
| 18 | Ropivacaine |  | — | TRPV3 | 0.28 ± 0.04 mM | A TRPV3 inhibitor that can be used as local anesthetics by extracellular interactions of their charged forms with the TRPV3 channel pore. | [118] |
| 19 | Dyclonine |  | — | TRPV3 | 3.2 μM | A TRPV3 inhibitor that can ameliorate the hyperactivity caused by itch/scratching behaviors. | [119] |
| 20 | Example #64 (WO 2006/122156) |  | — | TRPV3 | 0.2–1 μM | A TRPV3 inhibitor discovered by Hydra Biosciences that can reduce heat sensitivity after carrageenan injection in burns or hind paws. | [121] |
| 21 | DCP-THQ (WO 2007/056124) |  | — | TRPV3 | 117 nM | A TRPV3 inhibitor discovered by Hydra Biosciences. | [123] |
| 22 | Example #19 (US 2010/0292554A1) |  | — | TRPV3 | 200–1000 nM | A TRPV3 inhibitor discovered by Glenmark. | [124] |
| 23 | Example #58 (US 2009/0286811A1) |  | — | TRPV3 | < 500 nM | A TRPV3 inhibitor discovered by Glenmark. | [125] |
| 24 | Example #58 (WO 2009/130560) |  | — | TRPV3 | <250 nM | A TRPV3 inhibitor discovered by Glenmark. | [126] |
| 25 | Example #23 (WO 2010/055384) |  | — | TRPV3 | <50 nM | A TRPV3 inhibitor discovered by Glenmark. | [127] |
| 26 | Example #7 (US2010/0152192) |  | — | TRPV3 | <100 nM | A TRPV3 inhibitor discovered by Glenmark. | [128] |
| 27 | Example #37 (US2010/0152192) |  | — | TRPV3 | <100 nM | A TRPV3 inhibitor discovered by Glenmark. | [128] |
| 28 | Example #83 (WO2010/073128) |  | — | TRPV3 | <50 nM | A TRPV3 inhibitor discovered by Glenmark. | [129] |
| 29 | 74a |  | — | TRPV3 | 0.38 μM | A high potent TRPV3 inhibitor discovered by Glenmark that can be effective in rat neuropathic pain model. | [130] |
| Endogenous substance | |||||||
| 30 | 17R-RvD1 |  | — | TRPV3 | 0.4 μM | An endogenous TRPV3 inhibitor of endogenous lipid metabolites. | [142] |
| 31 | Isopentenyl pyrophosphate (IPP) |  | — | TRPV3 TRPA1 | 0.24 μM | An endogenous TRPA1 and TRPV3 inhibitor for topical analgesia. | [143] |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Cosens, D.J.; Manning, A. Abnormal Electroretinogram from a Drosophila Mutant. Nature 1969, 224, 285–287. [Google Scholar] [CrossRef] [PubMed]
- Minke, B.; Wu, C.-F.; Pak, W.L. Induction of photoreceptor voltage noise in the dark in Drosophila mutant. Nature 1975, 258, 84–87. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Maksaev, G.; Rau, M.; Xie, Z.; Hu, H.; Fitzpatrick, J.A.J.; Yuan, P. Gating of human TRPV3 in a lipid bilayer. Nat. Struct. Mol. Biol. 2020, 27, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cheng, X.; Tian, J.; Xiao, Y.; Tian, T.; Xu, F.; Hong, X.; Zhu, M.X. TRPC channels: Structure, function, regulation and recent advances in small molecular probes. Pharmacol. Ther. 2020, 209, 107497. [Google Scholar] [CrossRef]
- Chyb, S.; Raghu, P.; Hardie, R.C. Polyunsaturated fatty acids activate the Drosophila light-sensitive channels TRP and TRPL. Nature 1999, 397, 255–259. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.L.; Kwon, Y.; Adegbola, A.A.; Luo, J.; Chess, A.; Montell, C. Function of Rhodopsin in Temperature Discrimination in Drosophila. Science 2011, 331, 1333–1336. [Google Scholar] [CrossRef]
- Zhong, L.; Bellemer, A.; Yan, H.; Honjo, K.; Robertson, J.; Hwang, R.; Pitt, G.; Tracey, W. Thermosensory and nonther-mosensory isoforms of drosophila melanogaster trpa1 reveal heat-sensor domains of a thermotrp channel. Cell Rep. 2012, 1, 43–55. [Google Scholar] [CrossRef] [Green Version]
- Kang, K.; Pulver, S.R.; Panzano, V.C.; Chang, E.C.; Griffith, L.C.; Theobald, D.L.; Garrity, P.A. Analysis of Drosophila TRPA1 reveals an ancient origin for human chemical nociception. Nature 2010, 464, 597–600. [Google Scholar] [CrossRef] [Green Version]
- Julius, D. From peppers to peppermints: Natural products as probes of the pain pathway. Harvey Lect. 2007, 101, 89–115. [Google Scholar]
- Wei, Y.; Zhang, M.; Lyu, Z.; Yang, G.; Tian, T.; Ding, M.; Zeng, X.; Xu, F.; Wang, P.; Li, F.; et al. Benzothiazole amides as trpc3/6 inhibitors for gastric cancer treatment. ACS Omega 2021, 6, 9196–9203. [Google Scholar] [CrossRef]
- Montell, C. Physiology, phylogeny, and functions of the trp superfamily of cation channels. Sci. STKE 2001, 2001, re1. [Google Scholar] [CrossRef]
- Nilius, B.; Voets, T.; Peters, J. TRP Channels in Disease. Sci. STKE 2005, 2005, 8. [Google Scholar] [CrossRef]
- Minke, B.; Cook, B. TRP Channel Proteins and Signal Transduction. Physiol. Rev. 2002, 82, 429–472. [Google Scholar] [CrossRef] [Green Version]
- Qu, C.; Ding, M.; Zhu, Y.; Lu, Y.; Du, J.; Miller, M.; Tian, J.; Zhu, J.; Xu, J.; Wen, M.; et al. Pyrazolopyrimidines as potent stimulators for transient receptor potential canonical 3/6/7 channels. J. Med. Chem. 2017, 60, 4680–4692. [Google Scholar] [CrossRef]
- Zhu, Y.; Lu, Y.; Qu, C.; Miller, M.; Tian, J.; Thakur, D.P.; Zhu, J.; Deng, Z.; Hu, X.; Wu, M.; et al. Identification and optimization of 2-aminobenzimidazole derivatives as novel inhibitors of TRPC4 and TRPC5 channels. Br. J. Pharmacol. 2015, 172, 3495–3509. [Google Scholar] [CrossRef] [Green Version]
- Clapham, D.E. TRP channels as cellular sensors. Nature 2003, 426, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, L.; Tian, H.; Liu, M.; Jiang, S.; Shen, J.; Wang, K.; Cao, Z. Discovery of pyrroledione analogs as potent transient receptor potential canonical channel 5 inhibitors. Bioorganic. Med. Chem. Lett. 2022, 61, 128612. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.; Gu, L.; Hu, Y.; Ren, Y.; Yang, Z.; Chai, C.; Yu, B.; Ge, H.; Cao, Z.; Zhao, F. Inhibition of trpc6 suppressed tgf-beta-induced fibroblast-myofibroblast transdifferentiation in renal interstitial nrk-49f cells. Exp. Cell Res. 2022, 421, 113374. [Google Scholar] [CrossRef] [PubMed]
- Ding, M.; Wang, H.; Qu, C.; Xu, F.; Zhu, Y.; Lv, G.; Lu, Y.; Zhou, Q.; Zhou, H.; Zeng, X.; et al. Pyrazolo [1,5-a]pyrimidine TRPC6 antagonists for the treatment of gastric cancer. Cancer Lett. 2018, 432, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, G.; Raddatz, N.; Lorenzo, Y.; González, C.; Latorre, R. Biophysical and Molecular Features of Thermosensitive TRP Channels Involved in Sensory Transduction. 2015, pp. 1–39. Available online: https://link.springer.com/chapter/10.1007/978-3-319-18705-1_1 (accessed on 14 February 2022).
