
Antimicrobial Activity and Mechanisms of Walnut Green Husk Extract

Abstract
:1. Introduction
2. Results and Discussion
2.1. Antimicrobial Activity
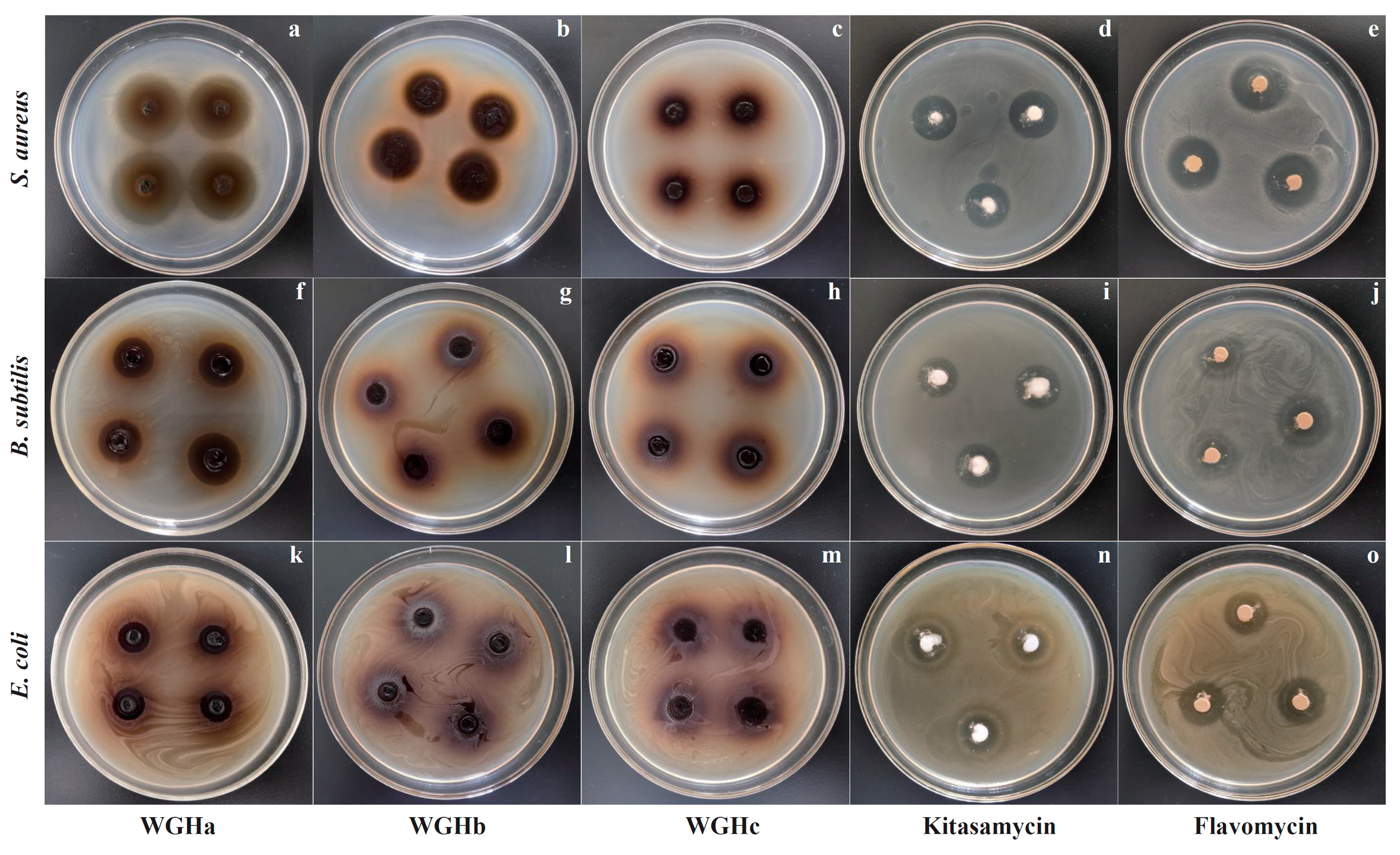
2.1.1. Inhibition Zones
2.1.2. Minimum Inhibitory Concentration (MIC)
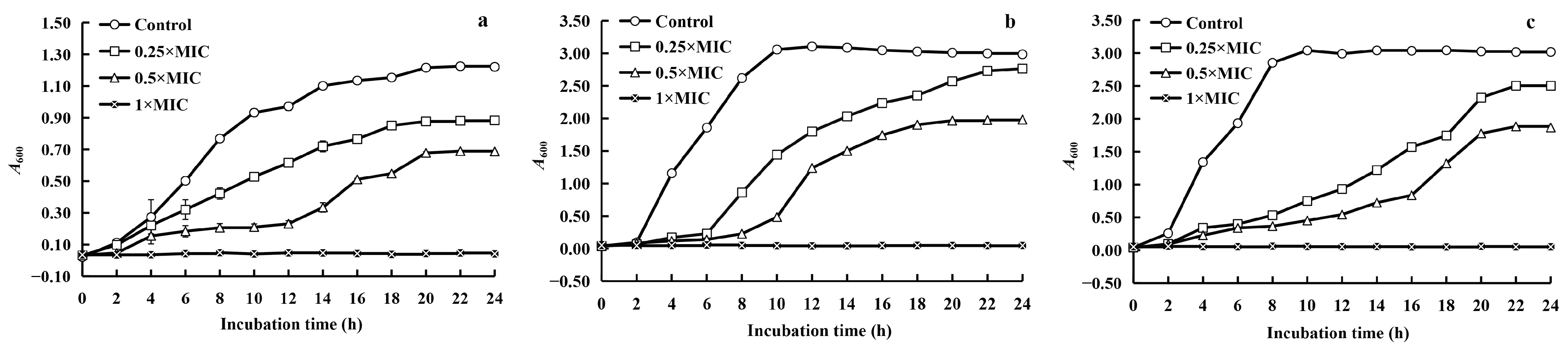
2.1.3. Growth Curve
2.2. Chemical Compositions of WGHa
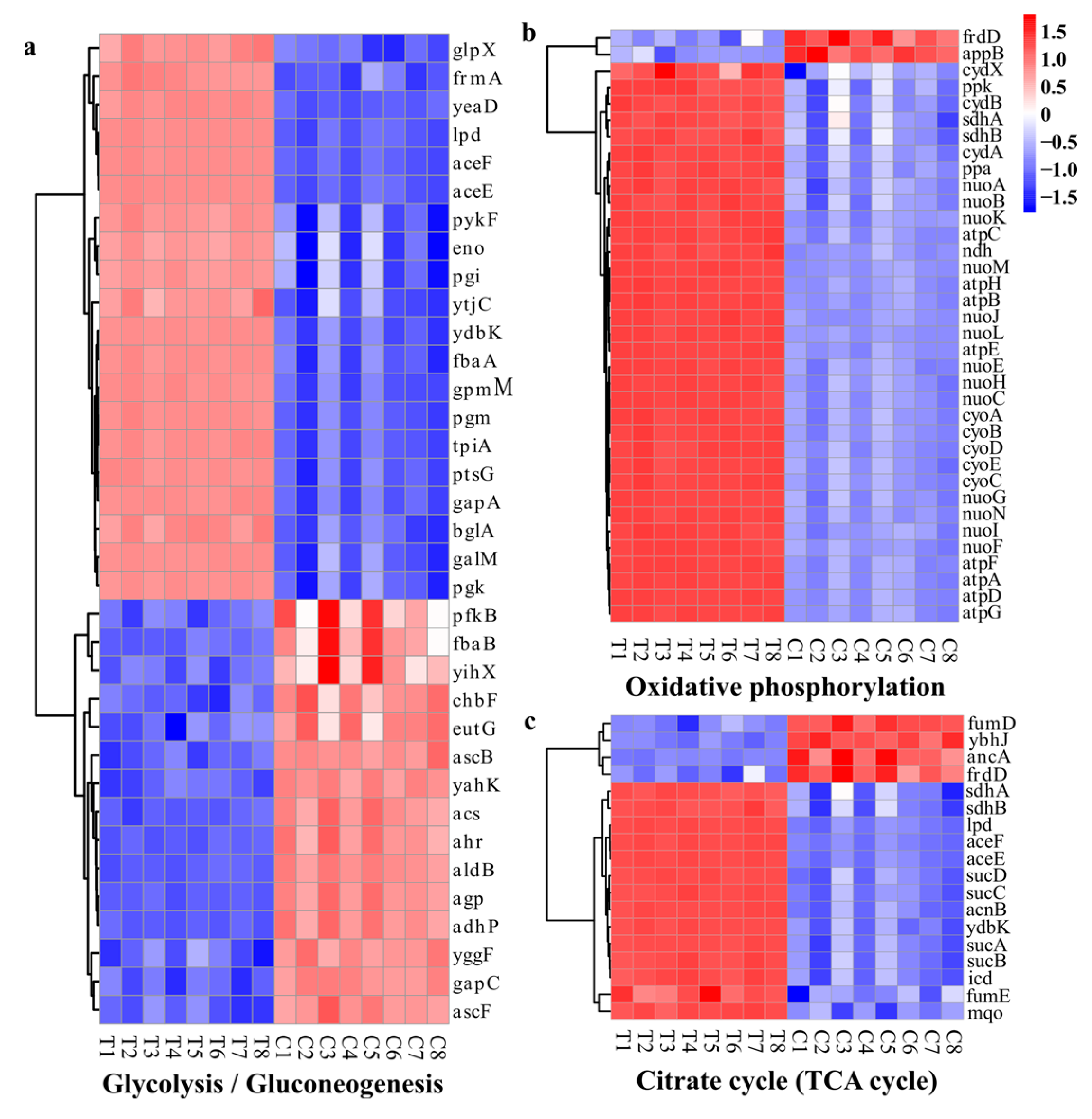
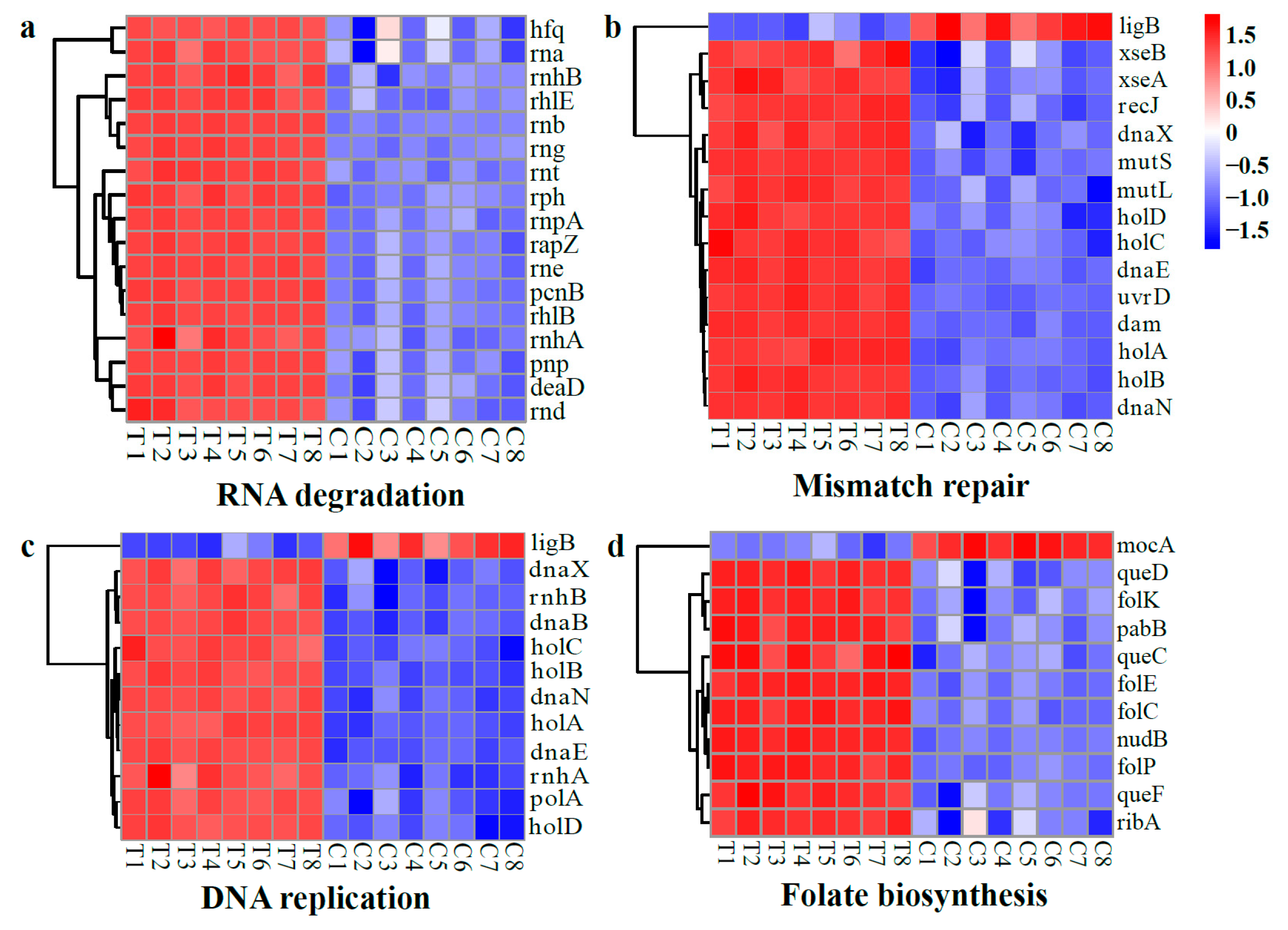
2.3. Antimicrobial Mechanisms of WGHa
2.3.1. Extracellular Alkaline Phosphatase (AKP) Activity
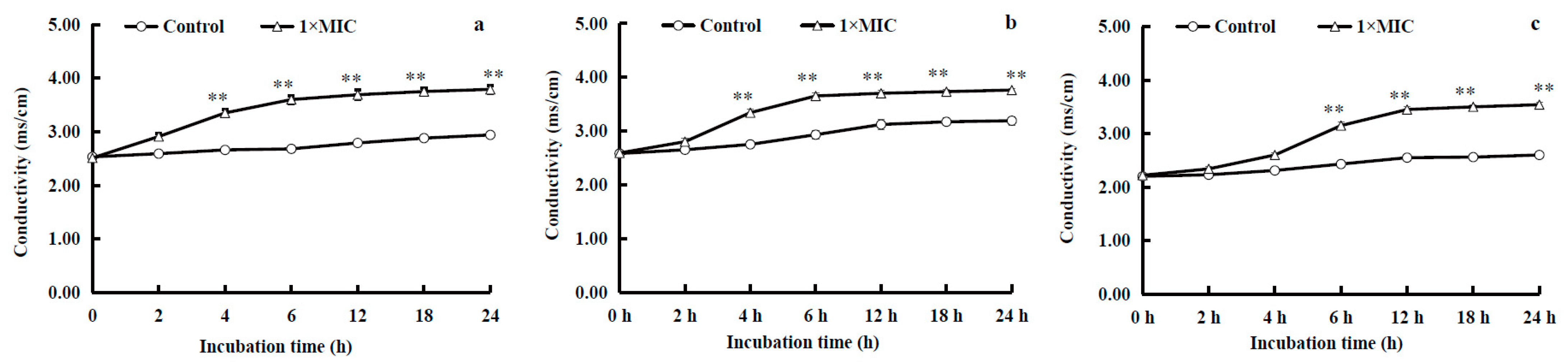
2.3.2. Electric Conductivity of the Medium
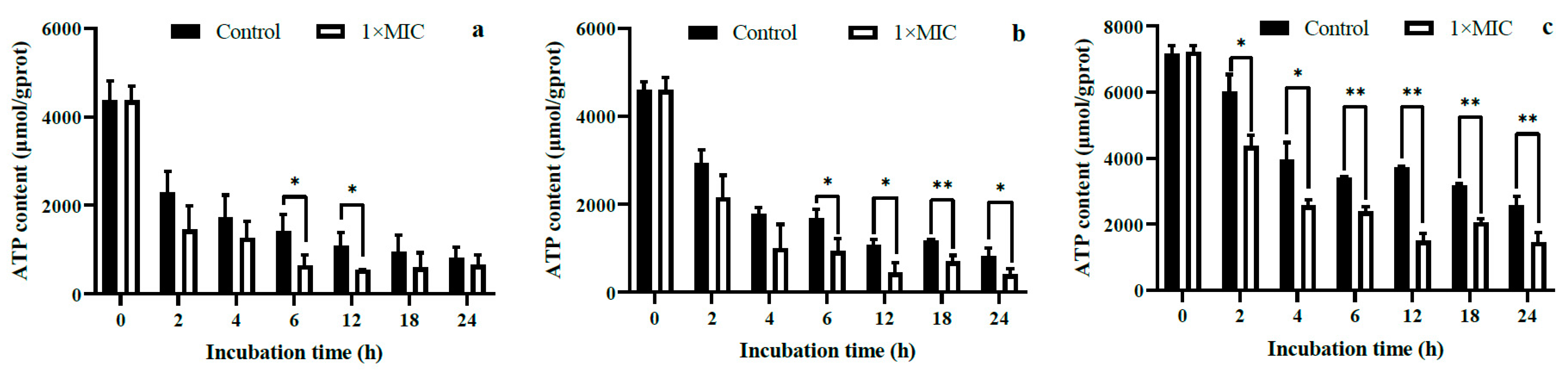
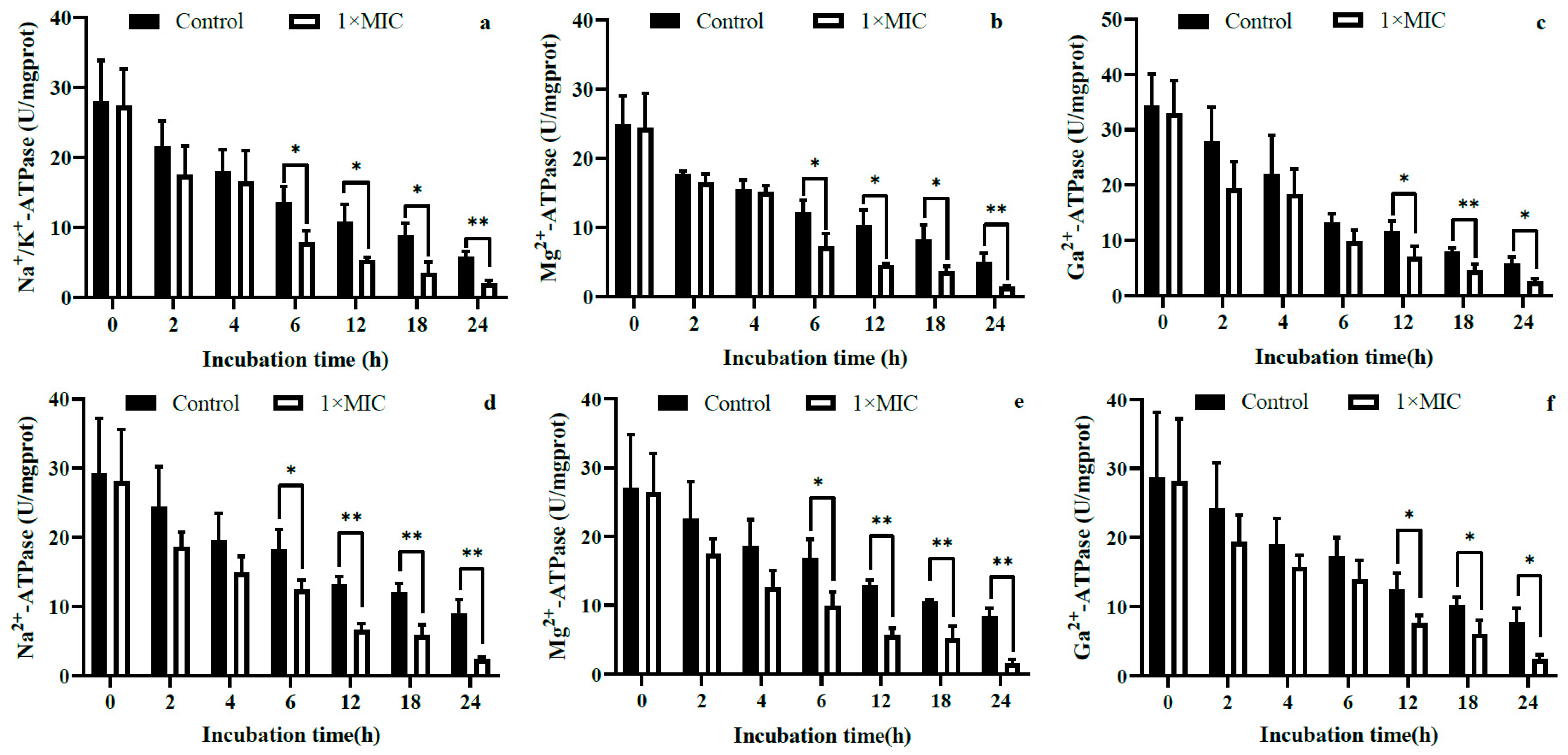
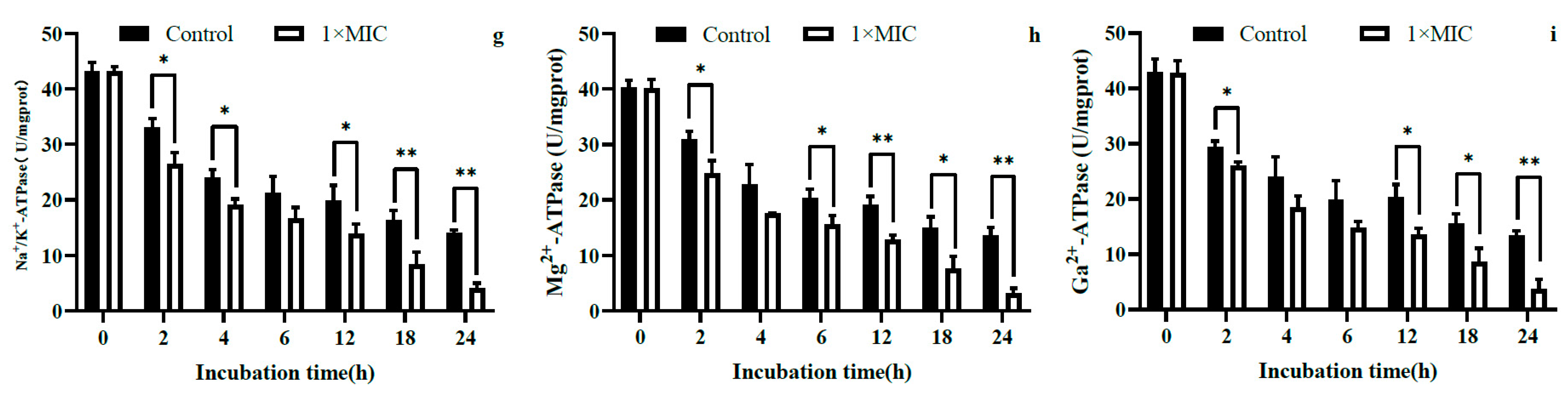
2.3.3. Intracellular ATP Content and ATPase Activity
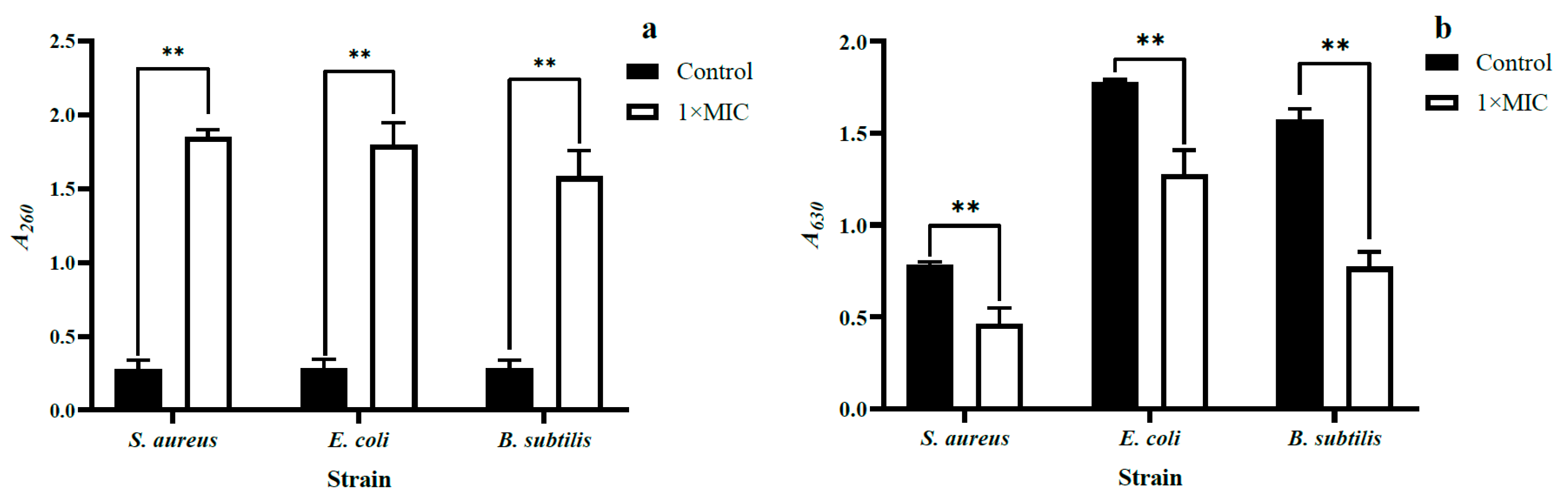
2.3.4. Loss of 260 nm Light-Absorbing Substances
2.3.5. Cellular Metabolic Vitality
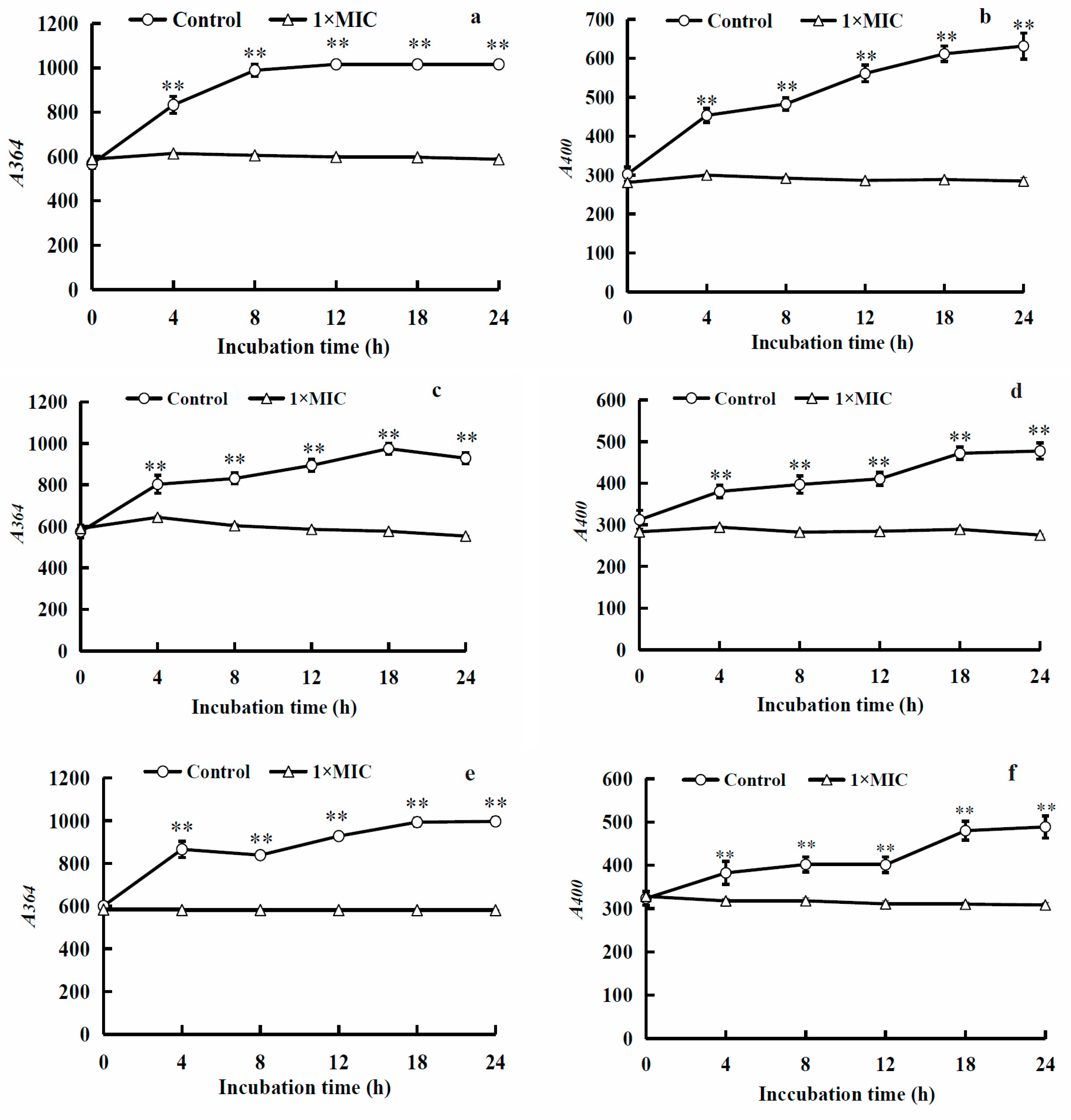
2.3.6. Fluorescence Intensity of Bacterial Nucleic Acids
2.3.7. Scanning Electron Microscopy (SEM)
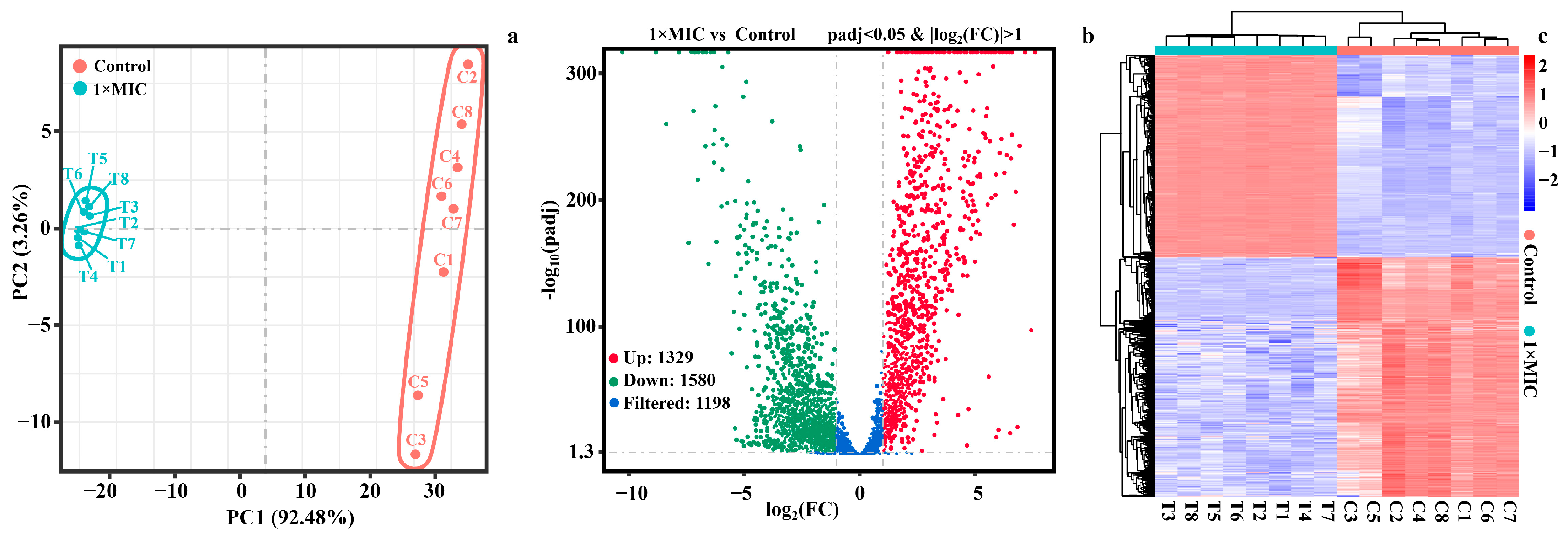
2.4. Transcriptome Analysis of Differentially Expressed Genes (DEGs)
2.4.1. Identification of DEGs
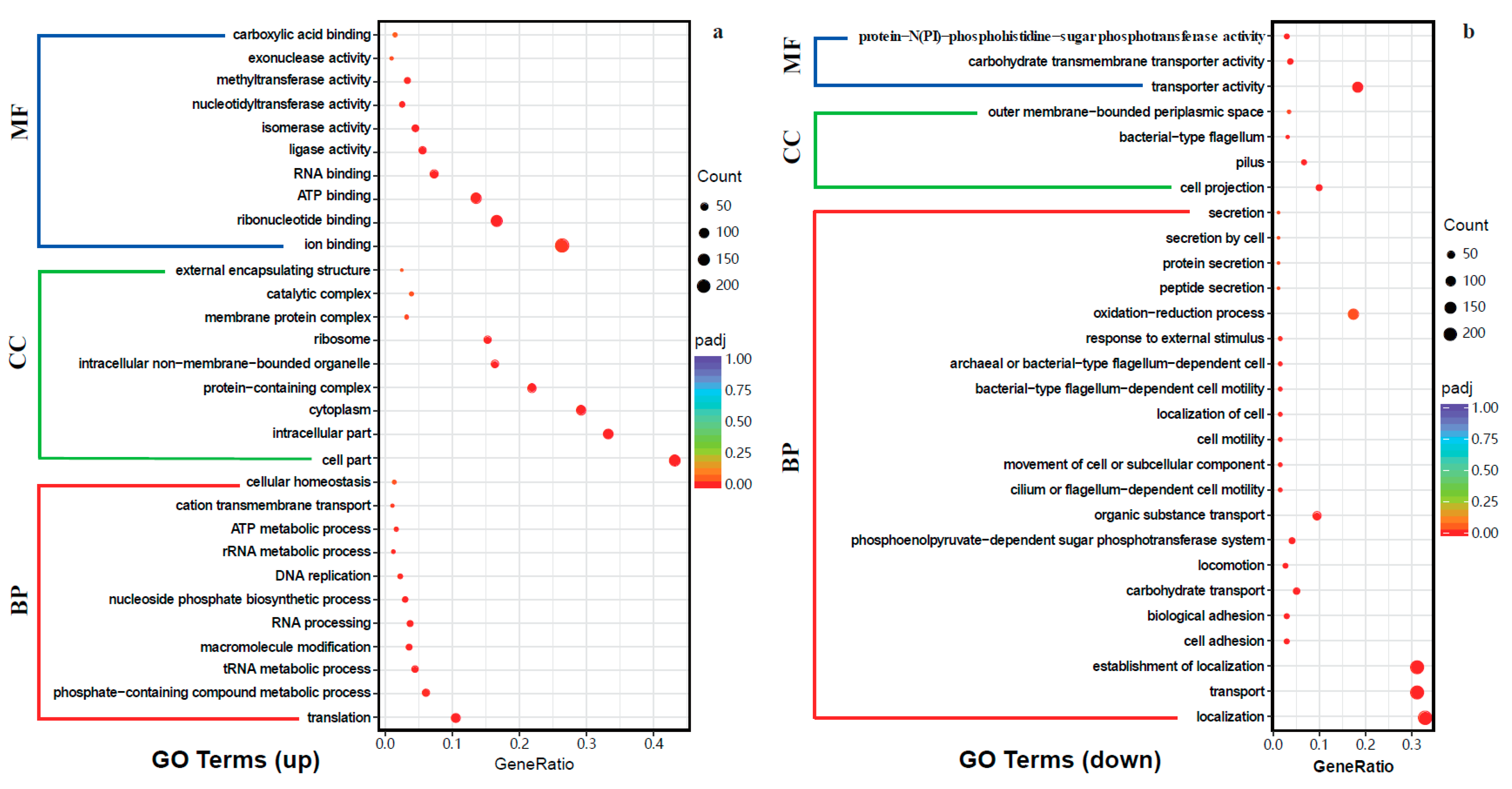
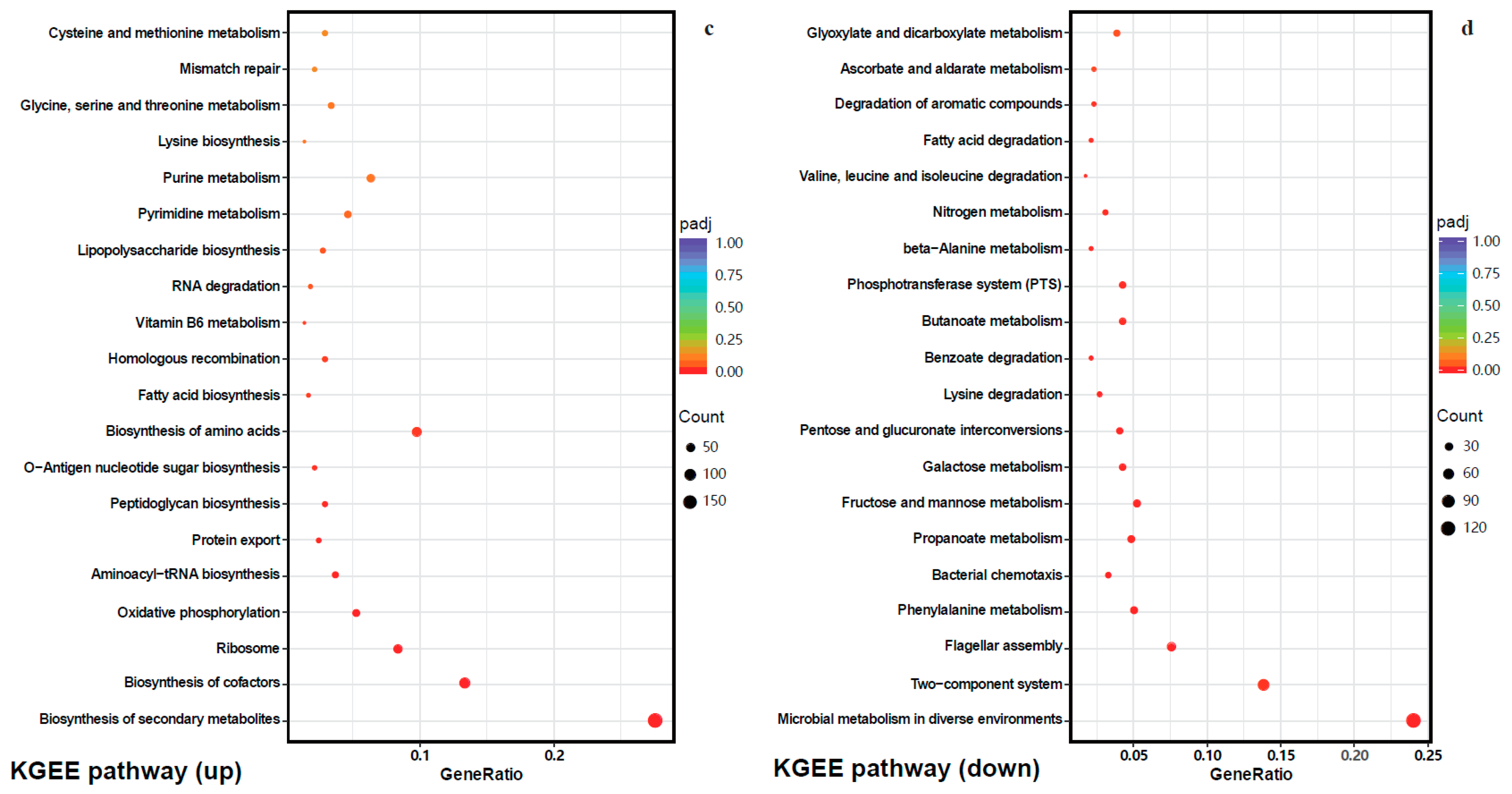
2.4.2. Gene Ontology (GO) Enrichment and Kyoto Encyclopedia of Genes and Genomes (KEGG) Pathway Analysis of DEGs
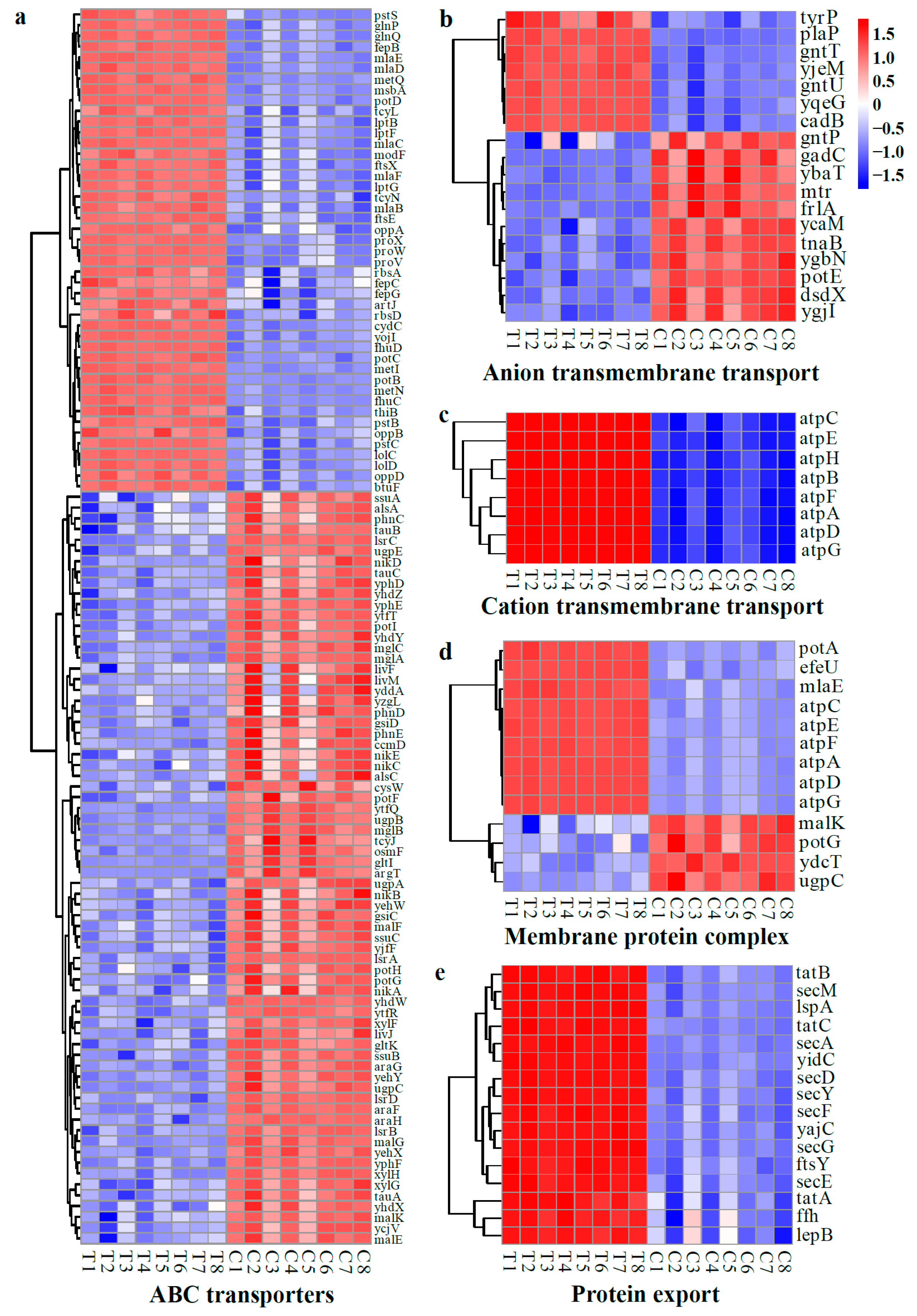
2.4.3. Gene Set Analysis of DEGs
DEGs Related to Cell Structures
DEGs Related to Cell Membrane Functions
DEGs Related to Environmental Adaptation
DEGs Related to Energy Metabolism
DEGs Related to Nucleic Acid Synthesis and Repair
3. Materials and Methods
3.1. Strains and Antibiotics
3.2. Preparation of WGH Extracts
3.3. Antimicrobial Activity of WGH Extracts
3.3.1. Diameter of Inhibition Zones
3.3.2. MIC
3.3.3. Kinetics of Growth
3.4. Identification of Compounds in WGHa via Ultrahigh-Performance Liquid Chromatography–Tandem Mass Spectrometry (UHPLC-MS/MS)
3.5. Antimicrobial Mechanisms of WGHa
3.5.1. Extracellular AKP Activity
3.5.2. Conductivity of the Medium, Intracellular ATP Content, and ATPase Activity
3.5.3. Loss of 260 nm Light-Absorbing Substances
3.5.4. Cellular Metabolic Vitality
3.5.5. Fluorescence Intensity of Bacterial Nucleic Acids
3.5.6. SEM
3.6. Transcriptome Sequencing of E. coli
3.7. Statistics
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cheng, G.; Hao, H.; Xie, S.; Wang, X.; Dai, M.; Huang, L.; Yuan, Z. Antibiotic alternatives: The substitution of antibiotics in animal husbandry? Front. Microbiol. 2014, 5, 217. [Google Scholar] [CrossRef] [PubMed]
