

Anti-Inflammatory Activity of 1,6,7-Trihydroxy-2-(1,1-dimethyl-2-propenyl)-3-methoxyxanthone Isolated from Cudrania tricuspidata via NF-κB, MAPK, and HO-1 Signaling Pathways in Lipopolysaccharide-Stimulated RAW 264.7 and BV2 Cells


 , ,
, ,
Abstract
:1. Introduction
2. Results
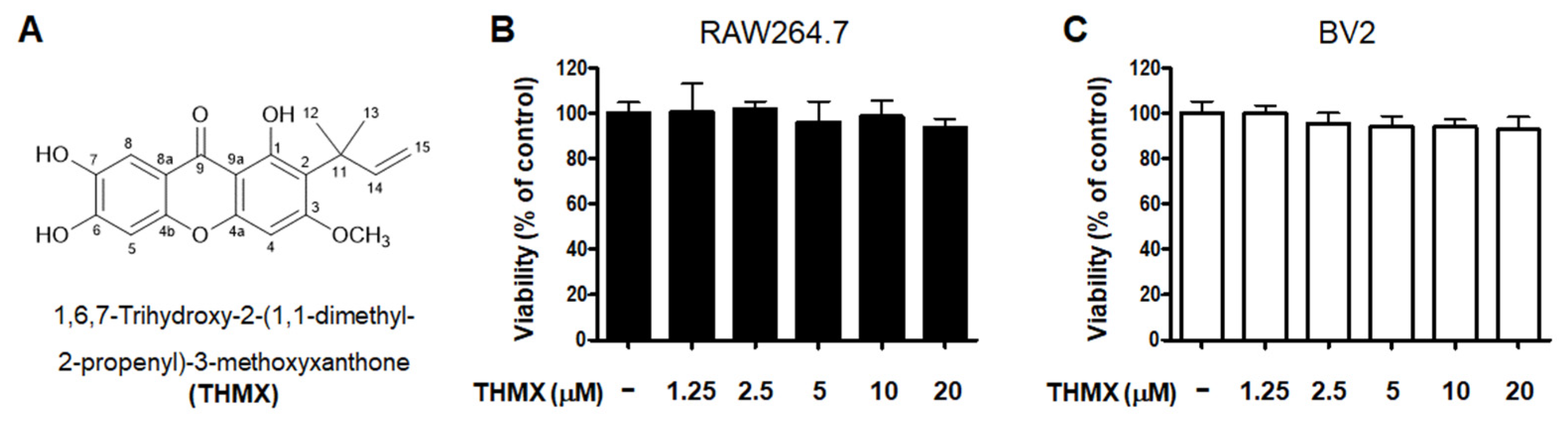
2.1. Effects of THMX on the Viability of RAW264.7 and BV2 Cells
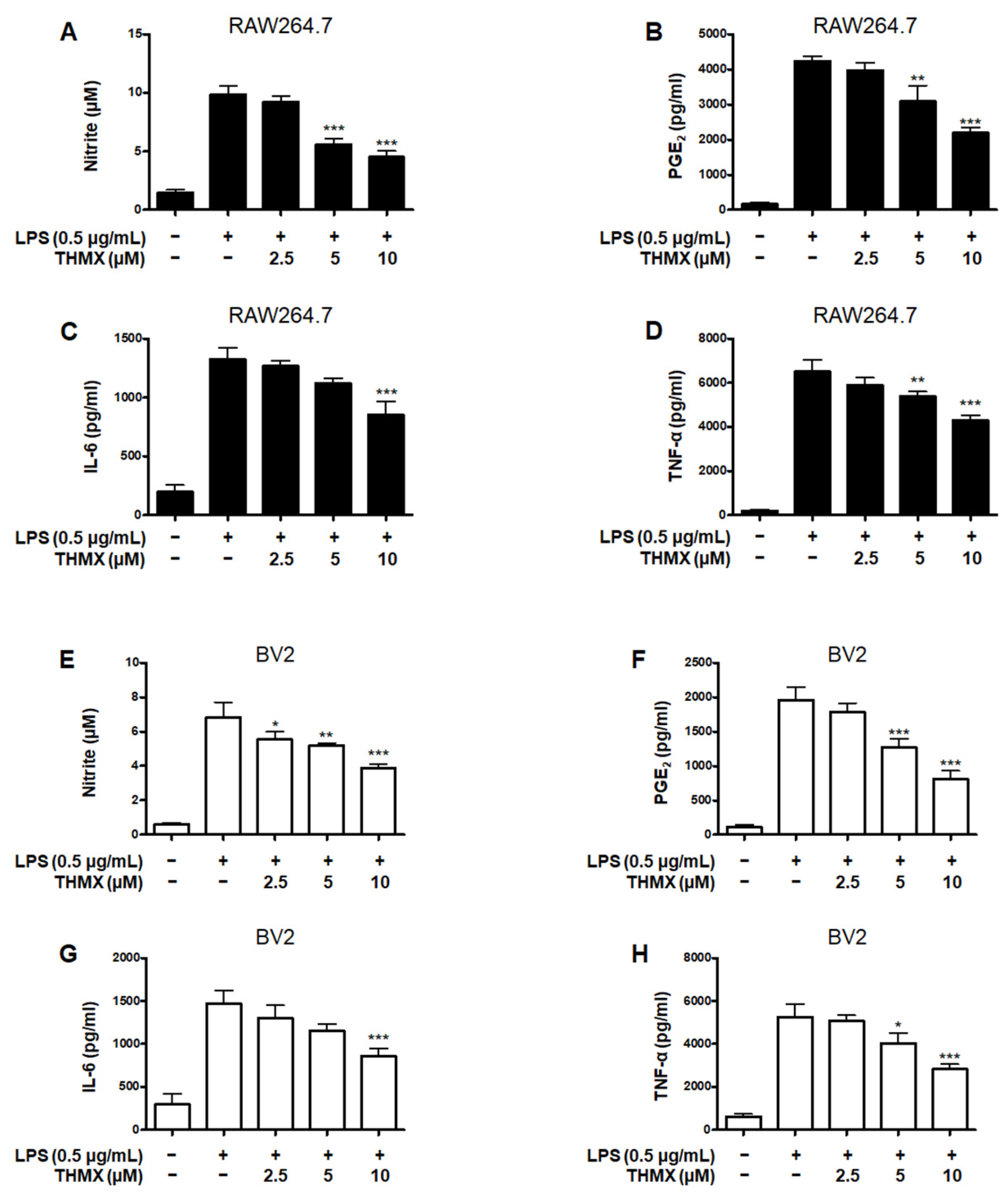
2.2. Effects of THMX on Pro-Inflammatory Factors in RAW 264.7 and BV2 Cells
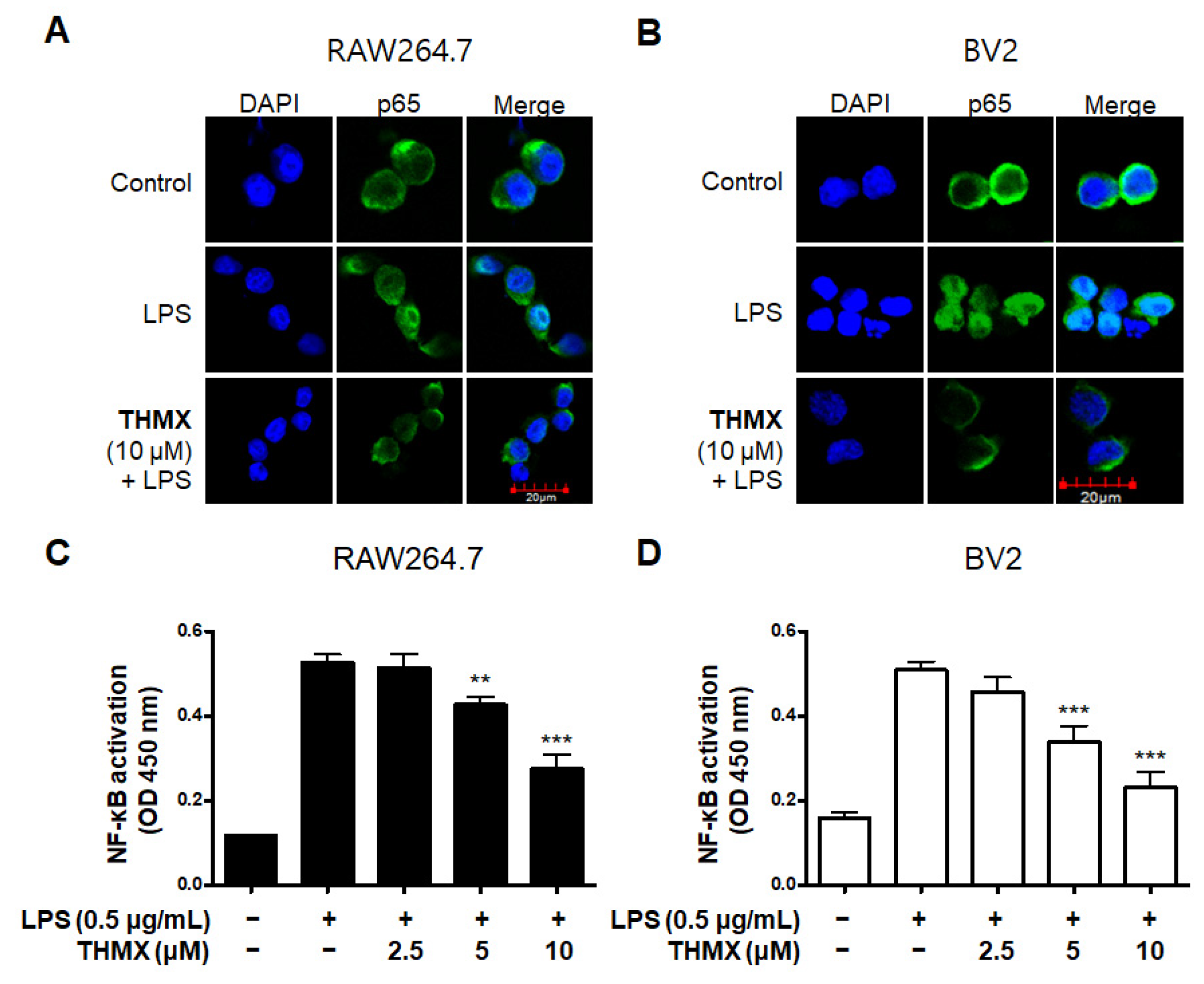
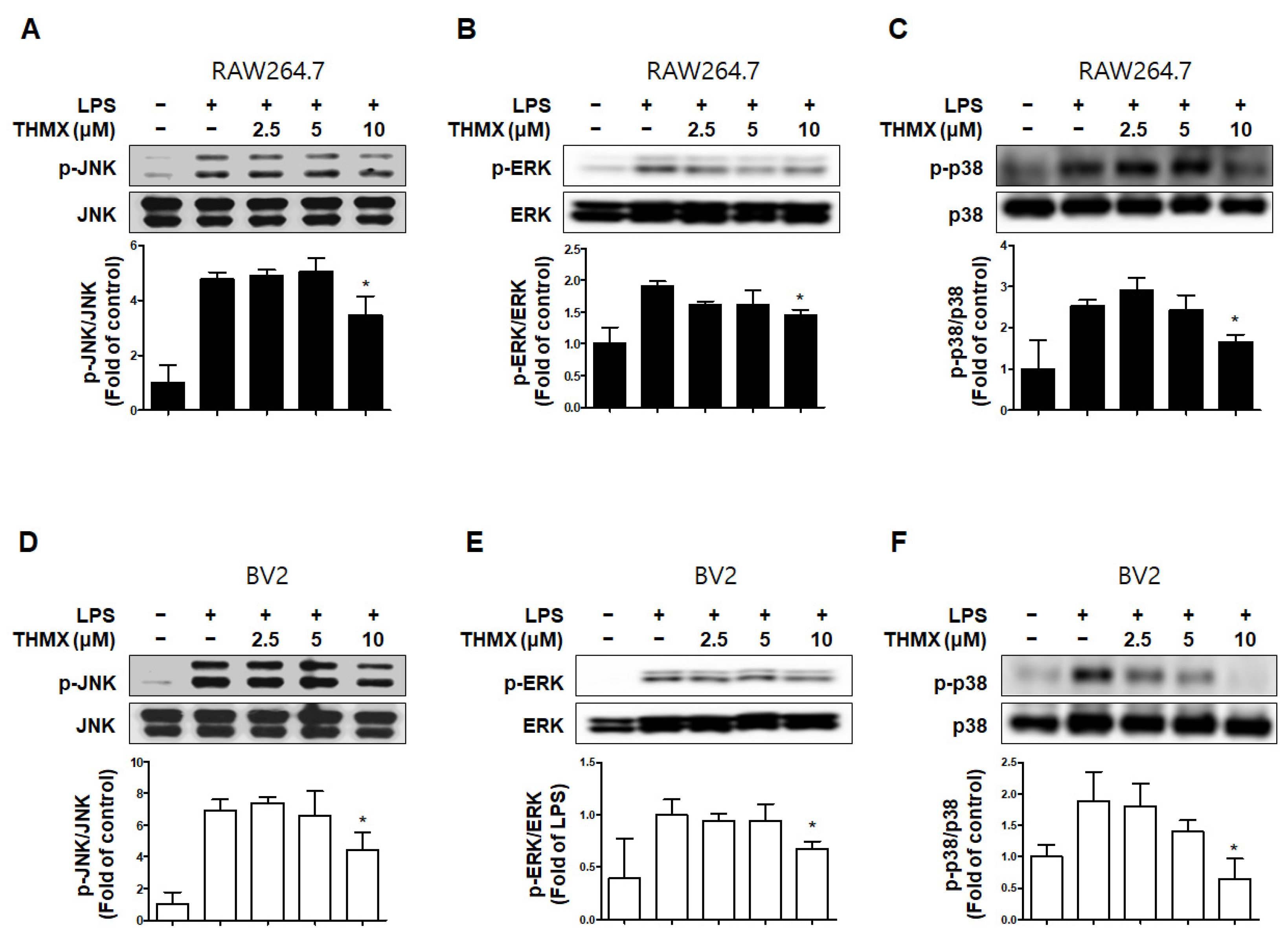
2.3. Effects of THMX on p65 Translation and MAPK Signaling Pathway in RAW 264.7 and BV2 Cells
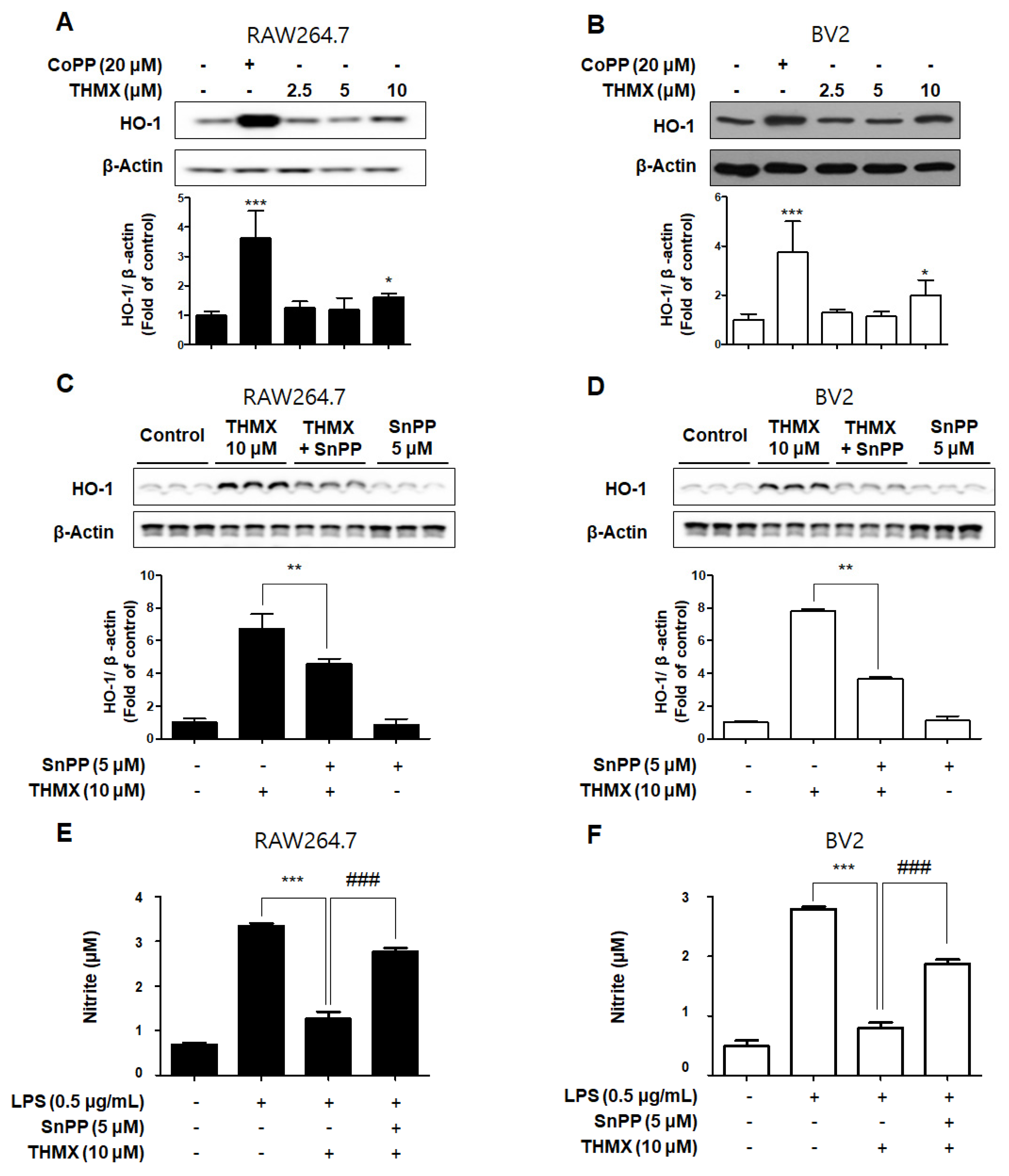
2.4. Effects of THMX on HO-1 in RAW 264.7 and BV2 Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture and Viability Assays
4.3. Measurement of Nitrite Oxide (NO) Generation
4.4. PGE2 Assay
4.5. Assays for IL-6 and TNF-α
4.6. Western Blotting Analysis
4.7. NF-κB Localization and Immunofluorescence
4.8. Measurement of DNA-Binding Activity of NF-κB
4.9. Statistical Analysis
5. Conclusion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Sampson, T.R.; Debelius, J.W.; Thron, T.; Janssen, S.; Shastri, G.G.; Ilhan, Z.E.; Challis, C.; Schretter, C.E.; Rocha, S.; Gradinaru, V.; et al. Gut microbiota regulate motor deficits and neuroinflammation in a model of Parkinson’s disease. Cell 2016, 167, 1469–1480.e12. [Google Scholar] [CrossRef] [PubMed]
- Shao, W.; Zhang, S.-Z.; Tang, M.; Zhang, X.-H.; Zhou, Z.; Yin, Y.-Q.; Zhou, Q.-B.; Huang, Y.-Y.; Liu, Y.-J.; Wawrousek, E.; et al. Suppression of neuroinflammation by astrocytic dopamine D2 receptors via αB-crystallin. Nature 2013, 494, 90–94. [Google Scholar] [CrossRef] [PubMed]