- Singh, A.K.; McGoldrick, L.L.; Sobolevsky, A.I. Structure and gating mechanism of the transient receptor potential channel TRPV3. Nat. Struct. Mol. Biol. 2018, 25, 805–813. [Google Scholar] [CrossRef]
- Xu, X.Z.S.; Li, H.S.; Guggino, W.B.; Montell, C. Coassembly of trp and trpl produces a distict store-operated conductance. Cell 1997, 89, 1155. [Google Scholar] [CrossRef] [PubMed]
- Seebohm, G.; Schreiber, J.A. Beyond Hot and Spicy: TRPV Channels and their Pharmacological Modulation. Cell Physiol. Biochem. 2021, 55, 108–130. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Ramsey, I.S.; Kotecha, S.A.; Moran, M.M.; Chong, J.A.; Lawson, D.; Ge, P.; Lilly, J.; Silos-Santiago, I.; Xie, Y.; et al. Trpv3 is a calcium-permeable temperature-sensitive cation channel. Nature 2002, 418, 181–186. [Google Scholar] [CrossRef]
- Toledo Mauriño, J.J.; Fonseca-Camarillo, G.; Furuzawa-Carballeda, J.; Barreto-Zuñiga, R.; Martínez Benítez, B.; Granados, J.; Yamamoto-Furusho, J.K. TRPV Subfamily (TRPV2, TRPV3, TRPV4, TRPV5, and TRPV6) Gene and Protein Expression in Patients with Ulcerative Colitis. J. Immunol. Res. 2020, 2020, 2906845. [Google Scholar] [CrossRef] [PubMed]
- Smith, G.D.; Gunthorpe, M.J.; Kelsell, R.E.; Hayes, P.D.; Reilly, P.; Facer, P.; Wright, J.E.; Jerman, J.C.; Walhin, J.-P.; Ooi, L.; et al. TRPV3 is a temperature-sensitive vanilloid receptor-like protein. Nature 2002, 418, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Vriens, J.; Appendino, G.; Nilius, B. Pharmacology of Vanilloid Transient Receptor Potential Cation Channels. Mol. Pharmacol. 2009, 75, 1262–1279. [Google Scholar] [CrossRef]
- Grandl, J.; Hu, H.; Bandell, M.; Bursulaya, B.; Schmidt, M.; Petrus, M.; Patapoutian, A. Pore region of TRPV3 ion channel is specifically required for heat activation. Nat. Neurosci. 2008, 11, 1007–1013. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.E.; Patapoutian, A.; Grandl, J. Single Residues in the Outer Pore of TRPV1 and TRPV3 Have Temperature-Dependent Conformations. PLoS ONE 2013, 8, e59593. [Google Scholar] [CrossRef]
- Nilius, B.; Bíró, T.; Owsianik, G. TRPV3: Time to decipher a poorly understood family member! J. Physiol. 2013, 592, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Yamada, T.; Ueda, T.; Ugawa, S.; Ishida, Y.; Imayasu, M.; Koyama, S.; Shimada, S. Functional expression of transient receptor potential vanilloid 3 (TRPV3) in corneal epithelial cells: Involvement in thermosensation and wound healing. Exp. Eye Res. 2010, 90, 121–129. [Google Scholar] [CrossRef]
- Liu, B.; Yao, J.; Zhu, M.X.; Qin, F. Hysteresis of gating underlines sensitization of TRPV3 channels. J. Gen. Physiol. 2011, 138, 509–520. [Google Scholar] [CrossRef] [PubMed]
- Peier, A.M.; Reeve, A.J.; Andersson, D.A.; Moqrich, A.; Patapoutian, A. A heat-sensitive trp channel expressed in keratinocytes. Science 2002, 296, 2046–2049. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Wang, K. The Ca2+-Permeable Cation Transient Receptor Potential TRPV3 Channel: An Emerging Pivotal Target for Itch and Skin Diseases. Mol. Pharmacol. 2017, 92, 193–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bang, S.; Yoo, S.; Yang, T.-J.; Cho, H.; Hwang, S.W. Farnesyl Pyrophosphate Is a Novel Pain-producing Molecule via Specific Activation of TRPV3. J. Biol. Chem. 2010, 285, 19362–19371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, T.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. TRPV3 regulates nitric oxide synthase-independent nitric oxide synthesis in the skin. Nat. Commun. 2011, 2, 369. [Google Scholar] [CrossRef] [Green Version]
- Duchatelet, S.; Pruvost, S.; de Veer, S.; Fraitag, S.; Nitschké, P.; Bole-Feysot, C.; Bodemer, C.; Hovnanian, A. A new TRPV3 missense mutation in a patient with Olmsted syndrome and erythromelalgia. JAMA Dermatol. 2014, 150, 303–306. [Google Scholar] [CrossRef] [Green Version]
- Lai-Cheong, J.E.; Sethuraman, G.; Ramam, M.; Stone, K.; Simpson, M.A.; McGrath, J.A. Recurrent heterozygous missense mutation, p.gly573ser, in the trpv3 gene in an indian boy with sporadic olmsted syndrome. Br. J. Dermatol. 2012, 167, 440–442. [Google Scholar] [CrossRef]
- Eytan, O.; Fuchs-Telem, D.; Mevorach, B.; Indelman, M.; Bergman, R.; Sarig, O.; Goldberg, I.; Adir, N.; Sprecher, E. Olmsted Syndrome Caused by a Homozygous Recessive Mutation in TRPV3. J. Investig. Dermatol. 2014, 134, 1752–1754. [Google Scholar] [CrossRef] [Green Version]
- Agarwala, M.; George, R.; Pramanik, R.; McGrath, J. Olmsted syndrome in an Indian male with a new de novo mutation in TRPV3. Br. J. Dermatol. 2015, 174, 209–211. [Google Scholar] [CrossRef]
- Ni, C.; Yan, M.; Zhang, J.; Cheng, R.; Liang, J.; Deng, D.; Wang, Z.; Li, M.; Yao, Z. A novel mutation in TRPV3 gene causes atypical familial Olmsted syndrome. Sci. Rep. 2016, 6, 21815. [Google Scholar] [CrossRef] [Green Version]
