
Anti-Neurodegenerating Activity: Structure–Activity Relationship Analysis of Flavonoids

 , , , ,
, , , ,
Abstract
:1. Introduction
Why Focus on Flavonoids
2. Results and Discussion
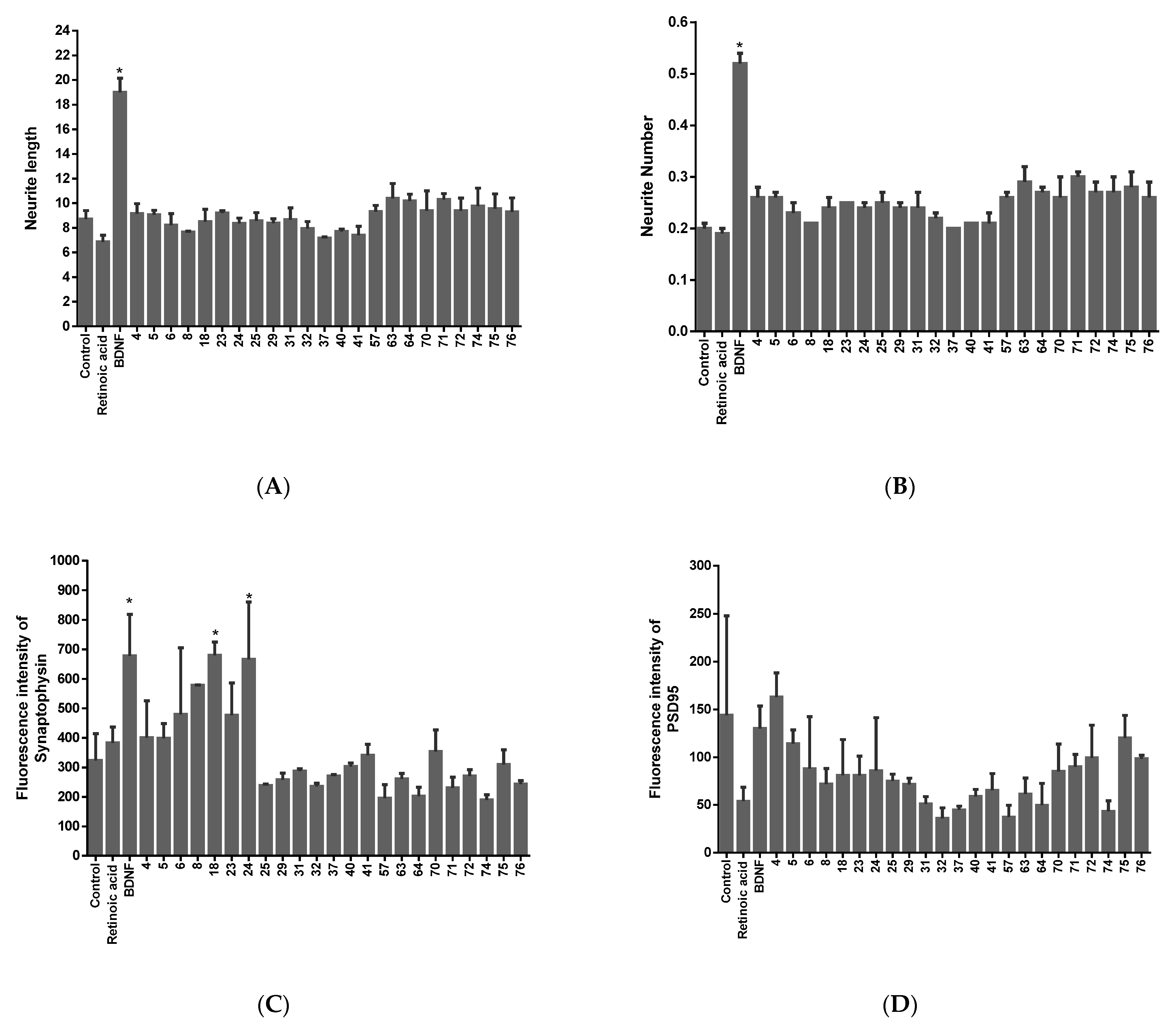
2.1. Screening of Neurotrophic Activity of the Compounds
2.2. Validation of the Neurotrophic and Synaptogenic Activities of the Candidates
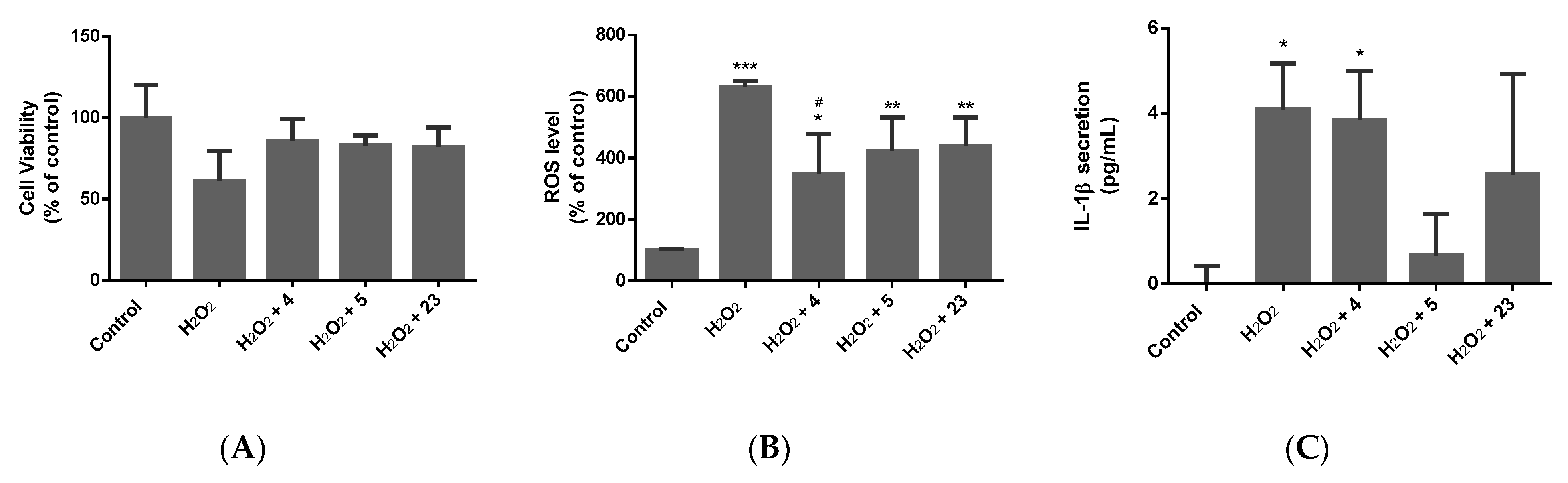
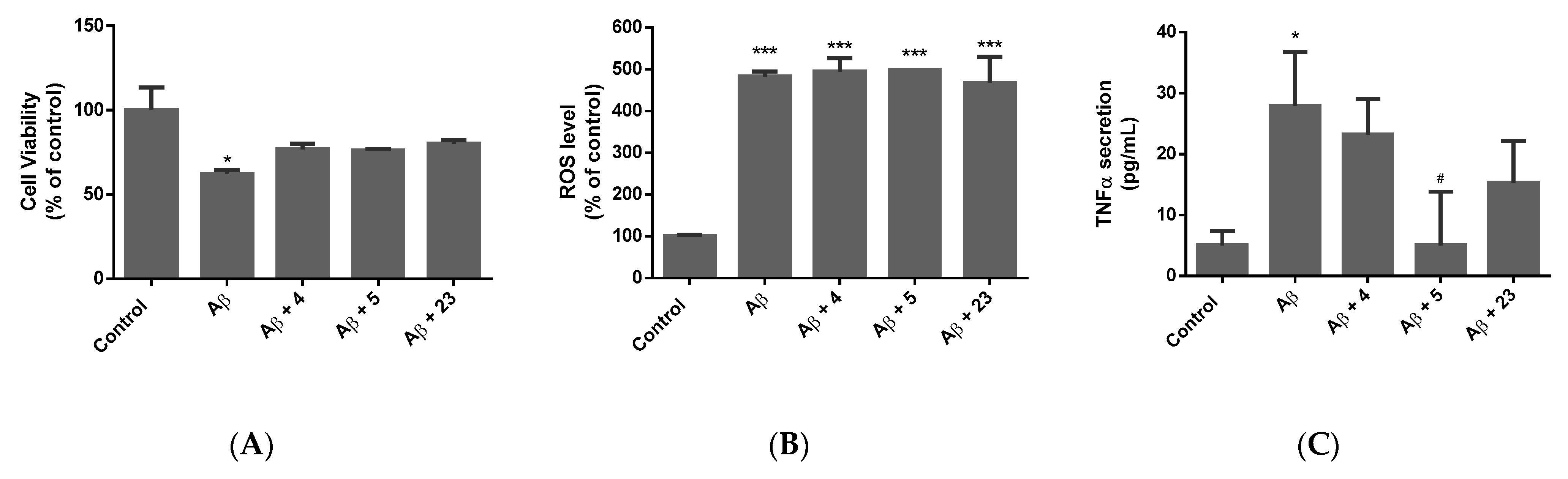
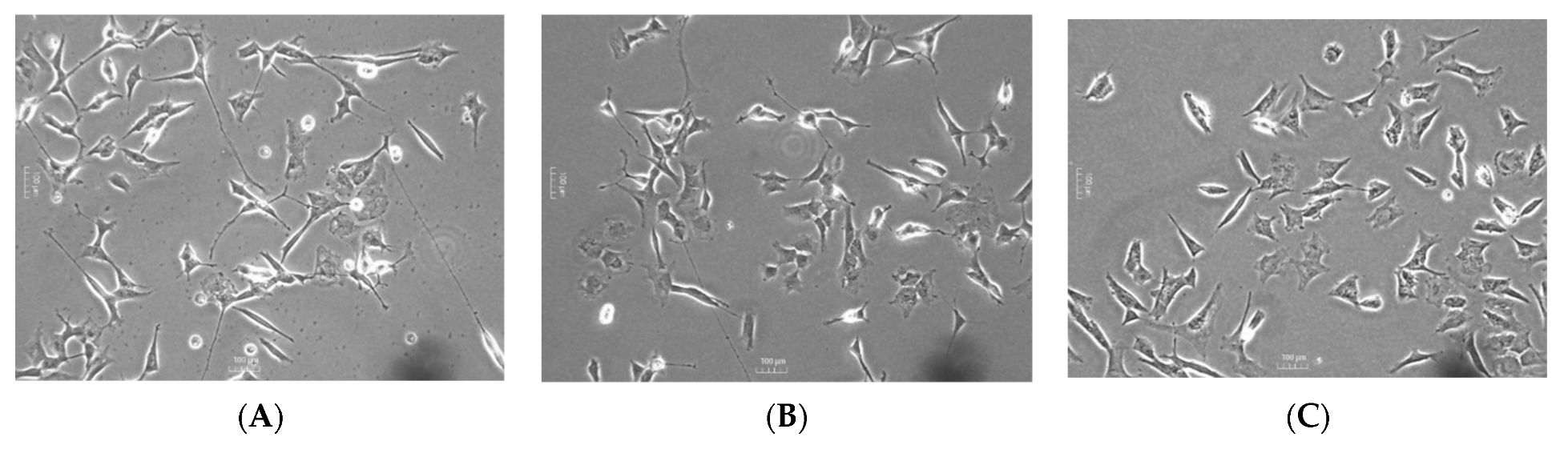
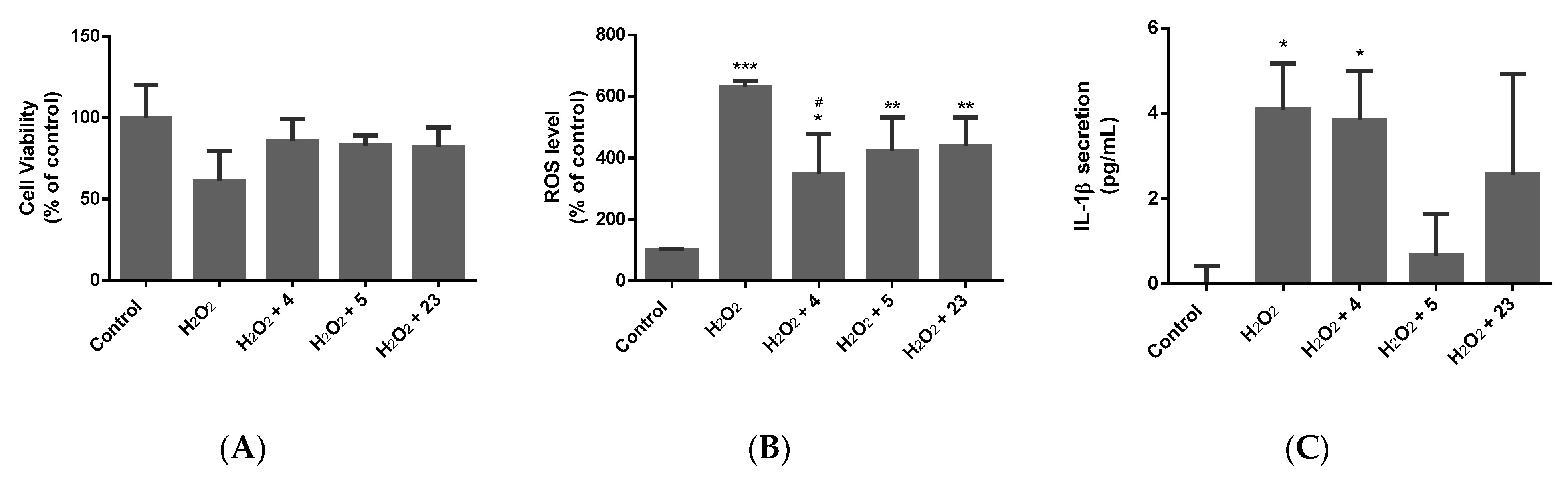
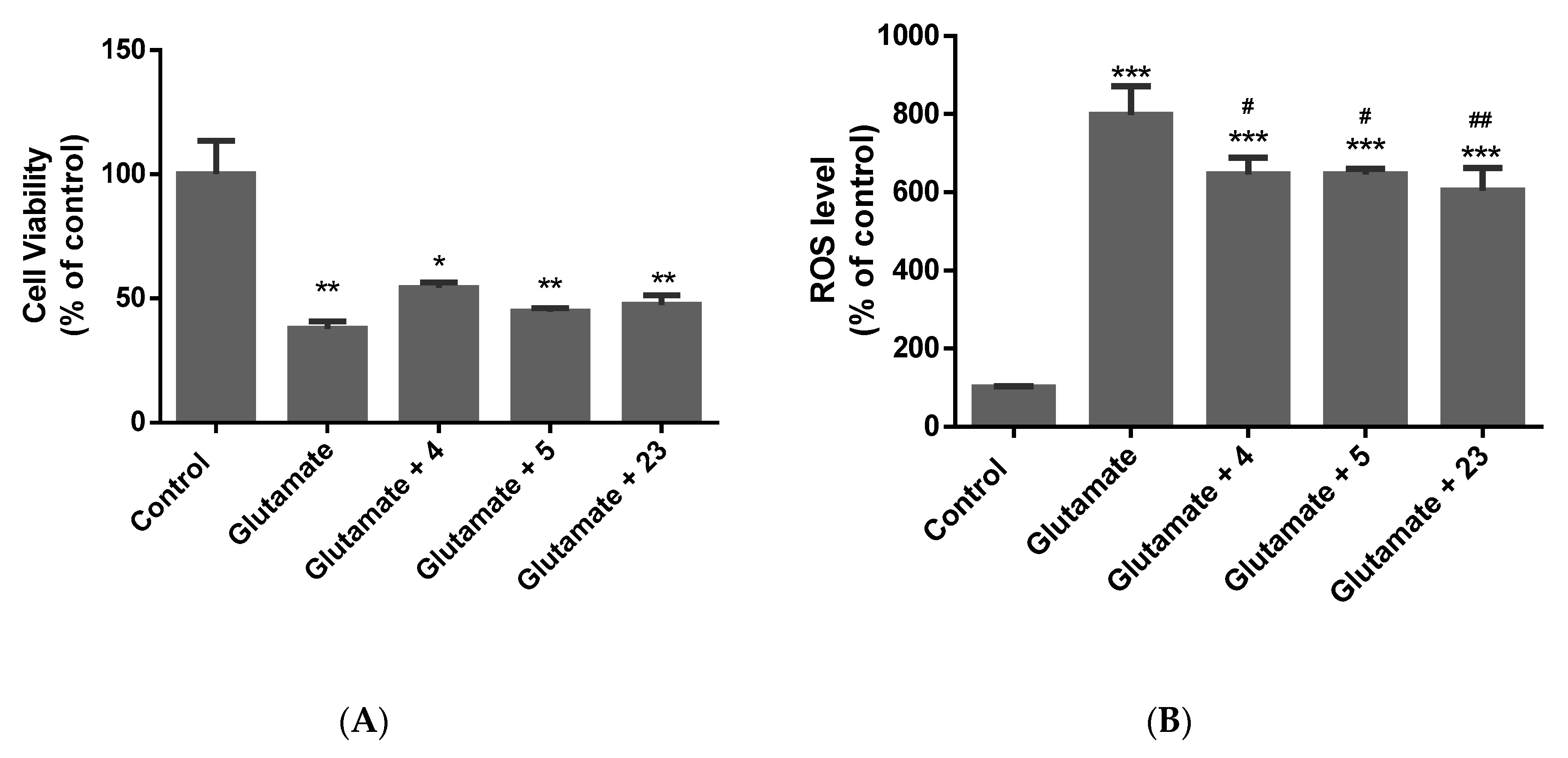
2.3. Determination of the Neuroprotective Activity
2.3.1. Oxidative Stress Model
2.3.2. Alzheimer’s Model
