Abstract
The present work investigates the potential for enhancing the NMR signals of DNA nucleobases by parahydrogen-based hyperpolarization. Signal amplification by reversible exchange (SABRE) and SABRE in Shield Enables Alignment Transfer to Heteronuclei (SABRE-SHEATH) of selected DNA nucleobases is demonstrated with the enhancement (ε) of 1H, 15N, and/or 13C spins in 3-methyladenine, cytosine, and 6-O-guanine. Solutions of the standard SABRE homogenous catalyst Ir(1,5-cyclooctadeine)(1,3-bis(2,4,6-trimethylphenyl)imidazolium)Cl (“IrIMes”) and a given nucleobase in deuterated ethanol/water solutions yielded low 1H ε values (≤10), likely reflecting weak catalyst binding. However, we achieved natural-abundance enhancement of 15N signals for 3-methyladenine of ~3300 and ~1900 for the imidazole ring nitrogen atoms. 1H and 15N 3-methyladenine studies revealed that methylation of adenine affords preferential binding of the imidazole ring over the pyrimidine ring. Interestingly, signal enhancements (ε~240) of both 15N atoms for doubly labelled cytosine reveal the preferential binding of specific tautomer(s), thus giving insight into the matching of polarization-transfer and tautomerization time scales. 13C enhancements of up to nearly 50-fold were also obtained for this cytosine isotopomer. These efforts may enable the future investigation of processes underlying cellular function and/or dysfunction, including how DNA nucleobase tautomerization influences mismatching in base-pairing.
1. Introduction
With an energy difference between nuclear spin states that is much less than the thermal energy, kT, NMR accesses only ~10−3% of available molecules (e.g., ~0.0032% for 1H spins at B0 = 9.4 T), and thus suffers from poor detection sensitivity. Hyperpolarization methods [1] such as dynamic nuclear polarization (DNP) [2,3], spin-exchange optical pumping (SEOP) [4,5], and parahydrogen-induced polarization (PHIP) [2,6,7] produce highly non-Boltzmann spin populations to greatly enhance NMR signals. With hyperpolarization, the rapid NMR signal acquisition of less-sensitive and low-natural-abundance nuclei (e.g., 15N, 13C, 129Xe, etc.) is greatly facilitated, making it feasible to perform the NMR/MRI of low-concentration species. Among these hyperpolarization techniques, PHIP-based approaches [6,7,8] are attractive because they can be conducted rapidly with low operational costs, merely requiring access to parahydrogen gas (p-H2, a nuclear spin isomer of ordinary molecular hydrogen), a catalyst, and an appropriate external magnetic field.
One such PHIP-based approach is signal amplification by reversible exchange (SABRE) [9,10]. Unlike hydrogenative PHIP [8], which requires the pairwise addition of p-H2 atoms across an unsaturated chemical bond, SABRE does not result in permanent chemical change of the substrate. Instead, SABRE involves the transient binding of the substrate and p-H2 to the catalyst to enable the transfer of spin order to the substrate’s nuclear spins through the J-coupling network, allowing bulk hyperpolarization of the “free” substrate to build up over time with subsequent exchange (Figure 1). Unlike 1H SABRE, hyperpolarization of nuclei with lower gyromagnetic ratios (e.g., non-quadrupolar ones like 15N and 13C) usually does not suffer from strong inter/intramolecular dipolar interactions, and thus often decays over longer time scales [11,12,13,14]; long-lived spin states can offer even longer hyperpolarization lifetimes [15]. Whereas the field-matching condition for 1H SABRE is in the mT regime, the efficient SABRE of heteronuclei requires microtesla fields [11,16], and thus is commonly performed within a magnetic shield with a constant or variable internal field, i.e., SABRE-SHEATH (SABRE in SHield Enables Alignment Transfer to Heteronuclei) [16].
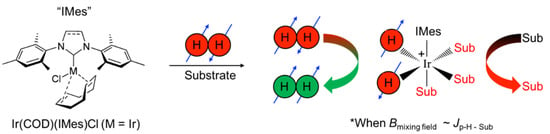
Figure 1.
In the presence of p-H2 and substrate, a pre-catalyst [e.g., Ir(1,5-cyclooctadeine)(1,3-bis(2,4,6-trimethylphenyl)imidazolium)Cl, Ir(COD)(IMes)Cl)] first undergoes a 4- to 6-coordinate transformation induced by the initial exposure to p-H2, giving rise to the standard “IrIMes” SABRE catalyst [17]. The formation of a transient hexa-coordinate center, wherein the symmetry of the nascent hydride spins is broken, allows spin order to be transferred to reversibly ligating substrates through the J-coupling network, particularly within a mixing field (Bmixing field) that roughly matches the frequency difference between source (parahydrogen-derived hydrides) and target spins (protons or other spin−1/2 nuclei of the exchangeable substrate) to the magnitude of the J coupling.
