

Comparative Metabolomics and Transcriptome Analysis Reveal the Fragrance-Related Metabolite Formation in Phoebe zhennan Wood
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
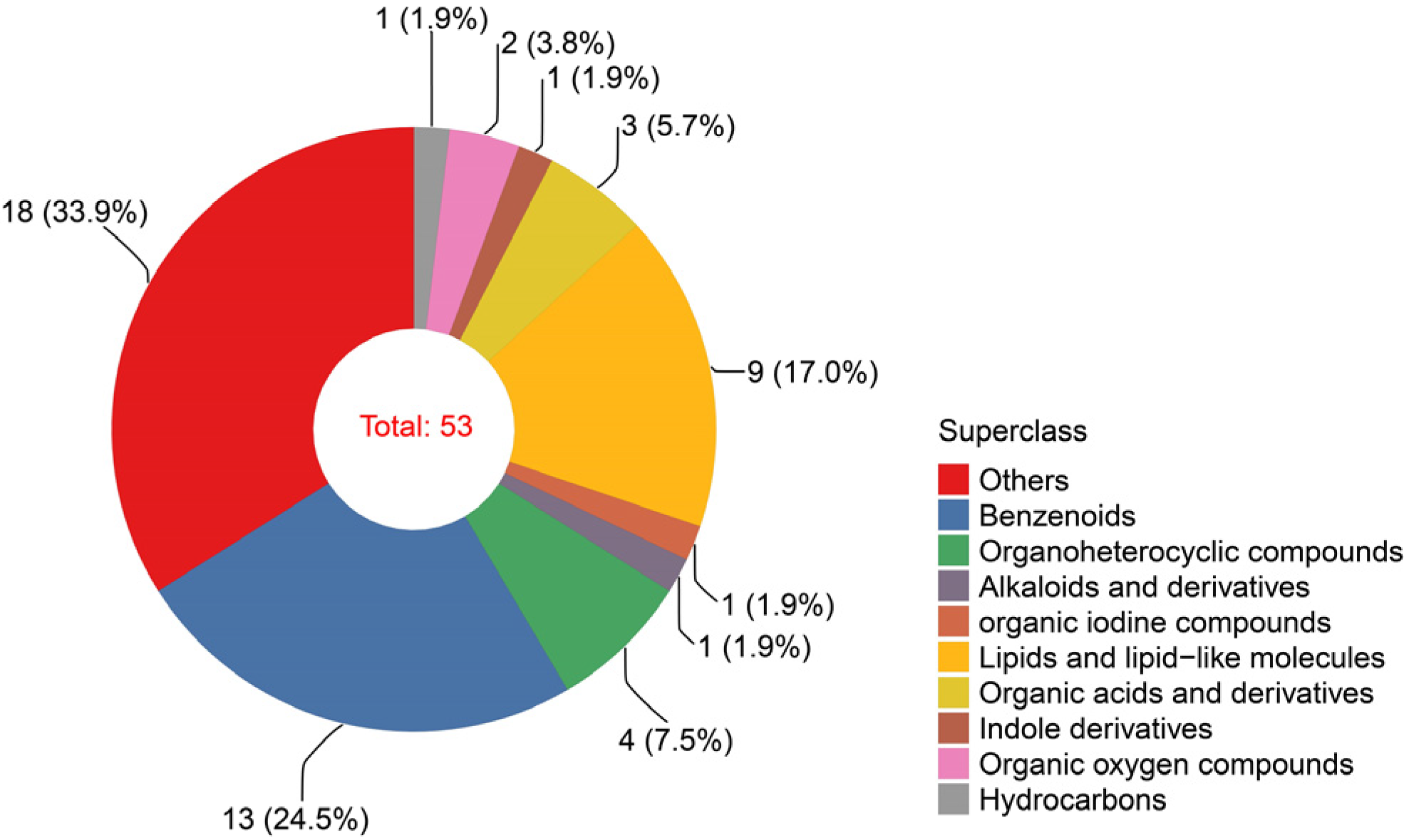
2.1. Metabolic Profiles
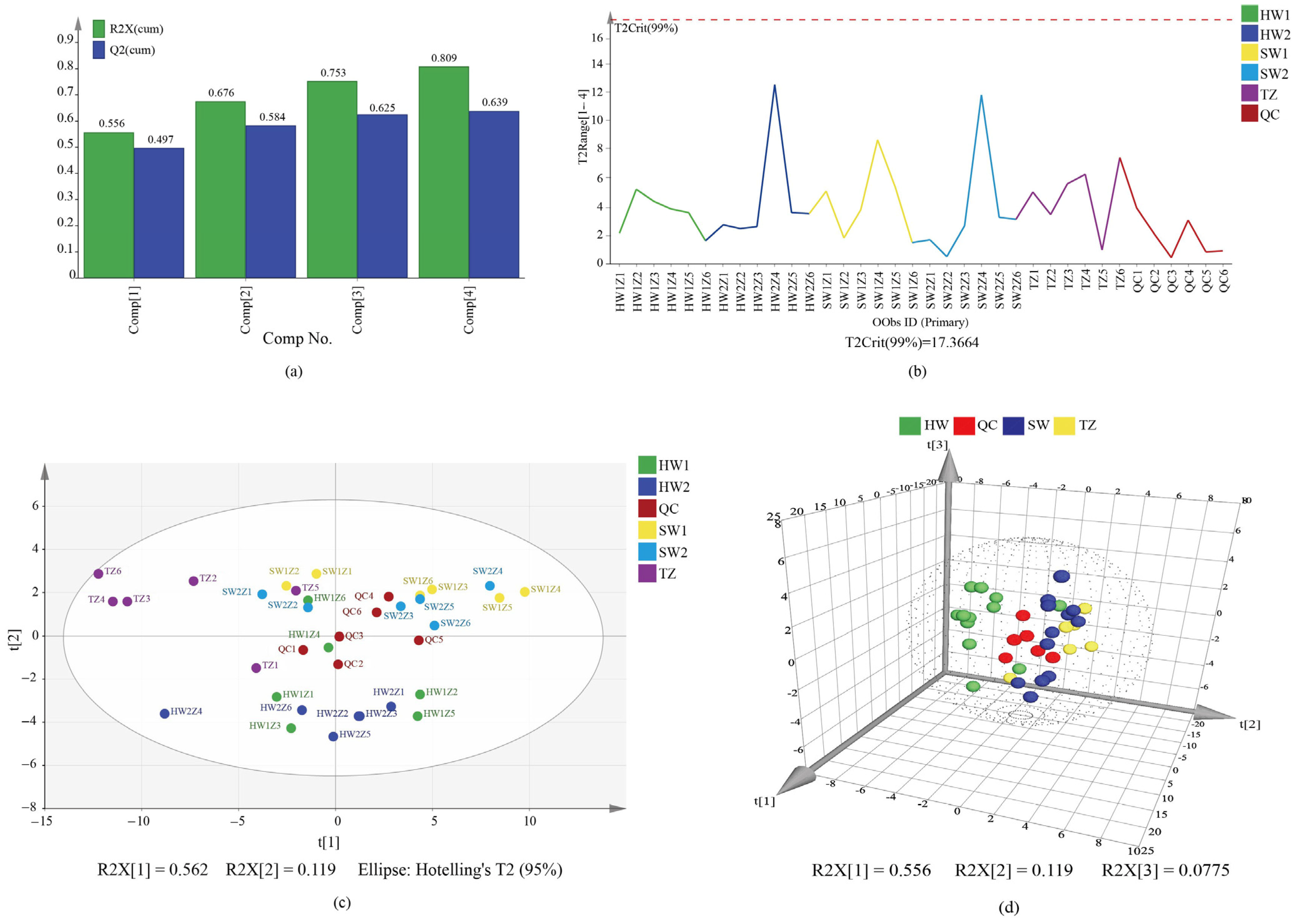
2.2. Principal Component Analysis (PCA) and Hierarchical Cluster Analysis (HCA) of All Tissues
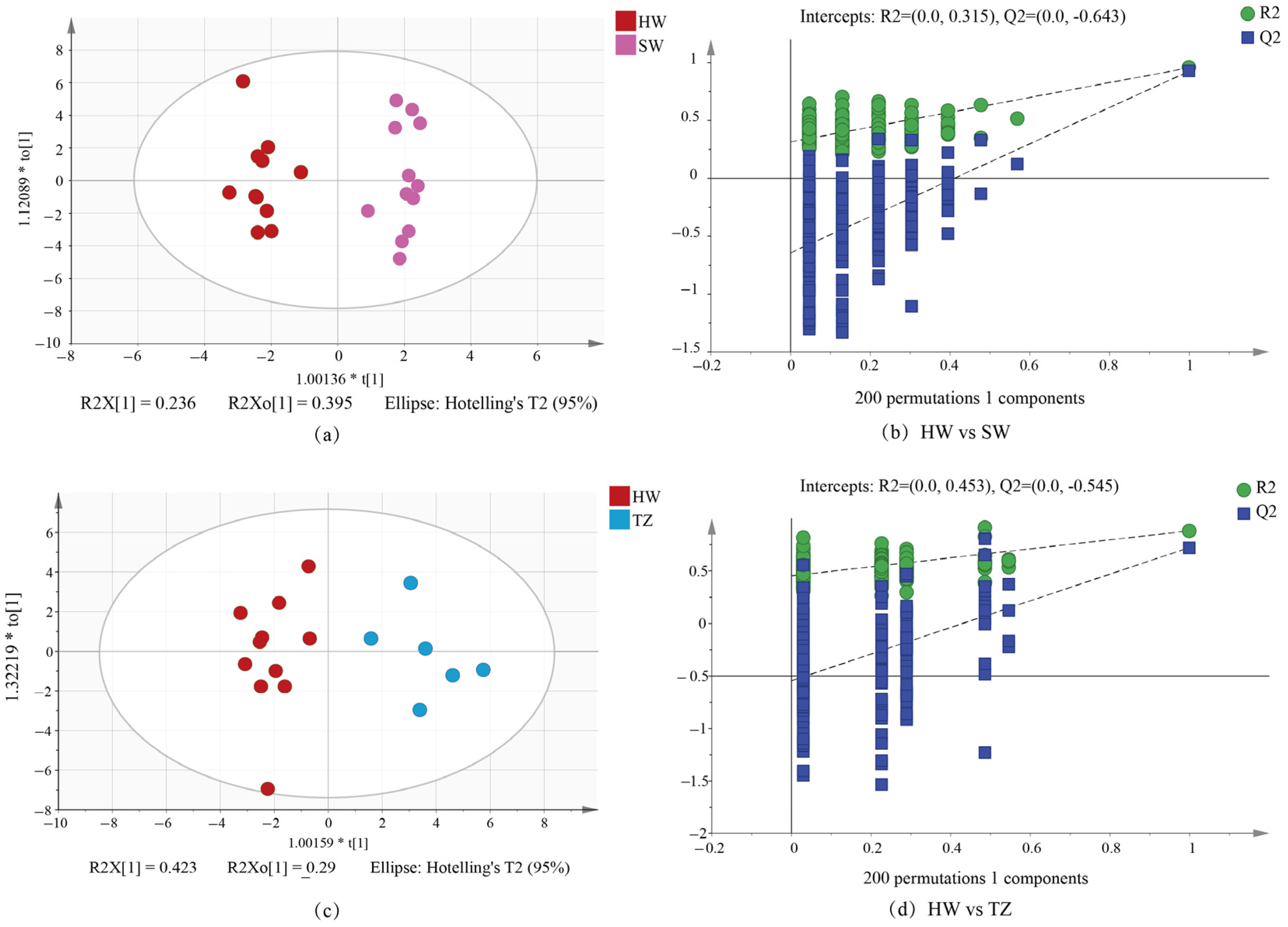
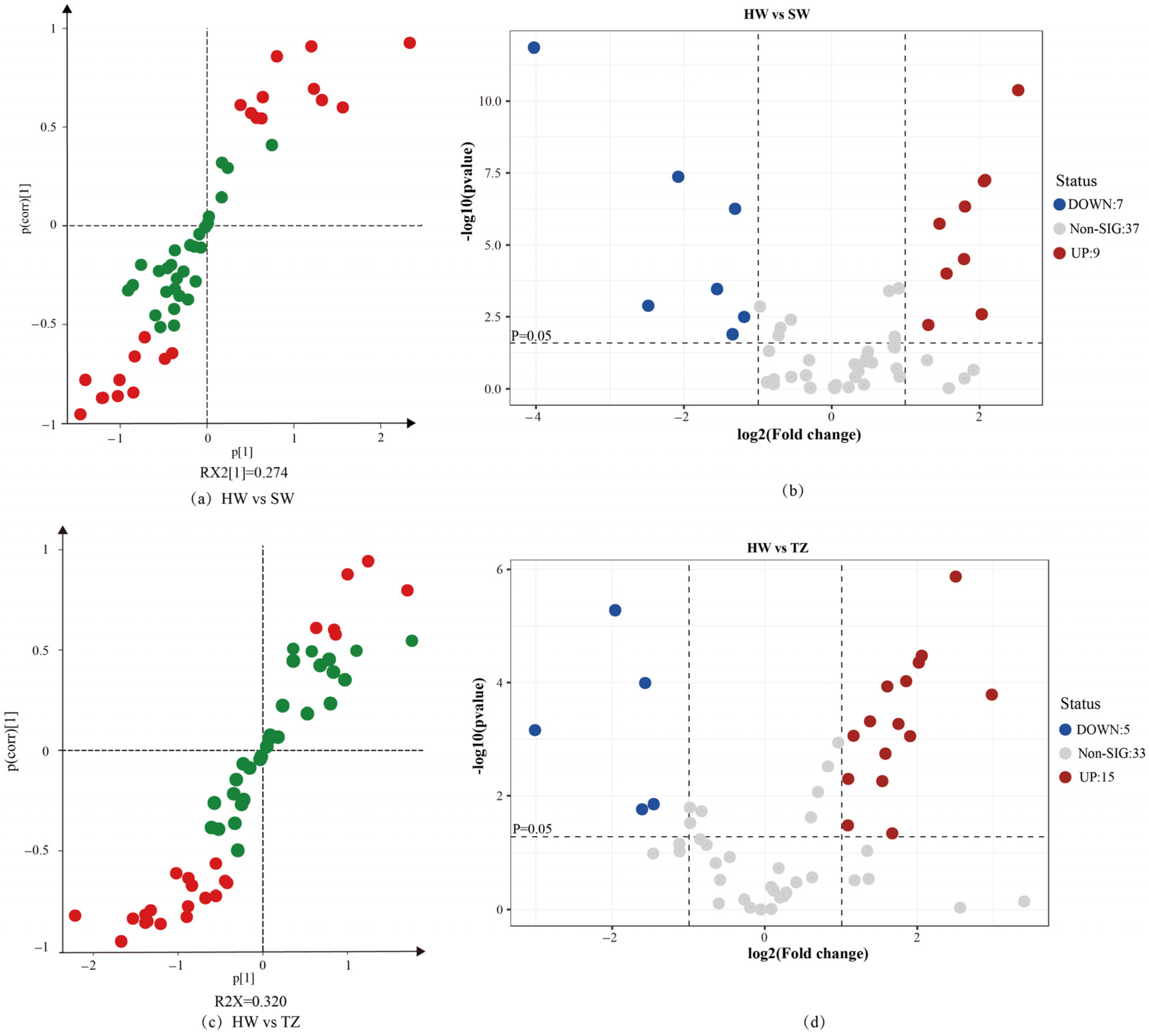
2.3. Orthogonal Partial Least Squares–Discrimination Analysis (OPLS-DA)
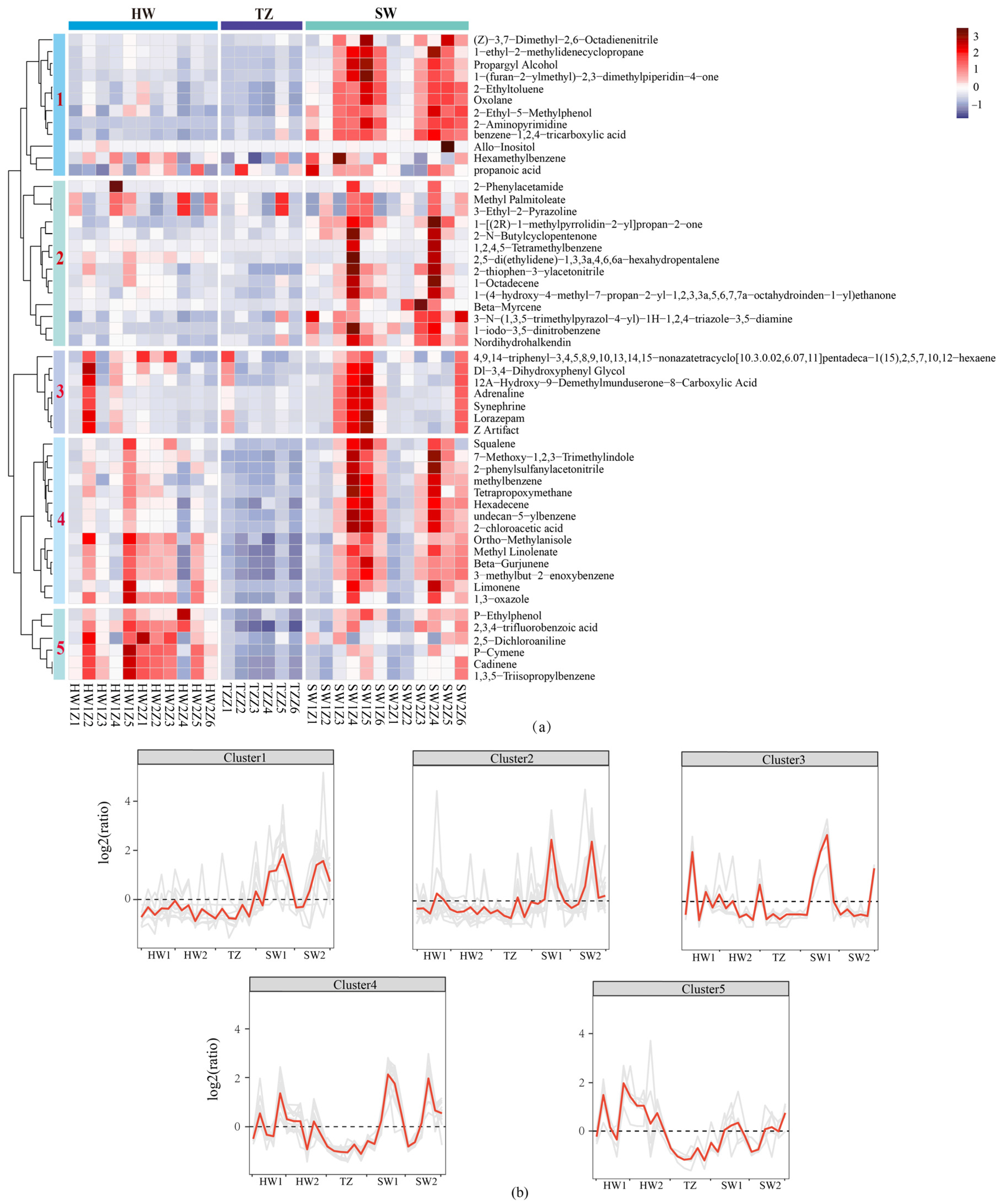
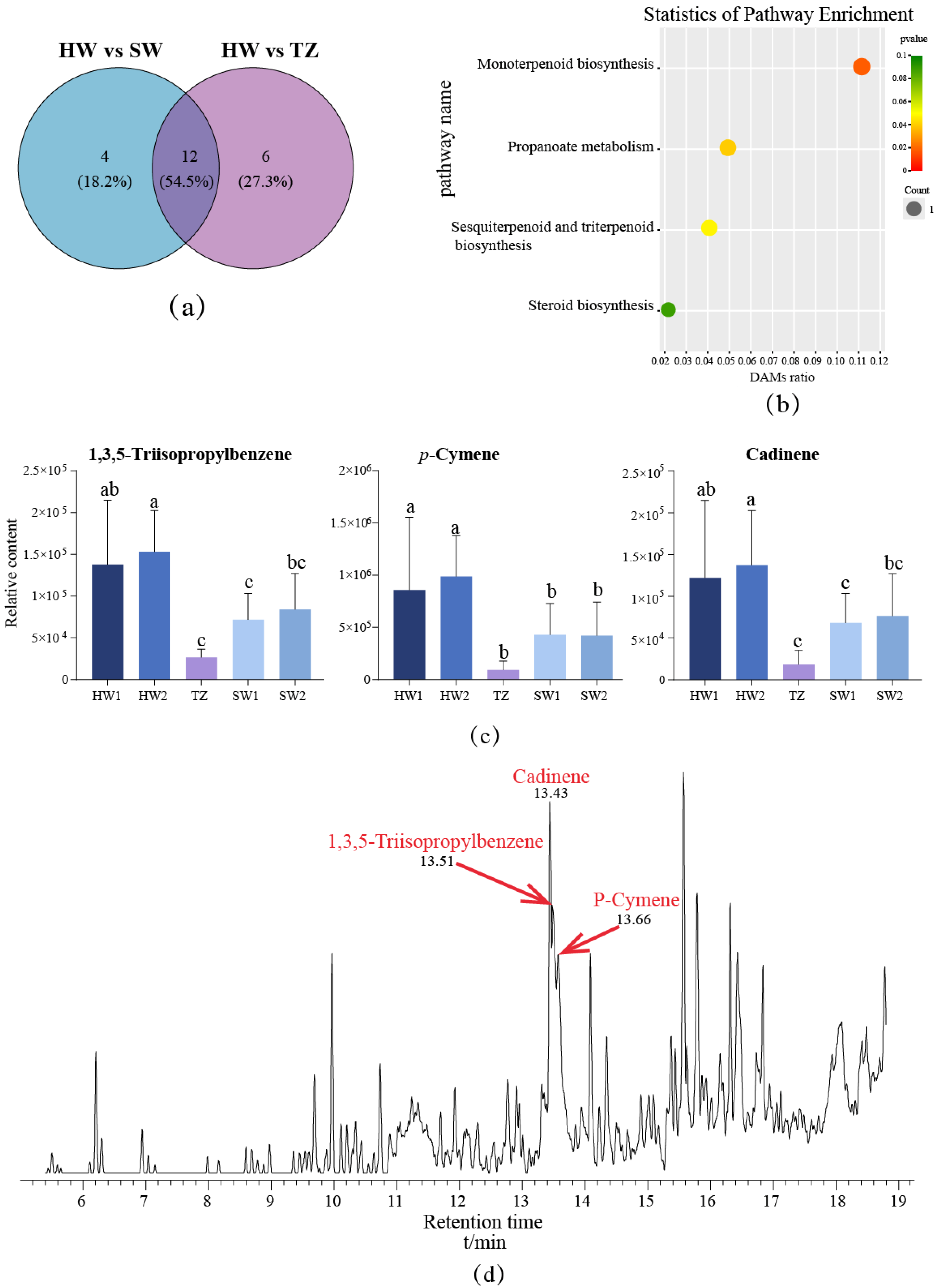
2.4. Identification of Differentially Accumulated Metabolites in Pairwise Comparisons
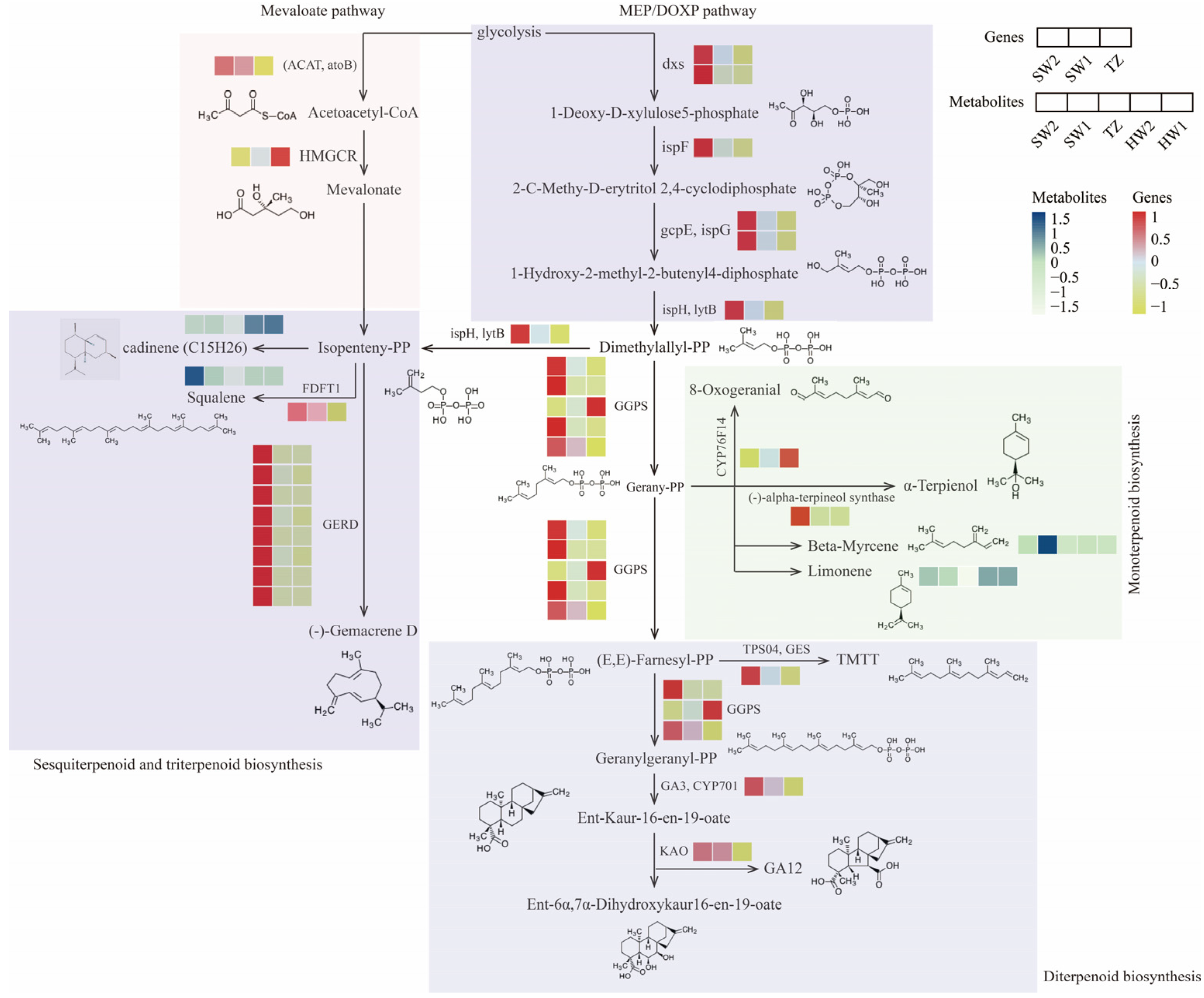
2.5. Analysis of the Key Pathway Involved in Terpenoid Biosynthesis in P. zhennan Wood
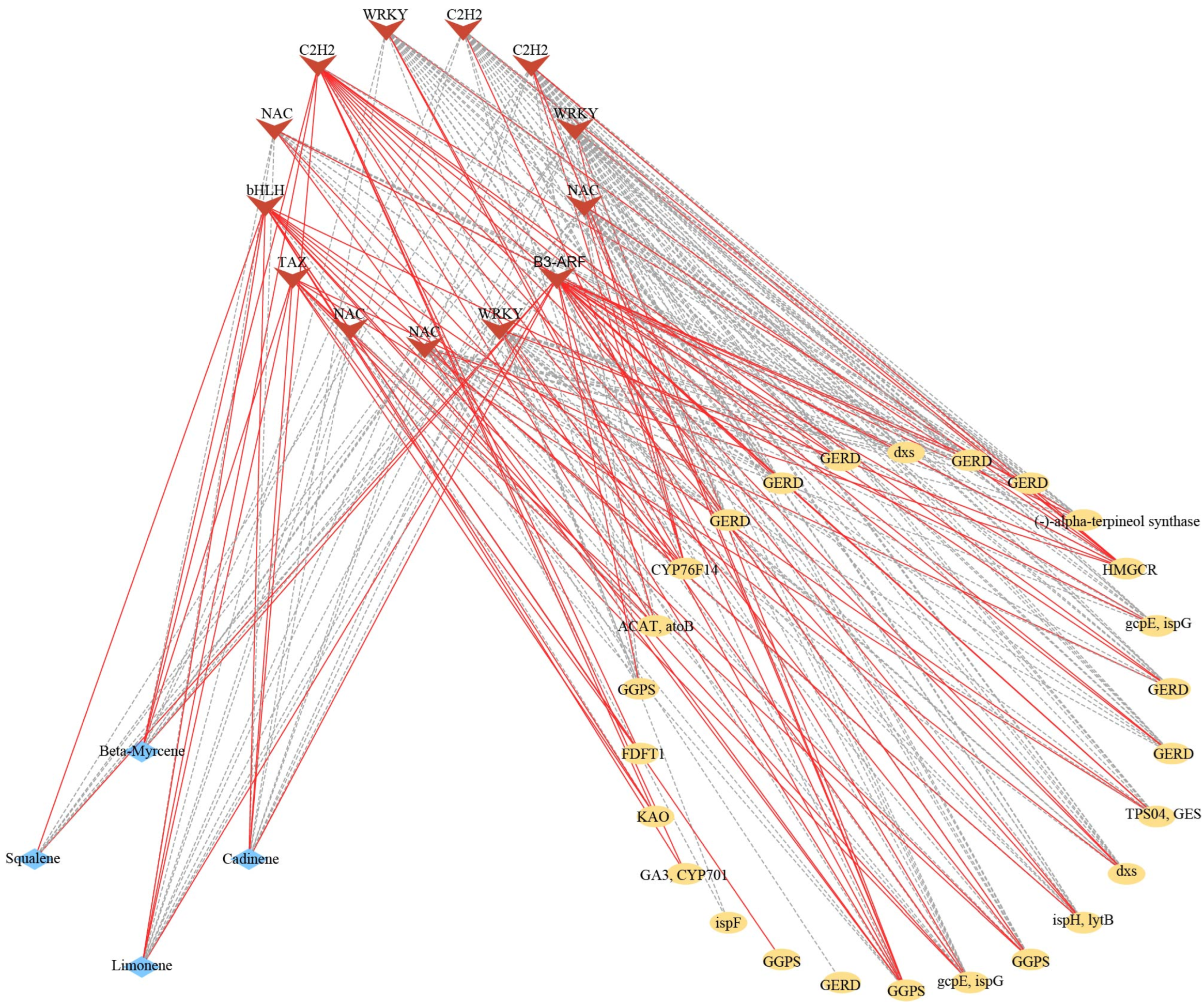
2.6. Transcription Factors Related to Terpenoid Biosynthesis
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. Sample Preparation for Metabolite Profiling
4.3. GC-MS Analysis Conditions
4.4. Metabolite Identification
4.5. Statistical Analysis of Fragrance-Related Metabolites
4.6. Terpenoid Biosynthesis Pathway Analysis According to RNA-seq Data
4.7. Transcription Factor Related to Terpenoid Biosynthesis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Zhu, Y.; An, W.; Peng, J.; Li, J.; Gu, Y.; Jiang, B.; Chen, L.; Zhu, P.; Yang, H. Genetic diversity of Nanmu (Phoebe zhennan S. Lee. et F. N. Wei) breeding population and extraction of core collection using nSSR, cpSSR and phenotypic markers. Forests 2022, 13, 1320. [Google Scholar] [CrossRef]
- Yang, H.; An, W.; Gu, Y.; Peng, J.; Jiang, Y.; Li, J.; Chen, L.; Zhu, P.; He, F.; Zhang, F.; et al. Integrative Metabolomic and Transcriptomic Analysis Reveals the Mechanism of Specific Color Formation in Phoebe zhennan Heartwood. Int. J. Mol. Sci. 2022, 23, 13569. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.-H.; Ding, X.; Li, L.; Ma, H.; Ci, X.-Q.; van der Merwe, M.; Conran, J.G.; Li, J. Miocene diversification of a golden-thread nanmu tree species (Phoebe zhennan, Lauraceae) around the Sichuan Basin shaped by the East Asian monsoon. Ecol. Evol. 2020, 10, 10543–10557. [Google Scholar] [CrossRef]