2.3.3. Parkinson’s Model
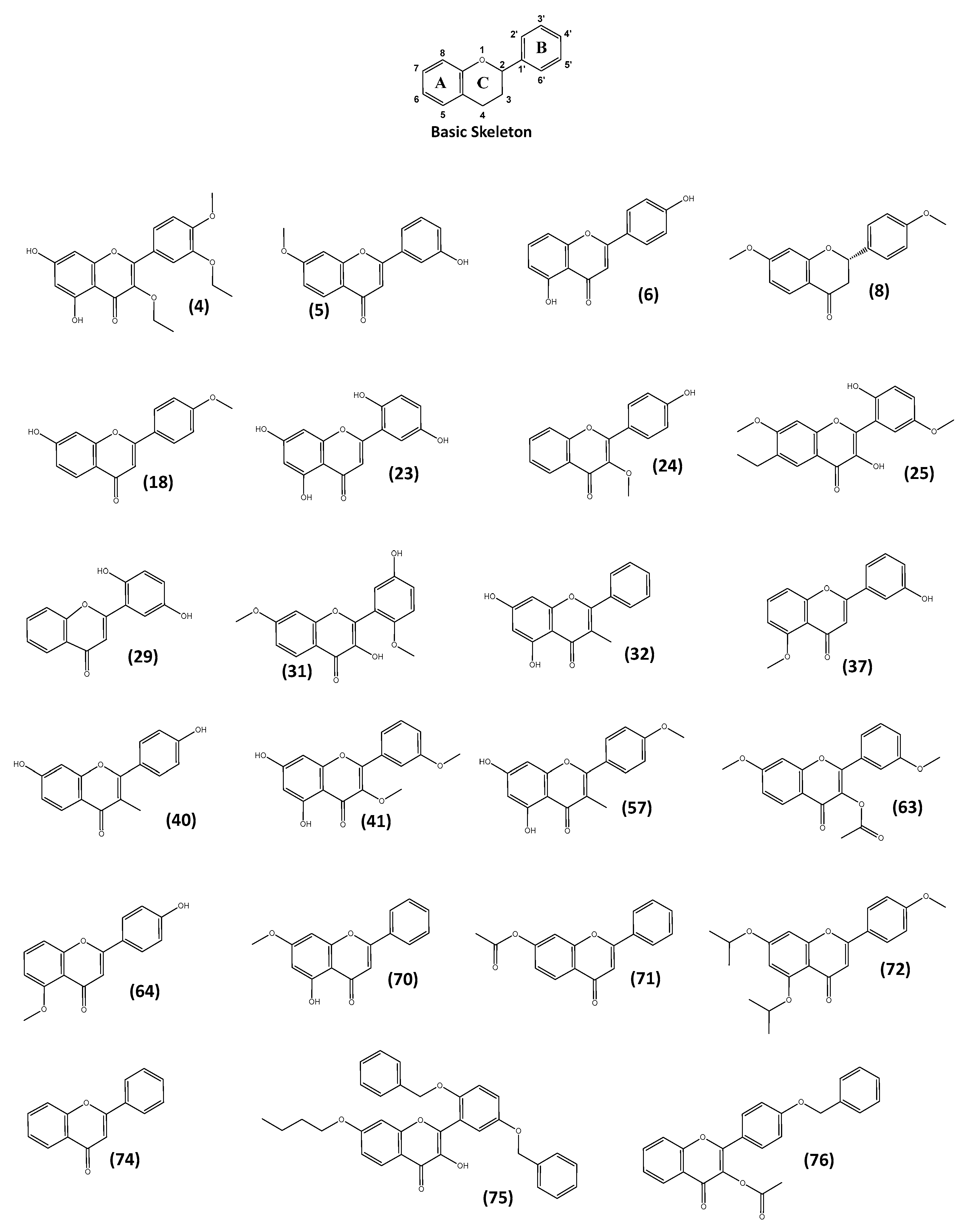
2.4. Structure–Activity Relationship (SAR) Analysis
2.4.1. Structure–Activity Relationship (SAR) Analysis Compared with Neuro-Differentiating Activity
2.4.2. Structure–Activity Relationship (SAR) Analysis Compared with Synaptogenic Activity
2.4.3. Structure–Activity Relationship (SAR) Analysis Compared with Antioxidant Activity
2.4.4. Structure–Activity Relationship (SAR) Analysis Compared with Anti-Inflammatory Activity
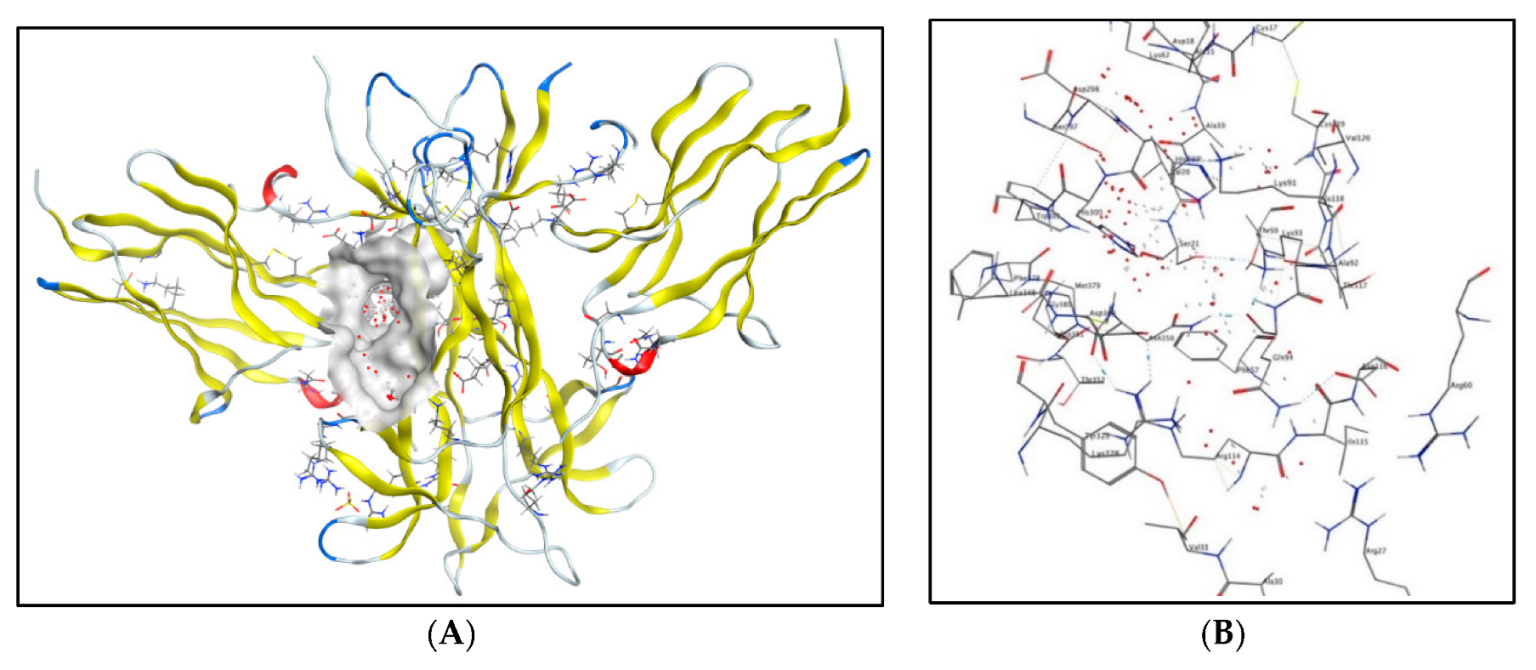
2.5. Molecular Docking Studies
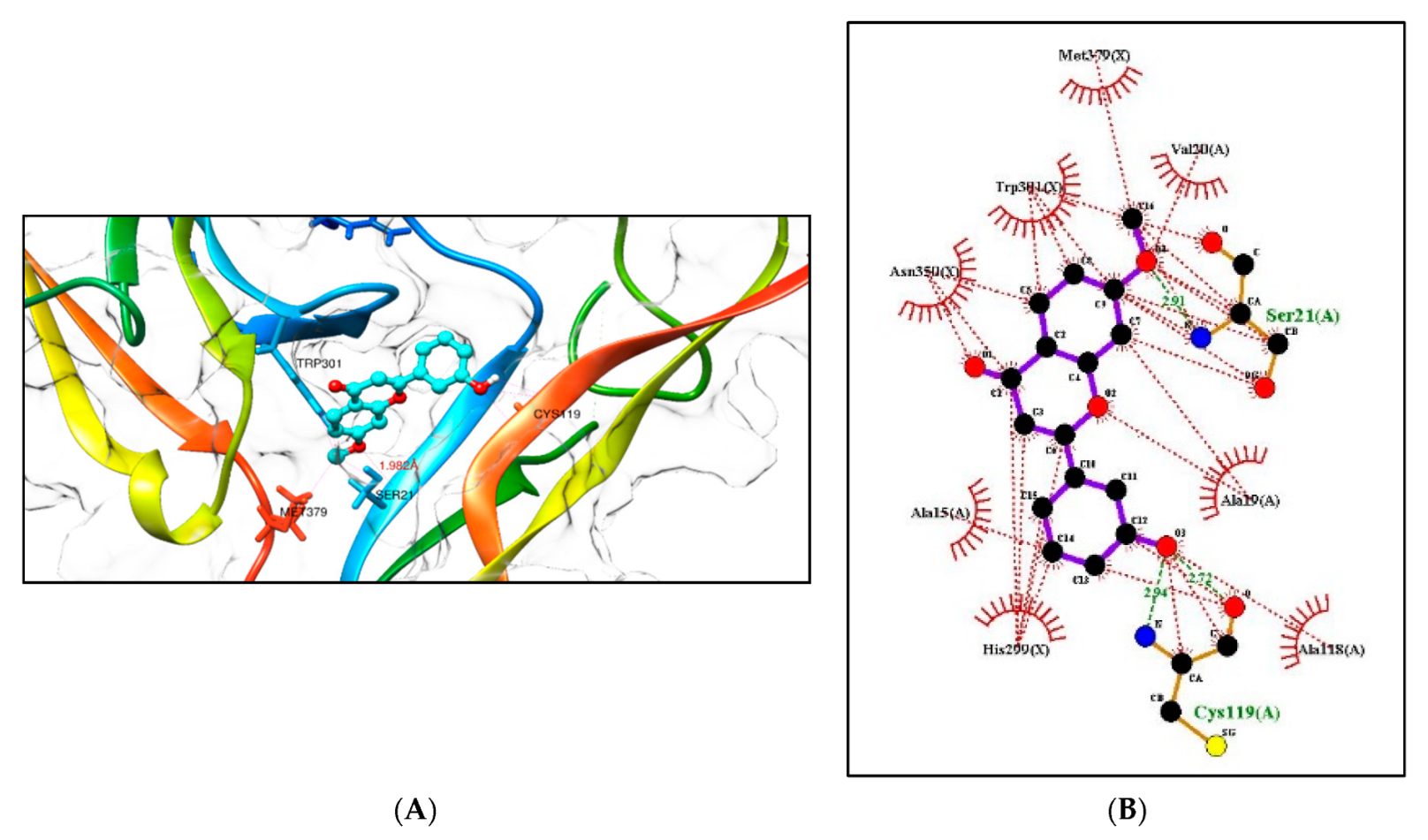
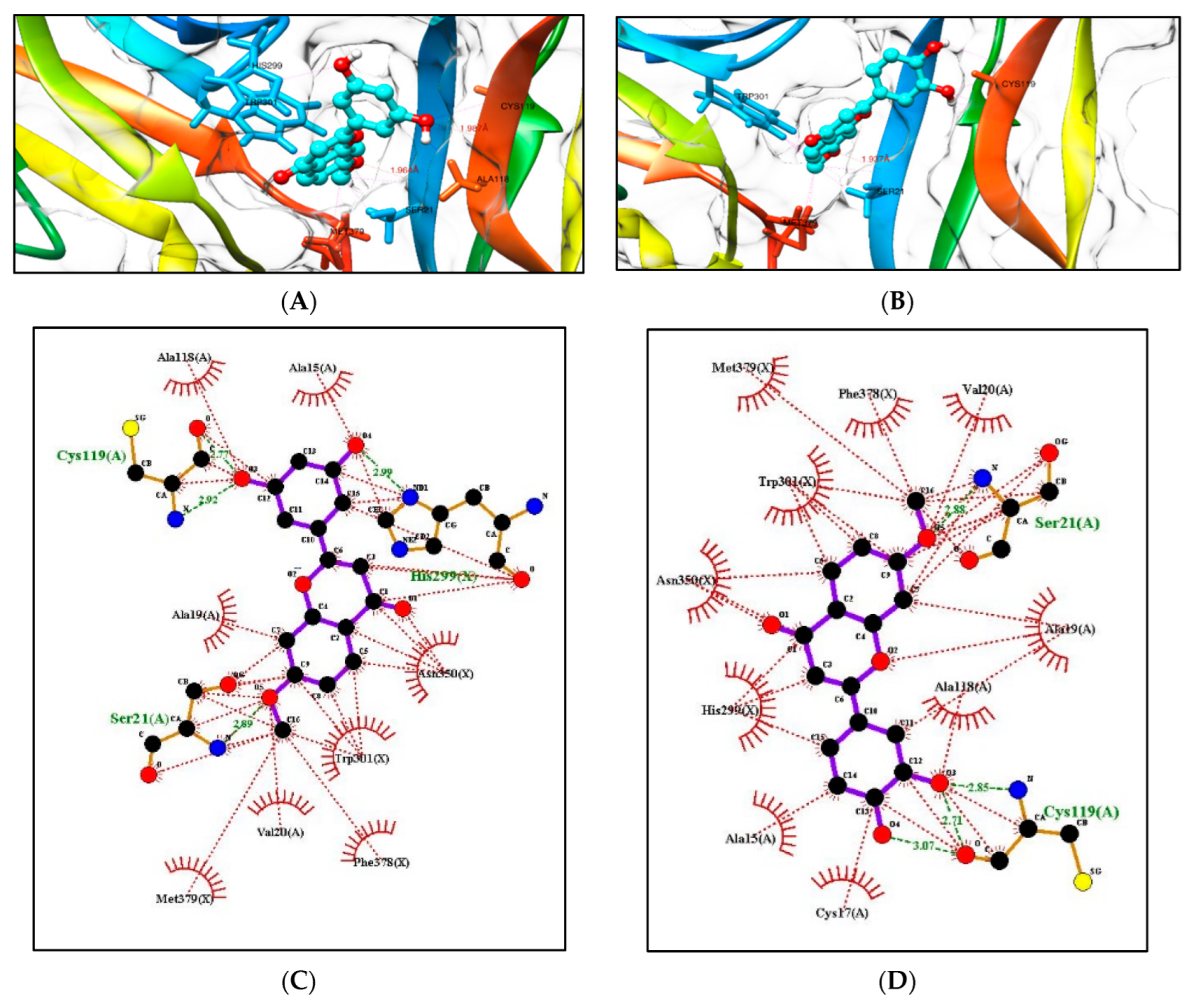
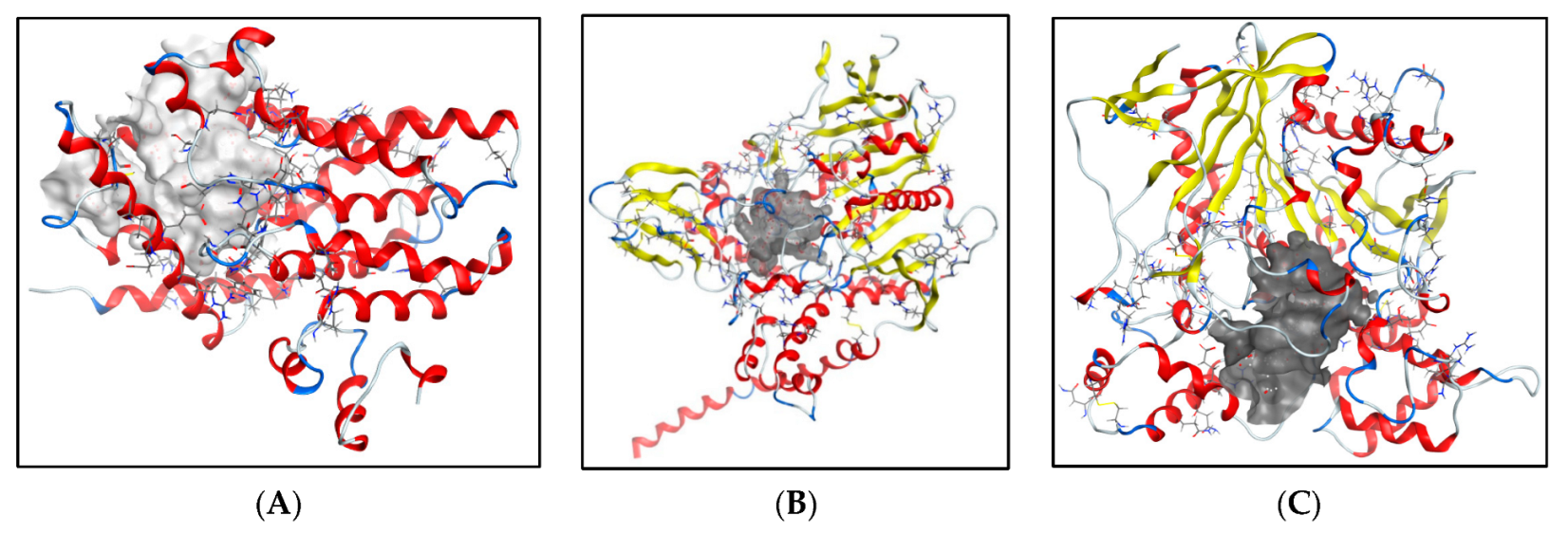
