
Effect of Salts on the Conformational Dynamics of the Cytochrome P450 OleP

 , , , , , , , , , ,
, , , , , , , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Equilibrium-Binding Analysis
2.3. Molecular Dynamics Simulations
2.4. Structural Analysis
3. Results
3.1. Divalent Cations Enhance the High-Spin State Subpopulation of the OleP-6DEB Complex
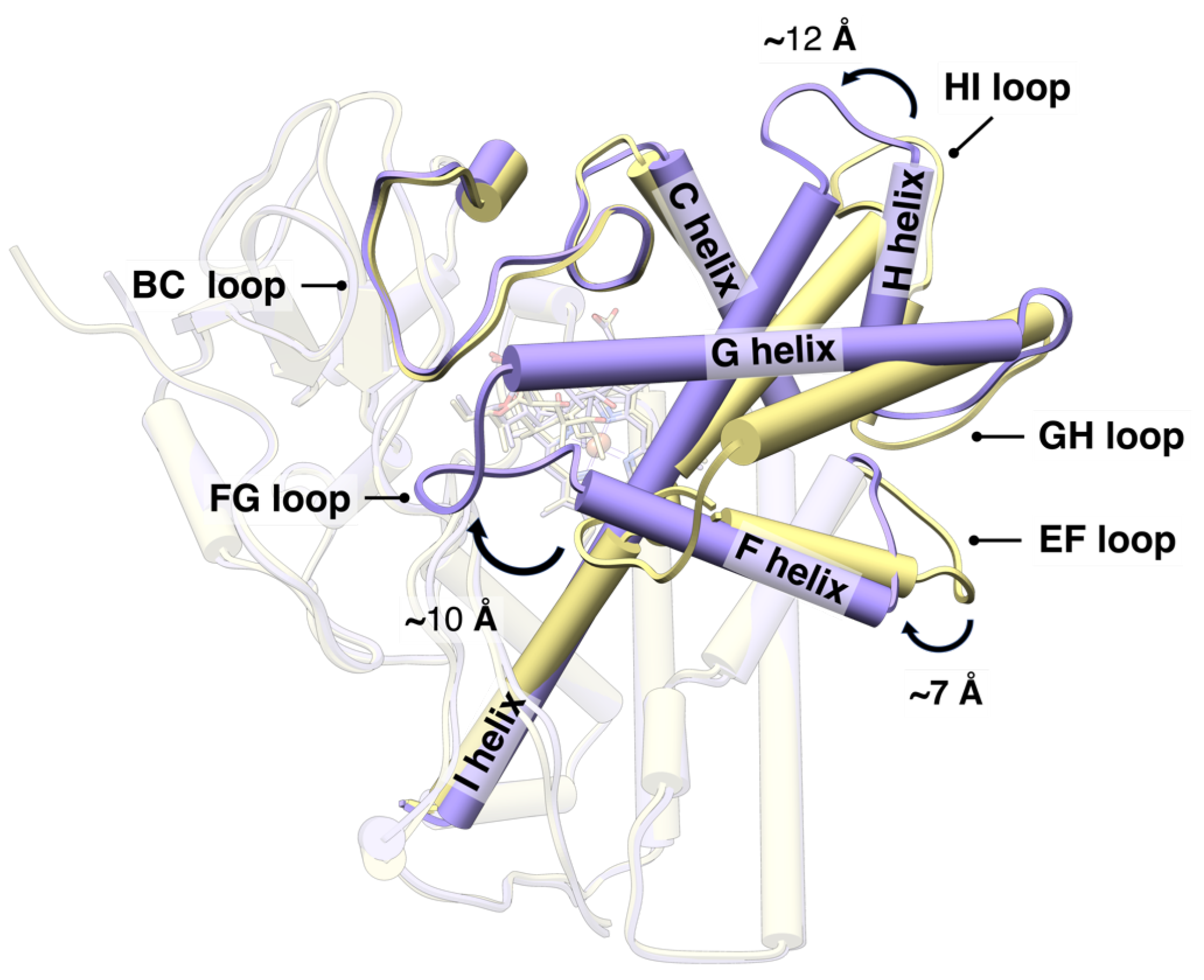
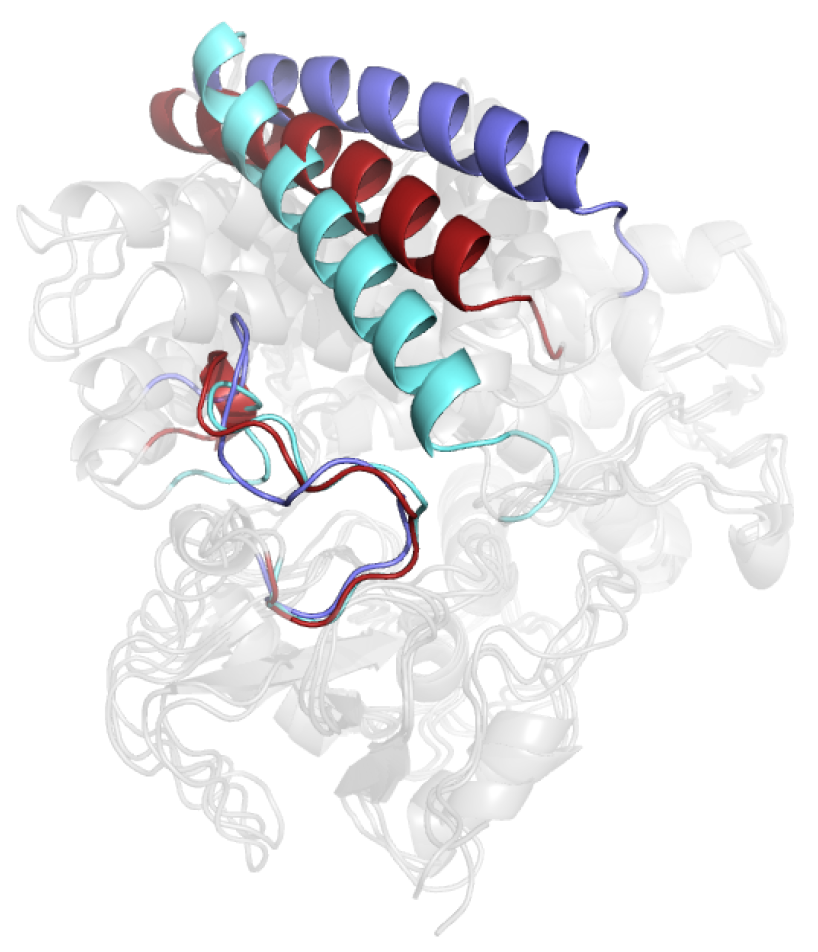
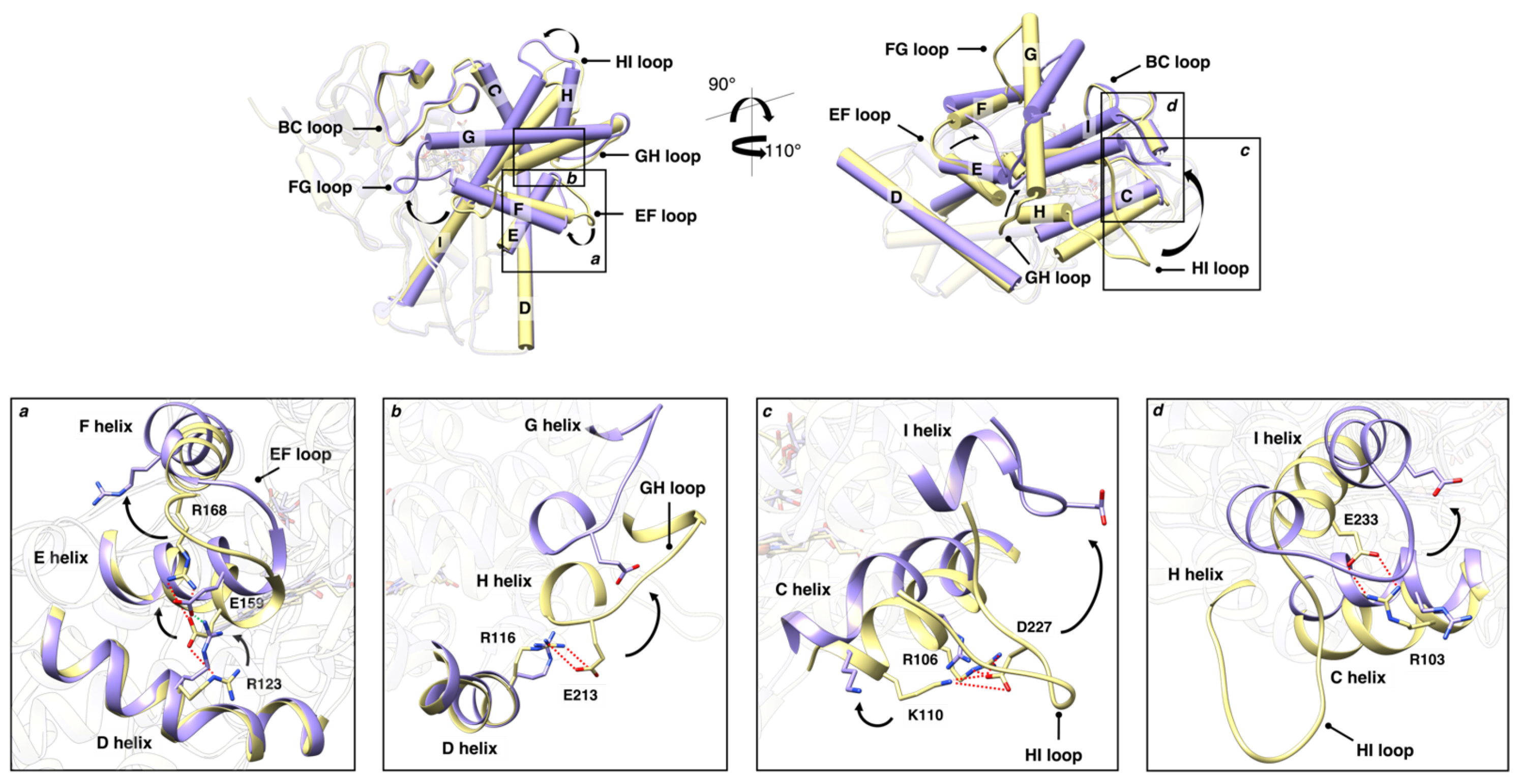
3.2. Structural Dynamic Analysis
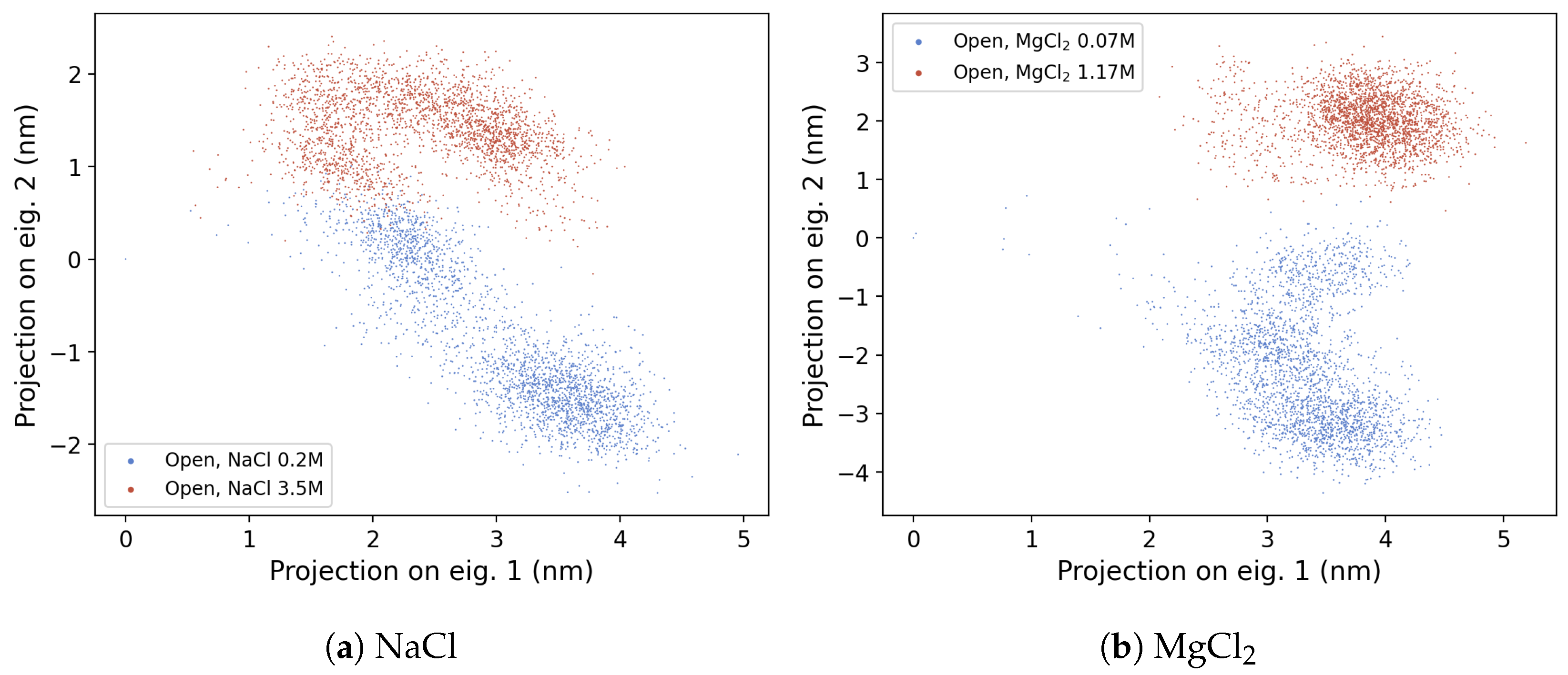
3.2.1. Principal Component Analysis
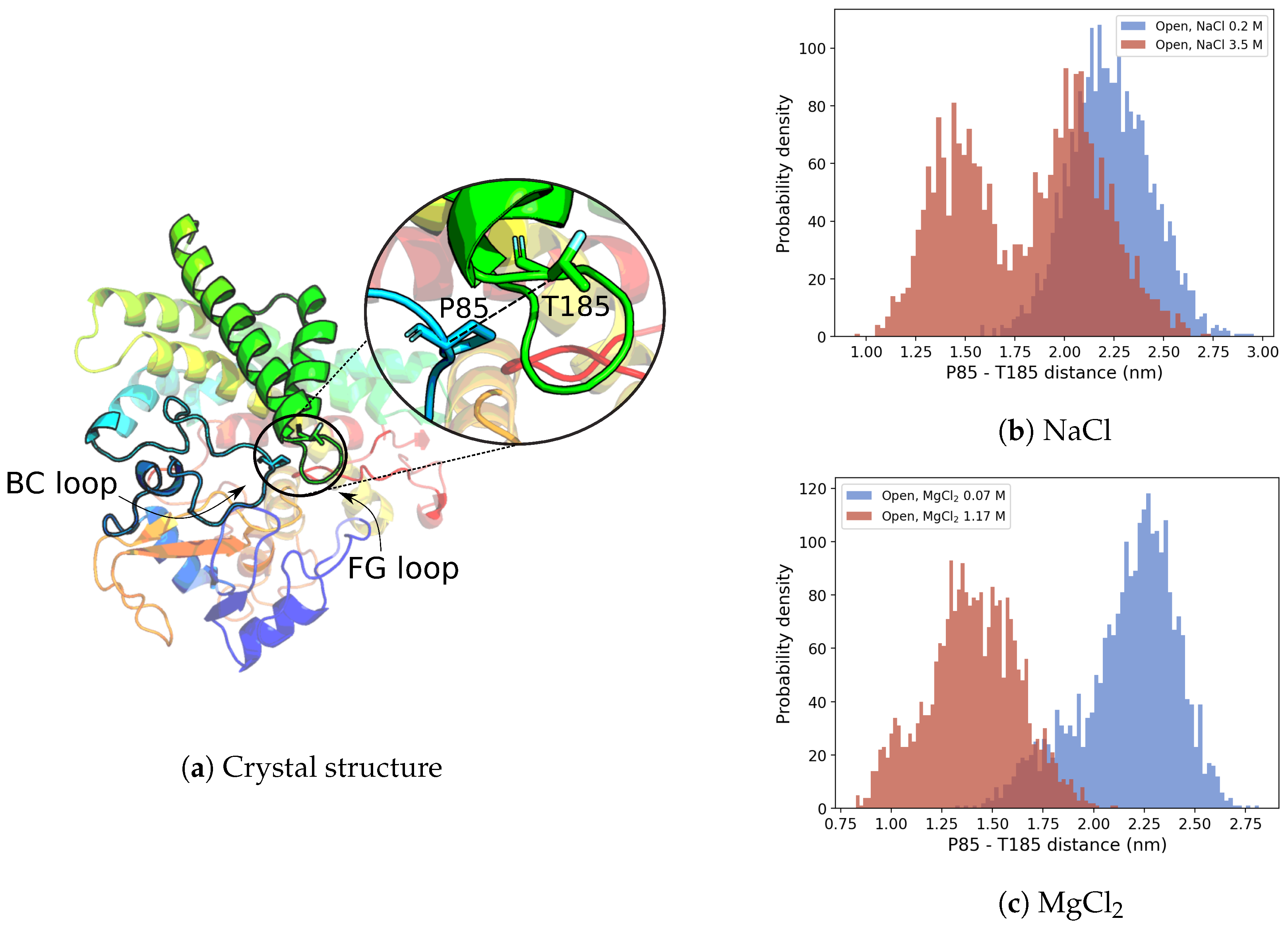
3.2.2. G-Helix–BC-Loop Distance
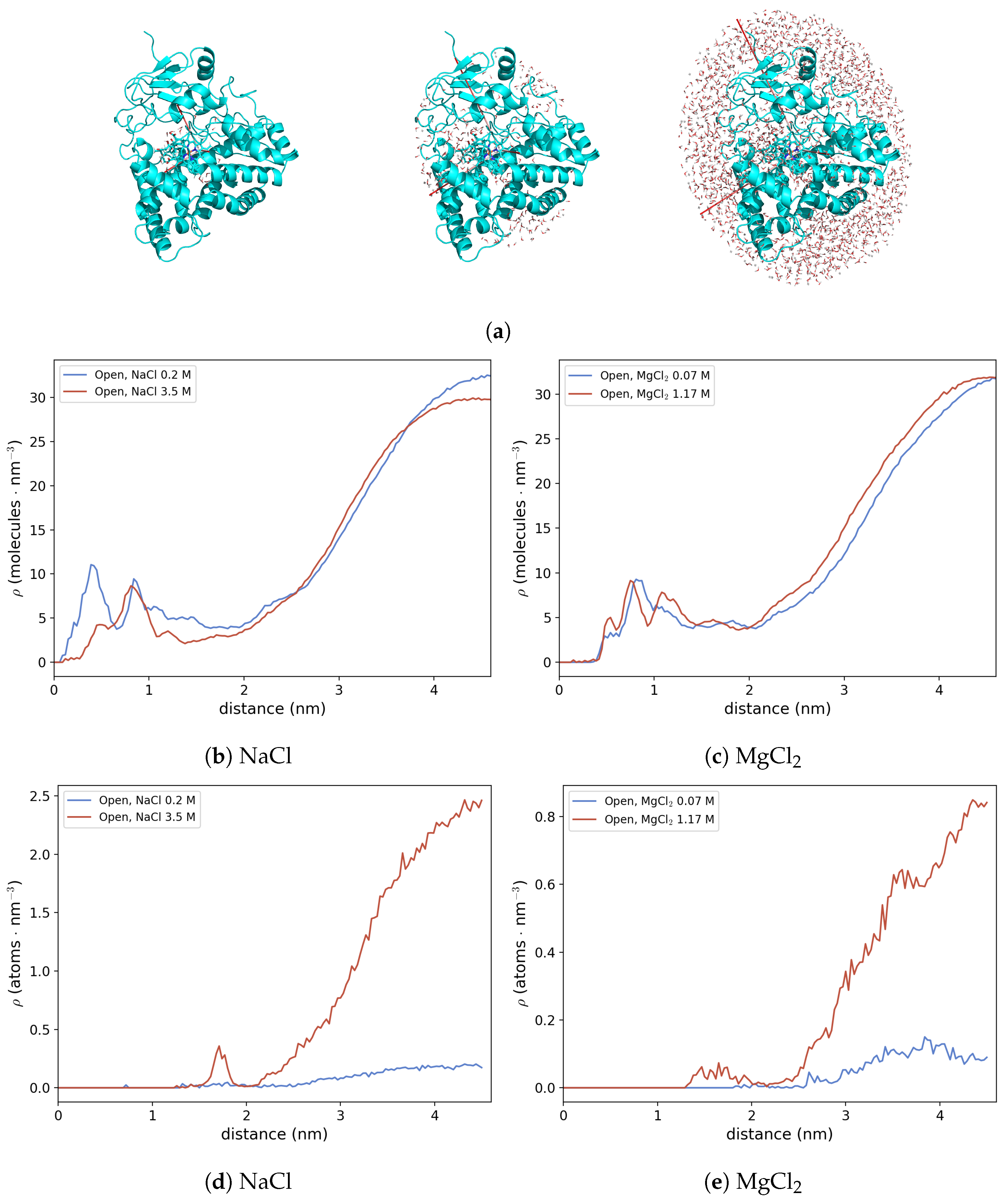
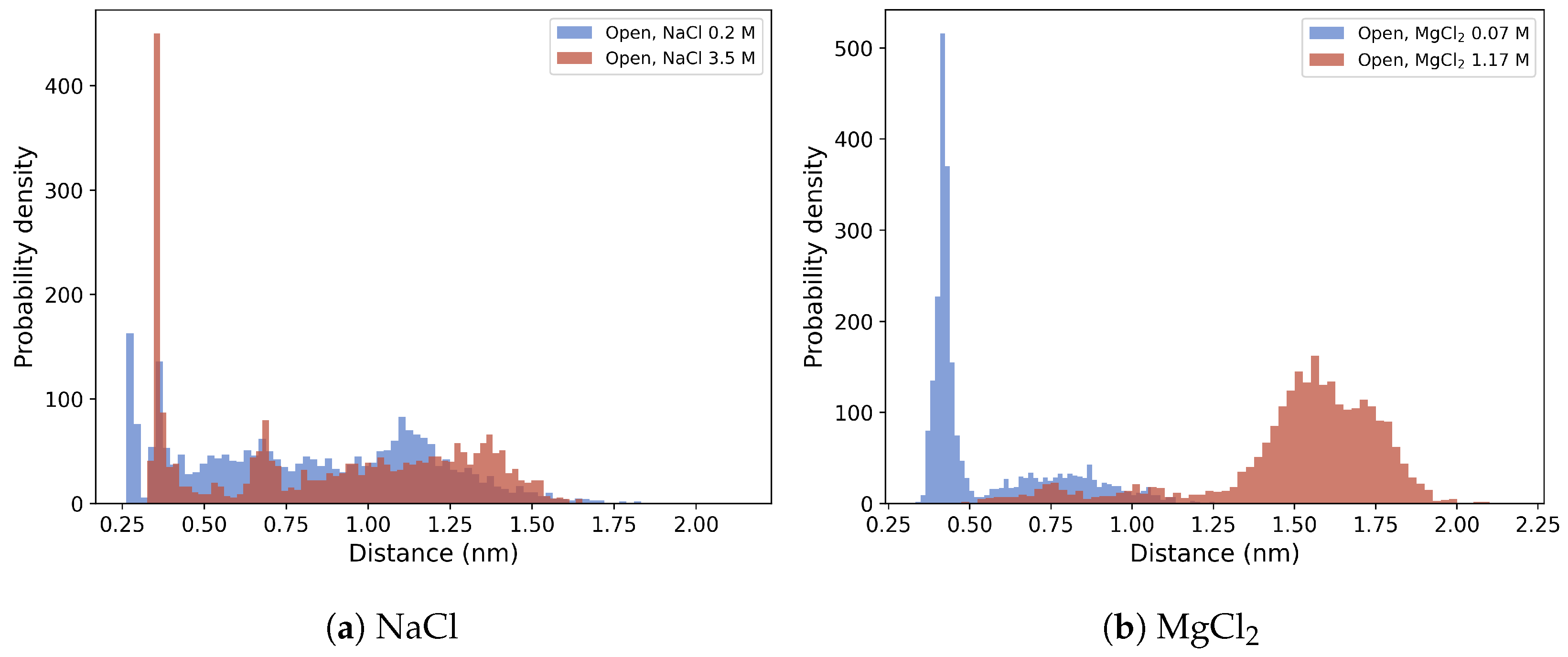
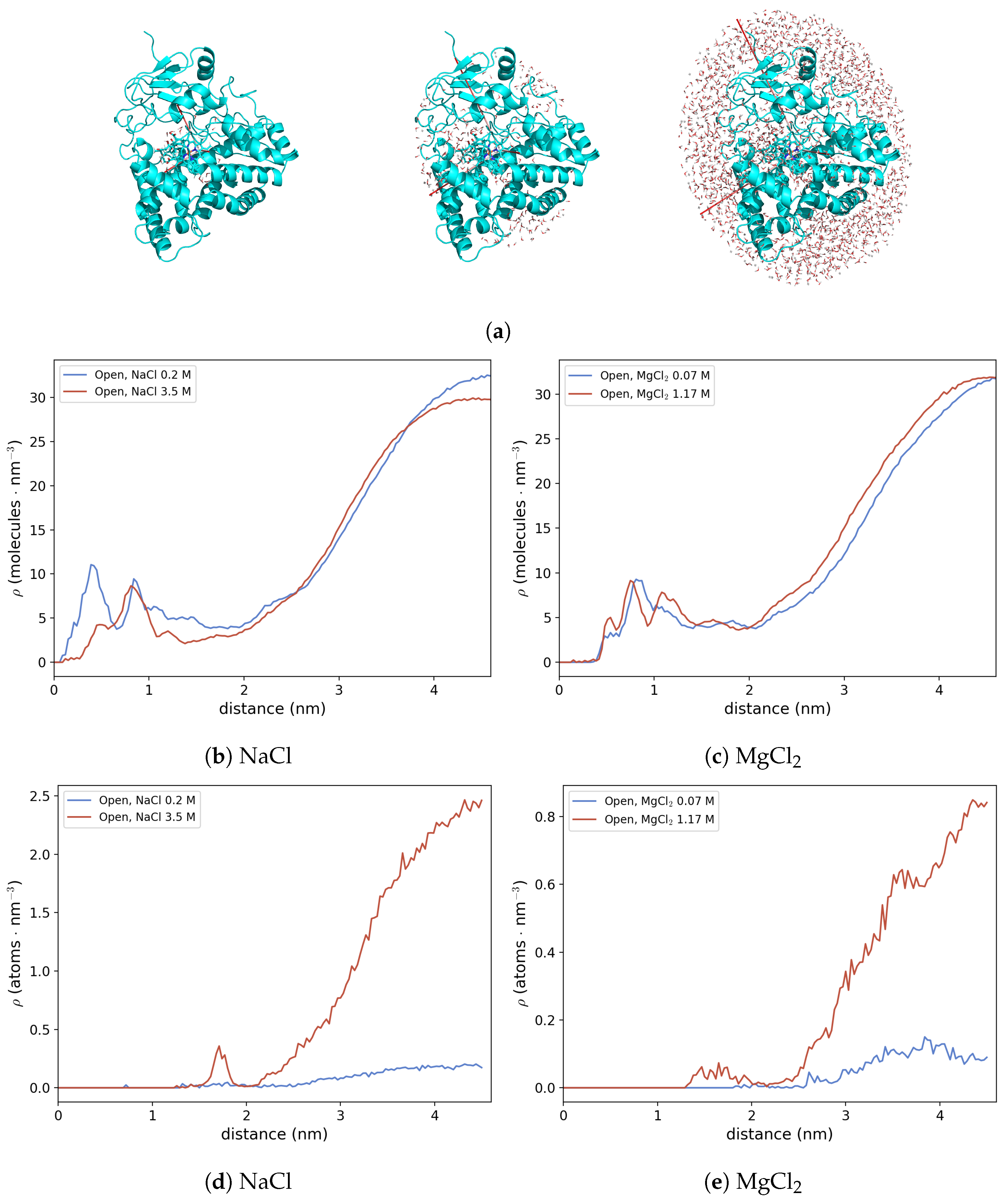
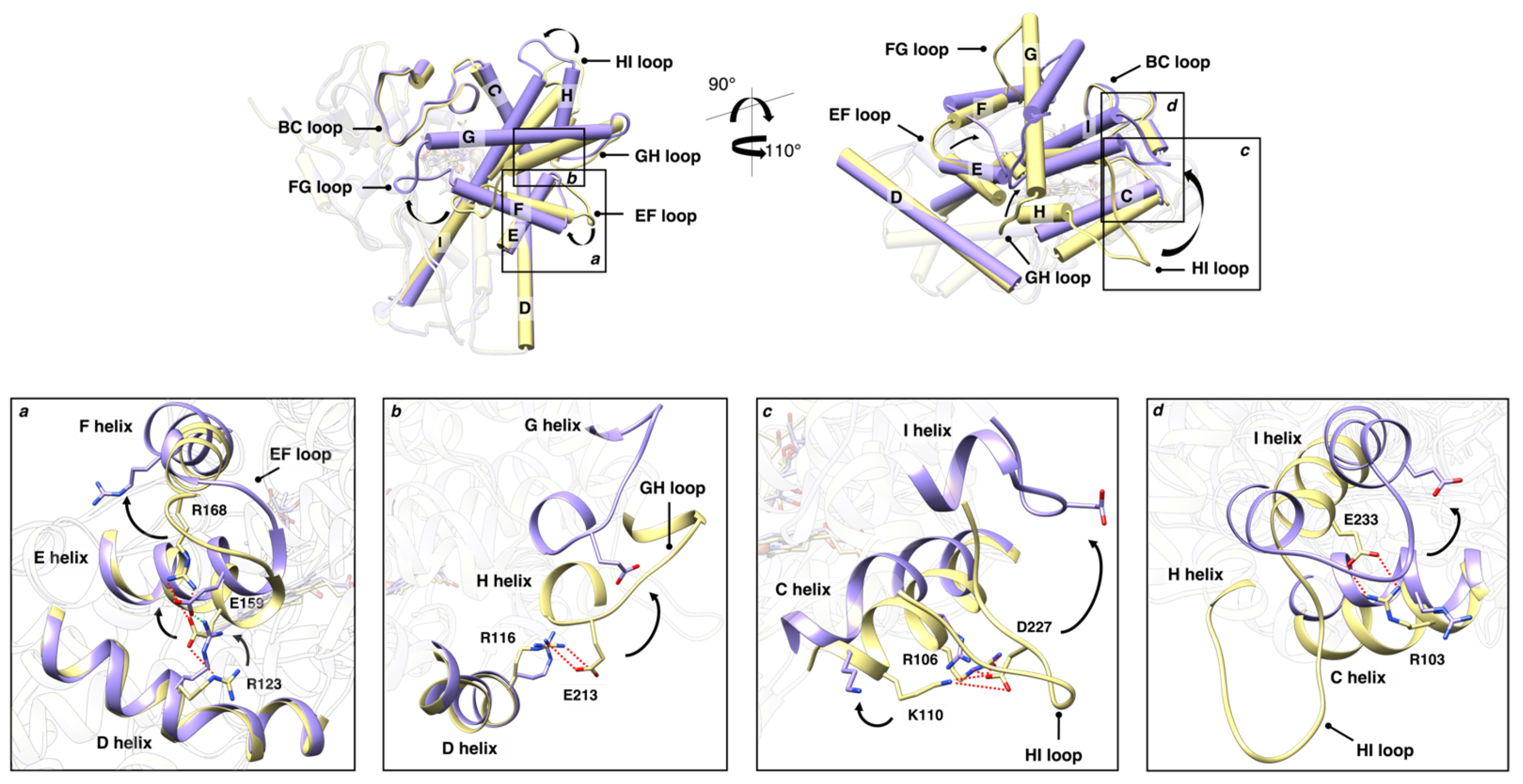
3.2.3. Protein Hydration Shell and Salt Bridges
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Shah, S.; Xue, Q.; Tang, L.; Carney, J.R.; Betlach, M.; Mcdaniel, R. Cloning, Characterization and Heterologous Expression of a Polyketide Synthase and P-450 Oxidase Involved in the Biosynthesis of the Antibiotic Oleandomycin. J. Antibiot. 