
Sinomenine Hydrochloride Can Ameliorate Benign Prostatic Hyperplasia by Lowering the 5α-Reductase 2 Level and Regulating the Balance between the Proliferation and Apoptosis of Cells
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of SIN Treatment on the Apoptosis of BPH-1 Cells
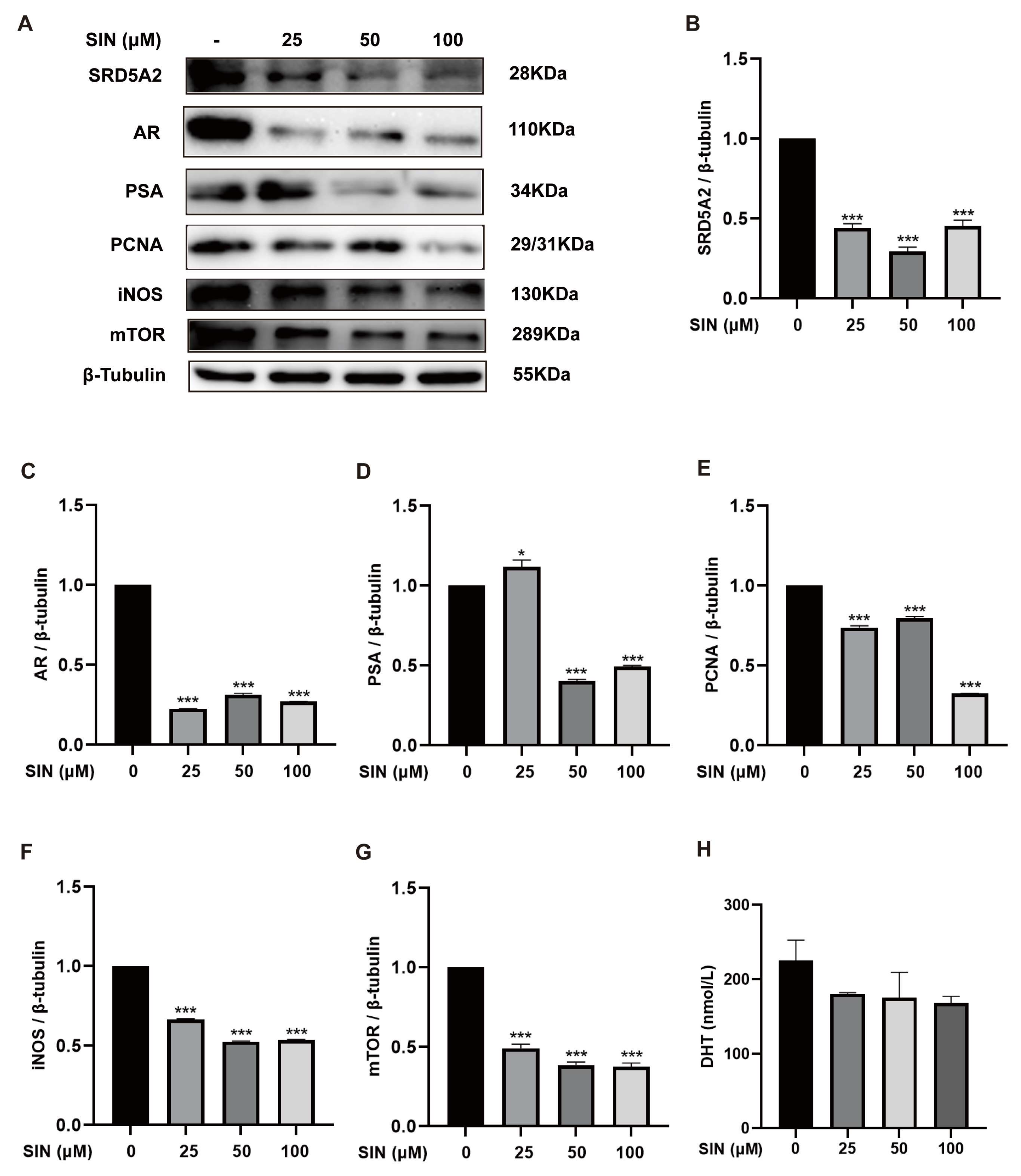
2.2. Effect of SIN Treatment on the AR Signaling Pathway in BPH-1 Cells
2.3. Effects of SIN Therapy on Prostatic Enlargement in Mice with TP-Induced BPH
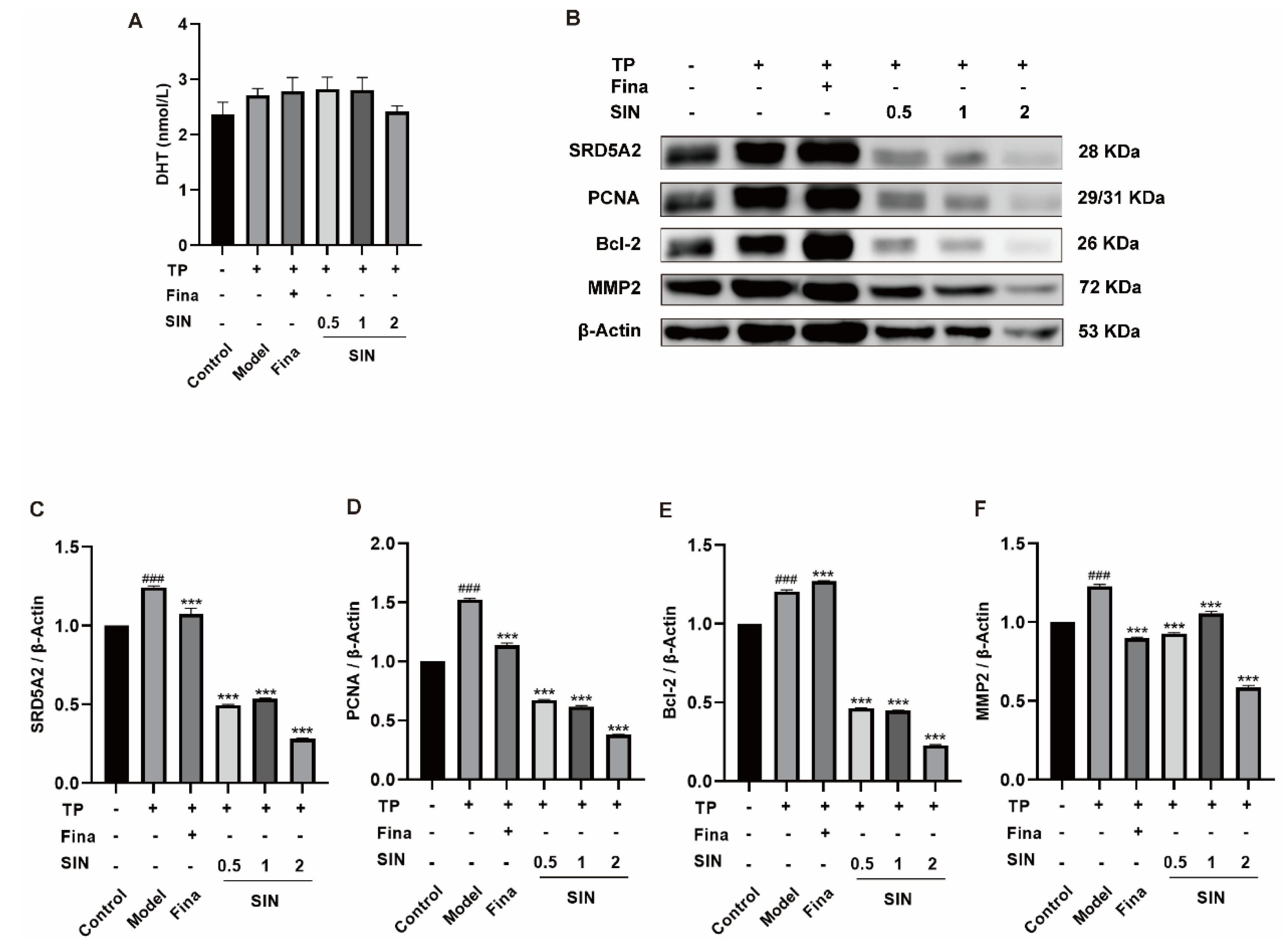
2.4. Effects of SIN Treatment on Protein Expression in the PG Tissue of Mice with TP-Induced BPH
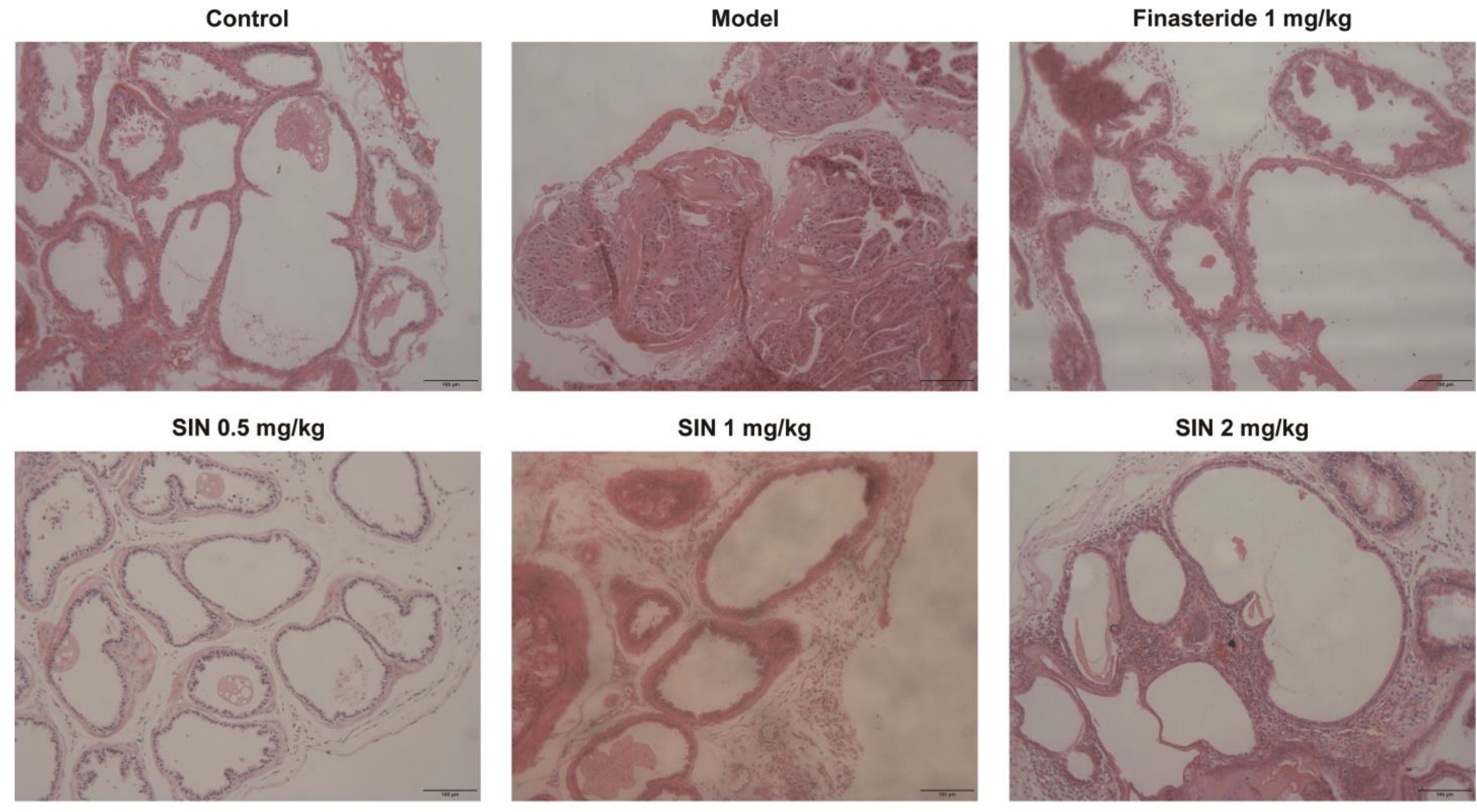
2.5. Effects of SIN Therapy on Histological Alterations of PG Tissue in Mice with TP-Induced BPH
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Cell Culture
4.3. Cell-Viability Assay
4.4. Staining
4.5. Western Blotting
4.6. ELISA Measurements in Cell Supernatants
4.7. Animals
4.8. Mouse Model of BPH
4.9. Histology
4.10. Statistical Analyses
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Foo, K.T. What is a disease? What is the disease clinical benign prostatic hyperplasia (BPH)? World J. Urol. 2019, 37, 1293–1296. [Google Scholar] [CrossRef] [Green Version]
- Devlin, C.M.; Simms, M.S.; Maitland, N.J. Benign prostatic hyperplasia—What do we know? BJU Int. 2021, 127, 389–399. [Google Scholar] [CrossRef] [PubMed]
- Jin, B.R.; An, H.J. Baicalin alleviates benign prostate hyperplasia through androgen-dependent apoptosis. Aging 2020, 12, 2142–2155. [Google Scholar] [CrossRef] [PubMed]