- Ransohoff, R.M. How neuroinflammation contributes to neurodegeneration. Science 2016, 353, 777. [Google Scholar] [CrossRef] [PubMed]
- Minter, M.R.; Taylor, J.M.; Crack, P.J. The contribution of neuroinflammation to amyloid toxicity in Alzheimer’s disease. J. Neurochem. 2016, 136, 457–474. [Google Scholar] [CrossRef]
- Li, W.; Suwanwela, N.C.; Patumraj, S. Curcumin by down-regulating NF-κB and elevating Nrf2, reduces brain edema and neurological dysfunction after cerebral I/R. Microvasc. Res. 2016, 106, 117–127. [Google Scholar] [CrossRef]
- Yang, T.; Zang, S.; Wang, Y.; Zhu, Y.; Jiang, L.; Chen, X.; Zhang, X.; Cheng, J.; Gao, R.; Xiao, H.; et al. Methamphetamine induced neuroinflammation in mouse brain and microglial cell line BV2: Roles of the TLR4/TRIF/Peli1 signaling axis. Toxicol. Lett. 2020, 333, 150–158. [Google Scholar] [CrossRef]
- Girard, C.A.; Lecacheur, M.; Jouira, R.B.; Berestjuk, I.; Diazzi, S.; Prod’Homme, V.; Mallavialle, A.; Larbret, F.; Gesson, M.; Schaub, S.; et al. A feed-forward mechanosignaling loop confers resistance to therapies targeting the MAPK pathway in BRAF-mutant melanoma. Cancer Res. 2020, 80, 1927–1941. [Google Scholar] [CrossRef]
- Zheng, Y.; Fang, W.; Fan, S.; Liao, W.; Xiong, Y.; Liao, S.; Li, Y.; Xiao, S.; Liu, J. Neurotropin inhibits neuroinflammation via suppressing NF-κB and MAPKs signaling pathways in lipopolysaccharide-stimulated BV2 cells. J. Pharmacol. Sci. 2018, 136, 242–248. [Google Scholar] [CrossRef]
- Li, S.-J.; Qin, W.-X.; Peng, D.-J.; Yuan, Z.-X.; He, S.-N.; Luo, Y.-N.; Aschner, M.; Jiang, Y.-M.; Liang, D.-Y.; Xie, B.-Y.; et al. Sodium P-aminosalicylic acid inhibits sub-chronic manganese-induced neuroinflammation in rats by modulating MAPK and COX-2. Neurotoxicology 2018, 64, 219–229. [Google Scholar] [CrossRef]
