

Composition of Polygonatum zanlanscianense Pamp. Steam and Leaf Phenolic Extract and Its Protective Mechanism on t-BHP-Induced Oxidative Damage of HepG2 Cells

Abstract
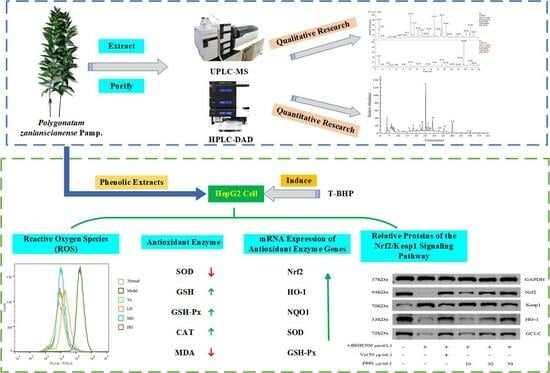
:
1. Introduction
2. Results
2.1. Qualitative and Quantitative Analysis of the Main Phenols in PPP
2.1.1. FTIR Analysis
2.1.2. UPLC-Q-Obtrip-MS Analysis
2.1.3. HPLC Analysis
2.2. The Effect of PPP and t-BHP on the Proliferation of HepG2 Cells
2.3. The Effect of PPP on the Level of Intracellular ROS Induced by t-BHP
2.4. The Effect of PPP on Antioxidant Enzyme Activity and MDA Content
2.5. Effect of PPP on the mRNA Expression of Antioxidant Enzyme Genes Induced by t-BHP
2.6. Western Blotting
3. Materials and Methods
3.1. Polygonatum Zanlanscianense Pamp. Stem and Leaf Samples
3.2. Sample Extraction
3.3. Identification and Quantification of Phenolic Compounds
3.3.1. FTIR
3.3.2. UPLC-Q-Obtrip-MS
3.3.3. HPLC-DAD
3.4. Cell Culture
3.5. Cell Viability Assay
3.6. Intracellular ROS Assay
3.7. Detection of Cellular Antioxidant Enzyme Activity and Malondialdehyde Content
3.8. RNA Extraction and Quantitative Real-Time polymerase Chain Reaction (qRT-PCR)
3.9. Western Blotting
3.10. Statistical Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, D.I.; Griendling, K.K. Regulation of signal transduction by reactive oxygen species in the cardiovascular system. Circ. Res. 2015, 116, 531–549. [Google Scholar] [CrossRef] [PubMed]
- Habtemariam, S. Modulation of reactive oxygen species in health and disease. Antioxidants 2019, 8, 513. [Google Scholar] [CrossRef] [PubMed]
- Nirmala, C.; Bisht, M.S.; Bajwa, H.K.; Santosh, O. Bamboo: A rich source of natural antioxidants and its applications in the food and pharmaceutical industry. Trends Food Sci. Technol. 2018, 77, 91–99. [Google Scholar] [CrossRef]
- Wu, Z.; Shang, X.; Liu, G.; Xie, Y. Comparative analysis of flavonoids, polyphenols and volatiles in roots, stems and leaves of five mangroves. PeerJ 2023, 11, e15529. [Google Scholar] [CrossRef] [PubMed]
- Polat, D.Ç.; Hürkul, M.M. Evaluation of Lonicera etrusca var. etrusca Santi (Caprifoliaceae) Stem and Leaf in Terms of Anatomical Structures and Some Phenolic Compounds. Turk. J. Pharm. Sci. 2022, 19, 636. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Chen, Y.; Xie, W.; Li, X.; Mei, G.; Xu, J.; Zhao, X.; Teng, H.; Yang, G. Phenolic Compounds from the Stems and Leaves of Berchemia lineata (L.) DC. Front. Chem. 2022, 10, 889441. [Google Scholar] [CrossRef] [PubMed]
- Tong, N.N.; Zhou, X.Y.; Peng, L.P.; Liu, Z.A.; Shu, Q.Y. A comprehensive study of three species of Paeonia stem and leaf phytochemicals, and their antioxidant activities. J. Ethnopharmacol. 2021, 273, 113985. [Google Scholar] [CrossRef] [PubMed]
- Goufo, P.; Singh, R.K.; Cortez, I. A reference list of phenolic compounds (including stilbenes) in grapevine (Vitis vinifera L.) roots, woods, canes, stems, and leaves. Antioxidants 2020, 9, 398. [Google Scholar] [CrossRef]
- Govindarajan, V.S.; Nadu, T. Critical reviews in food science and nutrition. Circ. Res. 2022, 62, 3833–3854. [Google Scholar]
- Suzuki, T.; Yamamoto, M. Stress-sensing mechanisms and the physiological roles of the Keap1-Nrf2 system during cellular stress. J. Biol. Chem. 2019, 29241, 16817–16824. [Google Scholar] [CrossRef]
- Niture, S.K.; Khatri, R.; Jaiswal, A.K. Regulation of Nrf2an update. Free. Radic. Biol. Med. 2014, 66, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.K.; Zhang, H.; Zhang, P.F.; Qin, K.; Liu, S.; Peng, D.; Li, L. Prediction of the Potential Distributions and Ecological Suitability of Polygonatum zanlanscianense Pamp. J. Agric. Sci. Technol. 2021, 23, 185–192. [Google Scholar]
- The Chinese Pharmacopoeia; Chinese Medicine Publishing House: Beijing, China, 2020.