2000, 53, 502–508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tatsuta, K.; Gunji, H.; Tajima, S.; Ishiyama, T.; Imai, S.; Okuyama, S.; Fukatsu, S. Biosynthetic studies on oleandomycin by incorporation of the chemically synthesized aglycones. J. Antibiot. 1990, 43, 909–911. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, A.M.; Olano, C.; Méndez, C.; Hutchinson, C.R.; Salas, J. A cytochrome P450-like gene possibly involved in oleandomycin biosynthesis by Streptomyces antibiot. FEMS Microbiol. Lett. 1995, 127, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Gaisser, S.; Lill, R.; Staunton, J.; Méndez, C.; Salas, J.; Leadlay, P.F. Parallel pathways for oxidation of 14-membered polyketide macrolactones in Saccharopolyspora erythraea. Mol. Microbiol. 2002, 44, 771–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parisi, G.; Montemiglio, L.C.; Giuffrè, A.; Macone, A.; Scaglione, A.; Cerutti, G.; Exertier, C.; Savino, C.; Vallone, B. Substrate-induced conformational change in cytochrome P450 OleP. FASEB J. 2019, 33, 1787–1800. [Google Scholar] [CrossRef] [PubMed]
- Montemiglio, L.C.; Parisi, G.; Scaglione, A.; Sciara, G.; Savino, C.; Vallone, B. Functional analysis and crystallographic structure of clotrimazole bound OleP, a cytochrome P450 epoxidase from Streptomyces antibioticus involved in oleandomycin biosynthesis. Biochim. Biophys. Acta 2016, 1860, 465–475. [Google Scholar] [CrossRef]