- Tariq, A.; Pan, K.; Olatunji, O.A.; Graciano, C.; Li, Z.; Sun, F.; Sun, X.; Song, D.; Chen, W.; Zhang, A.; et al. Phosphorous application improves drought tolerance of Phoebe zhennan. Front. Plant Sci. 2017, 8, 1561. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Zhang, W.; Li, J.; Long, H.; He, W.; Li, X. Amplified fragment length polymorphism analysis of the population structure and genetic diversity of Phoebe zhennan (Lauraceae), a native species to China. Biochem. Syst. Ecol. 2016, 64, 149–155. [Google Scholar] [CrossRef]
- Jiao, L.; Lu, Y.; Zhang, M.; Chen, Y.; Wang, Z.; Guo, Y.; Xu, C.; Guo, J.; He, T.; Ma, L.; et al. Ancient plastid genomes solve the tree species mystery of the imperial wood “Nanmu” in the Forbidden City, the largest existing wooden palace complex in the world. Plants People Planet 2022, 4, 696–709. [Google Scholar] [CrossRef]
- Xie, J.; Qi, J.; Huang, X.; Zhou, N.; Hu, Y. Comparative analysis of modern and ancient buried Phoebe zhennan wood: Surface color, chemical components, infrared spectroscopy, and essential oil composition. J. For. Res. 2015, 26, 501–507. [Google Scholar] [CrossRef]
- Shao, H.; Jiang, Y.; Pan, F.; Xie, J.; Qi, J.; Xiao, H.; Chen, Y. Chemical composition, UV/vis absorptivity, and antioxidant activity of essential oils from bark and leaf of Phoebe zhennan SK Lee & FN Wei. Nat. Prod. Res. 2020, 34, 876–879. [Google Scholar]
- Wang, X.; Wang, C.; Zhang, Q.; Quan, X. Heartwood and sapwood allometry of seven Chinese temperate tree species. Ann. For. Sci. 2010, 67, 410. [Google Scholar] [CrossRef]
- Hoch, G.; Richter, A.; KÖRner, C. Non-structural carbon compounds in temperate forest trees. Plant Cell Environ. 2003, 26, 1067–1081. [Google Scholar] [CrossRef]
- Yang, H.; An, W.; Wang, F.; Gu, Y.; Guo, H.; Jiang, Y.; Peng, J.; Liu, M.; Chen, L.; Zhang, F.; et al. Integrated Transcriptomic, Metabolomic, and Physiological Analyses Reveal New Insights into Fragrance Formation in the Heartwood of Phoebe hui. Int. J. Mol. Sci. 2022, 23, 14044. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.; Wang, J.; Yang, F.; Lei, C. Comparative analysis of essential oil components of two Cryptomeria species from China. Ind. Crops Prod. 2011, 34, 1226–1230. [Google Scholar] [CrossRef]