- Lee, K.M.; Kwon, T.Y.; Kang, U.; Seo, E.K.; Kim, Y.S. Tussilagonone-induced Nrf2 pathway activation protects Hep G2 cells from oxidative injury. Food Chem. Toxicol. Int. J. Publ. Br. Ind. Biol. Res. Assoc. 2017, 108, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Nurhanani, R.; Sarni, M.J.; Azlina, A.A. Polyphenols from the extract and fraction of T indica seeds protected Hep G2 cells against oxidative stress. BMC Complement. Altern. Med. 2015, 15, 438. [Google Scholar]
- Song, G.; Xia, J.; Xiao, Q. GC-MS Analysis and Antioxidant Activities Determination of Volatile Oil from Leaves of Polygonatum zanlanscianense Pamp. J. Hubei Minzu Univ. (Nat. Sci. Ed.) 2022, 40, 374–379+384. [Google Scholar]
- Niedzielska, E.; Smaga, I.; Gawlik, M.; Moniczewski, A.; Stankowicz, P.; Pera, J.; Filip, M. Oxidative stress in neurodegenerative diseases. Mol. Neurobiol. 2016, 53, 4094–4125. [Google Scholar] [CrossRef] [PubMed]
- Singh, K.S.; Patra, A. Evaluation of phenolic composition, antioxidant, anti-inflammatory and anticancer activities of Polygonatum verticillatum (L.). J. Integr. Med. 2018, 16, 273–282. [Google Scholar] [CrossRef]
- Yonghua, W.; Jianping, Z.H.A.N.G.; Jiechang, Z.H.A.O. Antioxidantion and composition analysis of purified polygonatum sibiricum polyphenols using macroporous resin. Trans. Chin. Soc. Agric. Eng. 2020, 36, 318–326. [Google Scholar]
- He, L.; He, T.; Farrar, S.; Ji, L.; Liu, T.; Ma, X. Antioxidants maintain cellular redox homeostasis by elimination of reactive oxygen species. Cell. Physiol. Biochem. 2017, 44, 532–553. [Google Scholar] [CrossRef]
- Cai, Y.; Li, B.; Peng, D.; Wang, X.; Li, P.; Huang, M.; Chen, J. Crm1-dependent nuclear export of Bach1 is involved in the protective effect of hyperoside on oxidative damage in hepatocytes and CCl4-induced acute liver injury. J. Inflamm. Res. 2021, 14, 551. [Google Scholar] [CrossRef]
- Xiao, L.; Lu, X.; Yang, H.; Lin, C.; Li, L.; Ni, C.; Yan, P. The antioxidant and hypolipidemic effects of mesona chinensis benth extracts. Molecules 2022, 27, 3423. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Gong, X.B.; Huang, L.G.; Wang, Z.X.; Wan, R.Z.; Zhang, P.; Zhang, B.S. Diosmetin exerts anti-oxidative, anti-inflammatory and anti-apoptotic effects to protect against endotoxin-induced acute hepatic failure in mice. Oncotarget 2017, 8, 30723. [Google Scholar] [CrossRef]
- Małgorzata, O.T.; Wianowska, D. Antioxidant Properties of Selected Flavonoids in Binary Mixtures-Considerations on Myricetin, Kaempferol and Quercetin. Int. J. Mol. Sci. 2023, 12, 24. [Google Scholar]
- Song, X.; Liu, C.; Zhang, Y.; Xiao, X.; Han, G.; Sun, K.; Liu, S.; Zhang, Z.; Dong, C.; Li, Y.; et al. Sustainable extraction of ligustilide and ferulic acid from Angelicae Sinensis Radix, for antioxidant and anti-inflammatory activities. Ultrason. Sonochemistry 2023, 94, 106344. [Google Scholar] [CrossRef]
