


A Natural Compound Containing a Disaccharide Structure of Glucose and Rhamnose Identified as Potential N-Glycanase 1 (NGLY1) Inhibitors
Abstract
:
1. Introduction
2. Results
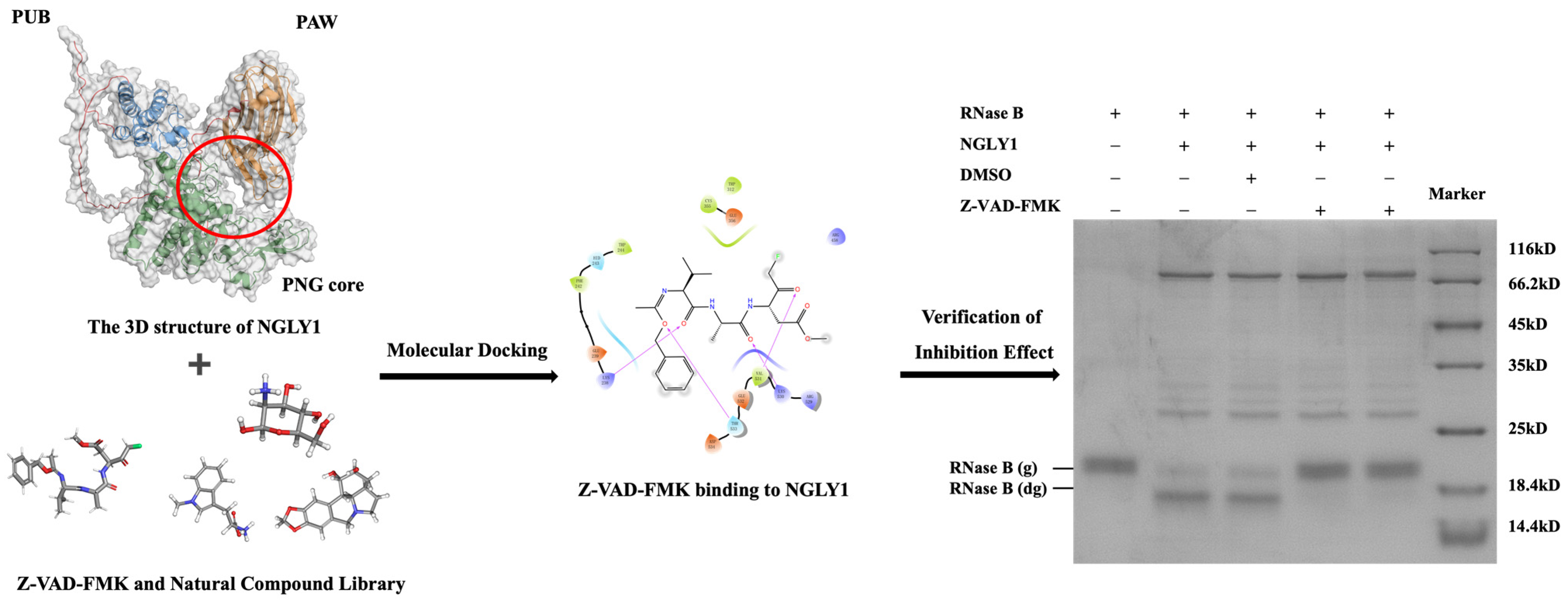
2.1. Establishment of a Screening Method for Targeting NGLY1 Lead Compounds
2.2. Verification of Inhibitory Effect of Primary Screening Compounds
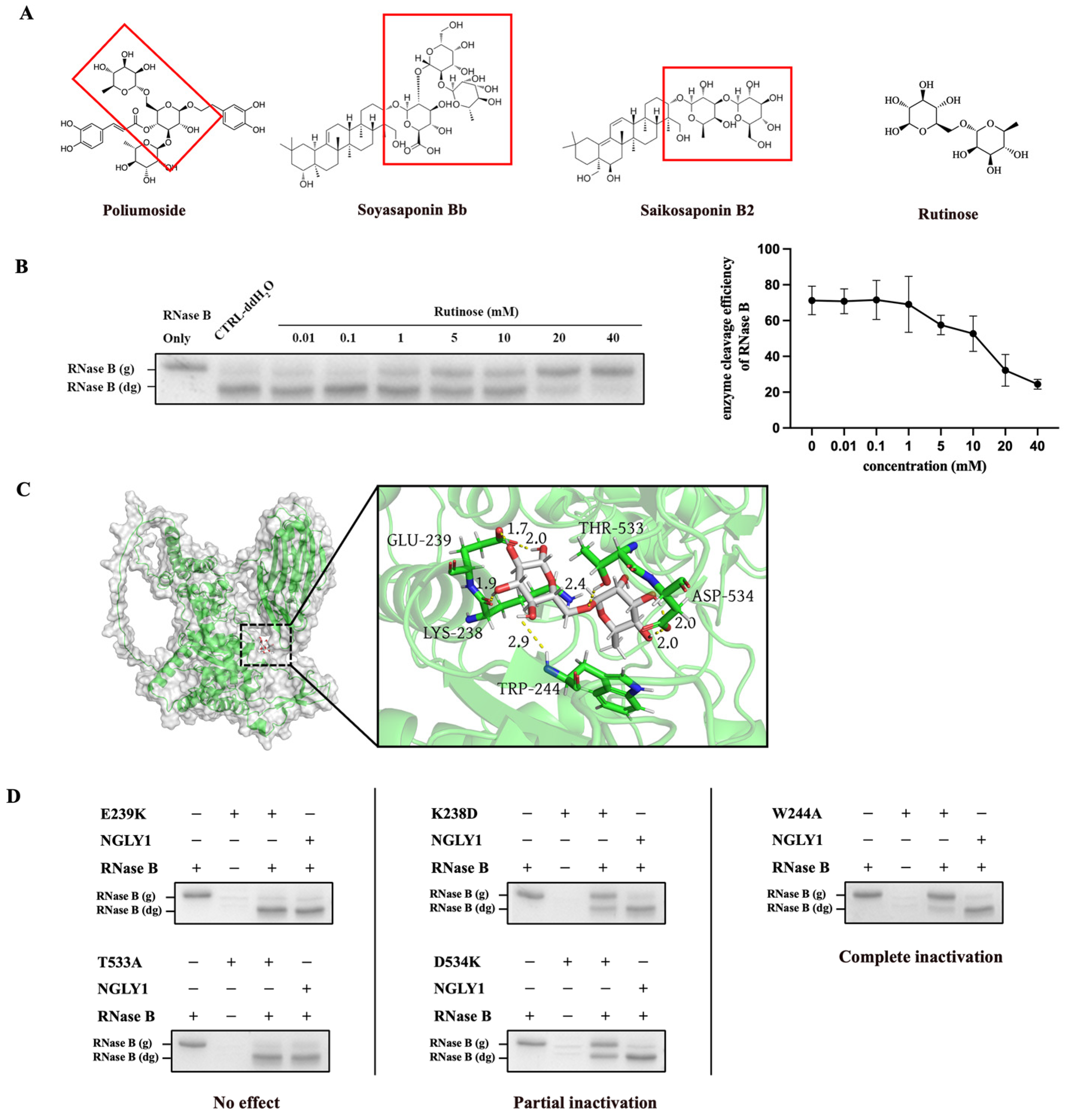
2.3. The Analysis of Inhibitory Mechanism of NGLY1 Inhibitor
2.4. The Inhibitory Effects of Rutinose as the Core Structural Element for NGLY1 Inhibitor
2.5. Toxicity Analysis
3. Discussion
4. Materials and Methods
4.1. Molecular Docking
4.2. Construction of NGLY1 Mutant
4.3. Protein Expression and Purification of NGLY1 and NGLY1 Mutant
4.4. The Electrophoretic Mobility Shift Assay
4.5. Toxicity Test
4.6. CCK-8 Analysis of Cell Viability Assay
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Srinivasan, B.; Zhou, H.; Mitra, S.; Skolnick, J. Novel small molecule binders of human N-glycanase 1, a key player in the endoplasmic reticulum associated degradation pathway. Bioorg. Med. Chem. 2016, 24, 4750–4758. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Huang, C.; Fujihira, H. The cytoplasmic peptide: N-glycanase (NGLY1)—Structure, expression and cellular functions. Gene 2016, 577, 1–7. [Google Scholar] [CrossRef]
- Tomlin, F.M.; Gerling-Driessen, U.I.M.; Liu, Y.C.; Flynn, R.A.; Vangala, J.R.; Lentz, C.S.; Clauder-Muenster, S.; Jakob, P.; Mueller, W.F.; Ordoñez-Rueda, D.; et al. Inhibition of NGLY1 Inactivates the Transcription Factor Nrf1 and Potentiates Proteasome Inhibitor Cytotoxicity. ACS Cent. Sci. 2017, 3, 1143–1155. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Raihan, M.A.; Reynoso, F.J.; He, M. Glycosylation Analysis for Congenital Disorders of Glycosylation. Curr. Protoc. Hum. Genet. 2015, 86, 17.18.1–17.18.22. [Google Scholar] [CrossRef] [PubMed]
- Asahina, M.; Fujinawa, R.; Nakamura, S.; Yokoyama, K.; Tozawa, R.; Suzuki, T. Ngly1−/− rats develop neurodegenerative phenotypes and pathological abnormalities in their peripheral and central nervous systems. Hum. Mol. Genet. 2020, 29, 1635–1647. [Google Scholar] [CrossRef] [PubMed]
- Might, M.; Wilsey, M. The shifting model in clinical diagnostics: How next-generation sequencing and families are altering the way rare diseases are discovered, studied, and treated. Genet. Med. 2014, 16, 736–737. [Google Scholar] [CrossRef] [PubMed]