- Duchatelet, S.; Guibbal, L.; Veer, S.; Fraitag, S.; Nitschké, P.; Zarhrate, M.; Bodemer, C.; Hovnanian, A. Olmsted syndrome with erythromelalgia caused by recessive transient receptor potential vanilloid 3 mutations. Br. J. Dermatol. 2014, 171, 675–678. [Google Scholar] [CrossRef] [PubMed]
- Yoshioka, T.; Imura, K.; Asakawa, M.; Suzuki, M.; Oshima, I.; Hirasawa, T.; Sakata, T.; Horikawa, T.; Arimura, A. Impact of the Gly573Ser Substitution in TRPV3 on the Development of Allergic and Pruritic Dermatitis in Mice. J. Investig. Dermatol. 2009, 129, 714–722. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto-Kasai, E.; Yasui, K.; Shichijo, M.; Sakata, T.; Yoshioka, T. Impact of TRPV3 on the development of allergic dermatitis as a dendritic cell modulator. Exp. Dermatol. 2013, 22, 820–824. [Google Scholar] [CrossRef]
- Vasas, N.; Pénzes, Z.; Kistamás, K.; Nánási, P.P.; Molnár, S.; Szegedi, A.; Szöllősi, A.G.; Bíró, T. Transient receptor potential vanilloid 3 expression is increased in non-lesional skin of atopic dermatitis patients. Exp. Dermatol. 2022, 31, 807–813. [Google Scholar] [CrossRef] [PubMed]
- Scott, V.; Patel, H.; Wetter, J.; Edlmayer, R.; Neelands, T.; Miller, L.; Huang, S.; Gauld, S.; Todorovic, V.; Gomtsian, A.; et al. 534 Defining a mechanistic link between TRPV3 activity and psoriasis through IL-1α and EGFR signaling pathways. J. Investig. Dermatol. 2016, 136, S94. [Google Scholar] [CrossRef]
- Yamamoto-Kasai, E.; Imura, K.; Yasui, K.; Shichijou, M.; Oshima, I.; Hirasawa, T.; Sakata, T.; Yoshioka, T. Trpv3 as a therapeutic target for itch. J. Investig. Dermatol. 2012, 132, 2109–2112. [Google Scholar] [CrossRef] [Green Version]
- Yang, Y.; Cho, S.; Choi, M.; Choi, Y.; Kwak, I.; Park, C.; Kim, H. Increased Expression of Three Types of Transient Receptor Potential Channels (TRPA1, TRPV4 and TRPV3) in Burn Scars with Post-burn Pruritus. Acta Derm.-Venereol. 2015, 95, 20–24. [Google Scholar] [CrossRef] [Green Version]
- Sulk, M.; Seeliger, S.; Aubert, J.; Schwab, V.D.; Cevikbas, F.; Rivier, M.; Nowak, P.; Voegel, J.J.; Buddenkotte, J.; Steinhoff, M. Distribution and expression of non-neuronal transient receptor potential (trpv) ion channels in rosacea. J. Investig. Dermatol. 2012, 132, 1253–1262. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zhang, Q.; Kai, F.; Li, B.; Li, H.; Qi, H.; Guo, J.; Cao, Y.; Sun, H. Overexpression of trpv3 correlates with tumor pro-gression in non-small cell lung cancer. Int. J. Mol. Sci. 2016, 17, 437. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.; Wang, Z.; Mu, J.; Zhu, M.; Zhen, Y.; Zhang, H. Upregulation of the transient receptor potential vanilloid 1 in colonic epithelium of patients with active inflammatory bowel disease. Int. J. Clin. Exp. Pathol. 2017, 10, 11335–11344. [Google Scholar]
- Sozucan, Y.; Kalender, M.E.; Sari, I.; Suner, A.; Oztuzcu, S.; Arman, K.; Yumrutas, O.; Bozgeyik, I.; Cengiz, B.; Igci, Y.Z.; et al. TRP genes family expression in colorectal cancer. Exp. Oncol. 2015, 37, 208–212. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Qi, H.; Cao, Y.; Shi, P.; Song, C.; Ba, L.; Chen, Y.; Gao, J.; Li, S.; Li, B.; et al. Activation of transient receptor potential vanilloid 3 channel (TRPV3) aggravated pathological cardiac hypertrophy via calcineurin/NFATc3 pathway in rats. J. Cell Mol. Med. 2018, 22, 6055–6067. [Google Scholar] [CrossRef]
- Liu, Y.; Qi, H.; E, M.; Shi, P.; Zhang, Q.; Li, S.; Wang, Y.; Cao, Y.; Chen, Y.; Ba, L.; et al. Transient receptor potential vanilloid-3 (trpv3) activation plays a central role in cardiac fibrosis induced by pressure overload in rats via tgf-beta1 pathway. Naunyn Schmiedebergs Arch. Pharmacol. 2018, 391, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, X.; Huang, W. MicroRNA-369 attenuates hypoxia-induced cardiomyocyte apoptosis and inflammation via targeting TRPV3. Braz. J. Med. Biol. Res. 2021, 54, e10550. [Google Scholar] [CrossRef] [PubMed]
- Aouizerat, B.; Dunn, L.; Cooper, B.; Paul, S.; West, C.; Miaskowski, C. Association between of temperature-sensitive transient receptor potential cation channel, subfamily V, member 3 (TRPV3) with preoperative pain in women with breast cancer. J. Pain 2011, 12, 25. [Google Scholar] [CrossRef]
- Gopinath, P.; Wan, E.; Holdcroft, A.; Facer, P.; Davis, J.B.; Smith, G.D.; Bountra, C.; Anand, P. Increased capsaicin receptor TRPV1 in skin nerve fibres and related vanilloid receptors TRPV3 and TRPV4 in keratinocytes in human breast pain. BMC Women’s Health 2005, 5, 2. [Google Scholar] [CrossRef] [Green Version]
- Huang, S.M.; Lee, H.; Chung, M.K.; Park, U.; Yu, Y.Y.; Bradshaw, H.B.; Coulombe, P.A.; Walker, J.M.; Caterina, M.J. Overexpressed transient receptor potential vanilloid 3 ion channels in skin keratinocytes modulate pain sensitivity via prostaglandin e2. J. Neurosci. 2008, 28, 13727–13737. [Google Scholar] [CrossRef] [Green Version]