SABRE can thus produce solution-phase hyperpolarization for various applications, including the creation of biologically-friendly hyperpolarized spectral probes for NMR, and metabolic contrast agents for MRI [13,14,18,19,20,21,22,23,24]. However, despite its advantages, a persistent challenge of the methodology has been the limited range of applicable substrates due to the necessity of requiring an sp- and sp2-hybridized lone pair to interact with the catalyst’s Ir-metal center, initially limiting SABRE mostly to nitrogen heterocycles [2,9,11,12,16]. While a number of approaches have recently been developed to greatly widen SABRE’s scope [18,19,20] (including biologically relevant carboxylic acid derivatives like α-ketoglutarate and pyruvate) [21,22,25,26,27,28], the myriad biologically relevant NHCs nevertheless make this structural motif of continued interest for SABRE targets for both fundamental studies and envisioned applications [23].
Of the NHCs studied thus far, derivatives of purine comprise the most widely occurring NHC family in nature and consist of fused pyrimidine and imidazole rings [29,30,31]. Moreover, derivatives of purine and pyrimidine, such as the nucleobases adenine, thymine, guanine, and cytosine, comprise DNA and participate in specific hydrogen bonding to complete canonical (A:T/G:C) Watson-Crick base pairing [32] (Figure 2). The resulting primary nucleic acid sequence gives rise to secondary structures composed of supercoils that result from protein complexes wound about a DNA helix [33,34]. These structures, along with other modifications (e.g., methylation, histone modification, etc.), are responsible for epigenetic gene regulation [34,35], which has far-reaching consequences in both normal (e.g., cell differentiation) and pathogenic development (e.g., cancer) [34,36,37]. This type of control primarily dictates which segments of the genetic sequence are available for reading (thereby acting as on-and-off switches [38]), depending on the shape, spacing, and composition of the helix and associated further levels of the structure. With RNA these effects are further compounded by the additional functions of RNA, ranging from signaling to catalysis [39,40].
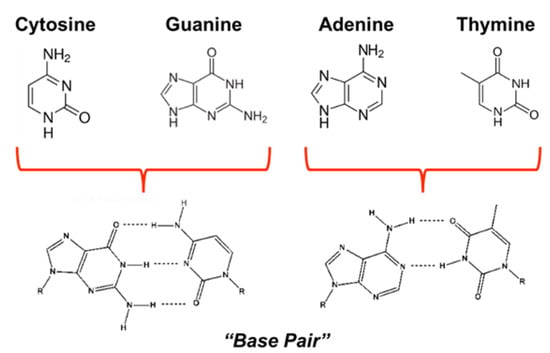
Figure 2.
Structures of primary DNA nucleobases that form base pairs through hydrogen-bonding.
Interestingly, nucleobase tautomers play a key role in epigenetic control, as they can alter the secondary level of the DNA’s structure [41,42]. For example, tautomerization of a nucleobase can lead to distortions in the shape of (or spacing between) DNA strands, which are then targeted by DNA repair enzymes. In some cases, an enzyme may fail to initiate the repair, thus epigenetically leading to silencing or changes in the gene’s expression [42], with implications for cancer and other diseases—depending on the location of the tautomerization [35]. Thus, any method that can probe epigenetic interactions has potential value in both clinical and research settings [35,37].
Due to the ubiquity of the nucleobases that comprise DNA and RNA, along with the myriad of other functions that nucleobases perform (e.g., cell signaling [43,44,45] and ATP [46]), such compounds are ripe for investigation by methods that may be able to sensitively and non-invasively report on nucleobase interactions in living systems or biological media. Such developments would be particularly relevant if those methods could complement the capabilities of other bioanalytical techniques under development (e.g., Refs. [43,47]). Given that the detection of nucleobase-containing biomarkers of disease have opened new areas of research, there should be natural interest in the investigation of rapid and inexpensive parahydrogen-based methods for hyperpolarizing nucleobases and their use in potential applications; however, to date there have only been a few such studies reported. In one type of approach, the unique resonances of catalyst-bound hyperpolarized hydrides [48,49,50] are used to detect the presence of specific nucleobases (and their tautomers) with high sensitivity, including in complex biological media [51,52]. Hövener et al. reported 1H hyperpolarization of adenine and adenosine as part of efforts to demonstrate continuous polarization in MRI applications [53]. More recently, as part of a larger effort to demonstrate SABRE enhancement in a large number of different molecules, Colell et al. [12] showed 15N enhancement of 15N-labelled adenine (ε~200) as an example where enamine-imine tautomers are sensitive to SABRE-SHEATH [12]; this sensitivity may be exploitable in potential biological applications. Finally, work has also been performed to optimize 1H and 15N hyperpolarization of pyrimidine, which is the framework for cytosine and thymine [54]. In the present work, we explore the applicability of SABRE and SABRE-SHEATH to various nucleobases, motivated by the desire to expand the current scope of these approaches to support the future development of new techniques for studying various biological systems and diseases—both for cellular studies and ultimate potential in vivo applications.