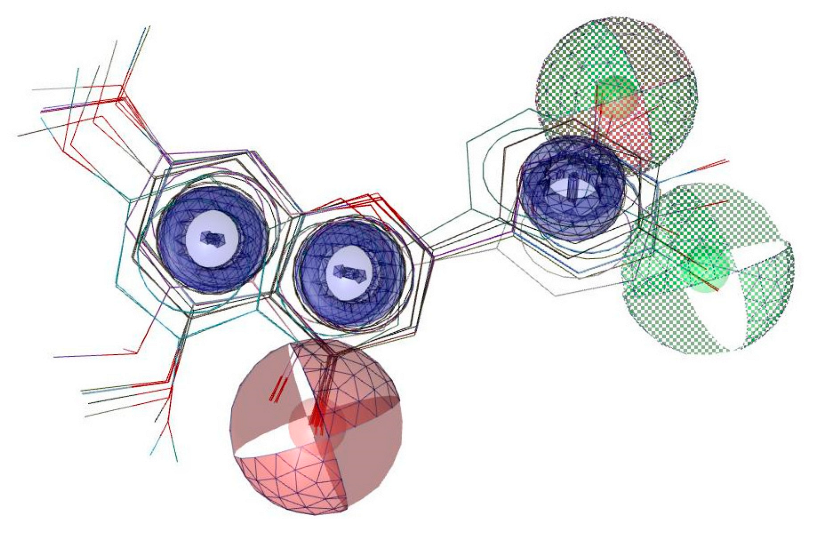
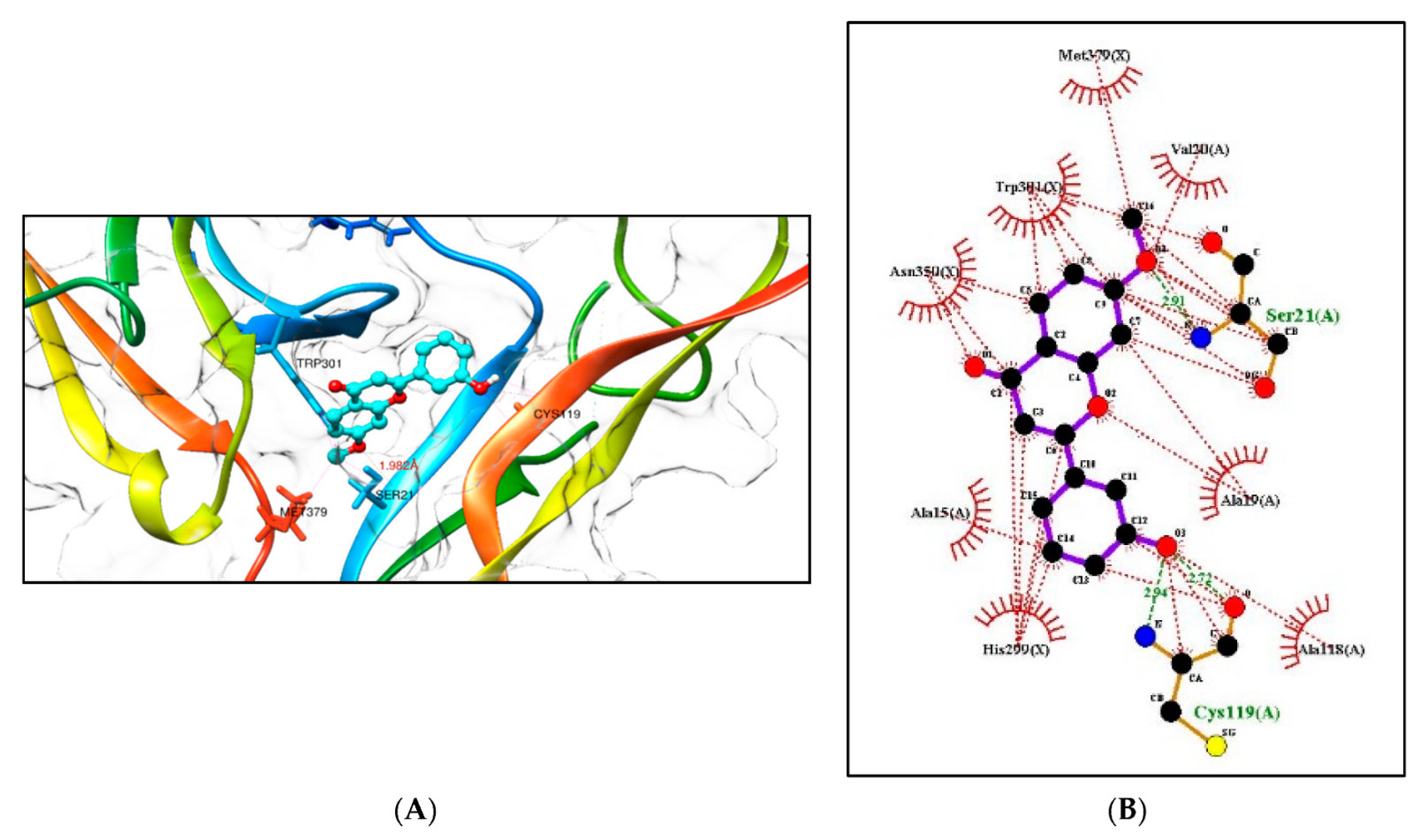
2.5.1. Molecular Docking Studies with the TRKB-d5 Domain (PDB: 1HCF)
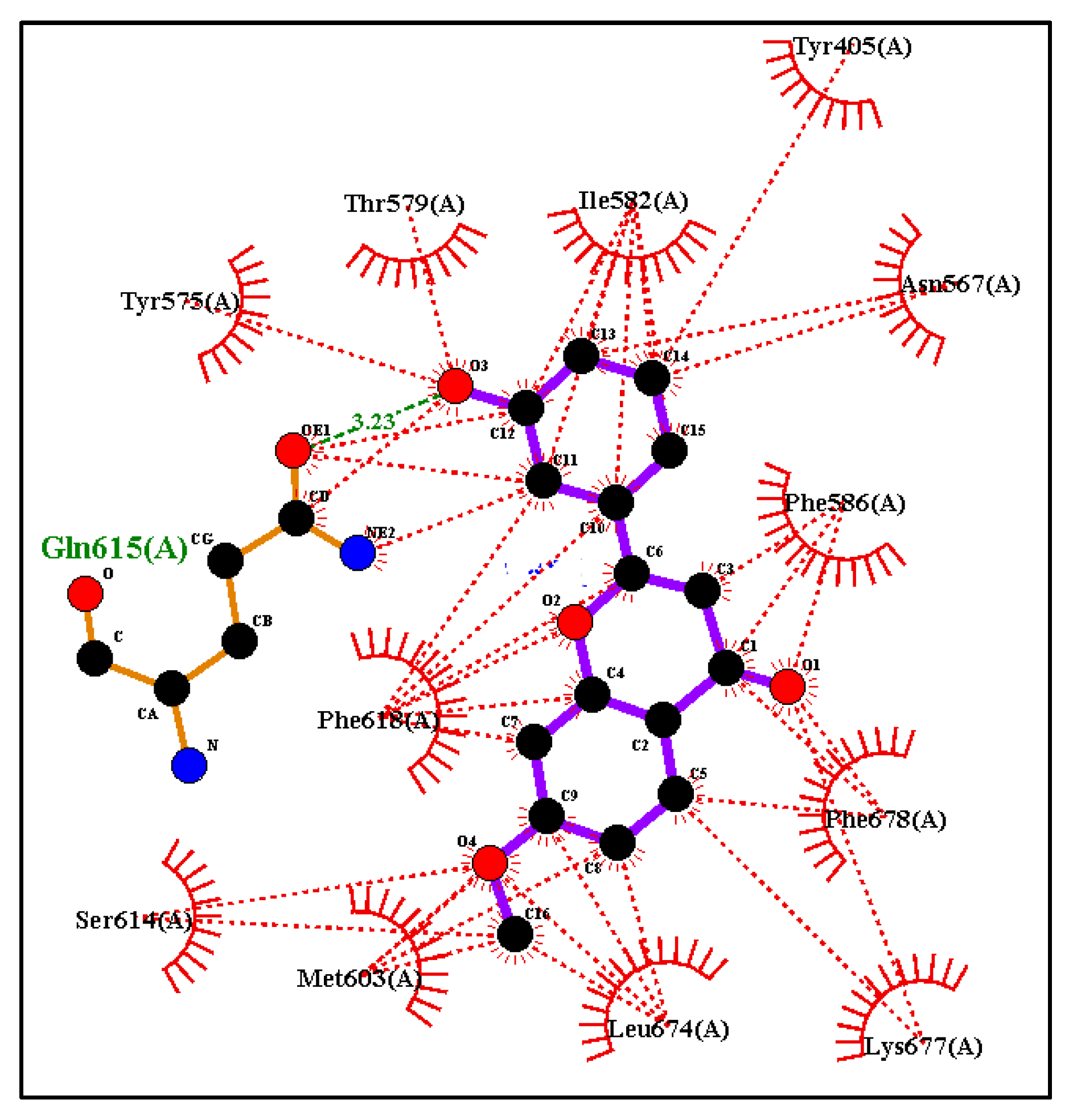
2.5.2. Additional Molecular Docking Studies with Human Phosphodiesterase 4B (PDB: 4MYQ) and AChE (PDB: 4BDT) and BuChE (PDB: 4BDS)
2.6. Pharmacophore Evaluation and ADMET Study
3. Materials and Methods
3.1. Origin of Compounds
3.2. Determination of the Highest Non-Toxic Dose of Compounds (Method S4.2)
3.3. Screening of Neurotrophic Activity of the Compounds (Method S4.3)
3.4. Assessment of the Neurotrophic and Synaptogenic Activities of the Candidates by High Content Screening (HCS) (Method S4.4)