- Abuduxikuer, K.; Zou, L.; Wang, L.; Chen, L.; Wang, J.S. Novel NGLY1 gene variants in Chinese children with global developmental delay, microcephaly, hypotonia, hypertransaminasemia, alacrimia, and feeding difficulty. J. Hum. Genet. 2020, 65, 387–396. [Google Scholar] [CrossRef]
- Kariminejad, A.; Shakiba, M.; Shams, M.; Namiranian, P.; Eghbali, M.; Talebi, S.; Makvand, M.; Jaeken, J.; Najmabadi, H.; Hennekam, R.C. NGLY1 deficiency: Novel variants and literature review. Eur. J. Med. Genet. 2021, 64, 104146. [Google Scholar] [CrossRef]
- Lam, C.; Wolfe, L.; Need, A.; Shashi, V.; Enns, G. NGLY1-Related Congenital Disorder of Deglycosylation. In GeneReviews®; Adam, M.P., Mirzaa, G.M., Pagon, R.A., Wallace, S.E., Bean, L.J.H., Gripp, K.W., Amemiya, A., Eds.; University of Washington: Seattle, WA, USA, 2018. [Google Scholar]
- Yang, K.; Huang, R.; Fujihira, H.; Suzuki, T.; Yan, N. N-glycanase NGLY1 regulates mitochondrial homeostasis and inflammation through NRF1. J. Exp. Med. 2018, 215, 2600–2616. [Google Scholar] [CrossRef]
- Wu, K.X.; Phuektes, P.; Kumar, P.; Goh, G.Y.; Moreau, D.; Chow, V.T.; Bard, F.; Chu, J.J. Human genome-wide RNAi screen reveals host factors required for enterovirus 71 replication. Nat. Commun. 2016, 7, 13150. [Google Scholar] [CrossRef]
- Misaghi, S.; Pacold, M.E.; Blom, D.; Ploegh, H.L.; Korbel, G.A. Using a small molecule inhibitor of peptide: N-glycanase to probe its role in glycoprotein turnover. Chem. Biol. 2004, 11, 1677–1687. [Google Scholar] [CrossRef] [PubMed]
- Needs, S.H.; Bootman, M.D.; Grotzke, J.E.; Kramer, H.B.; Allman, S.A. Off-target inhibition of NGLY1 by the polycaspase inhibitor Z-VAD-fmk induces cellular autophagy. FEBS J. 2022, 289, 3115–3131. [Google Scholar] [CrossRef] [PubMed]
- Bhambhani, S.; Kondhare, K.R.; Giri, A.P. Diversity in Chemical Structures and Biological Properties of Plant Alkaloids. Molecules 2021, 26, 3374. [Google Scholar] [CrossRef] [PubMed]
- Dubbu, S.; Chennaiah, A.; Verma, A.K.; Vankar, Y.D. Stereoselective synthesis of 2-deoxy-β-C-aryl/alkyl glycosides using Prins cyclization: Application in the synthesis of C-disaccharides and differently protected C-aryl glycosides. Carbohydr. Res. 2018, 468, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Chennaiah, A.; Bhowmick, S.; Vankar, Y.D. Conversion of glycals into vicinal-1,2-diazides and 1,2-(or 2,1)-azidoacetates using hypervalent iodine reagents and Me 3 SiN 3. Application in the synthesis of N-glycopeptides, pseudo-trisaccharides and an iminosugar. RSC Adv. 2017, 7, 41755–41762. [Google Scholar] [CrossRef]