2. Materials and Methods
The homogenous (Ir(COD)(IMes)Cl, MW = 639.67 g·mol−1) pre-catalyst was synthesized as previously described [17]. Each NMR solution consisted of 4 mM catalyst and 40 mM substrate in 600 µL solution of either 100% C2D5OD (3-methyladenine) or 92% C2D5OD: 8% D2O (cytosine and 6-O-methylguanine); D2O was necessary to increase the solubility of the latter nucleobases. Experimental setups at SIUC and Vanderbilt were described previously [11,16,19,55]. Each sample solution was transferred to a 5 mm O.D. NMR tube affixed with a 0.25-inch O.D. Teflon tube, sealed with a wye-connector, and activated at elevated temperatures (60–70 °C) for at least 10 min prior to acquisition. The experimental setup is schematically shown in Figure 3.
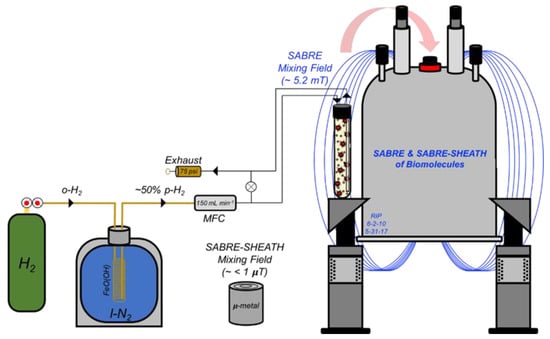
Figure 3.
1H SABRE and 15N/13C SABRE-SHEATH setup (used for experiments at SIUC). Hydrogen gas (H2) from a cylinder (with initial p-H2 fraction of 25%) is passed over a catalyst bed of FeO(OH) cooled in liquid nitrogen for ~50% conversion to p-H2. A mass flow controller (MFC) at 150 mL min−1 allows for the precise regulation of p-H2 bubbling rate within the NMR tube; pressure was typically maintained at 75 psi. 1H SABRE and 15N/13C SABRE-SHEATH experiments were performed by bubbling p-H2 through the sample while placed in either the NMR magnet’s fringe field (~5 mT) or a mu-metal magnetic shield (~1 µT), respectively, prior to rapid manual transfer of the sample to the 9.4 T magnet for high-field detection.
Parahydrogen generators used in this work provided either: p-H2 enrichment of ~50% (at 75 psi and 150 mL min−1 bubbling rate or at 75 psi and 110 mL min−1 bubbling rate at SIUC and Vanderbilt, respectively); or, ~90% p-H2 (for some heteronuclear studies at Vanderbilt). Most 15N/13C SABRE-SHEATH experiments were performed at Vanderbilt University using a µ-metal shield that was degaussed manually with a Variac and degaussing coil prior to use [56]; all of those experiments used medium-wall NMR tubes. All NMR experiments were performed on either an Agilent 400 MHz DD2 spectrometer with a wide-bore actively shielded (Oxford) magnet (SIUC) or a Bruker AVANCE III 400 MHz spectrometer with a narrow-bore actively shielded magnet (Vanderbilt). Single-scan acquisitions with 10°–pulses were used to acquire both SABRE-enhanced 1H spectra and 1H spectra from thermally polarized samples. Pulses of 90° of 18 µs (15N) or 10 µs (13C) and 1 scan were used to acquire 15N and 13C SABRE-SHEATH spectra. 15N thermally polarized reference spectra were acquired with a standard 8.65 M 15N2-imidazole solution (in D2O) using a 90°–pulse of 18 µs and 250 s pre-acquisition delay time, and one scan. 13C thermally polarized reference spectra were acquired with the activated cytosine solution at 70 °C using a 90°–pulse of 18 µs, 250 s delay time, and one scan.
3. Results and Discussion
3.1. 1H SABRE of DNA Nucleobases and Ethanol
Here we probe the efficacy of 1H SABRE enhancement for the DNA nucleobases 3-methyladenine, 6-O-methylguanine, and cytosine (others were attempted but lacked sufficient solubility under our conditions). We begin by showing 1H NMR spectra from thermally polarized and SABRE enhanced 3-methyladenine in 100% C2D5OD at 70 °C (Figure 4a). Interestingly, both aromatic hydrogens (labelled 1HA and 1HB) are hyperpolarized (albeit weakly), but with 1HA exhibiting slightly larger (factor of ~1.5) enhancement over 1HB, suggestive of preferential binding of the imidazole ring over the pyrimidine ring to the catalyst. Naively, such differential enhancement might have also been explained by differences in relaxation. However, 1HA exhibits a shorter T1 time constant of ~5 s in comparison to 1HB (~8 s, albeit measured at high field instead of the mixing field; Figure 4d; at milli-tesla and micro-tesla fields, the relaxation rates may be significantly different [57], although here we expect the qualitative trend to be the same). The faster rate of hyperpolarization decay for 1HA presumably reflects greater intermolecular interactions with the catalyst itself (which can act as a relaxation agent) [58], as well as possible contributions from hydrogen-bonding interactions with C2D5OD in the complex (discussed below).