3.5. Compound and Insult Treatment on Differentiated SH-SY5Y Cells (Method S4.5)
3.5.1. Assessment of Cell Viability and ROS Production (Method S4.5.1)
3.5.2. Investigation of Secretion of TNFα and IL-1β (Method S4.5.3)
3.6. Data Analysis and Visualisation
3.7. Molecular Docking Experiments
3.8. 3D Pharmacophore Model Generation and ADMET Study
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Przedborski, S.; Vila, M.; Jackson-Lewis, V. Neurodegeneration: What is it and where are we? J. Clin. Investig. 2003, 111, 3–10. [Google Scholar] [CrossRef]
- Young, J.J.; Lavakumar, M.; Tampi, D.; Balachandran, S.; Tampi, R.R. Frontotemporal dementia: Latest evidence and clinical implications. Ther. Adv. Psychopharmacol. 2017, 8, 33–48. [Google Scholar] [CrossRef]
- Alzheimer’s Disease Facts and Figures. Available online: https://www.alz.org/alzheimers-dementia/facts-figures (accessed on 21 June 2022).
- Parkinson’s Foundation. Available online: https://parkinson.org/Understanding-Parkinsons/Statistics (accessed on 21 June 2022).
- Huntington’s Disease. Available online: https://rarediseases.org/rare-diseases/huntingtons-disease (accessed on 21 June 2022).
- Centers for Disease Control and Prevention. Available online: https://www.cdc.gov/mmwr/volumes/67/wr/mm6707a3.htm (accessed on 21 June 2022).