- Van Asseldonk, B.; Barkin, J.; Elterman, D.S. Medical therapy for benign prostatic hyperplasia: A review. Can. J. Urol. 2015, 22 (Suppl. 1), 7–17. [Google Scholar]
- Rittmaster, R.S.; Norman, R.W.; Thomas, L.N.; Rowden, G. Evidence for atrophy and apoptosis in the prostates of men given finasteride. J. Clin. Endocrinol. Metab. 1996, 81, 814–819. [Google Scholar]
- Michel, M.C.; Vrydag, W. Alpha1-, alpha2- and beta-adrenoceptors in the urinary bladder, urethra and prostate. Br. J. Pharmacol. 2006, 147 (Suppl. 2), S88–S119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, I.M.; Goodman, P.J.; Tangen, C.M.; Lucia, M.S.; Miller, G.J.; Ford, L.G.; Lieber, M.M.; Cespedes, R.D.; Atkins, J.N.; Lippman, S.M.; et al. The influence of finasteride on the development of prostate cancer. N. Engl. J. Med. 2003, 349, 215–224. [Google Scholar] [CrossRef]
- Lepor, H. Alpha blockers for the treatment of benign prostatic hyperplasia. Rev. Urol. 2007, 9, 181–190. [Google Scholar] [CrossRef] [Green Version]
- Milani, S.; Djavan, B. Lower urinary tract symptoms suggestive of benign prostatic hyperplasia: Latest update on alpha-adrenoceptor antagonists. BJU Int. 2005, 95 (Suppl. 4), 29–36. [Google Scholar] [CrossRef]
- Ooi, S.L.; Pak, S.C. Serenoa repens for Lower Urinary Tract Symptoms/Benign Prostatic Hyperplasia: Current Evidence and Its Clinical Implications in Naturopathic Medicine. J. Altern. Complement. Med. 2017, 23, 599–606. [Google Scholar] [CrossRef]