- Álvarez-Martínez, F.J.; Barrajón-Catalán, E.; Herranz-López, M.; Micol, V. Antibacterial plant compounds, extracts and essential oils: An updated review on their effects and putative mechanisms of action. Phytomedicine 2021, 90, 153626. [Google Scholar] [CrossRef] [PubMed]
- Keita, K.; Darkoh, C.; Okafor, F. Secondary plant metabolites as potent drug candidates against antimicrobial-resistant pathogens. SN Appl. Sci. 2022, 4, 209. [Google Scholar] [CrossRef]
- Ajaykumar, C. Efficacy of herbal medicine in treating metabolic and endocrine disorders. In Treating Endocrine and Metabolic Disorders with Herbal Medicines; Hussain, A., Behl, S., Eds.; IGI Global: Hershey, PA, USA, 2021; pp. 236–255. [Google Scholar] [CrossRef]
- Oulahal, N.; Degraeve, P. Phenolic-rich plant extracts with antimicrobial activity: An alternative to food preservatives and biocides? Front. Microbiol. 2022, 12, 753518. [Google Scholar] [CrossRef] [PubMed]
- Shahrajabian, H.M.; Cheng, Q.; Sun, W. Application of herbal plants in organic poultry nutrition and production. Curr. Nutr. Food Sci. 2022, 18, 7. [Google Scholar] [CrossRef]
- Collares, L.J.; Turchen, L.M.; Guedes, R.N.C. Research trends, biases, and gaps in phytochemicals as insecticides: Literature survey and meta-analysis. Plants 2023, 12, 318. [Google Scholar] [CrossRef]
- Khir, R.; Pan, Z. Walnuts. In Integrated Processing Technologies for Food and Agricultural By-Products; Pan, Z., Zhang, R., Zicari, S., Eds.; Academic Press: San Diego, CA, USA, 2019; pp. 391–411. [Google Scholar] [CrossRef]
- Zicha, J. Global Walnut Production to Grow by 12% in 2022/23, USDA Says. Available online: https://www.mintecglobal.com/top-stories/global-walnut-production-to-grow-by-12-in-2022/23-usda-says (accessed on 15 July 2023).
- Barekat, S.; Nasirpour, A.; Keramat, J.; Dinari, M.; Meziane-Kaci, M.; Paris, C.; Desobry, S. Phytochemical composition, antimicrobial, anticancer properties, and antioxidant potential of green husk from several walnut varieties (Juglans regia L.). Antioxidants 2023, 12, 52. [Google Scholar] [CrossRef]
- Xi, M.; Hou, Y.; Cai, Y.; Shen, H.; Ao, J.; Li, M.; Wang, J.; Luo, A. Antioxidant and antimicrobial characteristics of ethyl acetate polar fractions from walnut green husk. J. Food Sci. 2023, 88, 1060–1074. [Google Scholar] [CrossRef]
- Vieira, V.; Pereira, C.; Abreu, R.M.V.; Calhelha, R.C.; Alves, M.J.; Coutinho, J.A.P.; Ferreira, O.; Barros, L.; Ferreira, I.C.F.R. Hydroethanolic extract of Juglans regia L. green husks: A source of bioactive phytochemicals. Food Chem. Toxicol. 2020, 137, 111189. [Google Scholar] [CrossRef]
- Popovici, C. Soxhlet extraction and characterisation of natura compounds from walnut (Juglans regia L.) by-products. Ukr. Food J. 2013, 2, 328–336. [Google Scholar]