- Shang, N.Y.; Li, X.; Liu, M.Y.; Chen, Q.Y.; Sun, Z.; Lan, J.Q.; Zhang, J.L.; Peng, Y. The protective effect and mechanism of caffeic acid on cerebral ischemic injury. Chin. J. Pharmacol. Toxicol. 2023, 37, 518. [Google Scholar]
- Cicek, B.; Hacimuftuoglu, A.; Yeni, Y.; Danisman, B.; Ozkaraca, M.; Mokhtare, B.; Lazopoulos, G.; Tsarouhas, K.; Tsatsakis, A.; Taghizadehghalehjoughi, A.; et al. Chlorogenic Acid Attenuates Doxorubi-cin-Induced Oxidative Stress and Markers of Apoptosis in Cardiomyocytes via Nrf2/HO-1 and Di-tyrosine Signaling. J. Pers. Med. 2023, 4, 13. [Google Scholar]
- Liu, R.; Li, X.D. Antioxidant and Anti-inflammatory Properties of the Citrus Flavonoids Hesperidin and Hesperetin: An Review of Their Molecular Mechanisms. Chin. J. Med. Guide 2019, 21, 749–752. [Google Scholar]
- Wei, X.; Nan, Z.; Zhang, Z.T. Effects of dietary cyanidin-3-diglucoside-5-glucoside complexes with rutin/Mg(II) against H2O2-induced cellular oxidative stress. Food Res. Int. 2019, 126, 108591. [Google Scholar]
- Liu, L.; Yuan, Y.; Zuo, J.; Tao, J. Composition and antioxidant activity of Paeonia lactiflora petal flavonoid extract and underlying mechanisms of the protective effect on H2O2-induced oxidative damage in BRL3A cells Hortic. Plant J. 2023, 9, 335–344. [Google Scholar] [CrossRef]
- Yi, S.; Zou, L.; Li, Z.; Sakao, K.; Wang, Y.; Hou, D.X. In Vitro Antioxidant Activity of Areca Nut Polyphenol Extracts on RAW264. 7 Cells. Foods 2022, 11, 3607. [Google Scholar] [CrossRef]
- Guo, S.; Szhang, Q. Paeonol protects melanocytes against hydrogen peroxide-induced oxidative stress through activation of Nrf2 signaling pathwa. Drug Dev. Res. 2021, 826, 861–869. [Google Scholar] [CrossRef]
- Ke, Z.; Zhao, Y.; Tan, S.; Chen, H.; Li, Y.; Zhou, Z.; Huang, C. Citrus reticulata Blanco peel extract ameliorates hepatic steatosis, oxidative stress and inflammation in HF and MCD diet-induced NASH C57BL/6 J mice. J. Nutr. Biochem. 2020, 83, 108426. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Rt/Min | Compounds | Molecular Formula | Additive Ion | Exact Mass | Theoretical Mass | ppm | Characteristic Fragmentation Ions |
|---|---|---|---|---|---|---|---|---|
| 1 | 0.87 | Caffeic acid | C9H8O4 | [M-H]− | 180.0352 | 180.0350 | 1.11 | 135.0443 |
| 2 | 6.89 | Chlorogenic acid | C16H18O9 | [M-H]− | 354.0874 | 354.1708 | 1.13 | 191.0191 |
| 3 | 8.45 | Kaempferol | C15H10O6 | [M+H]+ | 286.0408 | 286.0405 | 1.05 | 153.0181– 121.0284 |
| 4 | 9.13 | Ferulic acid | C10H10O4 | [M-H]− | 194.0504 | 194.0504 | −1.03 | 178.0548– 134.9392 |
| 5 | 9.39 | Taxifolin | C15H12O7 | [M-H]− | 304.0512 | 304.051 | 0.66 | 285.0406– 151.0393– 107.0235 |
| 6 | 9.64 | Kaempferol-3-O-rutinoside | C27H30O15 | [M-H]− | 594.1518 | 594.1511 | 1.18 | 285.0326 |
| 7 | 10.15 | Fisetin | C15H10O6 | [M+H]+ | 286.0555 | 286.055 | 1.75 | 213.0544– 137.0230– 121.0287 |
| 8 | 10.16 | Hesperidin | C28H34O15 | [M-H]− | 610.1825 | 610.1823 | −0.33 | 489.1451– 301.0275 |