- Han, N.; Zhang, B.; Wei, X.; Yu, L. The inhibitory function of icariin in cell model of benign prostatic hyperplasia by upregulation of miR-7. BioFactors, 2019; undefined. [Google Scholar] [CrossRef] [PubMed]
- Vásquez-Velásquez, C.; Gasco, M.; Fano-Sizgorich, D.; Gonzales, G.F. Inflammatory pathway employed by Red Maca to treat induced benign prostatic hyperplasia in rats. Andrologia 2020, 52, e13516. [Google Scholar] [CrossRef] [PubMed]
- Rho, J.; Seo, C.S.; Park, H.S.; Jeong, H.Y.; Moon, O.S.; Seo, Y.W.; Son, H.Y.; Won, Y.S.; Kwun, H.J. Asteris Radix et Rhizoma suppresses testosterone-induced benign prostatic hyperplasia in rats by regulating apoptosis and inflammation. J. Ethnopharmacol. 2020, 255, 112779. [Google Scholar] [CrossRef] [PubMed]
- Huang, G.; He, X.; Xue, Z.; Long, Y.; Liu, J.; Cai, J.; Tang, P.; Han, B.; Shen, B.; Huang, R.; et al. Rauwolfia vomitoria extract suppresses benign prostatic hyperplasia by inducing autophagic apoptosis through endoplasmic reticulum stress. BMC Complement. Med. Ther. 2022, 22, 125. [Google Scholar] [CrossRef]
- Xu, M.; Liu, L.; Qi, C.; Deng, B.; Cai, X. Sinomenine versus NSAIDs for the treatment of rheumatoid arthritis: A systematic review and meta-analysis. Planta Med. 2008, 74, 1423–1429. [Google Scholar] [CrossRef]
- Choi, H.M.; Jung, Y.; Park, J.; Kim, H.L.; Youn, D.H.; Kang, J.; Jeong, M.Y.; Lee, J.H.; Yang, W.M.; Lee, S.G.; et al. Cinnamomi Cortex (Cinnamomum verum) Suppresses Testosterone-induced Benign Prostatic Hyperplasia by Regulating 5α-reductase. Sci. Rep. 2016, 6, 31906. [Google Scholar] [CrossRef] [Green Version]