- Yan, K.; Sun, X.; Wang, G.; Liu, Y.; Wang, K. Pharmacological Activation of Thermo–Transient Receptor Potential Vanilloid 3 Channels Inhibits Hair Growth by Inducing Cell Death of Hair Follicle Outer Root Sheath. J. Pharmacol. Exp. Ther. 2019, 370, 299–307. [Google Scholar] [CrossRef]
- Greco, C.; Leclerc-Mercier, S.; Chaumon, S.; Doz, F.; Hadj-Rabia, S.; Molina, T.; Boucheix, C.; Bodemer, C. Use of Epidermal Growth Factor Receptor Inhibitor Erlotinib to Treat Palmoplantar Keratoderma in Patients with Olmsted Syndrome Caused by TRPV3 Mutations. JAMA Dermatol. 2020, 156, 191–195. [Google Scholar] [CrossRef]
- Lin, Z.; Chen, Q.; Lee, M.; Cao, X.; Zhang, J.; Ma, D.; Chen, L.; Hu, X.; Wang, H.; Wang, X.; et al. Exome Sequencing Reveals Mutations in TRPV3 as a Cause of Olmsted Syndrome. Am. J. Hum. Genet. 2012, 90, 558–564. [Google Scholar] [CrossRef] [Green Version]
- Phelps, C.B.; Wang, R.R.; Choo, S.S.; Gaudet, R. Differential regulation of trpv1, trpv3, and trpv4 sensitivity through a con-served binding site on the ankyrin repeat domain. J. Biol. Chem. 2010, 285, 731–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Delling, M.; Jun, J.C.; Clapham, D.E. Oregano, thyme and clove-derived flavors and skin sensitizers activate specific TRP channels. Nat. Neurosci. 2006, 9, 628–635. [Google Scholar] [CrossRef]
- Phelps, C.B.; Huang, R.J.; Lishko, P.V.; Wang, R.R.; Gaudet, R. Structural Analyses of the Ankyrin Repeat Domain of TRPV6 and Related TRPV Ion Channels. Biochemistry 2008, 47, 2476–2484. [Google Scholar] [CrossRef] [Green Version]
- Roufogalis, B.D.; Mandadi, S. ThermoTRP Channels in Nociceptors: Taking a Lead from Capsaicin Receptor TRPV1. Curr. Neuropharmacol. 2008, 6, 21–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mene, P.; Punzo, G.; Pirozzi, N. TRP Channels as Therapeutic Targets in Kidney Disease and Hypertension. Curr. Top. Med. Chem. 2013, 13, 386–397. [Google Scholar] [CrossRef] [PubMed]
- Benemei, S.; Patacchini, R.; Trevisani, M.; Geppetti, P. Trp channels. Curr. Opin. Pharmacol. 2015, 22, 18–23. [Google Scholar] [CrossRef]
- Lindy, A.S.; Parekh, P.K.; Zhu, R.; Kanju, P.; Chintapalli, S.V.; Tsvilovskyy, V.; Patterson, R.L.; Anishkin, A.; van Rossum, D.B.; Liedtke, W.B. TRPV channel-mediated calcium transients in nociceptor neurons are dispensable for avoidance behaviour. Nat. Commun. 2014, 5, 4734. [Google Scholar] [CrossRef] [Green Version]
- Hoeft, B.; Linseisen, J.; Beckmann, L.; Müller-Decker, K.; Canzian, F.; Hüsing, A.; Kaaks, R.; Vogel, U.; Jakobsen, M.U.; Overvad, K.; et al. Polymorphisms in fatty acid metabolism-related genes are associated with colorectal cancer risk. Carcinogenesis 2009, 31, 466–472. [Google Scholar] [CrossRef] [Green Version]
- Sakakibara, A.; Sakakibara, S.; Kusumoto, J.; Takeda, D.; Hasegawa, T.; Akashi, M.; Minamikawa, T.; Hashikawa, K.; Terashi, H.; Komori, T. Upregulated Expression of Transient Receptor Potential Cation Channel Subfamily V Receptors in Mucosae of Patients with Oral Squamous Cell Carcinoma and Patients with a History of Alcohol Consumption or Smoking. PLoS ONE 2017, 12, e0169723. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, H.; Xue, C.; Chen, H.; Xue, Y.; Zhao, F.; Zhu, M.X.; Cao, Z. TRPV3 enhances skin keratinocyte proliferation through EGFR-dependent signaling pathways. Cell Biol. Toxicol. 2021, 37, 313–330. [Google Scholar] [CrossRef]
- Qi, H.; Ren, J.; E, M.; Zhang, Q.; Cao, Y.; Ba, L.; Song, C.; Shi, P.; Fu, B.; Sun, H. MiR-103 inhibiting cardiac hypertrophy through inactivation of myocardial cell autophagy via targeting TRPV3 channel in rat hearts. J. Cell Mol. Med. 2019, 23, 1926–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duchatelet, S.; Hovnanian, A. Olmsted syndrome: Clinical, molecular and therapeutic aspects. Orphanet J. Rare Dis. 2015, 10, 33. [Google Scholar] [CrossRef] [PubMed]
- Mergler, S.; Garreis, F.; Sahlmüller, M.; Reinach, P.S.; Paulsen, F.; Pleyer, U. Thermosensitive transient receptor potential channels in human corneal epithelial cells. J. Cell Physiol. 2010, 226, 1828–1842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aijima, R.; Wang, B.; Takao, T.; Mihara, H.; Kashio, M.; Ohsaki, Y.; Zhang, J.; Mizuno, A.; Suzuki, M.; Yamashita, Y.; et al. The thermosensitive TRPV3 channel contributes to rapid wound healing in oral epithelia. FASEB J. 2014, 29, 182–192. [Google Scholar] [CrossRef]
- Cheng, X.; Jin, J.; Hu, L.; Shen, D.; Dong, X.-P.; Samie, M.A.; Knoff, J.; Eisinger, B.; Liu, M.-L.; Huang, S.M.; et al. TRP Channel Regulates EGFR Signaling in Hair Morphogenesis and Skin Barrier Formation. Cell 2010, 141, 331–343. [Google Scholar] [CrossRef] [Green Version]
- Borbiro, I.; Lisztes, E.; Tóth, B.I.; Czifra, G.; Oláh, A.; Szöllősi, A.G.; Szentandrássy, N.; Nánási, P.P.; Péter, Z.; Paus, R.; et al. Activation of Transient Receptor Potential Vanilloid-3 Inhibits Human Hair Growth. J. Investig. Dermatol. 2011, 131, 1605–1614. [Google Scholar] [CrossRef]