- Abeliovich, A.; Gitler, A.D. Defects in trafficking bridge Parkinson’s disease pathology and genetics. Nature 2016, 539, 207–216. [Google Scholar] [CrossRef]
- Canter, R.G.; Penney, J.; Tsai, L.-H. The road to restoring neural circuits for the treatment of Alzheimer’s disease. Nature 2016, 539, 187–196. [Google Scholar] [CrossRef]
- Taylor, J.P.; Brown, R.H., Jr.; Cleveland, D.W. Decoding ALS: From genes to mechanism. Nature 2016, 539, 197–206. [Google Scholar] [CrossRef]
- Wyss-Coray, T. Ageing, neurodegeneration and brain rejuvenation. Nature 2016, 539, 180–186. [Google Scholar] [CrossRef]
- Focus on neurodegenerative disease. Nat. Neurosci. 2018, 21, 1293. [CrossRef]
- Nakajima, Y.; Suzuki, S. Environmental stresses induce misfolded protein aggregation in plant cells in a microtubule-dependent manner. Int. J. Mol. Sci. 2013, 14, 7771–7783. [Google Scholar] [CrossRef]
- Sprenkle, N.T.; Sims, S.G.; Sánchez, C.L.; Meares, G.P. Endoplasmic reticulum stress and inflammation in the central nervous system. Mol. Neurodegener. 2017, 12, 1–18. [Google Scholar] [CrossRef]
- Singh, A.; Kukreti, R.; Saso, L.; Kukreti, S. Oxidative Stress: A Key Modulator in Neurodegenerative Diseases. Molecules 2019, 24, 1583. [Google Scholar] [CrossRef]
- Höhn, A.; Tramutola, A.; Cascella, R. Proteostasis Failure in Neurodegenerative Diseases: Focus on Oxidative Stress. Oxidative Med. Cell. Longev. 2020, 2020, 5497046. [Google Scholar] [CrossRef]
- Clarke, J.R.; Lyra, E.; Silva, N.M.; Figueiredo, C.P.; Frozza, R.L.; Ledo, J.H.; Beckman, D.; Katashima, C.K.; Razolli, D.; Car-Valho, B.M.; et al. Alzheimer-associated Aβ oligomers impact the central nervous system to induce pe-ripheral metabolic deregulation. EMBO Mol. Med. 2015, 7, 190–210. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.J.; Zhang, X.; Chen, W.W. Role of oxidative stress in Alzheimer’s disease. Biomed. Rep. 2016, 4, 519–522. [Google Scholar] [CrossRef] [PubMed]
- Álvarez, A.; Cacabelos, R.; Sanpedro, C.; García-Fantini, M.; Aleixandre, M. Serum TNF-alpha levels are increased and correlate negatively with free IGF-I in Alzheimer disease. Neurobiol. Aging 2007, 28, 533–536. [Google Scholar] [CrossRef] [PubMed]
- Chang, R.; Yee, K.-L.; Sumbria, R.K. Tumor necrosis factor α Inhibition for Alzheimer’s Disease. J. Central Nerv. Syst. Dis. 2017, 9, 1179573517709278. [Google Scholar] [CrossRef] [PubMed]
- Jalili-Baleh, L.; Babaei, E.; Abdpour, S.; Bukhari, S.N.A.; Foroumadi, A.; Ramazani, A.; Sharifzadeh, M.; Abdollahi, M.; Khoobi, M. A review on flavonoid-based scaffolds as multi-target-directed ligands (MTDLs) for Alzheimer’s disease. Eur. J. Med. Chem. 2018, 152, 570–589. [Google Scholar] [CrossRef]
- Adelina, C. The costs of dementia: Advocacy, media, and stigma. In World Alzheimer Report 2019: Attitudes to Dementia; Alzheimer’s Disease International: London, UK, 2019; Chapter 3; pp. 100–101. Available online: https://www.alzint.org/u/WorldAlzheimerReport2019.pdf (accessed on 10 October 2022).
- Campora, M.; Francesconi, V.; Schenone, S.; Tasso, B.; Tonelli, M. Journey on Naphthoquinone and Anthraquinone Derivatives: New Insights in Alzheimer’s Disease. Pharmaceuticals 2021, 14, 33. [Google Scholar] [CrossRef] [PubMed]
- Anand, P.; Singh, B. Flavonoids as lead compounds modulating the enzyme targets in Alzheimer’s disease. Med. Chem. Res. 2012, 22, 3061–3075. [Google Scholar] [CrossRef]
- McGeer, E.G.; McGeer, P.L. Clinically tested drugs for Alzheimer’s disease. Expert Opin. Investig. Drugs 2003, 12, 1143–1151. [Google Scholar] [CrossRef]
- Osborn, G.G.; Saunders, A.V. Current treatments for patients with Alzheimer disease. J. Am. Osteopat. Assoc. 2010, 110, S16–S26. [Google Scholar]
- Hardy, J.; Selkoe, D.J. The amyloid hypothesis of Alzheimer’s disease: Progress and problems on the road to therapeutics. Science 2002, 297, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, S.; Atluri, V.; Kaushik, A.; Yndart, A.; Nair, M. Alzheimer’s disease: Pathogenesis, diagnostics, and therapeu-tics. Int. J. Nanomed. 2019, 14, 5541–5554. [Google Scholar] [CrossRef] [PubMed]
- De Ferrari, G.V.; Canales, M.A.; Shin, I.; Weiner, L.M.; Silman, I.; Inestrosa, N.C. A structural motif of acetylcholinesterase that promotes amyloid β-peptide fibril formation. Biochemistry 2001, 40, 10447–10457. [Google Scholar] [CrossRef]
- Soreq, H.; Seidman, S. Acetylcholinesterase—New roles for an old actor. Nat. Rev. Neurosci. 2001, 2, 294–302. [Google Scholar] [CrossRef]
- Pera, M.; Martínez-Otero, A.; Colombo, L.; Salmona, M.; Ruiz-Molina, D.; Badia, A.; Clos, M. Acetylcholinesterase as an amyloid enhancing factor in PrP82-146 aggregation process. Mol. Cell. Neurosci. 2009, 40, 217–224. [Google Scholar] [CrossRef] [PubMed]
- Hanger, D.P.; Hughes, K.; Woodgett, J.R.; Brion, J.-P.; Anderton, B.H. Glycogen synthase kinase-3 induces Alzheimer’s disease-like phosphorylation of tau: Generation of paired helical filament epitopes and neuronal localisation of the kinase. Neurosci. Lett. 1992, 147, 58–62. [Google Scholar] [CrossRef]
- Tibbo, A.J.; Baillie, G.S. Phosphodiesterase 4B: Master Regulator of Brain Signaling. Cells 2020, 9, 1254. [Google Scholar] [CrossRef]
- Gurney, M.E.; D’amato, E.C.; Burgin, A.B. Phosphodiesterase-4 (PDE4) molecular pharmacology and Alzheimer’s disease. Neurotherapeutics 2014, 12, 49–56. [Google Scholar] [CrossRef]
- Tibbo, A.J.; Tejeda, G.S.; Baillie, G.S. Understanding PDE4′s function in Alzheimer’s disease; a target for novel therapeutic approaches. Biochem. Soc. Trans. 2019, 47, 1557–1565. [Google Scholar] [CrossRef]
- Cai, D.; Deng, K.; Mellado, W.; Lee, J.; Ratan, R.R.; Filbin, M.T. Arginase I and polyamines act downstream from cyclic AMP in overcoming inhibition of axonal growth MAG and myelin in vitro. Neuron 2002, 35, 711–719. [Google Scholar] [CrossRef]
- Sorce, S.; Krause, K.-H. NOX Enzymes in the Central Nervous System: From Signaling to Disease. Antioxid. Redox Signal. 2009, 11, 2481–2504. [Google Scholar] [CrossRef] [PubMed]
- Shimohamaa, S.; Taninob, H.; Kawakamib, N.; Okamurac, N.; Kodamac, H.; Yamaguchid, T.; Hayakawad, T.; Nunomurae, A.; Chibae, S.; Perryf, G.; et al. Activation of NADPH oxidase in Alzheimer’s disease brains. Biochem. Biophys. Res. Commun. 2000, 273, 5–9. [Google Scholar] [CrossRef] [PubMed]
- Bruce-Keller, A.J.; Gupta, S.; Parrino, T.E.; Knight, A.G.; Ebenezer, P.J.; Weidner, A.M.; LeVine, H., 3rd; Keller, J.N.; Markesbery, W.R.; Shi, G.-X.; et al. NOX Activity is increased in mild cognitive impairment. Antioxid. Redox Signal. 2010, 12, 1371–1382. [Google Scholar] [CrossRef] [PubMed]
- Ishige, K.; Schubert, D.; Sagara, Y. Flavonoids protect neuronal cells from oxidative stress by three distinct mechanisms. Free Radic. Biol. Med. 2001, 30, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Kang, Y.; Lee, J.-H.; Seo, Y.H.; Jang, J.-H.; Jeong, C.-H.; Lee, S.; Jeong, G.-S.; Park, B. Epicatechin Prevents Methamphetamine-Induced Neuronal Cell Death via Inhibition of ER Stress. Biomol. Ther. 2019, 27, 145–151. [Google Scholar] [CrossRef]
- Shen, N.; Wang, T.; Gan, Q.; Liu, S.; Wang, L.; Jin, B. Plant flavonoids: Classification, distribution, biosynthesis, and antioxidant activity. Food Chem. 2022, 383, 132531. [Google Scholar] [CrossRef]