- Fernández-Agulló, A.; Pereira, E.; Freire, M.S.; Valentão, P.; Andrade, P.B.; González-Álvarez, J.; Pereira, J.A. Influence of solvent on the antioxidant and antimicrobial properties of walnut (Juglans regia L.) green husk extracts. Ind. Crops Prod. 2013, 42, 126–132. [Google Scholar] [CrossRef]
- Movahhed, T.; Mehrabkhani, M.; Arefnezhad, M.; Hamedi, S.S.; Zare Mahmoudabadi, R.; Ghanbari, F.; Rostami, M. Antibacterial efficacy of walnut green husk (wgh) extract with zinc oxide nanoparticles on streptococcus mutans. Rep. Biochem. Mol. Biol. 2022, 11, 405–410. [Google Scholar] [CrossRef] [PubMed]
- Liao, Q.; Kim, E.J.; Tang, Y.; Xu, H.; Yu, D.-G.; Song, W.; Kim, B.J. Rational design of hyper-crosslinked polymers for biomedical applications. J. Polym. Sci. 2023, 1. [Google Scholar] [CrossRef]
- Song, W.; Zhang, M.; Huang, X.; Chen, B.; Ding, Y.; Zhang, Y.; Yu, D.G.; Kim, I. Smart L-borneol-loaded hierarchical hollow polymer nanospheres with antipollution and antibacterial capabilities. Mat. Today Chem. 2022, 26, 101252. [Google Scholar] [CrossRef]
- Petriccione, M.; Papa, S.; Ciniglia, C. Cell-programmed death induced by walnut husk washing waters in three horticultural crops. Environ. Sci. Pollut. Res. Int. 2014, 21, 3491–3502. [Google Scholar] [CrossRef] [PubMed]
- Green, M.R.; Sambrook, J. Alkaline phosphatase. Cold Spring Harb. Protoc. 2020, 2020, 100768. [Google Scholar] [CrossRef]
- Lan, W.; Zhao, X.; Chen, M.; Xie, J. Antimicrobial activity and mechanism of oregano essential oil against Shewanella putrefaciens. J. Food Saf. 2022, 42, e12952. [Google Scholar] [CrossRef]
- Wan, W.; Wang, Y.; Tan, J.; Qin, Y.; Zuo, W.; Wu, H.; He, H.; He, D. Alkaline phosphatase-harboring bacterial community and multiple enzyme activity contribute to phosphorus transformation during vegetable waste and chicken manure composting. Bioresour. Technol. 2020, 297, 122406. [Google Scholar] [CrossRef]
- Wang, S.; Yao, J.; Zhou, B.; Yang, J.; Chaudry, M.T.; Wang, M.; Xiao, F.; Li, Y.; Yin, W. Bacteriostatic effect of quercetin as an antibiotic alternative in vivo and its antibacterial mechanism in vitro. J. Food Prot. 2018, 81, 68–78. [Google Scholar] [CrossRef]
- Lin, L.; Gu, Y.; Li, C.; Vittayapadung, S.; Cui, H. Antibacterial mechanism of ε-Poly-lysine against Listeria monocytogenes and its application on cheese. Food Control 2018, 91, 76–84. [Google Scholar] [CrossRef]