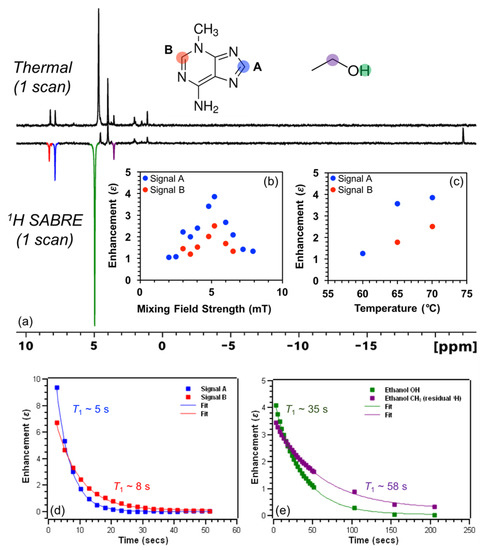
Figure 4.
(a) 1H NMR thermal (top) and SABRE enhanced (bottom) spectra of 3-methyl adenine at 70 °C show differential enhancement (and presumptive preferential binding) of the imidazole ring with a maximum ε of ~(-)4 at (b) ~5.2 mT and (c) 70 °C. HP 1H signals of 3-methyladenine exhibit T1 time constants (d) of ~5 s (1HA) and ~8 s (1HB), whereas HP signals from residual 1H spins of C2D5OD solvent molecules decay with T1 time constants (e) of ~35 s (-OH) and ~58 s (-CHD, residual 1H). Note that the -OH resonance of ethanol shifts due to a reduction of temperature when the solution is bubbled with p-H2. T1 values: in (d): 4.52 ± 0.03 (blue); 7.60 ± 0.13 (red); in (e): 35.1 ± 0.4 (green); 58.2 ± 1.2 (purple); similar T1 values were measured at the lower temperatures (not shown).
The present 3-methyladenine enhancement pattern is the reverse of that observed by Hövener et al. in adenine, which included a larger enhancement for the pyrimidine 1HB resonance (by a factor of approximately two) compared to that of the imidazole 1HA spin [53]. Such behavior might be rationalized by the fact that 3-methyladenine has one fewer available sp2-hybridized nitrogen atoms on the pyrimidine ring to bind to the catalyst; on the other hand, Hövener et al. also observed a larger enhancement for the imidazole resonance in adenosine despite the increased sterics likely caused by the sugar moiety [53]. More generally, binding with the 5-membered ring might be expected to be preferred because of the reduced sterics/stronger binding of 5-membered rings (e.g., imidazole) compared to 6-membered rings (e.g., pyridine). For example, high 15N polarization values in excess of 50% were reported for methylated imidazole with -CH3 in the ortho- position in the case of metronidazole [13,14,59], but no detectable 15N polarization (P15N < 0.01%) was seen for methylated pyridine in the ortho- position in the case of 2-picoline [60]. 1H SABRE of 3-methyladenine as a function of mixing field strength (Figure 4b) shows a maximum enhancement for both signals at ~5.2 mT, which was then selected as the mixing field for the remaining 1H SABRE experiments presented here. 1H SABRE of 3-methyladenine as a function of temperature (60–70 °C, Figure 4c) shows an increase in enhancement with temperature, which can be attributed to the better matching of exchange rates of 3-methlyadenine and p-H2 with the catalyst, once the temperature was raised sufficiently to help mitigate the otherwise poor solubility of the substrate in 100% C2D5OD below 60 °C.
Figure 4a also reveals a weak 1H SABRE of the resonance from residual -OH (slightly shifted due to temperature drop during p-H2 bubbling) and -CHD (residual 1H) of the bulk solvent, C2D5OD. There have been only a few previous studies of hyperpolarized alcohol solvents via SABRE (e.g., Ref. [61]). Currently, there are three proposed mechanisms for hyperpolarization of solvent alcohol molecules: (1) hyperpolarization through direct coordination to the metal center; (2) A SABRE-RELAY [18] type mechanism (particularly in a slightly acidic environment), wherein a hydrogen ion transfers to free substrate (post hyperpolarization), is hyperpolarized from spin coupling to the substrate protons, and then exchanges with the alcohol group on the solvent; and (3) a solvent molecule hydrogen-bonds to a free nitrogen of a (e.g., catalyst-bound) substrate, allowing the spin order to transfer via the extended scalar coupling network. Given that 1HA shows greater enhancement over 1HB, the imidazole ring of 3-methyladenine contains two nitrogen atoms, and only a single Ir-hydride signal is present (−22 ppm), we suggest that the ethanol solvent molecules may become hyperpolarized predominantly through hydrogen-bonding to 3-methyladenine while bound at the metal-center (Figure 5); direct binding may also contribute, though it is expected that SABRE-RELAY is unlikely to be a significant contributor because of the relatively low 1H enhancements observed for the substrate.