- Yamasaki, S.; Asakura, M.; Neogi, S.B.; Hinenoya, A.; Iwaoka, E.; Aoki, S. Inhibition of virulence potential of Vibrio cholerae by natural compounds. Indian J. Med. Res. 2011, 133, 232–239. [Google Scholar]
- Zhang, L.; Song, J.; Kong, L.; Yuan, T.; Li, W.; Zhang, W.; Hou, B.; Lu, Y.; Du, G. The strategies and techniques of drug discovery from natural products. Pharmacol. Ther. 2020, 216, 107686. [Google Scholar] [CrossRef]
- Guo, X.; Yin, X.; Liu, Z.; Wang, J. Non-Alcoholic Fatty Liver Disease (NAFLD) Pathogenesis and Natural Products for Prevention and Treatment. Int. J. Mol. Sci. 2022, 23, 15489. [Google Scholar] [CrossRef]
- Ma, N.; Zhang, Z.; Liao, F.; Jiang, T.; Tu, Y. The birth of artemisinin. Pharmacol. Ther. 2020, 216, 107658. [Google Scholar] [CrossRef]
- Yu, D.L.; Lou, Z.P.; Ma, F.Y.; Najafi, M. The interactions of paclitaxel with tumour microenvironment. Int. Immunopharmacol. 2022, 105, 108555. [Google Scholar] [CrossRef]
- Yang, H.; Landis-Piwowar, K.R.; Chen, D.; Milacic, V.; Dou, Q.P. Natural compounds with proteasome inhibitory activity for cancer prevention and treatment. Curr. Protein Pept. Sci. 2008, 9, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Choubdar, N.; Bhat, R.G.; Stubbs, K.A.; Yuzwa, S.; Pinto, B.M. Synthesis of 2-amido, 2-amino, and 2-azido derivatives of the nitrogen analogue of the naturally occurring glycosidase inhibitor salacinol and their inhibitory activities against O-GlcNAcase and NagZ enzymes. Carbohydr. Res. 2008, 343, 1766–1777. [Google Scholar] [CrossRef] [PubMed]
- Zhen, D.; Xuan, T.Q.; Hu, B.; Bai, X.; Fu, D.N.; Wang, Y.; Wu, Y.; Yang, J.; Ma, Q. Pteryxin attenuates LPS-induced inflammatory responses and inhibits NLRP3 inflammasome activation in RAW264.7 cells. J. Ethnopharmacol. 2022, 284, 114753. [Google Scholar] [CrossRef] [PubMed]
- Badshah, S.L.; Faisal, S.; Muhammad, A.; Poulson, B.G.; Emwas, A.H.; Jaremko, M. Antiviral activities of flavonoids. Biomed. Pharmacother. 2021, 140, 111596. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, A.; Kaleem, M.; Ahmed, Z.; Shafiq, H. Therapeutic potential of flavonoids and their mechanism of action against microbial and viral infections—A review. Food Res. Int. 2015, 77, 221–235. [Google Scholar] [CrossRef]
- Śledź, P.; Caflisch, A. Protein structure-based drug design: From docking to molecular dynamics. Curr. Opin. Struct. Biol. 2018, 48, 93–102. [Google Scholar] [CrossRef]