- Konaté, K.; Hilou, A.; Mavoungou, J.F.; Lepengué, A.N.; Souza, A.; Barro, N.; Datté, J.Y.; M’batchi, B.; Nacoulma, O.G. Antimicrobial activity of polyphenol-rich fractions from Sida alba L. (Malvaceae) against co-trimoxazol-resistant bacteria strains. Ann. Clin. Microbiol. Antimicrob. 2012, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- Farhadi, F.; Khameneh, B.; Iranshahi, M.; Iranshahy, M. Antibacterial activity of flavonoids and their structure-activity relationship: An update review. Phytother. Res. 2019, 33, 13–40. [Google Scholar] [CrossRef] [PubMed]
- Song, X.; Liu, T.; Wang, L.; Liu, L.; Li, X.; Wu, X. Antibacterial effects and mechanism of mandarin (Citrus reticulata L.) essential oil against Staphylococcus aureus. Molecules 2020, 25, 4956. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Yang, M.; Wang, T.; Sun, Z.; Liu, M.; Zhang, J.; Zeng, Q.; Cai, C.; Li, Y. Antibacterial mechanism of vanillic acid on physiological, morphological, and biofilm properties of carbapenem-resistant Enterobacter hormaechei. J. Food Prot. 2020, 83, 576–583. [Google Scholar] [CrossRef] [PubMed]
- Khan, F.; Bamunuarachchi, N.I.; Tabassum, N.; Kim, Y.M. Caffeic acid and its derivatives: Antimicrobial drugs toward microbial pathogens. J. Agric. Food Chem. 2021, 69, 2979–3004. [Google Scholar] [CrossRef]
- Hughes, T.; Azim, S.; Ahmad, Z. Inhibition of Escherichia coli ATP synthase by dietary ginger phenolics. Int. J. Biol. Macromol. 2021, 182, 2130–2143. [Google Scholar] [CrossRef]
- Lin, L.; Wang, X.; Li, C.; Cui, H. Inactivation mechanism of E. coli O157:H7 under ultrasonic sterilization. Ultrason Sonochem. 2019, 59, 104751. [Google Scholar] [CrossRef]
- Lee, S.; Kim, K.; Lim, S.; Ahn, D. Antibacterial mechanism of Myagropsis myagroides extract on Listeria monocytogenes. Food Control 2014, 42, 23–28. [Google Scholar] [CrossRef]
- Siriwong, S.; Teethaisong, Y.; Thumanu, K.; Dunkhunthod, B.; Eumkeb, G. The synergy and mode of action of quercetin plus amoxicillin against amoxicillin-resistant Staphylococcus epidermidis. BMC Pharmacol. Toxicol. 2016, 17, 39. [Google Scholar] [CrossRef]
- Cushnie, T.P.; Lamb, A.J. Antimicrobial activity of flavonoids. Int. J. Antimicrob. Agents 2005, 26, 343–356. [Google Scholar] [CrossRef]
- Sun, X.; Zhou, T.; Wei, C.; Lan, W.; Zhao, Y.; Pan, Y.; Wu, V. Antibacterial effect and mechanism of anthocyanin rich Chinese wild blueberry extract on various foodborne pathogens. Food Control 2018, 94, 155–161. [Google Scholar] [CrossRef]
- Ohadoma, S.; Akuodor, G.; Amazu, L.; Michael, H. Quantitative estimation of total phenolic and total flavonoid contents of ethylacetate fraction of Chikadoma as a bactericidal agent. Asian J. Sci. Technol. 2020, 11, 11012–11014. [Google Scholar] [CrossRef]