- Parisi, G.; Freda, I.; Exertier, C.; Cecchetti, C.; Gugole, E.; Cerutti, G.; D’Auria, L.; Macone, A.; Vallone, B.; Savino, C.; et al. Dissecting the Cytochrome P450 OleP Substrate Specificity: Evidence for a Preferential Substrate. Biomolecules 2020, 10, 1411. [Google Scholar] [CrossRef]
- Durham, B.; Fairris, J.L.; McLean, M.; Millett, F.; Scott, J.R.; Sligar, S.G.; Willie, A. Electron transfer from cytochromeb 5 to cytochromec. J. Bioenerg. Biomembr. 1995, 27, 331–340. [Google Scholar] [CrossRef]
- Meunier, B.; de Visser, S.P.; Shaik, S. Mechanism of Oxidation Reactions Catalyzed by Cytochrome P450 Enzymes. Chem. Rev. 2004, 104, 3947–3980. [Google Scholar] [CrossRef]
- Denisov, I.G.; Makris, T.M.; Sligar, S.G.; Schlichting, I. Structure and Chemistry of Cytochrome P450. Chem. Rev. 2005, 105, 2253–2278. [Google Scholar] [CrossRef]
- Montemiglio, L.C.; Gugole, E.; Freda, I.; Exertier, C.; D’Auria, L.; Chen, C.G.; Nardi, A.N.; Cerutti, G.; Parisi, G.; D’Abramo, M.; et al. Point Mutations at a Key Site Alter the Cytochrome P450 OleP Structural Dynamics. Biomolecules 2022, 12, 55. [Google Scholar] [CrossRef]