- Celedon, J.M.; Bohlmann, J. An extended model of heartwood secondary metabolism informed by functional genomics. Tree Physiol. 2018, 38, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.; Liping, N.; Xing, H.; Wei, Z.; Zhoua, Q.; Nong, R.; Chen, J. Essential oil extracted from leaf of Phoebe bournei (Hemsl.) yang: Chemical constituents, antitumor, antibacterial, hypoglycemic activities. Nat. Prod. Res. 2020, 34, 2524–2527. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Zhang, J.; Han, S.; Chong, S.L.; Meng, G.; Song, M.; Wang, Y.; Zhou, S.; Liu, C.; Lou, L.; et al. The chromosome-scale genome of Phoebe bournei reveals contrasting fates of terpene synthase (TPS)-a and TPS-b subfamilies. Plant Commun. 2022, 3, 100410. [Google Scholar] [CrossRef]
- Qi, J.; Wei, J.; Liao, D.; Ding, Z.; Yao, X.; Sun, P.; Li, X. Untargeted Metabolomics Analysis Revealed the Major Metabolites in the Seeds of four Polygonatum Species. Molecules 2022, 27, 1445. [Google Scholar] [CrossRef]
- Mudiam, M.K.R.; Ch, R.; Saxena, P.N. Gas Chromatography-Mass Spectrometry Based Metabolomic Approach for Optimization and Toxicity Evaluation of Earthworm Sub-Lethal Responses to Carbofuran. PLoS ONE 2013, 8, e81077. [Google Scholar] [CrossRef]
- Castro-Alves, V.; Kalbina, I.; Nilsen, A.; Aronsson, M.; Rosenqvist, E.; Jansen, M.A.K.; Qian, M.; Öström, Å.; Hyötyläinen, T.; Strid, Å. Integration of non-target metabolomics and sensory analysis unravels vegetable plant metabolite signatures associated with sensory quality: A case study using dill (Anethum graveolens). Food Chem. 2021, 344, 128714. [Google Scholar] [CrossRef]
- Peng, D.-Q.; Yu, Z.-X.; Wang, C.-H.; Gong, B.; Liu, Y.-Y.; Wei, J.-H. Chemical Constituents and Anti-Inflammatory Effect of Incense Smoke from Agarwood Determined by GC-MS. Int. J. Anal. Chem. 2020, 2020, 4575030. [Google Scholar] [CrossRef]
- Liu, Y.; Zhu, X.; Qin, X.; Wang, W.; Hu, Y.; Yuan, D. Identification and characterization of odorous volatile organic compounds emitted from wood-based panels. Environ. Monit. Assess. 2020, 192, 348. [Google Scholar] [CrossRef]