- Joseph, J.A.; Shukitt-Hale, B.; Denisova, N.A.; Prior, R.L.; Cao, G.; Martin, A.; Taglialatela, G.; Bickford, P.C. Long-term dietary strawberry, spinach, or vitamin e supplementation retards the onset of age-related neuronal signal-transduction and cognitive behavioral deficits. J. Neurosci. 1998, 18, 8047–8055. [Google Scholar] [CrossRef]
- Joseph, J.A.; Shukitt-Hale, B.; Denisova, N.A.; Bielinski, D.; Martin, A.; McEwen, J.J.; Bickford, P.C. Reversals of age-related declines in neuronal signal transduction, cognitive, and motor behavioral deficits with blueberry, spinach, or strawberry dietary supplementation. J. Neurosci. 1999, 19, 8114–8121. [Google Scholar] [CrossRef]
- Cichon, N.; Saluk-Bijak, J.; Gorniak, L.; Przyslo, L.; Bijak, M. Flavonoids as a Natural Enhancer of Neuroplasticity—An Overview of the Mechanism of Neurorestorative Action. Antioxidants 2020, 9, 1035. [Google Scholar] [CrossRef]
- Ichijo, H.; Nishida, E.; Irie, K.; ten Dijke, P.; Saitoh, M.; Moriguchi, T.; Takagi, M.; Matsumoto, K.; Miyazono, K.; Gotoh, Y. Induction of apoptosis by ASK1, a mammalian MAPKKK that activates SAPK/JNK and p38 signaling pathways. Science 1997, 275, 90–94. [Google Scholar] [CrossRef]
- Spencer, J.P.E.; Rice-Evans, C.; Williams, R.J. Modulation of pro-survival Akt/protein kinase B and ERK1/2 signaling cascades by quercetin and its in vivo metabolites underlie their action on neuronal viability. J. Biol. Chem. 2003, 278, 34783–34793. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.P.E.; Kuhnle, G.G.C.; Williams, R.J.; Rice-Evans, C. Intracellular metabolism and bioactivity of quercetin and its in vivo metabolites. Biochem. J. 2003, 372, 173–181. [Google Scholar] [CrossRef]
- Spencer, J.P.; Schroeter, H.; Crossthwaithe, A.J.; Kuhnle, G.; Williams, R.J.; Rice-Evans, C. Contrasting influences of glucuronidation and O -methylation of epicatechin on hydrogen peroxide-induced cell death in neurons and fibroblasts. Free Radic. Biol. Med. 2001, 31, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Miyake, Y.; Shimoi, K.; Kumazawa, S.; Yamamoto, K.; Kinae, N.; Osawa, T. Identification and antioxidant activity of flavonoid metabolites in plasma and urine of eriocitrin-treated rats. J. Agric. Food Chem. 2000, 48, 3217–3224. [Google Scholar] [CrossRef] [PubMed]
- Suganuma, M.; Okabe, S.; Oniyama, M.; Tada, Y.; Ito, H.; Fujiki, H. Wide distribution of [3H](-)-epigallocatechin gallate, a cancer preventive tea polyphenol, in mouse tissue. Carcinogen 1998, 19, 1771–1776. [Google Scholar] [CrossRef]
- De Gaetano, F.; Margani, F.; Barbera, V.; D’Angelo, V.; Germanò, M.P.; Pistarà, V.; Ventura, C.A. Characteriza-tion and In Vivo Antiangiogenic Activity Evaluation of Morin-Based Cyclodextrin Inclusion Complexes. Pharmaceutics 2023, 15, 2209. [Google Scholar] [CrossRef]
- Negahdari, R.; Bohlouli, S.; Sharifi, S.; Dizaj, S.M.; Saadat, Y.R.; Khezri, K.; Jafari, S.; Ahmadian, E.; Jahandizi, N.G.; Raeesi, S. Therapeutic benefits of rutin and its nanoformulations. Phytother. Res. 2020, 35, 1719–1738. [Google Scholar] [CrossRef]
- De Gaetano, F.; Celesti, C.; Paladini, G.; Venuti, V.; Cristiano, M.C.; Paolino, D.; Iannazzo, D.; Strano, V.; Gueli, A.M.; Tommasini, S.; et al. Solid Lipid Nanoparticles Containing Morin: Preparation, Characterization, and Ex Vivo Permeation Studies. Pharmaceutics 2023, 15, 1605. [Google Scholar] [CrossRef]
- Wang, C.-N.; Chi, C.-W.; Lin, Y.-L.; Chen, C.-F.; Shiao, Y.-J. The neuroprotective effects of phytoestrogens on amyloid β protein-induced toxicity are mediated by abrogating the activation of caspase cascade in rat cortical neurons. J. Biol. Chem. 2001, 276, 5287–5295. [Google Scholar] [CrossRef]
- Jung, M.; Park, M. Acetylcholinesterase inhibition by flavonoids from agrimonia Pilosa. Molecules 2007, 12, 2130–2139. [Google Scholar] [CrossRef]
- Kim, H.; Park, B.-S.; Lee, K.-G.; Choi, C.Y.; Jang, S.S.; Kim, Y.-H.; Lee, S.-E. Effects of naturally occurring compounds on fibril formation and oxidative stress of beta-amyloid. J. Agric. Food Chem. 2005, 53, 8537–8541. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.T.T.; Choi, R.C.Y.; Chu, G.K.Y.; Cheung, A.W.H.; Gao, Q.T.; Li, J.; Jiang, Z.Y.; Dong, T.T.X.; Tsim, K.W.K. Flavonoids possess neuroprotective effects on cultured pheochromocytoma pc12 cells: A comparison of different flavonoids in activating estrogenic effect and in preventing β-amyloid-induced Cell Death. J. Agric. Food Chem. 2007, 55, 2438–2445. [Google Scholar] [CrossRef]
- Jeon, S.-Y.; Bae, K.; Seong, Y.-H.; Song, K.-S. Green tea catechins as a BACE1 (β-Secretase) inhibitor. Bioorgan. Med. Chem. Lett. 2003, 13, 3905–3908. [Google Scholar] [CrossRef] [PubMed]
- Rezai-Zadeh, K.; Shytle, R.D.; Bai, Y.; Tian, J.; Hou, H.; Mori, T.; Zeng, J.; Obregon, D.; Town, T. Flavonoid-mediated presenilin-1 phosphorylation reduces Alzheimer’s disease β-amyloid production. J. Cell. Mol. Med. 2009, 13, 574–588. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Cui, J.; Lee, J.C.-M.; Ding, S.; Chalimoniuk, M.; Simonyi, A.; Sun, A.Y.; Gu, Z.; Weisman, G.A.; Wood, W.G.; et al. Prolonged exposure of cortical neurons to oligomeric amyloid-β impairs nmda receptor function via nadph oxidase-mediated ros production: Protective effect of green tea (-)-epigallocatechin-3-gallate. ASN Neuro 2010, 3, AN20100025. [Google Scholar] [CrossRef] [PubMed]
- Richetti, S.; Blank, M.; Capiotti, K.; Piato, A.; Bogo, M.; Vianna, M.; Bonan, C. Quercetin and rutin prevent scopolamine-induced memory impairment in zebrafish. Behav. Brain Res. 2011, 217, 10–15. [Google Scholar] [CrossRef]
- Hsieh, H.-M.; Wu, W.-M.; Hu, M.-L. Soy isoflavones attenuate oxidative stress and improve parameters related to aging and Alzheimer’s disease in C57BL/6J mice treated with d-galactose. Food Chem. Toxicol. 2009, 47, 625–632. [Google Scholar] [CrossRef]
- Zhu, J.T.; Choi, R.C.; Xie, H.Q.; Zheng, K.Y.; Guo, A.J.; Bi, C.W.; Lau, D.T.; Li, J.; Dong, T.T.; Lau, B.W.; et al. Hibifolin, a flavonol glycoside, prevents β-amyloid-induced neurotoxicity in cultured cortical neurons. Neurosci. Lett. 2009, 461, 172–176. [Google Scholar] [CrossRef]
- Zeng, K.-W.; Ko, H.; Yang, H.O.; Wang, X.-M. Icariin attenuates β-amyloid-induced neurotoxicity by inhibition of tau protein hyperphosphorylation in PC12 cells. Neuropharmacology 2010, 59, 542–550. [Google Scholar] [CrossRef]
- Kim, J.K.; Choi, S.J.; Cho, H.Y.; Hwang, H.-J.; Kim, Y.J.; Lim, S.T.; Kim, C.-J.; Kim, H.K.; Peterson, S.; Shin, D.-H. Protective effects of kaempferol (3,4′,5,7-tetrahydroxyflavone) against amyloid beta peptide (aβ)-induced neurotoxicity in ICR mice. Biosci. Biotechnol. Biochem. 2010, 74, 397–401. [Google Scholar] [CrossRef]
- Devi, S.; Kumar, V.; Singh, S.K.; Dubey, A.K.; Kim, J.J. Flavonoids: Potential Candidates for the Treatment of Neuro-degenerative Disorders. Biomedicines 2021, 9, 99. [Google Scholar] [CrossRef] [PubMed]
- Jones, Q.R.; Warford, J.; Rupasinghe, H.V.; Robertson, G.S. Target-based selection of flavonoids for neurodegenerative disorders. Trends Pharmacol. Sci. 2012, 33, 602–610. [Google Scholar] [CrossRef] [PubMed]
- Duarte, A.C.B.; Santana, M.G.; Orfali, G.d.C.; de Oliveira, C.T.P.; Priolli, D.G. Literature Evidence and ARRIVE Assessment on Neuroprotective Effects of Flavonols in Neurodegenerative Diseases’ Models. CNS Neurol. Disord. Drug Targets 2018, 17, 34–42. [Google Scholar] [CrossRef] [PubMed]