- Liu, N.; Wang, H.; Han, G.; Cheng, J.; Hu, W.; Zhang, J. Enhanced proliferation and differentiation of HO-1 gene-modified bone marrow-derived mesenchymal stem cells in the acute injured kidney. Int. J. Mol. Med. 2018, 42, 946–956. [Google Scholar] [CrossRef]
- Lou, J.; Cao, G.; Li, R.; Liu, J.; Dong, Z.; Xu, L. β-caryophyllene attenuates focal cerebral ischemia-reperfusion injury by Nrf2/HO-1 pathway in rats. Neurochem. Res. 2016, 41, 1291–1304. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Huang, L.; Yu, J. Effects of blueberry anthocyanins on retinal oxidative stress and inflammation in diabetes through Nrf2/HO-1 signaling. J. Neuroimmunol. 2016, 301, 1–6. [Google Scholar]
- Park, E.J.; Kim, Y.M.; Kim, H.J.; Chang, K.C. Luteolin activates ERK1/2- and ca2+-dependent HO-1 induction that reduces LPS-induced HMGB1, iNOS/NO, and COX-2 expression in RAW264.7 cells and mitigates acute lung injury of endotoxin mice. Inflamm. Res. 2018, 67, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Aparicio-Soto, M.; Sánchez-Hidalgo, M.; Cárdeno, A.; Rosillo, M.Á.; Sánchez-Fidalgo, S.; Utrilla, J.; Martín-Lacave, I.; Alarcón-de-la-Lastra, C. Dietary extra virgin olive oil attenuates kidney injury in pristane-induced SLE model via activation of HO-1/Nrf-2 antioxidant pathway and suppression of JAK/STAT, NF-κB and MAPK activation. J. Nutr. Biochem. 2016, 27, 278–288. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, H.; Lin, C.; Ren, J.; Zhang, S. Forsythiaside A exhibits anti-inflammatory effects in LPS-stimulated BV2 microglia cells through activation of Nrf2/HO-1 signaling pathway. Neurochem. Res. 2016, 41, 659–665. [Google Scholar] [CrossRef]
- Jeong, Y.H.; Oh, Y.C.; Cho, W.K.; Yim, N.H.; Ma, J.Y. Anti-inflammatory effect of Rhapontici radix ethanol extract via inhibition of NF-κB and MAPK and induction of HO-1 in macrophages. Mediat. Inflamm. 2016, 2016, 7216912. [Google Scholar] [CrossRef]