- Omelon, S.; Georgiou, J.; Habraken, W. A cautionary (spectral) tail: Red-shifted fluorescence by dapi-dapi interactions. Biochem. Soc. Trans. 2016, 44, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Tanious, F.A.; Veal, J.M.; Buczak, H.; Ratmeyer, L.S.; Wilson, W.D. DAPI (4′,6-diamidino-2-phenylindole) binds differently to DNA and RNA: Minor-groove binding at AT sites and intercalation at AU sites. Biochemistry 1992, 31, 3103–3112. [Google Scholar] [CrossRef] [PubMed]
- Azqueta, A.; Collins, A. Polyphenols and DNA damage: A mixed blessing. Nutrients 2016, 8, 785. [Google Scholar] [CrossRef]
- Arslan, H.; Ondul Koc, E.; Ozay, Y.; Canli, O.; Ozdemir, S.; Tollu, G.; Dizge, N. Antimicrobial and antioxidant activity of phenolic extracts from walnut (Juglans regia L.) green husk by using pressure-driven membrane process. J. Food Sci. Technol. 2023, 60, 73–83. [Google Scholar] [CrossRef]
- Friedman, M. Chemistry, antimicrobial mechanisms, and antibiotic activities of cinnamaldehyde against pathogenic bacteria in animal feeds and human foods. J. Agric. Food Chem. 2017, 65, 10406–10423. [Google Scholar] [CrossRef]
- Su, M.; Liu, F.; Luo, Z.; Wu, H.; Zhang, X.; Wang, D.; Zhu, Y.; Sun, Z.; Xu, W.; Miao, Y. The antibacterial activity and mechanism of chlorogenic acid against foodborne pathogen Pseudomonas aeruginosa. Foodborne Pathog. Dis. 2019, 16, 823–830. [Google Scholar] [CrossRef]
- Schroeder, A.; Mueller, O.; Stocker, S.; Salowsky, R.; Leiber, M.; Gassmann, M.; Lightfoot, S.; Menzel, W.; Granzow, M.; Ragg, T. The RIN: An RNA integrity number for assigning integrity values to RNA measurements. BMC Mol. Biol. 2006, 7, 3. [Google Scholar] [CrossRef]
- Wang, Y.; Malkmes, M.J.; Jiang, C.; Wang, P.; Zhu, L.; Zhang, H.; Zhang, Y.; Huang, H.; Jiang, L. Antibacterial mechanism and transcriptome analysis of ultra-small gold nanoclusters as an alternative of harmful antibiotics against Gram-negative bacteria. J. Hazard. Mater. 2021, 416, 126236. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Makino, F.; Miyata, T.; Minamino, T.; Kato, T.; Namba, K. Structure of the molecular bushing of the bacterial flagellar motor. Nat. Commun. 2021, 12, 4469. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, N.A.; Ton-That, H. Bacterial pili and fimbriae. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd: Hoboken, NJ, USA, 2020. [Google Scholar] [CrossRef]
- Di Lorenzo, F.; Duda, K.A.; Lanzetta, R.; Silipo, A.; De Castro, C.; Molinaro, A. A journey from structure to function of bacterial lipopolysaccharides. Chem. Rev. 2022, 122, 15767–15821. [Google Scholar] [CrossRef] [PubMed]
- Hrast, M.; Sosič, I.; Sink, R.; Gobec, S. Inhibitors of the peptidoglycan biosynthesis enzymes MurA-F. Bioorg. Chem. 2014, 55, 2–15. [Google Scholar] [CrossRef] [PubMed]