- Huang, Y.; Xie, F.-J.; Cao, X.; Li, M.-Y. Research progress in biosynthesis and regulation of plant terpenoids. Biotechnol. Biotechnol. Equip. 2021, 35, 1799–1808. [Google Scholar] [CrossRef]
- Chen, F.; Tholl, D.; Bohlmann, J.; Pichersky, E. The family of terpene synthases in plants: A mid-size family of genes for specialized metabolism that is highly diversified throughout the kingdom. Plant J. 2011, 66, 212–229. [Google Scholar] [CrossRef] [PubMed]
- Celedon, J.M.; Chiang, A.; Yuen, M.M.S.; Diaz-Chavez, M.L.; Madilao, L.L.; Finnegan, P.M.; Barbour, E.L.; Bohlmann, J. Heartwood-specific transcriptome and metabolite signatures of tropical sandalwood (Santalum album) reveal the final step of (Z)-santalol fragrance biosynthesis. Plant J. 2016, 86, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Yeh, T.-F.; Chu, J.-H.; Liu, L.-Y.; Chen, S.-Y. Differential Gene Profiling of the Heartwood Formation Process in Taiwania cryptomerioides Hayata Xylem Tissues. Int. J. Mol. Sci. 2020, 21, 960. [Google Scholar] [CrossRef]
- Dong, Y.; Zhang, W.; Ling, Z.; Li, J.; Bai, H.; Li, H.; Shi, L. Advances in Transcription Factors Regulating Plant Terpenoids Biosynthesis. Chin. Bull. Bot. 2020, 55, 340–350. [Google Scholar]
- Choi, C.W.; Choi, Y.H.; Cha, M.R.; Kim, Y.S.; Yon, G.H.; Kim, Y.K.; Choi, S.U.; Kim, Y.H.; Ryu, S.Y. Antitumor Components Isolated from the Heartwood Extract of Dalbergia odorifera. J. Korean Soc. Appl. Biol. Chem. 2009, 52, 375–379. [Google Scholar] [CrossRef]
- Ghadiriasli, R.; Mahmoud, M.A.A.; Wagenstaller, M.; Van de Kuilen, J.W.; Buettner, A. Molecular and sensory characterization of odorants in Cembran pine (Pinus cembra L.) from different geographic regions. Talanta 2020, 220, 121380. [Google Scholar] [CrossRef]
- Kamatou, G.P.P.; Viljoen, A.M.; Özek, T.; Başer, K.H.C. Chemical composition of the wood and leaf oils from the “Clanwilliam Cedar” (Widdringtonia cedarbergensis J.A. Marsh): A critically endangered species. S. Afr. J. Bot. 2010, 76, 652–654. [Google Scholar] [CrossRef]