- Noh, J.; Jung, E.; Lee, J.; Hyun, H.; Hong, S.; Lee, D. Engineered polymeric micelles for combinational oxidation anticancer therapy through concurrent HO-1 inhibition and ROS generation. Biomacromolecules 2019, 20, 1109–1117. [Google Scholar] [CrossRef]
- Quang, T.H.; Ngan, N.T.; Yoon, C.S.; Cho, K.H.; Kang, D.G.; Lee, H.S.; Kim, Y.C.; Oh, H. Protein tyrosine phosphatase 1B inhibitors from the Roots of Cudrania tricuspidata. Molecules 2015, 20, 11173–11183. [Google Scholar] [CrossRef]
- Xin, L.-T.; Yue, S.-J.; Fan, Y.-C.; Wu, J.-S.; Yan, D.; Guan, H.-S.; Wang, C.-Y. Cudrania tricuspidata: An updated review on ethnomedicine, phytochemistry and pharmacology. RSC Adv. 2017, 7, 31807–31832. [Google Scholar] [CrossRef]
- Jeong, J.Y.; Jo, Y.H.; Lee, K.Y.; Do, S.; Hwang, B.Y.; Lee, M.K. Optimization of pancreatic lipase inhibition by Cudrania tricuspidata fruits using response surface methodology. Bioorg. Med. Chem. Lett. 2014, 24, 2329–2333. [Google Scholar] [CrossRef]
- Jee, S.C.; Lee, K.M.; Kim, M.; Lee, Y.J.; Kim, S.; Park, J.O.; Sung, J.S. Neuroprotective effect of Cudrania tricuspidata fruit extracts on scopolamine-induced learning and memory impairment. Int. J. Mol. Sci. 2020, 21, 9202. [Google Scholar] [CrossRef]
- Kim, D.; Cheon, J.; Yoon, H.; Jun, H.S. Cudrania tricuspidata root extract prevents methylglyoxal-induced inflammation and oxidative stress via regulation of the PKC-NOX4 pathway in human kidney cells. Oxid. Med. Cell. Longev. 2021, 2021, 5511881. [Google Scholar] [CrossRef]
- Seo, W.G.; Pae, H.O.; Oh, G.S.; Chai, K.Y.; Yun, Y.G.; Chung, H.T.; Jang, K.K.; Kwon, T.O. Ethyl acetate extract of the stem bark of Cudrania tricuspidata induces apoptosis in human leukemia HL-60 cells. Am. J. Chin. Med. 2001, 29, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Tian, Y.H.; Kim, H.C.; Cui, J.M.; Kim, Y.C. Hepatoprotective constituents of Cudrania tricuspidata. Arch. Pharm. Res. 2005, 28, 44–48. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Lee, S.; Chung, Y.W.; Kim, B.M.; Kim, H.; Kim, K.; Yang, K.M. Antiobesity and antidiabetes effects of a Cudrania tricuspidata hydrophilic extract presenting PTP1B inhibitory potential. BioMed Res. Int. 2016, 2016, 8432759. [Google Scholar]
- Shi, L.; Dong, Q.; Ding, K. Structure elucidation and immunomodulatory activity in vitro of a xylan from roots of Cudrania tricuspidata. Food Chem. 2014, 152, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.; Ha, H.; Lee, J.K.; Seo, C.S.; Lee, N.H.; Jung, D.Y.; Park, S.J.; Shin, H.K. The fruits of Cudrania tricuspidata suppress development of atopic dermatitis in NC/Nga mice. Phytother. Res. 2012, 26, 594–599. [Google Scholar] [CrossRef] [PubMed]
- Ko, W.; Kim, N.; Lee, H.; Woo, E.R.; Kim, Y.C.; Oh, H.; Lee, D.S. Anti-inflammatory effects of compounds from Cudrania tricuspidata in HaCaT Human Keratinocytes. Int. J. Mol. Sci. 2021, 22, 7472. [Google Scholar] [CrossRef] [PubMed]
- Fulop, G.A.; Ahire, C.; Csipo, T.; Tarantini, S.; Kiss, T.; Balasubramanian, P.; Yabluchanskiy, A.; Farkas, E.; Toth, A.; Nyúl-Tóth, Á.; et al. Cerebral venous congestion promotes blood-brain barrier disruption and neuroinflammation, impairing cognitive function in mice. Geroscience 2019, 41, 575–589. [Google Scholar] [CrossRef]
- Wang, X.; Yu, H.; Wang, C.; Liu, Y.; Zhang, G. Chronic ethanol exposure induces neuroinflammation in H4 cells through TLR3/NF-κB pathway and anxiety-like behavior in male C57BL/6 mice. Toxicology 2020, 446, 152625. [Google Scholar] [CrossRef]