- Ballante, F.; Kooistra, A.J.; Kampen, S.; de Graaf, C.; Carlsson, J. Structure-Based Virtual Screening for Ligands of G Protein-Coupled Receptors: What Can Molecular Docking Do for You? Pharmacol. Rev. 2021, 73, 527–565. [Google Scholar] [CrossRef]
- Kitchen, D.B.; Decornez, H.; Furr, J.R.; Bajorath, J. Docking and scoring in virtual screening for drug discovery: Methods and applications. Nat. Rev. Drug Discov. 2004, 3, 935–949. [Google Scholar] [CrossRef]
- Zhang, L.; Xu, J.; Guo, J.; Wang, Y.; Wang, Q. Elucidation of Pharmacological Mechanism Underlying the Anti-Alzheimer’s Disease Effects of Evodia rutaecarpa and Discovery of Novel Lead Molecules: An In Silico Study. Molecules 2023, 28, 5846. [Google Scholar] [CrossRef]
- Lu, F.; Luo, G.; Qiao, L.; Jiang, L.; Li, G.; Zhang, Y. Virtual Screening for Potential Allosteric Inhibitors of Cyclin-Dependent Kinase 2 from Traditional Chinese Medicine. Molecules 2016, 21, 1259. [Google Scholar] [CrossRef]
- Luo, L.; Zhong, A.; Wang, Q.; Zheng, T. Structure-Based Pharmacophore Modeling, Virtual Screening, Molecular Docking, ADMET, and Molecular Dynamics (MD) Simulation of Potential Inhibitors of PD-L1 from the Library of Marine Natural Products. Mar. Drugs 2021, 20, 29. [Google Scholar] [CrossRef] [PubMed]
- Xiong, G.; Wu, Z.; Yi, J.; Fu, L.; Yang, Z.; Hsieh, C.; Yin, M.; Zeng, X.; Wu, C.; Lu, A.; et al. ADMETlab 2.0: An integrated online platform for accurate and comprehensive predictions of ADMET properties. Nucleic Acids Res. 2021, 49, W5–W14. [Google Scholar] [CrossRef]
- Alkahtani, H.M.; Almehizia, A.A.; Al-Omar, M.A.; Obaidullah, A.J.; Zen, A.A.; Hassan, A.S.; Aboulthana, W.M. In Vitro Evaluation and Bioinformatics Analysis of Schiff Bases Bearing Pyrazole Scaffold as Bioactive Agents: Antioxidant, Anti-Diabetic, Anti-Alzheimer, and Anti-Arthritic. Molecules 2023, 28, 7125. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.Q.; Chen, H.; Chen, L.; Tang, D.D.; Miao, H.; Zhao, Y.Y. Metabolomic application in toxicity evaluation and toxicological biomarker identification of natural product. Chem. Biol. Interact. 2016, 252, 114–130. [Google Scholar] [CrossRef] [PubMed]