- Jung, C.; Ristau, O.; Rein, H. The high-spin/low-spin equilibrium in cytochrome P-450–a new method for determination of the high-spin content. Biochim. Biophys. Acta 1991, 1076, 130–136. [Google Scholar] [CrossRef] [PubMed]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1–2, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Best, R.B.; Zhu, X.; Shim, J.; Lopes, P.E.M.; Mittal, J.; Feig, M.; MacKerell, A.D.J. Optimization of the Additive CHARMM All-Atom Protein Force Field Targeting Improved Sampling of the Backbone φ, ψ and Side-Chain χ1 and χ2 Dihedral Angles. J. Chem. Theory Comput. 2012, 8, 3257–3273. [Google Scholar] [CrossRef] [Green Version]
- Soteras Gutiérrez, I.; Lin, F.Y.; Vanommeslaeghe, K.; Lemkul, J.A.; Armacost, K.A.; Brooks, C.L.; MacKerell, A.D. Parametrization of halogen bonds in the CHARMM general force field: Improved treatment of ligand–protein interactions. Bioorg. Med. Chem. 2016, 24, 4812–4825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mark, P.; Nilsson, L. Structure and dynamics of the TIP3P, SPC, and SPC/E water models at 298 K. J. Phys. Chem. A 2001, 105, 9954–9960. [Google Scholar] [CrossRef]
- Del Galdo, S.; Marracino, P.; D’Abramo, M.; Amadei, A. In silico characterization of protein partial molecular volumes and hydration shells. Phys. Chem. Chem. Phys. 2015, 17, 31270–31277. [Google Scholar] [CrossRef]