- Hung, W.L.; Ho, C.T.; Pan, M.H. Targeting the NLRP3 inflammasome in neuroinflammation: Health promoting effects of dietary phytochemicals in neurological disorders. Mol. Nutr. Food Res. 2020, 64, 1900550. [Google Scholar] [CrossRef]
- Tripathi, A.; Whitehead, C.; Surrao, K.; Pillai, A.; Pillai, A. Type 1 interferon mediates chronic stress-induced neuroinflammation and behavioral deficits via complement component 3-dependent pathway. Mol. Psychiatry 2021, 26, 3043–3059. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Korhonen, E.A.; Merlini, A.; Strauss, J.; Alitalo, K. Angiopoietin-2 blockade ameliorates autoimmune neuroinflammation by inhibiting leukocyte recruitment into the CNS. J. Clin. Investig. 2020, 130, 1977–1990. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Wang, H.; Wang, T.; Luo, C.; Wang, Z.; Zhang, M.; Chen, X.; Tao, L. Platelet regulates neuroinflammation and restores blood–brain barrier integrity in a mouse model of traumatic brain injury. J. Neurochem. 2020, 154, 190–204. [Google Scholar] [CrossRef] [PubMed]
- Liang, B.; Li, H.R.; Xu, L.Z.; Yang, S.L. Xanthones from the roots of Cudrania fruticosa wight. J. Asian Nat. Prod. Res. 2007, 9, 393–397. [Google Scholar] [CrossRef]
- Yang, L.; Zhou, R.; Tong, Y.; Chen, P.; Shen, Y.; Miao, S.; Liu, X. Neuroprotection by dihydrotestosterone in LPS-induced neuroinflammation. Neurobiol. Dis. 2020, 140, 104814. [Google Scholar] [CrossRef]
- Zou, M.; Yang, L.; Niu, L.; Zhao, Y.; Sun, Y.; Fu, Y.; Peng, X. Baicalin ameliorates Mycoplasma gallisepticum -induced lung inflammation in chicken by inhibiting TLR6-mediated NF-κB signalling. Br. Poult. Sci. 2020, 62, 199–210. [Google Scholar] [CrossRef]
- Li, Z.; Wang, C.; Mao, Y.; Cui, J.; Wang, X.; Dang, J.; Wang, S. The expression of STAT3 inhibited the NF-κB signalling pathway and reduced inflammatory responses in mice with viral myocarditis. Int. Immunopharmacol. 2021, 95, 107534. [Google Scholar] [CrossRef]