- Rendedula, D.; Doddipalla, R.; Shaik, H.; Gupta, H.P.; Godugu, D.; Kristipati, R.R.; Mudiam, M.K.R. Multiplatform Metabolomics to Understand the Imidacloprid-Induced Toxicity in Drosophila. Chem. Res. Toxicol. 2023, 36, 1574–1583. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Chen, B.; Cai, Z. Metabolomics-based approach for assessing the toxicity mechanisms of dibutyl phthalate to abalone (Haliotis diversicolor supertexta). Environ. Sci. Pollut. Res. Int. 2015, 22, 5092–5099. [Google Scholar] [CrossRef]
- Decout, A.; Katz, J.D.; Venkatraman, S.; Ablasser, A. The cGAS-STING pathway as a therapeutic target in inflammatory diseases. Nat. Rev. Immunol. 2021, 21, 548–569. [Google Scholar] [CrossRef] [PubMed]
- Manasanch, E.E.; Orlowski, R.Z. Proteasome inhibitors in cancer therapy. Nat. Rev. Clin. Oncol. 2017, 14, 417–433. [Google Scholar] [CrossRef]
- Dou, Q.P.; Zonder, J.A. Overview of proteasome inhibitor-based anti-cancer therapies: Perspective on bortezomib and second generation proteasome inhibitors versus future generation inhibitors of ubiquitin-proteasome system. Curr. Cancer Drug Targets 2014, 14, 517–536. [Google Scholar] [CrossRef]
- Radhakrishnan, S.K.; Lee, C.S.; Young, P.; Beskow, A.; Chan, J.Y.; Deshaies, R.J. Transcription factor Nrf1 mediates the proteasome recovery pathway after proteasome inhibition in mammalian cells. Mol. Cell 2010, 38, 17–28. [Google Scholar] [CrossRef]
- Radhakrishnan, S.K.; den Besten, W.; Deshaies, R.J. p97-dependent retrotranslocation and proteolytic processing govern formation of active Nrf1 upon proteasome inhibition. eLife 2014, 3, e01856. [Google Scholar] [CrossRef]
- Suzuki, T.; Yoshida, Y. Ever-expanding NGLY1 biology. J. Biochem. 2022, 171, 141–143. [Google Scholar] [CrossRef] [PubMed]
- Uhlén, M.; Björling, E.; Agaton, C.; Szigyarto, C.A.; Amini, B.; Andersen, E.; Andersson, A.C.; Angelidou, P.; Asplund, A.; Asplund, C.; et al. A human protein atlas for normal and cancer tissues based on antibody proteomics. Mol. Cell. Proteom. 2005, 4, 1920–1932. [Google Scholar] [CrossRef]
- Zolekar, A.; Lin, V.J.T.; Mishra, N.M.; Ho, Y.Y.; Hayatshahi, H.S.; Parab, A.; Sampat, R.; Liao, X.; Hoffmann, P.; Liu, J.; et al. Stress and interferon signalling-mediated apoptosis contributes to pleiotropic anticancer responses induced by targeting NGLY1. Br. J. Cancer 2018, 119, 1538–1551. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.Y.; Lee, I.S.; Jung, S.H.; Lee, Y.M.; Lee, Y.R.; Kim, J.H.; Sun, H.; Kim, J.S. Caffeoylated phenylpropanoid glycosides from Brandisia hancei inhibit advanced glycation end product formation and aldose reductase in vitro and vessel dilation in larval zebrafish in vivo. Planta Med. 2013, 79, 1705–1709. [Google Scholar] [CrossRef]