- Andriole, G.; Bruchovsky, N.; Chung, L.W.; Matsumoto, A.M.; Rittmaster, R.; Roehrborn, C.; Russell, D.; Tindall, D. Dihydrotestosterone and the prostate: The scientific rationale for 5alpha-reductase inhibitors in the treatment of benign prostatic hyperplasia. J. Urol. 2004, 172 Pt 1, 1399–1403. [Google Scholar] [CrossRef]
- Thigpen, A.E.; Silver, R.I.; Guileyardo, J.M.; Casey, M.L.; McConnell, J.D.; Russell, D.W. Tissue distribution and ontogeny of steroid 5 alpha-reductase isozyme expression. J. Clin. Investig. 1993, 92, 903–910. [Google Scholar] [CrossRef]
- Levitt, J.M.; Slawin, K.M. Prostate-specific antigen and prostate-specific antigen derivatives as predictors of benign prostatic hyperplasia progression. Curr. Urol. Rep. 2007, 8, 269–274. [Google Scholar] [CrossRef]
- Zhong, X.Y.; Lin, J.M.; Zhou, J.H.; Xu, W.; Hong, Z.F.; Peng, J. Qianliening capsule treats benign prostatic hyperplasia (BPH) by down-egulating the expression of PCNA, CyclinD1 and CDK4. Afr. J. Biotechnol. 2012, 11, 7731–7737. [Google Scholar]
- Xu, C.; Sun, G.; Yuan, G.; Wang, R.; Sun, X. Effects of platycodin D on proliferation, apoptosis and PI3K/Akt signal pathway of human glioma U251 cells. Molecules 2014, 19, 21411–21423. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Kim, B.H.; Jin, B.R.; Park, S.J.; An, H.J. Purple Corn Extract Improves Benign prostatic hyperplasia by Regulating Prostate Cell Proliferation and Apoptosis. J. Agric. Food Chem. 2022, 70, 5561–5569. [Google Scholar] [CrossRef]