- de Andrade Teles, R.B.; Diniz, T.C.; Costa Pinto, T.C.; de Oliveira Junior, R.G.; Gama, E.S.M.; de Lavor, E.M.; Fernandes, A.W.C.; de Oliveira, A.P.; de Almeida Ribeiro, F.P.R.; da Silva, A.A.M.; et al. Flavonoids as Therapeutic Agents in Alzheimer’s and Parkinson’s Diseases: A Systematic Review of Preclinical Evidences. Oxid. Med. Cell. Longev. 2018, 2018, 7043213. [Google Scholar] [CrossRef]
- Kujawska, M.; Jodynis-Liebert, J. Polyphenols in Parkinson’s Disease: A Systematic Review of In Vivo Studies. Nutrients 2018, 10, 642. [Google Scholar] [CrossRef]
- Jang, S.-W.; Liu, X.; Yepes, M.; Shepherd, K.R.; Miller, G.W.; Liu, Y.; Wilson, W.D.; Xiao, G.; Blanchi, B.; Sun, Y.E.; et al. A selective TrkB agonist with potent neurotrophic activities by 7,8-dihydroxyflavone. Proc. Natl. Acad. Sci. USA 2010, 107, 2687–2692. [Google Scholar] [CrossRef]
- Sander, T.; Freyss, J.; Von Korff, M.; Rufener, C. DataWarrior: An Open-Source Program for Chemistry Aware Data Visualization and Analysis. J. Chem. Inf. Model. 2015, 55, 460–473. [Google Scholar] [CrossRef]
- Wang, T.-Y.; Li, Q.; Bi, K.-S. Bioactive flavonoids in medicinal plants: Structure, activity and biological fate. Asian J. Pharm. Sci. 2017, 13, 12–23. [Google Scholar] [CrossRef]
- Echeverry, C.; Arredondo, F.; Abin-Carriquiry, J.A.; Midiwo, J.O.; Ochieng, C.; Kerubo, L.; Dajas, F. Pretreatment with natural flavones and neuronal cell survival after oxidative stress: A structure−activity relationship study. J. Agric. Food Chem. 2010, 58, 2111–2115. [Google Scholar] [CrossRef]
- Ullah, A.; Munir, S.; Badshah, S.L.; Khan, N.; Ghani, L.; Poulson, B.G.; Emwas, A.-H.; Jaremko, M. Important Flavonoids and Their Role as a Therapeutic Agent. Molecules 2020, 25, 5243. [Google Scholar] [CrossRef]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.-H.; Yang, Z.-S.; Wen, C.-C.; Chang, Y.-S.; Wang, B.-C.; Hsiao, C.-A.; Shih, T.-L. Evaluation of the structure–activity relationship of flavonoids as antioxidants and toxicants of zebrafish larvae. Food Chem. 2012, 134, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Yoshida, N.; Ishibashi, K.; Haratake, M.; Arano, Y.; Mori, H.; Nakayama, M. Radioiodinated Flavones for In Vivo Imaging of β-Amyloid Plaques in the Brain. J. Med. Chem. 2005, 48, 7253–7260. [Google Scholar] [CrossRef]
- Chirumbolo, S. The Role of Quercetin, Flavonols and Flavones in Modulating Inflammatory Cell Function. Inflamm. Allergy-Drug Targets 2010, 9, 263–285. [Google Scholar] [CrossRef]
- Çelik, H.; Koşar, M. Inhibitory effects of dietary flavonoids on purified hepatic NADH-cytochrome b5 reductase: Structure–activity relationships. Chem. Interact. 2012, 197, 103–109. [Google Scholar] [CrossRef] [PubMed]
- During, A.; Larondelle, Y. The O-methylation of chrysin markedly improves its intestinal anti-inflammatory properties: Structure–activity relationships of flavones. Biochem. Pharmacol. 2013, 86, 1739–1746. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A.J. AutoDock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef]
- Molecular Operating Environment (MOE); Chemical Computing Group ULC: Montreal, QC, Canada, 2022.
- Banfield, M.J.; Naylor, R.L.; Robertson, A.G.; Allen, S.J.; Dawbarn, D.; Brady, R. Specificity in trk receptor: Neurotrophin interactions: The crystal structure of trkb-d5 in complex with neurotrophin-4/5. Structure 2001, 9, 1191–1199. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera? A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef]
- Schrödinger, L.; De Lano, W. PyMOL. 2020. Available online: http://www.pymol.org/pymol (accessed on 10 October 2022).
- Laskowski, R.A.; Swindells, M.B. LigPlot+: Multiple ligand–protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 2011, 51, 2778–2786. [Google Scholar] [CrossRef]
- Fox, D.; Burgin, A.B.; Gurney, M.E. Structural basis for the design of selective phosphodiesterase 4B inhibitors. Cell. Signal. 2014, 26, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Nachon, F.; Carletti, E.; Ronco, C.; Trovaslet, M.; Nicolet, Y.; Jean, L.; Renard, P.-Y. Crystal structures of human cholinesterases in complex with huprine W and tacrine: Elements of specificity for anti-Alzheimer’s drugs targeting acetyl- and butyryl-cholinesterase. Biochem. J. 2013, 453, 393–399. [Google Scholar] [CrossRef] [PubMed]
- Wolber, G.; Langer, T. LigandScout: 3-D pharmacophores derived from protein-bound ligands and their use as virtual screening filters. J. Chem. Inf. Model. 2004, 45, 160–169. [Google Scholar] [CrossRef] [PubMed]
- Seidel, T.; Ibis, G.; Bendix, F.; Wolber, G. Strategies for 3D pharmacophore-based virtual screening. Drug Discov. Today Technol. 2011, 7, e221–e228. [Google Scholar] [CrossRef] [PubMed]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Ring A | Ring B | Docking Score (kcal/mol) | ||||
|---|---|---|---|---|---|---|---|
| C5 | C7 | C2′ | C3′ | C4′ | C5′ | ||
| 5 | OCH3 | OH | −9.5 | ||||
| 4 | OH | OH | OCH3 | OCH2CH3 | −8.4 | ||
| 23 | OH | OH | OH | OH | −9.4 | ||
| Compound | Ring A | Ring B | Docking Score (kcal/mol) | ||||
|---|---|---|---|---|---|---|---|
| C5 | C7 | C2′ | C3′ | C4′ | C5′ | ||
| 5 | OCH3 | OH | −9.5 | ||||
| 5A | OCH3 | OCH3 | OH | −8.5 | |||
| 5B | OCH3 | OH | −8.8 | ||||
| 5C | OCH3 | OH | OH | −9.7 | |||
| 5D | OCH3 | OCH3 | OH | OH | −8.7 | ||
| 5E | OCH3 | OH | OH | −8.9 | |||
| 5F | OCH3 | OH | −9.4 | ||||
| 5G | OCH3 | OCH3 | OH | −8.3 | |||
| 5H | OCH3 | OH | −8.7 | ||||
| 5I | OCH3 | OH | OH | −9.6 | |||
| 5J | OCH3 | OCH3 | OH | OH | −8.6 | ||
| 5K | OCH3 | OH | OH | −9 | |||
| 5L | OCH3 | OH | OH | −9.5 | |||
| 5M | OCH3 | OCH3 | OH | OH | −8.6 | ||
| 5N | OCH3 | OH | OH | −9 | |||
| 5O | OCH3 | OH | OH | OH | −8.9 | ||
| 5P | OCH3 | OCH3 | OH | OH | OH | −8.4 | |
| 5Q | OCH3 | OH | OH | OH | −8.2 | ||
| Docking Score (with -OCH3) on Ring A (kcal/mol) | Docking Score (Replacing -OCH3 with -OH) on Ring A (kcal/mol) |
|---|---|
| −9.7 | −9.6 |
| −9.4 | −9.3 |
| −9.6 | −9.5 |
| −9.5 | −9.5 |
| Compound | Ring A | Ring B | Docking Score (kcal/mol) | ||||
|---|---|---|---|---|---|---|---|
| C5 | C7 | C2′ | C3′ | C4′ | C5′ | ||
| 5 | OCH3 | OH | −10.2 | ||||
| 4 | OH | OH | OCH3 | OCH2CH3 | −8.9 | ||
| 23 | OH | OH | OH | OH | −10.0 | ||
| Compound | Ring A | Ring B | Docking Score (kcal/mol) PDB:4BDT | Docking Score (kcal/mol) PDB:4BDS | ||||
|---|---|---|---|---|---|---|---|---|
| C5 | C7 | C2′ | C3′ | C4′ | C5′ | |||
| 5 | OCH3 | OH | −9.5 | −9.3 | ||||
| 4 | OH | OH | OCH3 | OCH2CH3 | −8.0 | −8.5 | ||
| 23 | OH | OH | OH | OH | −9.8 | −9.1 | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Preet, G.; Haj Hasan, A.; Ramlagan, P.; Fawdar, S.; Boulle, F.; Jaspars, M. Anti-Neurodegenerating Activity: Structure–Activity Relationship Analysis of Flavonoids. Molecules 2023, 28, 7188. https://doi.org/10.3390/molecules28207188
Preet G, Haj Hasan A, Ramlagan P, Fawdar S, Boulle F, Jaspars M. Anti-Neurodegenerating Activity: Structure–Activity Relationship Analysis of Flavonoids. Molecules. 2023; 28(20):7188. https://doi.org/10.3390/molecules28207188
Chicago/Turabian StylePreet, Gagan, Ahlam Haj Hasan, Piteesha Ramlagan, Shameem Fawdar, Fabien Boulle, and Marcel Jaspars. 2023. "Anti-Neurodegenerating Activity: Structure–Activity Relationship Analysis of Flavonoids" Molecules 28, no. 20: 7188. https://doi.org/10.3390/molecules28207188
APA StylePreet, G., Haj Hasan, A., Ramlagan, P., Fawdar, S., Boulle, F., & Jaspars, M. (2023). Anti-Neurodegenerating Activity: Structure–Activity Relationship Analysis of Flavonoids. Molecules, 28(20), 7188. https://doi.org/10.3390/molecules28207188