- Zhu, L.; Fu, R.; Liu, X.; He, Y.; Wang, B.; Ma, T. Soyasaponin Bb Protects Rat Hepatocytes from Alcohol-Induced Oxidative Stress by Inducing Heme Oxygenase-1. Pharmacogn. Mag. 2016, 12, 302–306. [Google Scholar]
- Jiang, K.; Lu, Q.; Li, Q.; Ji, Y.; Chen, W.; Xue, X. Astragaloside IV inhibits breast cancer cell invasion by suppressing Vav3 mediated Rac1/MAPK signaling. Int. Immunopharmacol. 2017, 42, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Zhu, J.; Xu, Y.W.; Mou, F.F.; Shan, X.L.; Wang, Q.L.; Liu, B.N.; Ning, K.; Liu, J.J.; Wang, Y.C.; et al. Notoginsenoside R1-loaded mesoporous silica nanoparticles targeting the site of injury through inflammatory cells improves heart repair after myocardial infarction. Redox Biol. 2022, 54, 102384. [Google Scholar] [CrossRef]
- Wu, L.; Li, Q.; Liu, Y. Polyphyllin D induces apoptosis in K562/A02 cells through G2/M phase arrest. J. Pharm. Pharmacol. 2014, 66, 713–721. [Google Scholar] [CrossRef]
- Li, S.; Li, X.; He, F.; Jiao, R.; Zhang, S.; Li, Z. Amarogentin promotes osteoblast differentiation in oestrogen-deficiency-induced osteoporosis rats by modulating the Nrf-2/MAPK/ERK signalling pathway. Arch. Med. Sci. 2023, 19, 452–457. [Google Scholar] [CrossRef]
- Siraj, F.M.; SathishKumar, N.; Kim, Y.J.; Kim, S.Y.; Yang, D.C. Ginsenoside F2 possesses anti-obesity activity via binding with PPARγ and inhibiting adipocyte differentiation in the 3T3-L1 cell line. J. Enzym. Inhib. Med. Chem. 2015, 30, 9–14. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Shen, H.; Li, J.; Gong, Y.; Bao, H.; Zhang, J.; Hu, L.; Wang, Z.; Gong, J. Loganin inhibits macrophage M1 polarization and modulates sirt1/NF-κB signaling pathway to attenuate ulcerative colitis. Bioengineered 2020, 11, 628–639. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Deng, C.; Pan, G.; Wang, X.; Zhang, K.; Dong, Z.; Zhao, G.; Tan, M.; Hu, X.; Shi, S.; et al. Lycorine hydrochloride inhibits cell proliferation and induces apoptosis through promoting FBXW7-MCL1 axis in gastric cancer. J. Exp. Clin. Cancer Res. 2020, 39, 230. [Google Scholar] [CrossRef]
- Lin, L.T.; Chung, C.Y.; Hsu, W.C.; Chang, S.P.; Hung, T.C.; Shields, J.; Russell, R.S.; Lin, C.C.; Li, C.F.; Yen, M.H.; et al. Saikosaponin b2 is a naturally occurring terpenoid that efficiently inhibits hepatitis C virus entry. J. Hepatol. 2015, 62, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Xu, X. Advanced research on anti-tumor effects of amygdalin. J. Cancer Res. Ther. 2014, 10 (Suppl. S1), 3–7. [Google Scholar]