- Karunasagara, S.; Hong, G.L.; Jung, D.Y.; Kim, K.H.; Cho, K.; Jung, J.Y. Protective effects of combination of Stauntonia hexaphylla and Cornus officinalis on testosterone-induced benign prostatic hyperplasia through inhibition of 5α- reductase type 2 and induced cell apoptosis. PLoS ONE 2020, 15, e0236879. [Google Scholar] [CrossRef]
- Vignozzi, L.; Rastrelli, G.; Corona, G.; Gacci, M.; Forti, G.; Maggi, M. Benign prostatic hyperplasia: A new metabolic disease? J. Endocrinol. Investig. 2014, 37, 313–322. [Google Scholar] [CrossRef]
- La Vignera, S.; Condorelli, R.A.; Russo, G.I.; Morgia, G.; Calogero, A.E. Endocrine control of benign prostatic hyperplasia. Andrology 2016, 4, 404–411. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.Y.; Kim, S.; Kim, S.; Kim, J.H.; Bae, B.S.; Koo, G.B.; So, S.H.; Lee, J.; Lee, Y.H. Effects of red ginseng oil (KGC11o) on testosterone-propionate-induced benign prostatic hyperplasia. J. Ginseng. Res. 2022, 46, 473–480. [Google Scholar] [CrossRef]
- Wang, Z.; Jia, G.; Li, Y.; Liu, J.; Luo, J.; Zhang, J.; Xu, G.; Chen, G. Clinicopathological signature of p21-activated kinase 1 in prostate cancer and its regulation of proliferation and autophagy via the mTOR signaling pathway. Oncotarget 2017, 8, 22563–22580. [Google Scholar] [CrossRef] [Green Version]
- Swanton, E.; Savory, P.; Cosulich, S.; Clarke, P.; Woodman, P. Bcl-2 regulates a caspase-3/caspase-2 apoptotic cascade in cytosolic extracts. Oncogene 1999, 18, 1781–1787. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Ren, F.; Song, Y. Upregulation of SPOCK2 inhibits the invasion and migration of prostate cancer cells by regulating the MT1-MMP/MMP2 pathway. Peer J. 2019, 7, e7163. [Google Scholar] [CrossRef]