| 9 | 10.45 | Hyperin | C21H20O12 | [M-H]− | 464.0887 | 464.0881 | 1.29 | 301.0275– 271.0248– 179.0055– 151.0344 |
| 10 | 10.08 | Isoquercitrin | C21H20O12 | [M-H]− | 464.0882 | 464.0882 | 0 | 300.0275– 271.0248– 255.0298 |
| 11 | 10.98 | Rutin | C27H30O16 | [M-H]− | 610.1467 | 610.1461 | 0.98 | 300.0277– 271.0247– 255.0299 |
| 12 | 12.19 | Astragalin | C21H20O11 | [M-H]− | 448.0933 | 448.0930 | −0.67 | 285.0408– 151.0029– 107.0125 |
| 13 | 12.30 | Epicatechin | C15H14O6 | [M-H]− | 290.0714 | 290.0718 | −1.38 | 245.0816– 203.0722– 123.0465 |
| 14 | 13.37 | Quercetin | C15H10O7 | [M-H]− | 302.0359 | 302.0354 | 1.66 | 151.0024 |
| 15 | 13.84 | Diosmetin | C16H12O6 | [M-H]− | 300.0567 | 300.0561 | 2.00 | 284.0326– 151.0029 |
| 16 | 15.02 | Luteolin | C15H10O6 | [M-H]− | 286.0407 | 286.0404 | 1.05 | 257.0444– 151.0029– 133.0287 |
| 17 | 15.43 | 3-Methoxy-5,7,3′,4′-tetrahydroxy-flavone | C16H12O7 | [M-H]− | 316.0512 | 316.051 | 0.63 | 300.0276– 283.0535– 283.0535– |
| 18 | 18.8 | Farrerol | C17H16O5 | [M-H]− | 299.0929 | 299.0925 | 1.33 | 271.1834– 179.0342 |
| 19 | 19.16 | Dalspinosin | C18H16O7 | [M+H]+ | 345.0962 | 345.0969 | −2.03 | 316.1031– 179.0423 |
| 20 | 25.24 | Gardenin B | C19H18O7 | [M-H]− | 360.1129 | 360.1125 | 1.11 | 344.0543– 329.0305– 326.0883– 298.0556 |
| Peak | Compounds | Regression Equation | R2 | Content (mg/g) |
|---|---|---|---|---|
| 5 | hyperin | y = 410.1x + 0.185 | 0.9999 | 76.6 |
| 9 | astragalin | y = 633.29x + 0.1012 | 0.9999 | 21.4 |
| 12 | diosmetin | y = 507.27x − 0.0038 | 0.9999 | 26.4 |
| Genes | Primer Sequences |
|---|---|
| GAPDH | forward: GGAGCGAGATCCCTCCAAAA reverse: GGCTGTTGTCATACTTCTCATGG |
| Nrf2 | forward: AGTCCAGAAGCCAAACTGACAGAAG reverse: GGAGAGGATGCTGCTGAAGGAATC |
| HO-1 | forward: TGCCAGTGCCACCAAGTTCAAG reverse: TGTTGAGCAGGAACGCAGTCTTG |
| NQO1 | forward: GGATGGGAGGTGGTGGAGTCG reverse: AATATCACAAGGTCTGCGGCTTCC |
| SOD | forward: AGCAGATGACTTGGGCAAAGGTG reverse: ACCACAAGCCAAACGACTTCCAG |
| GSH-Px | forward: GCAACCAGTTTGGGCATCAGGAG reverse: GCACCGTTCACCTCGCACTTC |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, S.; Yong, J.; Yan, J.; Peng, T.; Long, F.; Chen, H. Composition of Polygonatum zanlanscianense Pamp. Steam and Leaf Phenolic Extract and Its Protective Mechanism on t-BHP-Induced Oxidative Damage of HepG2 Cells. Molecules 2023, 28, 7487. https://doi.org/10.3390/molecules28227487
Tang S, Yong J, Yan J, Peng T, Long F, Chen H. Composition of Polygonatum zanlanscianense Pamp. Steam and Leaf Phenolic Extract and Its Protective Mechanism on t-BHP-Induced Oxidative Damage of HepG2 Cells. Molecules. 2023; 28(22):7487. https://doi.org/10.3390/molecules28227487
Chicago/Turabian StyleTang, Shuang, Jin Yong, Jin Yan, Teng Peng, Fei Long, and Hulan Chen. 2023. "Composition of Polygonatum zanlanscianense Pamp. Steam and Leaf Phenolic Extract and Its Protective Mechanism on t-BHP-Induced Oxidative Damage of HepG2 Cells" Molecules 28, no. 22: 7487. https://doi.org/10.3390/molecules28227487