- Xu, H. Camphor activates and strongly desensitizes the transient receptor potential vanilloid subtype 1 channel in a vanilloid-independent mechanism. J. Neurosci. 2005, 25, 8924–8937. [Google Scholar] [CrossRef] [Green Version]
- Bang, S.; Kim, K.Y.; Yoo, S.; Kim, Y.G.; Hwang, S.W. Transient receptor potential A1 mediates acetaldehyde-evoked pain sensation. Eur. J. Neurosci. 2007, 26, 2516–2523. [Google Scholar] [CrossRef]
- Selescu, T.; Ciobanu, A.C.; Dobre, C.; Reid, G.; Babes, A. Camphor activates and sensitizes transient receptor potential me-lastatin 8 (trpm8) to cooling and icilin. Chem. Senses 2013, 38, 563–575. [Google Scholar] [CrossRef]
- Moussaieff, A.; Rimmerman, N.; Bregman, T.; Straiker, A.; Felder, C.C.; Shoham, S.; Kashman, Y.; Huang, S.M.; Lee, H.; Shohami, E.; et al. Incensole acetate, an incense component, elicits psychoactivity by activating TRPV3 channels in the brain. FASEB J. 2008, 22, 3024–3034. [Google Scholar] [CrossRef] [Green Version]
- Pollastro, F.; Golin, S.; Chianese, G.; Putra, M.Y.; Moriello, A.S.; De Petrocellis, L.; García, V.; Munoz, E.; Taglialatela-Scafati, O.; Appendino, G. Neuroactive and Anti-inflammatory Frankincense Cembranes: A Structure–Activity Study. J. Nat. Prod. 2016, 79, 1762–1768. [Google Scholar] [CrossRef] [PubMed]
- Yang, B.H.; Piao, Z.G.; Kim, Y.-B.; Lee, C.-H.; Lee, J.K.; Park, K.; Kim, J.S.; Oh, S.B. Activation of Vanilloid Receptor 1 (VR1) by Eugenol. J. Dent. Res. 2003, 82, 781–785. [Google Scholar] [CrossRef]
- Niu, C.; Sun, X.; Hu, F.; Tang, X.; Wang, K. Molecular determinants for the chemical activation of the warmth-sensitive TRPV3 channel by the natural monoterpenoid carvacrol. J. Biol. Chem. 2022, 298, 111706. [Google Scholar] [CrossRef] [PubMed]
- Stotz, S.C.; Vriens, J.; Martyn, D.; Clardy, J.; Clapham, D.E. Citral sensing by transient [corrected] receptor potential channels in dorsal root ganglion neurons. PLoS ONE 2008, 3, e2082. [Google Scholar] [CrossRef]
- Vogt-Eisele, A.K.; Weber, K.; A Sherkheli, M.; Vielhaber, G.; Panten, J.; Gisselmann, G.; Hatt, H. Monoterpenoid agonists of TRPV3. Br. J. Pharmacol. 2007, 151, 530–540. [Google Scholar] [CrossRef] [Green Version]
- Sherkheli, M.A.; Benecke, H.; Doerner, J.F.; Kletke, O.; Vogt-Eisele, A.K.; Gisselmann, G.; Hatt, H. Monoterpenoids Induce Agonist-Specific Desensitization of Transient Receptor Potential Vanilloid-3 (TRPV3) ion Channels. J. Pharm. Pharm. Sci. 2009, 12, 116–128. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.-L.; Lei, M.; Zhou, L.-P.; Zeng, B.; Zou, F. Borneol Is a TRPM8 Agonist that Increases Ocular Surface Wetness. PLoS ONE 2016, 11, e0158868. [Google Scholar] [CrossRef] [Green Version]
- Peier, A.M.; Moqrich, A.; Hergarden, A.C.; Reeve, A.J.; Andersson, D.A.; Story, G.M.; Earley, T.J.; Dragoni, I.; McIntyre, P.; Bevan, S.; et al. A TRP Channel that Senses Cold Stimuli and Menthol. Cell 2002, 108, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Xiao, B.; Dubin, A.E.; Bursulaya, B.; Viswanath, V.; Jegla, T.; Patapoutian, A. Identification of Transmembrane Domain 5 as a Critical Molecular Determinant of Menthol Sensitivity in Mammalian TRPA1 Channels. J. Neurosci. 2008, 28, 9640–9651. [Google Scholar] [CrossRef] [Green Version]
- Andrade, E.; Meotti, F.; Calixto, J. TRPA1 antagonists as potential analgesic drugs. Pharmacol. Ther. 2011, 133, 189–204. [Google Scholar] [CrossRef]
- De Petrocellis, L.; Orlando, P.; Moriello, A.S.; Aviello, G.; Stott, C.; Izzo, A.; Di Marzo, V. Cannabinoid actions at TRPV channels: Effects on TRPV3 and TRPV4 and their potential relevance to gastrointestinal inflammation. Acta Physiol. 2011, 204, 255–266. [Google Scholar] [CrossRef] [PubMed]
- Etemad, L.; Karimi, G.; Alavi, M.S.; Roohbakhsh, A. Pharmacological effects of cannabidiol by transient receptor potential channels. Life Sci. 2022, 300, 120582. [Google Scholar] [CrossRef]
- Landucci, E.; Pellegrini-Giampietro, D.; Gianoncelli, A.; Ribaudo, G. Cannabidiol preferentially binds TRPV2: A novel mechanism of action. Neural Regen. Res. 2022, 17, 2693. [Google Scholar] [CrossRef]
- Hu, H.-Z.; Gu, Q.; Wang, C.; Colton, C.K.; Tang, J.; Kinoshita-Kawada, M.; Lee, L.-Y.; Wood, J.D.; Zhu, M.X. 2-Aminoethoxydiphenyl Borate Is a Common Activator of TRPV1, TRPV2, and TRPV3. J. Biol. Chem. 2004, 279, 35741–35748. [Google Scholar] [CrossRef] [Green Version]
- Chung, M.K.; Lee, H.; Mizuno, A.; Suzuki, M.; Caterina, M.J. 2-aminoethoxydiphenyl borate activates and sensitizes the heat-gated ion channel trpv3. J. Neurosci. 2004, 24, 5177–5182. [Google Scholar] [CrossRef] [Green Version]
- Scheuble, J.; Rössler, O.G.; Ulrich, M.; Thiel, G. Pharmacological and genetic inhibition of TRPC6-induced gene transcription. Eur. J. Pharmacol. 2020, 886, 173357. [Google Scholar] [CrossRef]