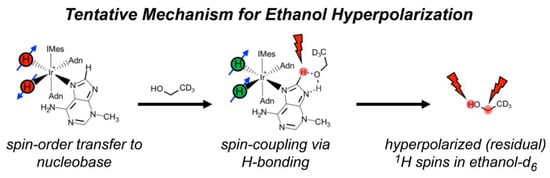
Figure 5.
Illustration of the proposed mechanism for hyperpolarization of residual protons of deuterated ethanol solvent molecules. In this scenario, bound 3-methyladenine becomes hyperpolarized, deuterated ethanol (with residual 1H spins) hydrogen-bonds to 3-methyladenine, and then the solvent molecule becomes hyperpolarized via spin-relayed SABRE.
The shorter T1 time constant of 1HA for 3-methyladenine (Figure 4d) may reflect intermolecular interactions, which may include exchange with hydrogen-bonding solvent molecules. Interestingly, deuterated ethanol residual -OH and -CHD spins exhibit hyperpolarization decay (T1) time constants of ~35 s and ~58 s at 70 °C (Figure 4e), respectively, which is substantially longer than the 1H’s of 3-methyladenine. For residual 1H of C2D5OD, the contribution from intra- and intermolecular dipole-dipole interactions to T1 relaxation are minimized since the gyromagnetic ratio of 2H is ~6.5 times smaller than 1H. The shorter T1 for the residual -OH resonance likely reflects the greater participation in exchange.
Expanding the scope of 1H SABRE to other DNA nucleobases, we also studied cytosine (Figure 6a) and 6-O-methylguanine (Figure 6b). Small but clear 1H SABRE enhancements were observed for 1H resonances of both of these substrates. The weakness of the effects can partly be attributed to the need for 8% D2O to help solubilize the substrates, even at elevated temperatures; the use of aqueous media is challenging for SABRE due to the poor solubility of H2 in water and a reduced substrate exchange rate with the catalyst [62,63,64,65,66,67,68], thus often resulting in lower SABRE enhancements.
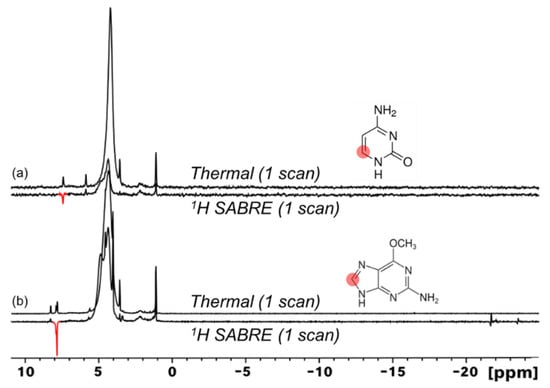
Figure 6.
1H SABRE spectra of cytosine ((a), amino tautomer shown) and 6-O-methylguanine (b) show minimal hyperpolarization of 1H nuclei adjacent to binding nitrogen atoms. In (b), the bottom spectrum is scaled by a factor of ~1.9 compared to the top spectrum.
Analogous to 3-methyladenine, 6-O-methylguanine appears to show preferential catalyst binding of the imidazole ring, likely due to less steric hindrance. Hydride resonances showed generally weaker signals compared to those obtained with 3-methyladenine (Figure 4a), particularly with cytosine, where virtually no detectable hydride resonances were observed. The weaker hydride signals are generally consistent with lower SABRE enhancements. Moreover, whereas 3-methyladenine gave rise to a single hydride peak (indicating magnetic equivalence of the complex’s two hydride sites on the NMR time scale), the multiple (weak) hydride signals observed with 6-O-methylguanine (comprising an asymmetric, antiphase doublet and a weak absorptive singlet) likely indicate the simultaneous presence of at least two different hydrogenated metal complexes (i.e., having different ligands) in significant concentration (see, for example, Refs. [20,22]). Note also the lack of SABRE enhancement of residual solvent resonances. Tautomerization has been suggested to result in a decrease in 1H enhancements [69]. Indeed, tautomerization, along with elevated temperatures [70,71] and the weak binding of these substrates, likely contributes to the weakness of the 1H SABRE effects observed for this set of molecules. Nonetheless, we have shown successful 1H SABRE for three nucleobases, including the DNA base cytosine and the modified base 6-O-methylguanine for the first time.