- Xing, L.; Sun, L.; Liu, S.; Zhang, L.; Sun, J.; Yang, H. Metabolomic analysis of white, green and purple morphs of sea cucumber Apostichopus japonicus during body color pigmentation process. Comp. Biochem. Physiol. Part D Genom. Proteom. 2021, 39, 100827. [Google Scholar] [CrossRef]
- Zeng, X.; Li, J.X.; Lyu, X.; Chen, J.; Chen, X.M.; Guo, S.X. Untargeted Metabolomics Reveals Multiple Phytometabolites in the Agricultural Waste Materials and Medicinal Materials of Codonopsis pilosula. Front. Plant Sci. 2022, 12, 814011. [Google Scholar] [CrossRef]
- Hanson, J.R. Natural Products: The Secondary Metabolites; The Royal Society of Chemistry: London, UK, 2003. [Google Scholar]
- Cherri-Martin, M.; Jullien, F.; Heizmann, P.; Baudino, S. Fragrance heritability in hybrid tea roses. Sci. Hortic. 2007, 113, 177–181. [Google Scholar] [CrossRef]
- Pichersky, E.; Raguso, R.A. Why do plants produce so many terpenoid compounds? New Phytol. 2018, 220, 692–702. [Google Scholar] [CrossRef] [PubMed]
- Surendran, S.; Qassadi, F.; Surendran, G.; Lilley, D.; Heinrich, M. Myrcene-What Are the Potential Health Benefits of This Flavouring and Aroma Agent? Front. Nutr. 2021, 8, 699666. [Google Scholar] [CrossRef] [PubMed]
- Sterba, K.; Cejka, P.; Culik, J.; Jurkova, M.; Krofta, K.; Pavlovic, M.; Mikyska, A.; Olsovska, J. Determination of Linalool in Different Hop Varieties Using a New Method Based on Fluidized-Bed Extraction with Gas Chromatographic-Mass Spectrometric Detection. J. Am. Soc. Brew. Chem. 2015, 73, 151–158. [Google Scholar]
- Huo, H.X.; Zhu, Z.X.; Pang, D.R.; Li, Y.T.; Huang, Z.; Shi, S.P.; Zheng, J.; Zhang, Q.; Zhao, Y.F.; Tu, P.F.; et al. Anti-neuroinflammatory sesquiterpenes from Chinese eaglewood. Fitoterapia 2015, 106, 115–121. [Google Scholar] [CrossRef]
- Balaban-Ucar, M.; Gonultas, O. Volatile compounds of archaeological wood from the ancient harbor Thedosius in Istanbul. Eur. J. Wood Wood Prod. 2019, 77, 475–481. [Google Scholar] [CrossRef]
- Joshi, S.C.; Padalia, R.C.; Bisht, D.S.; Mathela, C.S. Terpenoid Diversity in the Leaf Essential Oils of Himalayan lauraceae Species. Chem. Biodivers. 2009, 6, 1364–1373. [Google Scholar] [CrossRef]