- Yelshanskaya, M.V.; Nadezhdin, K.D.; Kurnikova, M.G.; Sobolevsky, A.I. Structure and function of the calcium-selective TRP channel TRPV6. J. Physiol. 2020, 599, 2673–2697. [Google Scholar] [CrossRef]
- Oda, M.; Fujiwara, Y.; Ishizaki, Y.; Shibasaki, K. Oxidation sensitizes TRPV2 to chemical and heat stimuli, but not mechanical stimulation. Biochem. Biophys. Rep. 2021, 28, 101173. [Google Scholar] [CrossRef]
- Deeringrice, C.E.; Mitchell, V.K.; Romero, E.G.; Aziz, M.; Ryskamp, D.A.; Križaj, D.; Gopal, V.R.; Reilly, C.A. Drofenine: A 2-apb analogue with greater selectivity for human trpv3. Pharmacol. Res. Perspect. 2014, 2, e62. [Google Scholar] [CrossRef]
- Ma, L.; Lee, B.H.; Clifton, H.; Schaefer, S.; Zheng, J. Nicotinic Acid is a Common Regulator of Heat-Sensing TRPV1-4 Ion Channels. Sci. Rep. 2015, 5, 8906. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Inoue, R.; Morii, T.; Takahashi, N.; Yamamoto, S.; Hara, Y.; Tominaga, M.; Shimizu, S.; Sato, Y.; Mori, Y. Nitric oxide activates TRP channels by cysteine S-nitrosylation. Nat. Chem. Biol. 2006, 2, 596–607. [Google Scholar] [CrossRef]
- Hu, H.Z.; Xiao, R.; Wang, C.; Gao, N.; Colton, C.K.; Wood, J.D.; Zhu, M.X. Potentiation of trpv3 channel function by un-saturated fatty acids. J. Cell Physiol. 2006, 208, 201–212. [Google Scholar] [CrossRef]
- Han, Y.; Luo, A.; Kamau, P.M.; Takomthong, P.; Hu, J.; Boonyarat, C.; Luo, L.; Lai, R. A plant-derived trpv3 inhibitor sup-presses pain and itch. Br. J. Pharmacol. 2021, 178, 1669–1683. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.-Y.; Sun, L.-L.; Qi, H.; Gao, Q.; Wang, G.-X.; Wei, N.-N.; Wang, K. Antipruritic Effect of Natural Coumarin Osthole through Selective Inhibition of Thermosensitive TRPV3 Channel in the Skin. Mol. Pharmacol. 2018, 94, 1164–1173. [Google Scholar] [CrossRef] [Green Version]
- Neuberger, A.; Nadezhdin, K.D.; Zakharian, E.; Sobolevsky, A.I. Structural mechanism of TRPV3 channel inhibition by the plant-derived coumarin osthole. EMBO Rep. 2021, 22, e53233. [Google Scholar] [CrossRef]
- Qi, H.; Shi, Y.; Wu, H.; Niu, C.; Sun, X.; Wang, K. Inhibition of temperature-sensitive trpv3 channel by two natural iso-chlorogenic acid isomers for alleviation of dermatitis and chronic pruritus. Acta Pharm. Sin. B 2022, 12, 723–734. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, X.; Qi, H.; Ma, Q.; Zhou, Q.; Wang, W.; Wang, K. Pharmacological inhibition of the temperature-sensitive and ca(2+)-permeable transient receptor potential vanilloid trpv3 channel by natural forsythoside b attenuates pruritus and cytotoxicity of keratinocytes. J. Pharmacol. Exp. Ther. 2019, 368, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Sun, X.; Qi, H.; Wu, H.; Qu, Y.; Wang, K. Anti-pruritic and anti-inflammatory effects of natural verbascoside through selective inhibition of temperature-sensitive Ca2+-permeable TRPV3 channel. J. Dermatol. Sci. 2020, 97, 229–231. [Google Scholar] [CrossRef]
- Makarieva, T.N.; Ogurtsova, E.K.; Korolkova, Y.V.; Andreev, Y.; Mosharova, I.V.; Tabakmakher, K.M.; Guzii, A.G.; Denisenko, V.A.; Dmitrenok, P.S.; Lee, H.-S.; et al. Pulchranins B and C, New Acyclic Guanidine Alkaloids from the Far-Eastern Marine Sponge Monanchora Pulchra. Nat. Prod. Commun. 2013, 8, 1229–1232. [Google Scholar] [CrossRef] [Green Version]
- Guzii, A.G.; Makarieva, T.N.; Korolkova, Y.V.; Andreev, Y.A.; Mosharova, I.V.; Tabakmaher, K.M.; Denisenko, V.A.; Dmitrenok, P.S.; Ogurtsova, E.K.; Antonov, A.S.; et al. Pulchranin A, isolated from the Far-Eastern marine sponge, Monanchora pulchra: The first marine non-peptide inhibitor of TRPV-1 channels. Tetrahedron Lett. 2013, 54, 1247–1250. [Google Scholar] [CrossRef]
- Korolkova, Y.; Makarieva, T.; Tabakmakher, K.; Shubina, L.; Kudryashova, E.; Andreev, Y.; Mosharova, I.; Lee, H.-S.; Lee, Y.-J.; Kozlov, S. Marine Cyclic Guanidine Alkaloids Monanchomycalin B and Urupocidin A Act as Inhibitors of TRPV1, TRPV2 and TRPV3, but not TRPA1 Receptors. Mar. Drugs 2017, 15, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vennekens, R.; Owsianik, G.; Nilius, B. Vanilloid Transient Receptor Potential Cation Channels: An Overview. Curr. Pharm. Des. 2008, 14, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Pierre, M.S.; Reeh, P.; Zimmermann, K. Differential effects of TRPV channel block on polymodal activation of rat cutaneous nociceptors in vitro. Exp. Brain Res. 2009, 196, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.-K.; Güler, A.D.; Caterina, M.J. Biphasic Currents Evoked by Chemical or Thermal Activation of the Heat-gated Ion Channel, TRPV3. J. Biol. Chem. 2005, 280, 15928–15941. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Lin, Y.; Min, W.; Hou, Y.; Yuan, K.; Wang, J.; Yang, P. Computational discovery, structural optimization and biological evaluation of novel inhibitors targeting transient receptor potential vanilloid type 3 (trpv3). Bioorg. Chem. 2021, 114, 105093. [Google Scholar] [CrossRef]