- Matsuzaki, Y.; Koyama, M.; Hitomi, T.; Yokota, T.; Kawanaka, M.; Nishikawa, A.; Germain, D.; Sakai, T. Arctiin induces cell growth inhibition through the down-regulation of cyclin D1 expression. Oncol. Rep. 2008, 19, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.A.; Hwang, Y.H.; Yang, H.; Ryuk, J.A.; Gu, D.R.; Ha, H. Ethanolic extract of Pyrrosia lingua (Thunb.) Farw. ameliorates OVX-induced bone loss and RANKL-induced osteoclastogenesis. Biomed. Pharmacother. 2022, 147, 112640. [Google Scholar] [CrossRef]
- Manciu, F.S.; Guerrero, J.; Bennet, K.E.; Chang, S.Y.; Rahman, M.; Martinez Lopez, L.V.; Chantigian, S.; Castellanos, M.; Manciu, M. Assessing Nordihydroguaiaretic Acid Therapeutic Effect for Glioblastoma Multiforme. Sensors 2022, 22, 2643. [Google Scholar] [CrossRef]
- Yamashita, N.; Taga, C.; Ozawa, M.; Kanno, Y.; Sanada, N.; Kizu, R. Camalexin, an indole phytoalexin, inhibits cell proliferation, migration, and mammosphere formation in breast cancer cells via the aryl hydrocarbon receptor. J. Nat. Med. 2022, 76, 110–118. [Google Scholar] [CrossRef]
- Tan, C.; Zhu, W.; Lu, Y. Aloin, cinnamic acid and sophorcarpidine are potent inhibitors of tyrosinase. Chin. Med. J. 2002, 115, 1859–1862. [Google Scholar]
- Cao, B.; Zhao, R.Y.; Li, H.H.; Xu, X.M.; Cui, H.; Deng, H.; Chen, L.; Wei, B. Oral administration of asparagine and 3-indolepropionic acid prolongs survival time of rats with traumatic colon injury. Mil. Med. Res. 2022, 9, 37. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Docking Score | Molecular Formula | Weight (g/mol) | Noncovalent Interactions | Amino Acid Sites | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Z-VAD-FMK | −4.560 | C22H30FN3O7 | 467.494 | 4 H–bond | Lys238 | / | / | / | / | Lys530 | Val531 | Thr533 | |
| Poliumoside | −10.088 | C35H46O19 | 770.734 | 1 Pi–pi 6 H–bond | Lys238 | Glu239 | Trp244 | / | Trp369 | / | Val531 | / | Glu544 |
| Soyasaponin Bb | −8.497 | C48H78O18 | 943.134 | 3 H–bond | Lys238 | / | Trp244 | Glu356 | / | / | / | / | / |
| Saikosaponin B2 | −6.007 | C42H68O13 | 780.993 | 5 H–bond | Lys238 | Glu239 | / | Glu356 | / | / | / | / | / |
| Target | H-HT | DILI | Skin Sensitization | Carcinogenicity | Eye Corrosion | Eye Irritation |
|---|---|---|---|---|---|---|