- Bussi, G.; Donadio, D.; Parrinello, M. Canonical sampling through velocity rescaling. J. Chem. Phys. 2007, 126, 014101. [Google Scholar] [CrossRef] [Green Version]
- Darden, T.; York, D.; Pedersen, L.G. Particle mesh Ewald: An N-log(N) method for Ewald sums in large systems. J. Chem. Phys. 1993, 98, 10089–10092. [Google Scholar] [CrossRef] [Green Version]
- Essmann, U.; Perera, L.; Berkowitz, M.L.; Darden, T.; Lee, H.; Pedersen, L.G. A smooth particle mesh Ewald method. J. Chem. Phys. 1995, 103, 8577–8593. [Google Scholar] [CrossRef]
- Daidone, I.; Amadei, A. Essential dynamics: Foundation and applications. Wiley Interdiscip. Rev. Comput. Mol. Sci. 2012, 2, 762–770. [Google Scholar] [CrossRef]
- Del Galdo, S.; Amadei, A. The unfolding effects on the protein hydration shell and partial molar volume: A computational study. Phys. Chem. Chem. Phys. 2016, 18, 28175–28182. [Google Scholar] [CrossRef]
- Isin, E.M.; Guengerich, F.P. Substrate binding to cytochromes P450. Anal. Bioanal. Chem. 2008, 392, 1019–1030. [Google Scholar] [CrossRef] [Green Version]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. Biochemistry (Loose-Leaf). MacMillan: London, UK, 2007. [Google Scholar]