- Tonisi, S.; Okaiyeto, K.; Hoppe, H.; Mabinya, L.V.; Nwodo, U.U.; Okoh, A.I. Chemical constituents, antioxidant and cytotoxicity properties of Leonotis leonurus used in the folklore management of neurological disorders in the Eastern Cape, South Africa. 3 Biotech 2020, 10, 141. [Google Scholar] [CrossRef]
- Kumar, L.R.G.; Chatterjee, N.S.; Tejpal, C.S.; Vishnu, K.V.; Anas, K.K.; Asha, K.K.; Anandan, R.; Mathew, S. Evaluation of chitosan as a wall material for microencapsulation of squalene by spray drying: Characterization and oxidative stability studies. Int. J. Biol. Macromol. 2017, 104, 1986–1995. [Google Scholar] [CrossRef]
- Singh, H.P.; Kaur, S.; Mittal, S.; Batish, D.R.; Kohli, R.K. In vitro screening of essential oil from young and mature leaves of Artemisia scoparia compared to its major constituents for free radical scavenging activity. Food Chem. Toxicol. 2010, 48, 1040–1044. [Google Scholar] [CrossRef]
- Li, Y.Y.; Wan, Y.M.; Sun, Z.H.; Li, T.Q.; Liu, X.F.; Ma, H.; Liu, X.X.; He, R.; Ma, Y.; Li, Z.H. Floral Scent Chemistry of Luculia yunnanensis (Rubiaceae), a Species Endemic to China with Sweetly Fragrant Flowers. Molecules 2017, 22, 879. [Google Scholar] [CrossRef] [PubMed]
- Miyazawa, M.; Nakashima, Y.; Nakahashi, H.; Hara, N.; Nakagawa, H.; Usami, A.; Chavasiri, W. Volatile Compounds with Characteristic Odor of Essential Oil from Magnolia obovata Leaves by Hydrodistillation and Solvent-assisted Flavor Evaporation. J. Oleo Sci. 2015, 64, 999–1007. [Google Scholar] [CrossRef]
- Plagemann, I.; Krings, U.; Berger, R.G.; Marostica, M.R. Volatile constituents of jabuticaba (Myrciaria jaboticaba (Vell.) O. Berg) fruits. J. Essent. Oil Res. 2012, 24, 45–51. [Google Scholar] [CrossRef]
- Agulló, L.; Romero-Silva, M.J.; Domenech, M.; Seeger, M. p-Cymene Promotes Its Catabolism through the p-Cymene and the p-Cumate Pathways, Activates a Stress Response and Reduces the Biofilm Formation in Burkholderia xenovorans LB400. PLoS ONE 2017, 12, e0169544. [Google Scholar] [CrossRef] [PubMed]
- Chung, M.; Cheng, S.; Lin, C.; Chang, S. Profiling of aroma compounds released from cooking Dendrocalamus latiflorus shoots. BioResources 2020, 15, 8744–8755. [Google Scholar] [CrossRef]
- Kabir, Y.; Akasaka-Hashimoto, Y.; Kubota, K.; Komai, M. Volatile compounds of black cumin (Nigella sativa L.) seeds cultivated in Bangladesh and India. Heliyon 2020, 6, e05343. [Google Scholar] [CrossRef]