- Higgins, C.F. ABC transporters: Physiology, structure and mechanism—An overview. Res. Microbiol. 2001, 152, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Bogdanov, M.; Lu, S.; Guan, Z.; Margolin, W.; Weiss, J.; Zheng, L. The phospholipid-repair system LplT/Aas in Gram-negative bacteria protects the bacterial membrane envelope from host phospholipase A2 attack. J. Biol. Chem. 2018, 293, 3386–3398. [Google Scholar] [CrossRef] [PubMed]
- Jeckelmann, J.M.; Erni, B. Transporters of glucose and other carbohydrates in bacteria. Pflugers Arch. 2020, 472, 1129–1153. [Google Scholar] [CrossRef]
- Karmakar, R. State of the art of bacterial chemotaxis. J. Basic Microbiol. 2021, 61, 366–379. [Google Scholar] [CrossRef] [PubMed]
- Ziege, R.; Tsirigoni, A.M.; Large, B.; Serra, D.O.; Blank, K.G.; Hengge, R.; Fratzl, P.; Bidan, C.M. Adaptation of Escherichia coli biofilm growth, morphology, and mechanical properties to substrate water content. ACS Biomater. Sci. Eng. 2021, 7, 5315–5325. [Google Scholar] [CrossRef]
- Penesyan, A.; Paulsen, I.T.; Kjelleberg, S.; Gillings, M.R. Three faces of biofilms: A microbial lifestyle, a nascent multicellular organism, and an incubator for diversity. NPJ Biofilms Microbiomes 2021, 7, 80. [Google Scholar] [CrossRef]
- Sharma, G.; Sharma, S.; Sharma, P.; Chandola, D.; Dang, S.; Gupta, S.; Gabrani, R. Escherichia coli biofilm: Development and therapeutic strategies. J. Appl. Microbiol. 2016, 121, 309–319. [Google Scholar] [CrossRef]
- Niba, E.T.; Naka, Y.; Nagase, M.; Mori, H.; Kitakawa, M. A genome-wide approach to identify the genes involved in biofilm formation in E. coli. DNA Res. 2007, 14, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Xue, T.; Zhao, L.; Sun, H.; Zhou, X.; Sun, B. LsrR-binding site recognition and regulatory characteristics in Escherichia coli AI-2 quorum sensing. Cell Res. 2009, 19, 1258–1268. [Google Scholar] [CrossRef] [PubMed]
- Rühle, T.; Leister, D. Assembly of F1F0-ATP synthases. Biochim. Biophys. Acta 2015, 1847, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Chen, L.; Ouyang, K.; Zhang, Q.; Wang, W. Antibacterial activity and mechanism of flavonoids from Chimonanthus salicifolius S. Y. Hu. and its transcriptome analysis against Staphylococcus aureus. Front. Microbiol. 2023, 13, 1103476. [Google Scholar] [CrossRef] [PubMed]
- Dogovski, C.; Xie, S.C.; Burgio, G.; Bridgford, J.; Mok, S.; McCaw, J.M.; Chotivanich, K.; Kenny, S.; Gnädig, N.; Straimer, J.; et al. Targeting the cell stress response of Plasmodium falciparum to overcome artemisinin resistance. PLoS Biol. 2015, 13, e1002132. [Google Scholar] [CrossRef]
- Jain, C. Degradation of mRNA in Escherichia coli. IUBMB Life. 2002, 54, 315–321. [Google Scholar] [CrossRef]