- Luthra, A.; Denisov, I.G.; Sligar, S.G. Spectroscopic features of cytochrome P450 reaction intermediates. Arch. Biochem. Biophys. 2011, 507, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Wade, R.C.; Gabdoulline, R.R.; Lüdemann, S.K.; Lounnas, V. Electrostatic steering and ionic tethering in enzyme–ligand binding: Insights from simulations. Proc. Natl. Acad. Sci. USA 1998, 95, 5942–5949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deprez, E.; Gill, E.; Helms, V.; Wade, R.C.; Hoa, G.H.B. Specific and non-specific effects of potassium cations on substrate–protein interactions in cytochromes P450cam and P450lin. J. Inorg. Biochem. 2002, 91, 597–606. [Google Scholar] [CrossRef]
- Davydov, D.R.; Botchkareva, A.E.; Kumar, S.; He, Y.Q.; Halpert, J.R. An electrostatically driven conformational transition is involved in the mechanisms of substrate binding and cooperativity in cytochrome P450eryF. Biochemistry 2004, 43, 6475–6485. [Google Scholar] [CrossRef]
- Yun, C.H.; Song, M.; Ahn, T.; Kim, H. Conformational change of cytochrome P450 1A2 induced by sodium chloride. J. Biol. Chem. 1996, 271, 31312–31316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, C.H.; Ahn, T.; Guengerich, F.P. Conformational change and activation of cytochrome P450 2B1 induced by salt and phospholipid. Arch. Biochem. Biophys. 