- Loreto, F.; Schnitzler, J.-P. Abiotic stresses and induced BVOCs. Trends Plant Sci. 2010, 15, 154–166. [Google Scholar] [CrossRef]
- Arimura, G.-I.; Huber, D.P.W.; Bohlmann, J. Forest tent caterpillars (Malacosoma disstria) induce local and systemic diurnal emissions of terpenoid volatiles in hybrid poplar (Populus trichocarpa × deltoides): cDNA cloning, functional characterization, and patterns of gene expression of (−)-germacrene D synthase, PtdTPS1. Plant J. Cell Mol. Biol. 2004, 37, 603–616. [Google Scholar]
- Broun, P. Transcription factors as tools for metabolic engineering in plants. Curr. Opin. Plant Biol. 2004, 7, 202–209. [Google Scholar] [CrossRef]
- Ma, D.; Pu, G.; Lei, C.; Ma, L.; Wang, H.; Guo, Y.; Chen, J.; Du, Z.; Wang, H.; Li, G.; et al. Isolation and Characterization of AaWRKY1, an Artemisia annua Transcription Factor that Regulates the Amorpha-4,11-diene Synthase Gene, a Key Gene of Artemisinin Biosynthesis. Plant Cell Physiol. 2009, 50, 2146–2161. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wang, H.; Yang, J.; Deng, K.; Wang, T. RNA sequencing on Amomum villosum Lour. induced by MeJA identifies the genes of WRKY and terpene synthases involved in terpene biosynthesis. Genome 2018, 61, 91–102. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.-H.; Wang, J.-W.; Wang, S.; Wang, J.-Y.; Chen, X.-Y. Characterization of GaWRKY1, a cotton transcription factor that regulates the sesquiterpene synthase gene (+)-delta-cadinene synthase-A. Plant Physiol. 2004, 135, 507–515. [Google Scholar] [CrossRef]
- Chuang, Y.C.; Hung, Y.C.; Tsai, W.C.; Chen, W.H.; Chen, H.H. PbbHLH4 regulates floral monoterpene biosynthesis in Phalaenopsis orchids. J. Exp. Bot. 2018, 69, 4363–4377. [Google Scholar] [CrossRef]
- Nieuwenhuizen, N.J.; Chen, X.; Wang, M.Y.; Matich, A.J.; Perez, R.L.; Allan, A.C.; Green, S.A.; Atkinson, R.G. Natural Variation in Monoterpene Synthesis in Kiwifruit: Transcriptional Regulation of Terpene Synthases by NAC and Ethylene-Insensitive3-Like Transcription Factors. Plant Physiol. 2015, 167, 1243–1258. [Google Scholar] [CrossRef] [PubMed]