- Shi, Q.; Cao, J.; Fang, L.; Zhao, H.; Liu, Z.; Ran, J.; Zheng, X.; Li, X.; Zhou, Y.; Ge, D.; et al. Geniposide suppresses LPS-induced nitric oxide, PGE2 and inflammatory cytokine by downregulating NF-κB, MAPK and AP-1 signaling pathways in macrophages. Int. Immunopharmacol. 2014, 20, 298–306. [Google Scholar] [CrossRef]
- Kishimoto, Y.; Niki, H.; Saita, E.; Ibe, S.; Umei, T.; Miura, K.; Ikegami, Y.; Ohmori, R.; Kondo, K.; Momiyama, Y. Blood levels of heme oxygenase-1 versus bilirubin in patients with coronary artery disease. Clin. Chim. Acta. 2020, 504, 30–35. [Google Scholar] [CrossRef]
- Ko, W.; Sohn, J.H.; Jang, J.H.; Ahn, J.S.; Kang, D.G.; Lee, H.S.; Kim, J.S.; Kim, Y.C.; Oh, H. Inhibitory effects of alternaramide on inflammatory mediator expression through TLR4-MyD88-mediated inhibition of NF-κB and MAPK pathway signaling in lipopolysaccharide-stimulated RAW264.7 and BV2 cells. Chem. Biol. Interact. 2016, 244, 16–26. [Google Scholar] [CrossRef] [PubMed]
- Titheradge, M.A. The enzymatic measurement of nitrate and nitrite. Methods Mol. Biol. 1998, 100, 83–91. [Google Scholar] [PubMed]
- Kim, D.C.; Yoon, C.S.; Quang, T.H.; Ko, W.; Kim, J.S.; Oh, H.; Kim, Y.C. Prenylated flavonoids from Cudrania tricuspidata suppress lipopolysaccharide-induced neuroinflammatory activities in BV2 microglial cells. Int. J. Mol. Sci. 2016, 17, 255. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pro-Inflammatory Mediators | Name | RAW264.7 (μM) | BV2 (μM) |
|---|---|---|---|
| NO (IC50) | THMX | 5.77 ± 0.66 | 11.93 ± 2.90 |
| Dexamethasone a | 5.38 ± 1.43 | 7.14 ± 2.16 | |
| PGE2 (IC50) | THMX | 9.70 ± 1.46 | 7.53 ± 1.88 |
| Dexamethasone a | 7.06 ± 0.88 | 4.41 ± 0.59 | |
| IL-6 (IC50) | THMX | 13.34 ± 4.92 | 10.87 ± 3.23 |
| Dexamethasone a | 7.02 ± 0.78 | 8.03 ± 0.65 | |