1998, 356, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Mansuy, D.; Carlier, M.; Bertrand, J.C.; Azoulay, E. Spectral Characterization of Cytochrome P-450 of a Strain of Candida tropicalis Grown on Tetradecane. Eur. J. Biochem. 1980, 109, 103–108. [Google Scholar] [CrossRef]
- Di Primo, C.; Deprez, E.; Sligar, S.G.; Hui Bon Hoa, G. Origin of the Photoacoustic Signal in Cytochrome P-450cam: Role of the Arg186- Asp251- Lys178 Bifurcated Salt Bridge. Biochemistry 1997, 36, 112–118. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KD (μM) | (kcal · mol−1) | % HS (substrate-free) | % HS @ [6DEB] = 90 µM | ||
|---|---|---|---|---|---|
| No salt | 3.83 ± 0.06 | −7.39 ± 0.01 | 0.05/0.04 | 32 | 46 |
| LiCl | 2.89 ± 0.02 | −7.55 ± 0.01 | 0.09/0.07 | 34 | 56 |
| NaCl | 5.43 ± 0.08 | −7.18 ± 0.01 | 0.07/0.06 | 33 | 55 |
| KCl | 3.71 ± 0.03 | −7.40 ± 0.01 | 0.07/0.06 | 32 | 55 |
| NH4Cl | 7.52 ± 0.05 | −6.99 ± 0.01 | 0.08/0.06 | 33 | 53 |
| MgCl2 | 3.18 ± 0.07 | −7.50 ± 0.01 | 0.11/0.10 | 33 | 67 |
| CaCl2 | 1.54 ± 0.02 | −7.92 ± 0.01 | 0.11/0.09 | 38 | 67 |