- Durica-Mitic, S.; Göpel, Y.; Amman, F.; Görke, B. Adaptor protein RapZ activates endoribonuclease RNase E by protein-protein interaction to cleave a small regulatory RNA. RNA 2020, 26, 1198–1215. [Google Scholar] [CrossRef]
- Zeman, M.K.; Cimprich, K.A. Causes and consequences of replication stress. Nat. Cell Biol. 2014, 16, 2–9. [Google Scholar] [CrossRef]
- Nazki, F.H.; Sameer, A.S.; Ganaie, B.A. Folate: Metabolism, genes, polymorphisms and the associated diseases. Gene 2014, 533, 11–20. [Google Scholar] [CrossRef]
- Liang, W.; Li, H.; Zhou, H.; Wang, M.; Zhao, X.; Sun, X.; Li, C.; Zhang, X. Effects of Taraxacum and Astragalus extracts combined with probiotic Bacillus subtilis and Lactobacillus on Escherichia coli-infected broiler chickens. Poult. Sci. 2021, 100, 101007. [Google Scholar] [CrossRef]
- Zgoda, J.R.; Porter, J.R. A convenient microdilution method for screening natural products against bacteria and fungi. Pharm. Biol. 2001, 39, 221–225. [Google Scholar] [CrossRef]
- Kang, J.; Liu, L.; Wu, X.; Sun, Y.; Liu, Z. Effect of thyme essential oil against Bacillus cereus planktonic growth and biofilm formation. Appl. Microbiol. Biotechnol. 2018, 102, 10209–10218. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Lin, S.; Jia, Y.; Rehman, F.; Zeng, S.; Wang, Y. Anthocyanin and spermidine derivative hexoses coordinately increase in the ripening fruit of Lycium ruthenicum. Food Chem. 2020, 311, 125874. [Google Scholar] [CrossRef]
- Wu, Q.; Cui, D.; Chao, X.; Chen, P.; Liu, J.; Wang, Y.; Su, T.; Li, M.; Xu, R.; Zhu, Y.; et al. Transcriptome analysis identifies strategies targeting immune response-related pathways to control enterotoxigenic Escherichia coli infection in porcine intestinal epithelial cells. Front. Vet. Sci. 2021, 8, 677897. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef]
- Anders, S.; Huber, W. Differential expression of RNA-Seq data at the gene level-the DESeq package. Eur. Mol. Biol. Lab. (EMBL) 2012, 10, f1000research. [Google Scholar]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef]
- Bu, D.; Luo, H.; Huo, P.; Wang, Z.; Zhang, S.; He, Z.; Wu, Y.; Zhao, L.; Liu, J.; Guo, J.; et al. KOBAS-i: Intelligent prioritization and exploratory visualization of biological functions for gene enrichment analysis. Nucleic Acids Res. 2021, 49, W317–W325. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bacteria | WGHa (200 mg/mL) | WGHb (200 mg/mL) | WGHc (200 mg/mL) | Kitasamycin (50 mg/mL) | Flavomycin (50 mg/mL) |
|---|---|---|---|---|---|
| S. aureus | 18.08 ± 1.62 a | 12.50 ± 1.68 c | 0.00 ± 0.00 d | 11.92 ± 0.90 c | 16.50 ± 0.79 b |
| B. subtilis | 9.25 ± 1.36 c | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 13.5 ± 1.00 a | 10.58 ± 0.99 b |