- Sun, Z.R.; Liu, H.R.; Hu, D.; Fan, M.S.; Wang, M.Y.; An, M.F.; Zhao, Y.L.; Xiang, Z.M.; Sheng, J. Ellagic Acid Exerts Beneficial Effects on Hyperuricemia by Inhibiting Xanthine Oxidase and NLRP3 Inflammasome Activation. J. Agric. Food Chem. 2021, 69, 12741–12752. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fan, M.-S.; Xia, Y.-F.; Ye, R.-H.; Sun, Z.-R.; Wang, M.-Y.; An, M.-F.; Zhang, S.-S.; Zhang, L.-J.; Zhao, Y.-L.; Xiang, Z.-M.; et al. Sinomenine Hydrochloride Can Ameliorate Benign Prostatic Hyperplasia by Lowering the 5α-Reductase 2 Level and Regulating the Balance between the Proliferation and Apoptosis of Cells. Molecules 2023, 28, 803. https://doi.org/10.3390/molecules28020803
Fan M-S, Xia Y-F, Ye R-H, Sun Z-R, Wang M-Y, An M-F, Zhang S-S, Zhang L-J, Zhao Y-L, Xiang Z-M, et al. Sinomenine Hydrochloride Can Ameliorate Benign Prostatic Hyperplasia by Lowering the 5α-Reductase 2 Level and Regulating the Balance between the Proliferation and Apoptosis of Cells. Molecules. 2023; 28(2):803. https://doi.org/10.3390/molecules28020803
Chicago/Turabian StyleFan, Mao-Si, Yue-Fei Xia, Rui-Han Ye, Ze-Rui Sun, Ming-Yue Wang, Meng-Fei An, Shao-Shi Zhang, Li-Juan Zhang, Yun-Li Zhao, Ze-Min Xiang, and et al. 2023. "Sinomenine Hydrochloride Can Ameliorate Benign Prostatic Hyperplasia by Lowering the 5α-Reductase 2 Level and Regulating the Balance between the Proliferation and Apoptosis of Cells" Molecules 28, no. 2: 803. https://doi.org/10.3390/molecules28020803