3.2. 15N and 13C SABRE-SHEATH of Nucleobases
While 1H SABRE represents a rapid screening method for molecules of interest (and was used as such here), 1H hyperpolarization can suffer rapid T1 decay due to strong inter- and intramolecular interactions. Non-quadrupolar heteronuclei (e.g., 15N and 13C) suffer weaker dipolar interactions and thus generally allow greater accumulation and retention of hyperpolarization because of longer T1 values. Moreover, accessing such nuclei allows for nuclei-specific investigations into biological processes. 15N and 13C SABRE-SHEATH of DNA nucleobases may thus prove to be more useful in evaluating tautomerization dynamics—in addition to potentially enabling future applications. Here we begin by showing natural-abundance 15N (0.365%) SABRE-SHEATH of 3-methyladenine at 70 °C in 100% C2D5OD (Figure 7a). SABRE-SHEATH shows the enhancement of two adjacent 15N NMR signals from two nitrogen atoms (sites/resonances “A” and “B”) that integrate to a ~2:1 ratio for 15NA compared to 15NB, which when compared to the thermally polarized signal from 15N2-imidazole, corresponds to enhancement values of ε ~3300 (corresponding to 15N polarization just above 1%) and ε ~1900, respectively. The ~230 ppm chemical shifts of these two resonances are consistent with the values expected for the N1, N3, and N7 positions of adenine, with the N9 and -NH2 positions expected ~160 ppm and ~78 ppm, respectively [72,73,74]. However, no enhancements are observed for the NMR signals in those lower ranges. Moreover, given that no enhancement would be expected for the N3 site in 3-methyladenine (because of steric inhibition of catalyst binding), the enhanced sites are likely associated with the N1 (pyrimidine ring) and N7 (imidazole ring) positions. Moreover, to be consistent with the 1H SABRE results in Figure 4a (where the larger 1H enhancement was observed on the imidazole proton), we tentatively assign the more-downfield peak (NA, with the greater SABRE enhancement) in Figure 7a to N7, and the more-upfield peak (NB) to N1. The absence of any signal attributable to the N9 site would be consistent with the prevalence of a tautomer wherein the N9 position is protonated (as it is commonly depicted).
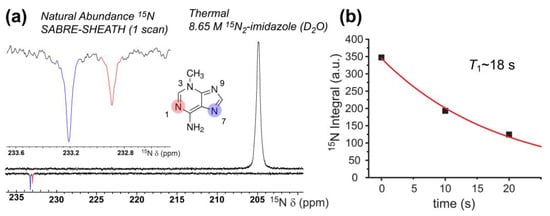
Figure 7.
(a) Natural-abundance 15N SABRE-SHEATH of 40 mM 3-methyladenine (bottom) reveals ε values of ~3300 (15NA) and ~1900 (15NB), respectively, when compared to a thermally polarized signal from a 15N2-imidazole reference sample (top); a close-up of the enhanced resonances is shown in the inset. Tentative 15N shift assignments were made in light of Refs. [72,73,74]. (b) Hyperpolarization decay curve obtained from single-point acquisitions with varying delay at 9.4 T prior to acquisition (the fit was constrained by including a fourth point set to zero at 1000 s).
This interpretation of the 15N results would be consistent with the 1H SABRE results for 3-methyladenine and C2D5OD (Figure 4), as well as the notion of preferential binding of the imidazole ring to the catalyst. Figure 7b shows a 15N T1 decay curve, wherein a somewhat rapid decay in signal (T1 = 18.4 ± 1.8 s) is observed. These results represent the first natural abundance 15N enhancement (spin concentration: ~150 µM) of a nucleobase via SABRE-SHEATH.
We also investigated the 15N SABRE-SHEATH of doubly-labelled cytosine (2-13C; 1,3-15N2). The enhancement of cytosine 15N signals from a 40 mM solution (92% C2D5OD: 8% D2O) at 70 °C is shown in Figure 8a, following polarization transfer in a mixing field of ~1 µT for 10 s; a short bubbling time was used to prevent a rapid decrease in temperature that could cause the substrate to crash out of the solution. In the figure, we see a modest selectivity in relative enhancement (1:0.75) for 15NA (206 ppm) to 15NB (141 ppm) compared to expectations based on a thermally polarized spectrum from a sample containing only labelled cytosine (i.e., without catalyst) with relative integrals of 1:1 (not shown); the absolute enhancements were ~240-fold and ~200-fold, respectively. A T1 measurement (performed as part of a different experimental run; discussed further below) saw an even-greater variance in the intensity of the two sites (Figure 8a inset).