| TNF-α (IC50) | THMX | 16.14 ± 2.19 | 9.28 ± 0.40 |
| Dexamethasone a | 9.31 ± 0.45 | 8.53 ± 0.93 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ko, W.; Baek, J.-S.; Liu, Z.; Dong, L.; Kim, N.; Lee, H.; Yoon, C.-S.; Kim, N.Y.; Kim, S.C.; Lee, D.-S. Anti-Inflammatory Activity of 1,6,7-Trihydroxy-2-(1,1-dimethyl-2-propenyl)-3-methoxyxanthone Isolated from Cudrania tricuspidata via NF-κB, MAPK, and HO-1 Signaling Pathways in Lipopolysaccharide-Stimulated RAW 264.7 and BV2 Cells. Molecules 2023, 28, 7299. https://doi.org/10.3390/molecules28217299
Ko W, Baek J-S, Liu Z, Dong L, Kim N, Lee H, Yoon C-S, Kim NY, Kim SC, Lee D-S. Anti-Inflammatory Activity of 1,6,7-Trihydroxy-2-(1,1-dimethyl-2-propenyl)-3-methoxyxanthone Isolated from Cudrania tricuspidata via NF-κB, MAPK, and HO-1 Signaling Pathways in Lipopolysaccharide-Stimulated RAW 264.7 and BV2 Cells. Molecules. 2023; 28(21):7299. https://doi.org/10.3390/molecules28217299
Chicago/Turabian StyleKo, Wonmin, Jong-Suep Baek, Zhiming Liu, Linsha Dong, Nayeon Kim, Hwan Lee, Chi-Su Yoon, Na Young Kim, Sam Cheol Kim, and Dong-Sung Lee. 2023. "Anti-Inflammatory Activity of 1,6,7-Trihydroxy-2-(1,1-dimethyl-2-propenyl)-3-methoxyxanthone Isolated from Cudrania tricuspidata via NF-κB, MAPK, and HO-1 Signaling Pathways in Lipopolysaccharide-Stimulated RAW 264.7 and BV2 Cells" Molecules 28, no. 21: 7299. https://doi.org/10.3390/molecules28217299
APA StyleKo, W., Baek, J.-S., Liu, Z., Dong, L., Kim, N., Lee, H., Yoon, C.-S., Kim, N. Y., Kim, S. C., & Lee, D.-S. (2023). Anti-Inflammatory Activity of 1,6,7-Trihydroxy-2-(1,1-dimethyl-2-propenyl)-3-methoxyxanthone Isolated from Cudrania tricuspidata via NF-κB, MAPK, and HO-1 Signaling Pathways in Lipopolysaccharide-Stimulated RAW 264.7 and BV2 Cells. Molecules, 28(21), 7299. https://doi.org/10.3390/molecules28217299