- Lv, M.; Wu, H.; Qu, Y.; Wu, S.; Wang, J.; Wang, C.; Luan, Y.; Zhang, Z. The design and synthesis of transient receptor potential vanilloid 3 inhibitors with novel skeleton. Bioorganic. Chem. 2021, 114, 105115. [Google Scholar] [CrossRef]
- Horishita, R.; Ogata, Y.; Fukui, R.; Yamazaki, R.; Moriwaki, K.; Ueno, S.; Yanagihara, N.; Uezono, Y.; Yokoyama, Y.; Minami, K.; et al. Local Anesthetics Inhibit Transient Receptor Potential Vanilloid Subtype 3 Channel Function in Xenopus Oocytes. Obstet. Anesth. Dig. 2021, 132, 1756–1767. [Google Scholar] [CrossRef]
- Liu, Q.; Wang, J.; Wei, X.; Hu, J.; Ping, C.; Gao, Y.; Xie, C.; Wang, P.; Cao, P.; Cao, Z.; et al. Therapeutic inhibition of keratinocyte TRPV3 sensory channel by local anesthetic dyclonine. eLife 2021, 10, e68128. [Google Scholar] [CrossRef]
- Neuberger, A.; Nadezhdin, K.D.; Sobolevsky, A.I. Structural mechanism of trpv3 channel inhibition by the anesthetic dyclonine. Nat. Commun. 2022, 13, 2795. [Google Scholar] [CrossRef]
- Chong, J.A.; Fanger, C.; Larsen, G.R.; Lumma, W.C.; Moran, M.M.; Ripka, A.; Underwood, D.J.; Weigele, M.; Zhen, X. Compounds for Modulating trpv3 Function. Patent WO-2006122156-A2, 9 May 2005. [Google Scholar]
- Grubisha, O.; Mogg, A.J.; Sorge, J.L.; Ball, L.-J.; Sanger, H.; Ruble, C.L.A.; Folly, E.A.; Ursu, D.; Broad, L.M. Pharmacological profiling of the TRPV3 channel in recombinant and native assays. Br. J. Pharmacol. 2014, 171, 2631–2644. [Google Scholar] [CrossRef] [Green Version]
- Moran, M.M.; Chong, J.A.; Fanger, C.; Ripka, A.; Larsen, G.R.; Zhen, X.; Underwood, D.J.; Weigele, M. Compounds for Modulating trpv3 Function. Patent WO-2007056124, 3 November 2006. [Google Scholar]
- Prasadarao, L.V.S.; Abraham, T.; Anandrao, M.D.; Yusuf, K.J.; Neelima, K.J.; Ganapati, K.V. Fused Pyrimidine Derivatives as trpv3 Modulators. Patent US-20100292254, 18 November 2010. [Google Scholar]
- Lingam, V.; Thomas, A.; More, D.A.; Khatik, J.Y.; Khairatkar-Joshi, N.; Kattige, V.G. Fused Pyrimidine Derivatives as trpv3 Modulators. Patent US-20090286811, 19 November 2009. [Google Scholar]
- Lingam, V.; Thomas, A.; Khatik, J.Y.; Khairatkar-Joshi, N.; Kattige, V.G. Chromane Derivatives as trpv3 Modulators. U.S. Patent 8,487,121, 16 July 2013. [Google Scholar]
- Chaudhari, S.S.; Thomas, A.; Kadam, A.B.; Dhone, S.V.; Adik, B.G.; Khairatkar-Joshi, N.; Kattige, V.G. Chromenone Derivatives as trpv3 Antagonists. Patent WO-2010055384, 20 May 2010. [Google Scholar]
- Sunarlal, C.S.; Abraham, T.; Bhausaheb, K.A.; Vasantrao, D.S.; Mahadev, K.S.; Neelima, K.J.; Ganapati, K.V. Fused imidazole Carboxamides as trpv3 Modulators. Patent US-8318928-B2, 27 November 2012. [Google Scholar]
- Prasadarao, L.V.S.; Abraham, T.; Kashinath, P.S.; Shankar, M.A.; Yusuf, K.J.; Neelima, K.J.; Ganapati, K.V. Fused Imidazole Derivatives as trpv3 Antagonist. Patent WO-2010073128, 1 July 2010. [Google Scholar]
- Gomtsyan, A.R.; Schmidt, R.G.; Bayburt, E.K.; Gfesser, G.A.; Voight, E.A.; Daanen, J.F.; Schmidt, D.L.; Cowart, M.D.; Liu, H.; Altenbach, R.J. Synthesis and pharmacology of (pyridin-2-yl)methanol derivatives as novel and selective transient receptor potential vanilloid 3 (trpv3) antagonists. J. Med. Chem. 2016, 59, 4926–4947. [Google Scholar] [CrossRef] [PubMed]
- Ortar, G.; Morera, L.; Moriello, A.S.; Morera, E.; Nalli, M.; Di Marzo, V.; De Petrocellis, L. Modulation of thermo-transient receptor potential (thermo-TRP) channels by thymol-based compounds. Bioorganic Med. Chem. Lett. 2012, 22, 3535–3539. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.H.D.; Itoh, S.G.; Okumura, H.; Tominaga, M. Structural basis for promiscuous action of monoterpenes on TRP channels. Commun. Biol. 2021, 4, 293. [Google Scholar] [CrossRef] [PubMed]
- Sherkheli, M.A.; Schreiner, B.; Haq, R.U.; Werner, M.; Hatt, H. Borneol inhibits TRPA1, a proinflammatory and noxious pain-sensing cation channel. Pak. J. Pharm. Sci. 2015, 28, 1357–1363. [Google Scholar] [PubMed]
- Wang, S.; Zhang, D.; Hu, J.; Jia, Q.; Xu, W.; Su, D.; Song, H.; Xu, Z.; Cui, J.; Zhou, M.; et al. A clinical and mechanistic study of topical borneol-induced analgesia. EMBO Mol. Med. 2017, 9, 802–815. [Google Scholar] [CrossRef]
- Macpherson, L.J.; Hwang, S.W.; Miyamoto, T.; Dubin, A.E.; Patapoutian, A.; Story, G.M. More than cool: Promiscuous relationships of menthol and other sensory compounds. Mol. Cell Neurosci. 2006, 32, 335–343. [Google Scholar] [CrossRef]
- McKemy, D.D. TRPM8: The Cold and Menthol Receptor. In TRP Ion Channel Function in Sensory Transduction and Cellular Signaling Cascades; Liedtke, W.B., Heller, S., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2007; Chapter 13; ISBN 978-0-8493-4048-2. [Google Scholar]