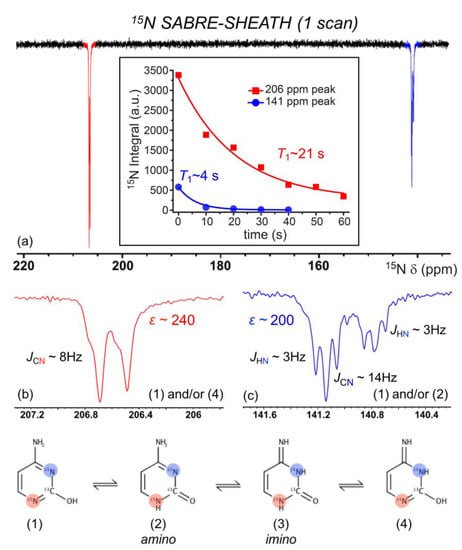
Figure 8.
(a). 15N NMR SABRE-SHEATH of doubly-labeled cytosine appears to reveal the preferential binding of tautomers (1), (2), and/or (4) via 15NA (b) over 15NB (c). Steric hindrance from the adjacent ketone and amine to 15NB likely reduces the binding ability of tautomers (1) and (2). Enhancements calculated from the 8.65 M 15N2-imidazole thermally polarized reference section shown in Figure 7. 15N spectra here were taken without 1H decoupling. The inset of (a) shows that the NA (206 ppm) and NB (141 ppm) sites decay with different 15N T1 values (measured at 9.4 T; each point is a separate experiment with variable delay time at high field prior to acquisition) of 4.3 ± 0.5 s (blue) and 20.6 ± 4.2 s (red).
Such deviation may suggest the preferential binding of specific tautomers of cytosine (Figure 8). For both 15N atoms to become hyperpolarized, tautomerization of the labile 1H about the ketone must take place to generate both sp2-hybridized 15N atoms. Thus, tautomers (1), (2), and/or (4) (Figure 8) can potentially bind to the catalyst. We suggest that the larger ε of 15NA over 15NB is due to preferential binding of (1) and/or (4) to 15NA because of steric hindrance from the adjacent ketone and amine to 15NB, thus reducing interactions of 15NB with the catalyst. This is further supported by our weak but non-zero 1H SABRE for the adjacent 1H (Figure 6). The signal at ~206 ppm (NA) shows what appears to be two overlapping doublets that are not well-resolved, each with a J-coupling of ~8 Hz that likely arises from JCN for that 15N coupled to the labeled 13C site. Interestingly, the signal at ~141 ppm (NB) shows two unique triplets (split by a presumed JCN for that site of ~14.5 Hz). Previous work by Shchepin et al. on the sensitivity of 15N of imidazole to pH revealed a ~30 ppm up-field shift in the 15N resonance as pH is varied from 1 to 12, thus reporting on protonated and unprotonated 15N [75]. The finer splitting arises from JNH; the above J assignments are consistent with thermally polarized 1H-decoupled 15N experiments (not shown), where the decoupling collapsed the ~3 Hz splittings but not the larger (unequal) splittings. The 15N relaxation measurements performed at 9.4 T (Figure 8a inset) showed that the NA site had a similar high-field T1 (20.6 ± 4.2 s) as that of the enhanced 15N site in 3-methyladenine. However, the NB site’s hyperpolarization decayed much more quickly (T1 = 4.3 ± 0.5 s), suggesting the contribution of additional relaxation mechanisms for this spin (e.g., greater contributions from exchange) that could serve to exacerbate the selective 15N SABRE enhancement.
Finally, we present cytosine 13C enhancements via SABRE-SHEATH. Figure 9a shows a 13C SABRE-SHEATH spectrum from doubly-labeled cytosine obtained under conditions of 1H decoupling, compared to a thermally polarized reference spectrum from the same sample. A close-up of these spectra is shown in Figure 9c. There, the spectra manifest a non-first-order doublet of doublets arising from the two JCN couplings to NA and NB. Although the four peaks exhibit equal intensities for the 13C thermally polarized spectrum, the SABRE-SHEATH spectrum shows a highly asymmetric pattern of enhancement, which likely arises from unequal efficiencies of polarization transfer through the level anti-crossing regime achieved while the sample was in the magnetic shield. Taken together, the average enhancement (integrating over all four peaks) is ~21-fold. In another series of experiments, 13C SABRE-SHEATH spectra were obtained from doubly-labeled cytosine, but without 1H decoupling (Figure 9b,d). For these experiments, symmetric multiplets were observed (Figure 9d) with average effective splittings of ~6–8 Hz, reflecting both JCN and JCH contributions. The larger 13C enhancements observed (up to nearly ~50-fold) likely reflect the improved experimental efficiency of polarization transfer and are likely unrelated to the decoupling condition. A series of such spectra were obtained with a variable delay at 9.4 T prior to acquisition, allowing the high-field 13C hyperpolarization lifetime to be measured for the 13C cytosine resonance (T1 = 22.9 ± 0.8 s).