| SrCl2 | 2.82 ± 0.01 | −7.57 ± 0.01 | 0.12/0.09 | 34 | 65 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Sciscio, M.L.; Nardi, A.N.; Parisi, G.; Bulfaro, G.; Costanzo, A.; Gugole, E.; Exertier, C.; Freda, I.; Savino, C.; Vallone, B.; et al. Effect of Salts on the Conformational Dynamics of the Cytochrome P450 OleP. Molecules 2023, 28, 832. https://doi.org/10.3390/molecules28020832
De Sciscio ML, Nardi AN, Parisi G, Bulfaro G, Costanzo A, Gugole E, Exertier C, Freda I, Savino C, Vallone B, et al. Effect of Salts on the Conformational Dynamics of the Cytochrome P450 OleP. Molecules. 2023; 28(2):832. https://doi.org/10.3390/molecules28020832
Chicago/Turabian StyleDe Sciscio, Maria Laura, Alessandro Nicola Nardi, Giacomo Parisi, Giovanni Bulfaro, Antonella Costanzo, Elena Gugole, Cécile Exertier, Ida Freda, Carmelinda Savino, Beatrice Vallone, and et al. 2023. "Effect of Salts on the Conformational Dynamics of the Cytochrome P450 OleP" Molecules 28, no. 2: 832. https://doi.org/10.3390/molecules28020832
APA StyleDe Sciscio, M. L., Nardi, A. N., Parisi, G., Bulfaro, G., Costanzo, A., Gugole, E., Exertier, C., Freda, I., Savino, C., Vallone, B., Montemiglio, L. C., & D’Abramo, M. (2023). Effect of Salts on the Conformational Dynamics of the Cytochrome P450 OleP. Molecules, 28(2), 832. https://doi.org/10.3390/molecules28020832