- Earley, S.; Gonzales, A.L.; Garcia, Z.I. A dietary agonist of transient receptor potential cation channel v3 elicits endothelium-dependent vasodilation. Mol. Pharmacol. 2010, 77, 612–620. [Google Scholar] [CrossRef] [Green Version]
- Ben, A.M. Cannabinoids in medicine: A review of their therapeutic potential. J. Ethnopharmacol. 2006, 105, 1–25. [Google Scholar]
- De Petrocellis, L.; Ligresti, A.; Moriello, A.S.; Allara, M.; Bisogno, T.; Petrosino, S.; Stott, C.G.; Di Marzo, V. Effects of canna-binoids and cannabinoid-enriched cannabis extracts on trp channels and endocannabinoid metabolic enzymes. Br. J. Phamacol. 2011, 163, 1479–1494. [Google Scholar] [CrossRef] [Green Version]
- Ligresti, A.; Moriello, A.S.; Starowicz, K.; Matias, I.; Pisanti, S.; De Petrocellis, L.; Laezza, C.; Portella, G.; Bifulco, M.; Di Marzo, V. Antitumor Activity of Plant Cannabinoids with Emphasis on the Effect of Cannabidiol on Human Breast Carcinoma. Experiment 2006, 318, 1375–1387. [Google Scholar] [CrossRef] [Green Version]
- Yang, N.-N.; Shi, H.; Yu, G.; Wang, C.-M.; Zhu, C.; Yang, Y.; Yuan, X.-L.; Tang, M.; Wang, Z.-L.; Gegen, T.; et al. Osthole inhibits histamine-dependent itch via modulating TRPV1 activity. Sci. Rep. 2016, 6, 25657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bang, S.; Yoo, S.; Yang, T.; Cho, H.; Hwang, S. 17(R)-resolvin D1 specifically inhibits transient receptor potential ion channel vanilloid 3 leading to peripheral antinociception. Br. J. Pharmacol. 2011, 165, 683–692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bang, S.; Yoo, S.; Yang, T.-J.; Cho, H.; Hwang, S.W. Isopentenyl pyrophosphate is a novel antinociceptive substance that inhibits TRPV3 and TRPA1 ion channels. Pain 2011, 152, 1156–1164. [Google Scholar] [CrossRef] [PubMed]


| Diseases | Research Evidence | Reference(s) |
|---|---|---|
| Olmsted syndrome (OS) | A series of independent clinical reports are identified mutations in the TRPV3 gene as the cause of Olmsted’s syndrome. | [37,38,39,40,41,42] |
| Pruritic and atopic dermatitis (AD) | 1. The expression of TRPV3 is significantly increased in skin lesions and non-diseased skin of patients with specific dermatitis. 2. The pharmacological activation of TRPV3 leads to the development of AD in wild-type mice, and it has no effect on TRPV3 knockout mice. 3. The inhibition of TRPV3 treats inflammatory dorsal skin in a dose-dependent manner. | [43,44,45] |
| Psoriasis | 1. The expression level of TRPV3 in psoriasis patients is significantly higher than it is in people without psoriasis. 2. TRPV3 antagonists relieve the symptoms in patients with moderate-to-severe psoriasis in a dose-dependent manner. | [46] |
| Cutaneous pruritus | 1. TRPV3 knockout mice show no increase in scratching behavior after itch modeling. 2. The expression of TRPV3 is significantly up-regulated in the epidermis of patients with pruritus after having been burnt. | [47,48] |
| Rosacea | TRPV3 gene expression is significantly increased in rosacea. | [49] |
| Cancer | 1. TRPV3 expression is increased in pancreatic, bone, breast, lung and oral squamous cell cancers. 2. TRPV3 expression is decreased in colorectal cancer. | [21,50,51,52] |
| Myocardial hypertrophy | 1. The expression of TRPV3 is increased in pathological cardiac hypertrophy. 2. TRPV3 expression is increased in cardiomyocyte hypertrophy induced by Ang-II in vitro. 3. TRPV3 inhibitors can significantly aggravate cardiomyocyte hypertrophy and TRPV3 antagonists can slow cardiomyocyte hypertrophy. | [53] |
| Cardiac fibrosis | TRPV3 activation exacerbate cardiac dysfunction and interstitial fibrosis in pressure-overloaded rats. | [54] |
| Myocardial infarction | The expression of TRPV3 is significantly up-regulated in neonatal rat cardiomyocytes after myocardial infarction and in hypoxia-treated rats. | [55] |
| Pain | TRPV3 is overexpressed in skin pain, breast pain, cancer pain and other pain-experiencing tissues. | [56,57,58] |
| Alopecia | TRPV3 agonists inhibit hair growth, whereas TRPV3 inhibitors significantly reverse the symptom. | [59] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, W.; Qiao, X.; Wang, W.; He, S.; Liang, K.; Hong, X. TRPV3: Structure, Diseases and Modulators. Molecules 2023, 28, 774. https://doi.org/10.3390/molecules28020774
Su W, Qiao X, Wang W, He S, Liang K, Hong X. TRPV3: Structure, Diseases and Modulators. Molecules. 2023; 28(2):774. https://doi.org/10.3390/molecules28020774
Chicago/Turabian StyleSu, Wuyue, Xue Qiao, Wumei Wang, Shengnan He, Ke Liang, and Xuechuan Hong. 2023. "TRPV3: Structure, Diseases and Modulators" Molecules 28, no. 2: 774. https://doi.org/10.3390/molecules28020774
APA StyleSu, W., Qiao, X., Wang, W., He, S., Liang, K., & Hong, X. (2023). TRPV3: Structure, Diseases and Modulators. Molecules, 28(2), 774. https://doi.org/10.3390/molecules28020774