| E. coli | 3.75 ± 0.75 c | 0.00 ± 0.00 d | 0.00 ± 0.00 d | 7.08 ± 0.79 b | 11.00 ± 0.85 a |
| Bacteria | MIC (mg/mL) | |||
|---|---|---|---|---|
| WGHa | WGHb | WGHc | Extraction Solvents | |
| S. aureus | 6.25 | 200 | >200 | >200 |
| B. subtilis | 6.25 | >200 | >200 | >200 |
| E. coli | 25.00 | >200 | >200 | >200 |
| Compound Name | RT (min) | m/z | Adducts | Relative Content (%) | Classification | Formula |
|---|---|---|---|---|---|---|
| α-Cyperone | 0.9220 | 241.1533 | [M + Na]+ | 4.60 | Terpenoid | C15H22O |
| Chlorogenic acid | 2.5530 | 377.0841 | [M + Na]+ | 2.64 | Polyphenol | C16H18O9 |
| Syringic acid | 4.8165 | 199.0585 | [M + H]+ | 7.60 | Monophenol | C9H10O5 |
| Gentiopicrin | 5.2351 | 379.0995 | [M + Na]+ | 1.29 | Terpenoid | C16H20O9 |
| Trans-ferulic acid | 5.2351 | 177.0530 | [M + H – H2O]+ | 2.90 | Monophenol | C10H10O4 |
| Myricetin | 5.9873 | 319.0421 | [M + H]+ | 1.61 | Flavonoid | C15H10O8 |
| Picroside II | 6.6416 | 535.1387 | [M + Na]+ | 1.36 | Terpenoid | C23H28O13 |
| Quercetin | 6.9312 | 303.0491 | [M + H]+ | 12.05 | Flavonoid | C15H10O7 |
| Taxifolin | 7.2128 | 305.0643 | [M + H]+ | 4.72 | Flavonoid | C15H12O7 |
| Cyanidin-3-glucoside | 7.5995 | 449.1042 | [M]+ | 2.17 | Terpenoid | C21H21O11 |
| Kaempferol-3-O-α-l-arabinoside | 8.1943 | 419.0941 | [M + H]+ | 3.82 | Terpenoid | C35H52O9 |
| Brucine | 8.6146 | 395.2022 | [M + H]++ | 1.86 | Alkaloid | C23H26N2O4 |
| Coniferyl aldehyde | 8.7457 | 179.0691 | [M + H]+ | 1.52 | Monophenol | C10H10O3 |
| 6-Methylcoumarin | 9.4412 | 161.0596 | [M + H]+ | 3.48 | Coumarin | C10H8O2 |
| Licoricidin | 10.3541 | 447.2166 | [M + Na]+ | 2.24 | Flavonoid | C26H32O5 |
| Baccatin III | 10.5884 | 609.2280 | [M + Na]+ | 1.62 | Terpenoid | C31H38O11 |
| Dehydroandrographolide | 17.1554 | 333.1991 | [M + H]+ | 1.62 | Terpenoid | C20H28O4 |
| Fluoren-9-one | 0.8525 | 179.0470 | [M – H]− | 8.95 | Ketone | C13H8O |
| Succinic acid | 1.2660 | 117.0127 | [M – H]− | 12.20 | Dicarboxylic acid | C4H6O4 |
| Gallic acid | 1.4213 | 169.0070 | [M – H]− | 65.89 | Polyphenols | C7H6O5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Li, W.; Li, X.; Liu, J.; Chen, Y. Antimicrobial Activity and Mechanisms of Walnut Green Husk Extract. Molecules 2023, 28, 7981. https://doi.org/10.3390/molecules28247981
Wang L, Li W, Li X, Liu J, Chen Y. Antimicrobial Activity and Mechanisms of Walnut Green Husk Extract. Molecules. 2023; 28(24):7981. https://doi.org/10.3390/molecules28247981
Chicago/Turabian StyleWang, Lei, Wenjing Li, Xuanyue Li, Jiancheng Liu, and Yong Chen. 2023. "Antimicrobial Activity and Mechanisms of Walnut Green Husk Extract" Molecules 28, no. 24: 7981. https://doi.org/10.3390/molecules28247981