
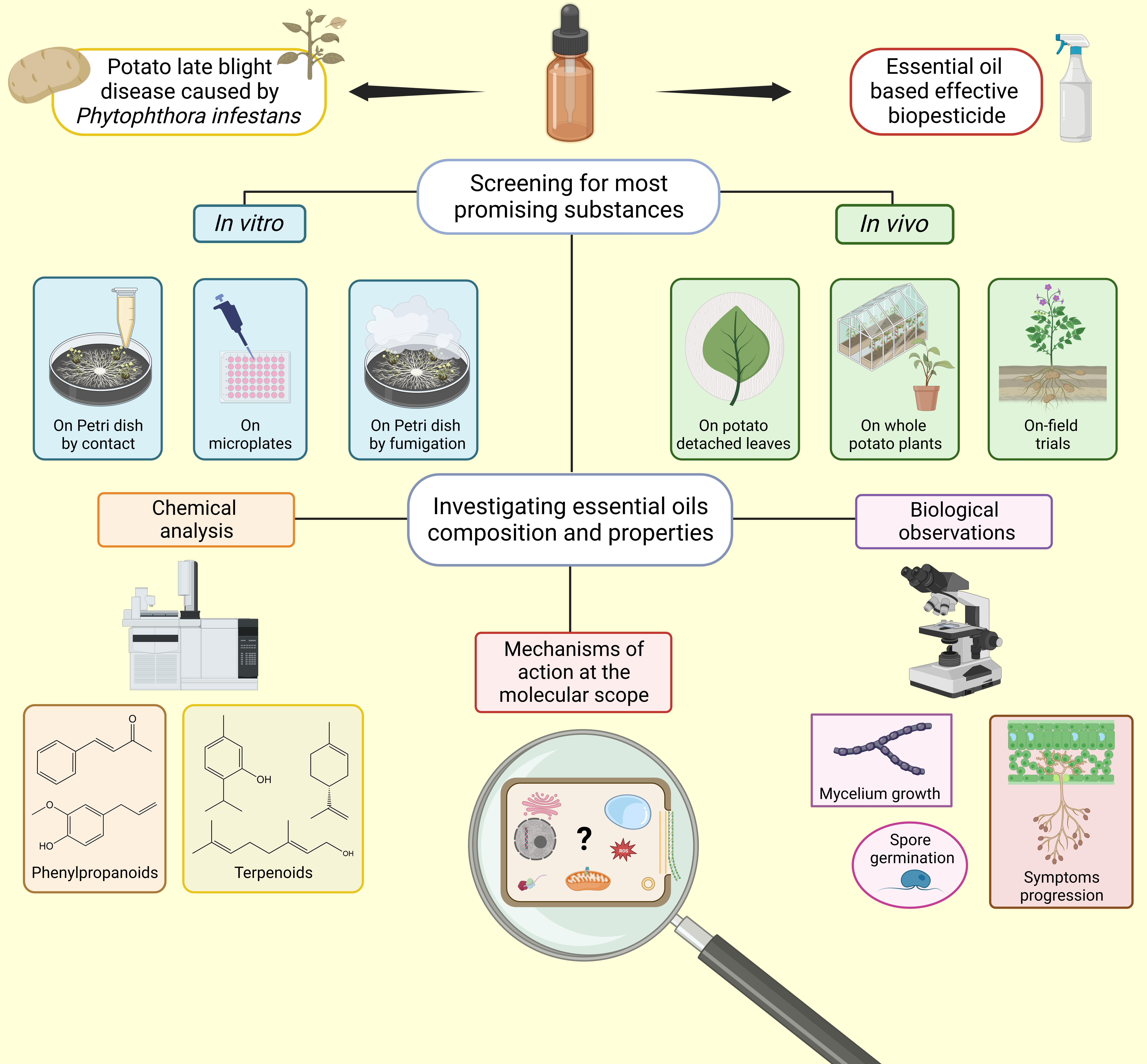
Harnessing Plant’s Arsenal: Essential Oils as Promising Tools for Sustainable Management of Potato Late Blight Disease Caused by Phytophthora infestans—A Comprehensive Review


 ,
,  ,
, Abstract

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Share and Cite
Martini, F.; Jijakli, M.H.; Gontier, E.; Muchembled, J.; Fauconnier, M.-L. Harnessing Plant’s Arsenal: Essential Oils as Promising Tools for Sustainable Management of Potato Late Blight Disease Caused by Phytophthora infestans—A Comprehensive Review. Molecules 2023, 28, 7302. https://doi.org/10.3390/molecules28217302
Martini F, Jijakli MH, Gontier E, Muchembled J, Fauconnier M-L. Harnessing Plant’s Arsenal: Essential Oils as Promising Tools for Sustainable Management of Potato Late Blight Disease Caused by Phytophthora infestans—A Comprehensive Review. Molecules. 2023; 28(21):7302. https://doi.org/10.3390/molecules28217302
Chicago/Turabian StyleMartini, Florian, M. Haïssam Jijakli, Eric Gontier, Jérôme Muchembled, and Marie-Laure Fauconnier. 2023. "Harnessing Plant’s Arsenal: Essential Oils as Promising Tools for Sustainable Management of Potato Late Blight Disease Caused by Phytophthora infestans—A Comprehensive Review" Molecules 28, no. 21: 7302. https://doi.org/10.3390/molecules28217302
APA StyleMartini, F., Jijakli, M. H., Gontier, E., Muchembled, J., & Fauconnier, M.-L. (2023). Harnessing Plant’s Arsenal: Essential Oils as Promising Tools for Sustainable Management of Potato Late Blight Disease Caused by Phytophthora infestans—A Comprehensive Review. Molecules, 28(21), 7302. https://doi.org/10.3390/molecules28217302