| Z-VAD-FMK | 0.377 * (−) | 0.924 (+++) | 0.401 (−) | 0.238 (−−) | 0.004 (−−−) | 0.01 (−−−) |
| Poliumoside | 0.375 (−) | 0.953 (+++) | 0.971 (+++) | 0.031 (−−−) | 0.003 (−−−) | 0.311 (−) |
| Soyasaponin Bb | 0.178 (−−) | 0.044 (−−−) | 0.2 (−−) | 0.025 (−−−) | 0.003 (−−−) | 0.008 (−−−) |
| Saikosaponin B2 | 0.281 (−−) | 0.008 (−−−) | 0.053 (−−−) | 0.024 (−−−) | 0.003 (−−−) | 0.006 (−−−) |
| Mutant | Primer 5′-3′ | |
|---|---|---|
| K238D | Forward | CACCCAGTGAAAAAATTCTTCGTCAAACCAGTGCAAAAGCTCCAG |
| Reverse | CTGGAGCTTTTGCACTGGTTTGACGAAGAATTTTTTCACTGGGTG | |
| E239K | Forward | TCACCCAGTGAAAAAATTCTTTCTTAAACCAGTGCAAAAGCTC |
| Reverse | GAGCTTTTGCACTGGTTTAAGAAAGAATTTTTTCACTGGGTGA | |
| W244A | Forward | TGCTGCACAAAACGTTATTCACCGCGTGAAAAAATTCTTCCTTAAACC |
| Reverse | GGTTTAAGGAAGAATTTTTTCACGCGGTGAATAACGTTTTGTGCAGCA | |
| T533A | Forward | CCATGTGCCAGTCTGCTTCAACTTTTCTGAATATAGATTCCATT |
| Reverse | AATGGAATCTATATTCAGAAAAGTTGAAGCAGACTGGCACATGG | |
| D534K | Forward | CAAATATACCATGTGCCACTTTGTTTCAACTTTTCTGAATATAGATTCCATTTTCC |
| Reverse | GGAAAATGGAATCTATATTCAGAAAAGTTGAAACAAAGTGGCACATGGTATATTTG | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, R.; Gu, J.; Ye, Y.; Zhang, Y.; Zhang, S.; Lin, Q.; Yuan, S.; Chen, Y.; Lu, X.; Tong, Y.; et al. A Natural Compound Containing a Disaccharide Structure of Glucose and Rhamnose Identified as Potential N-Glycanase 1 (NGLY1) Inhibitors. Molecules 2023, 28, 7758. https://doi.org/10.3390/molecules28237758
Liu R, Gu J, Ye Y, Zhang Y, Zhang S, Lin Q, Yuan S, Chen Y, Lu X, Tong Y, et al. A Natural Compound Containing a Disaccharide Structure of Glucose and Rhamnose Identified as Potential N-Glycanase 1 (NGLY1) Inhibitors. Molecules. 2023; 28(23):7758. https://doi.org/10.3390/molecules28237758
Chicago/Turabian StyleLiu, Ruijie, Jingjing Gu, Yilin Ye, Yuxin Zhang, Shaoxing Zhang, Qiange Lin, Shuying Yuan, Yanwen Chen, Xinrong Lu, Yongliang Tong, and et al. 2023. "A Natural Compound Containing a Disaccharide Structure of Glucose and Rhamnose Identified as Potential N-Glycanase 1 (NGLY1) Inhibitors" Molecules 28, no. 23: 7758. https://doi.org/10.3390/molecules28237758
APA StyleLiu, R., Gu, J., Ye, Y., Zhang, Y., Zhang, S., Lin, Q., Yuan, S., Chen, Y., Lu, X., Tong, Y., Lv, S., Chen, L., & Sun, G. (2023). A Natural Compound Containing a Disaccharide Structure of Glucose and Rhamnose Identified as Potential N-Glycanase 1 (NGLY1) Inhibitors. Molecules, 28(23), 7758. https://doi.org/10.3390/molecules28237758