- Sharma, B.; Seth, R.; Thakur, S.; Parmar, R.; Masand, M.; Devi, A.; Singh, G.; Dhyani, P.; Choudhary, S.; Sharma, R.K. Genome-wide transcriptional analysis unveils the molecular basis of organ-specific expression of isosteroidal alkaloids biosynthesis in critically endangered Fritillaria roylei Hook. Phytochemistry 2021, 187, 112772. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Yu, G.; Shi, C.; Liu, L.; Guo, Q.; Han, C.; Zhang, D.; Zhang, L.; Liu, B.; Gao, H.; et al. Majorbio Cloud: A one-stop, comprehensive bioinformatic platform for multiomics analyses. iMeta 2022, 1, e12. [Google Scholar] [CrossRef]
- Varet, H.; Brillet-Guéguen, L.; Coppée, J.-Y.; Dillies, M.-A. SARTools: A DESeq2- and EdgeR-Based R Pipeline for Comprehensive Differential Analysis of RNA-Seq Data. PLoS ONE 2016, 11, e0157022. [Google Scholar] [CrossRef] [PubMed]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, H.; Wang, F.; An, W.; Gu, Y.; Jiang, Y.; Guo, H.; Liu, M.; Peng, J.; Jiang, B.; Wan, X.; et al. Comparative Metabolomics and Transcriptome Analysis Reveal the Fragrance-Related Metabolite Formation in Phoebe zhennan Wood. Molecules 2023, 28, 7047. https://doi.org/10.3390/molecules28207047
Yang H, Wang F, An W, Gu Y, Jiang Y, Guo H, Liu M, Peng J, Jiang B, Wan X, et al. Comparative Metabolomics and Transcriptome Analysis Reveal the Fragrance-Related Metabolite Formation in Phoebe zhennan Wood. Molecules. 2023; 28(20):7047. https://doi.org/10.3390/molecules28207047
Chicago/Turabian StyleYang, Hanbo, Fang Wang, Wenna An, Yunjie Gu, Yongze Jiang, Hongying Guo, Minhao Liu, Jian Peng, Bo Jiang, Xueqin Wan, and et al. 2023. "Comparative Metabolomics and Transcriptome Analysis Reveal the Fragrance-Related Metabolite Formation in Phoebe zhennan Wood" Molecules 28, no. 20: 7047. https://doi.org/10.3390/molecules28207047
APA StyleYang, H., Wang, F., An, W., Gu, Y., Jiang, Y., Guo, H., Liu, M., Peng, J., Jiang, B., Wan, X., Chen, L., Huang, X., He, F., & Zhu, P. (2023). Comparative Metabolomics and Transcriptome Analysis Reveal the Fragrance-Related Metabolite Formation in Phoebe zhennan Wood. Molecules, 28(20), 7047. https://doi.org/10.3390/molecules28207047