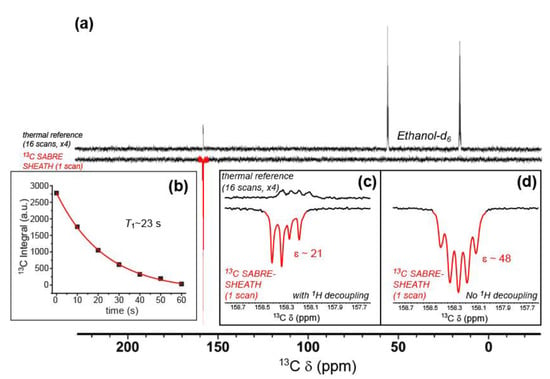
Figure 9.
(a) 13C NMR SABRE-SHEATH of doubly-labeled cytosine (bottom), compared to a thermally polarized reference scan from the same sample (16 scans, top). Close-ups of the cytosine resonances from these spectra, both taken under conditions of broadband 1H-decoupling, are shown in (c); enhancement (ε) ~21-fold. (b) The hyperpolarization decay curve for the integrated cytosine carbonyl 13C resonance, obtained from a second set of SABRE-SHEATH experiments (measured at 9.4 T; each point is a separate experiment with variable delay time at high field prior to acquisition), yielding T1 of 22.9 ± 0.8 s. Data points were taken from spectra obtained without 1H decoupling, an example of which is shown in (d); ε~48-fold; that spectrum also has a very weak, broad peak at 150 ppm (not shown) that is tentatively assigned to cytosine bound to the catalyst.
A summary of the measurements obtained in this work—SABRE enhancements and T1 relaxation measurements—is contained below in Table 1. Taken together, the hyperpolarization lifetimes of the heteronuclei measured for these nucleobases is shorter than may be expected. However, when contemplating the rapid T1 decays for hyperpolarized 15N and 13C signals of the present nucleobases, it should be noted that chemical shift anisotropy (CSA) is likely to be a significant contributor to T1 relaxation of 15N and 13C in these systems. Because CSA relaxation is a function of the square of field strength, SABRE-hyperpolarized DNA nucleobases may indeed prove useful at lower field strengths (e.g., clinical MRI or benchtop NMR), wherein greater enhancements and longer T1 time constants are likely.

Table 1.
Summary of nucleobase SABRE enhancements and T1 values.
It is also worth noting that the order-of-magnitude lower 15N SABRE-SHEATH enhancements for (doubly-labeled) cytosine compared to (naturally abundant) 3-methyladenine are qualitatively consistent with the 1H SABRE results for these molecules (Figure 4 and Figure 6). Such weaker 15N enhancements would also be much more difficult to observe (and hence optimize) with naturally abundant cytosine. Moreover, it may be difficult to achieve large 13C SABRE-SHEATH enhancements in nucleobases without the 15N labeling of the nitrogen sites, because abundant quadrupolar 14N spins can greatly accelerate 13C relaxation in the magnetic shield due to scalar relaxation of the second kind [76].
4. Conclusions
In this study, we have expanded the accessibility of SABRE and SABRE-SHEATH of DNA nucleobases by demonstrating NMR signal enhancement of: 1H spins in 3-methyladenine, cytosine, and 6-O-guanine; 15N spins in 3-methyladenine and cytosine; and 13C spins in cytosine. 1H SABRE generally exhibited weak enhancements for each studied nucleobase. However, 1H SABRE studies of 3-methyladenine revealed that simple chemical modification (compared to previous work on adenine) gave rise to preferential binding of the imidazole ring over the pyrimidine ring. Moreover, 1H SABRE also showed hyperpolarization of the solvent molecules (residual protons of deuterated ethanol), ostensibly through hydrogen-bonding to 3-methyladenine, with T1 values approaching ~60 s; the proposed mechanism will be explored in more detail with future studies. Preferential binding of the imidazole ring is further supported by the natural-abundance 15N SABRE-SHEATH enhancement of 3-methyladenine, upwards of ε~3300—the first natural-abundance result for a DNA nucleobase. Finally, 15N and 13C enhancements of cytosine (upwards of ε~240 and ε~50, respectively) via SABRE-SHEATH suggest a balance between tautomerization and catalyst binding kinetics. It is likely that significantly larger enhancements could be achieved by utilizing more-recently optimized hyperpolarizer platforms and conditions [22], greater p-H2 enrichment (in some cases), and alternative pulse sequences designed to improve SABRE-SHEATH efficiency [77]. Thus, while DNA nucleobases may represent a family of challenging SABRE substrates, these results help pave the way for a variety of envisioned biological studies in the future, including efforts to further the fundamental understanding of the interplay of nucleobase tautomerization, base pairing, and disease. For example, hyperpolarized nucleobases may be useful in cellular or in vivo cell-signaling studies. It may also become possible to probe how certain genetic sequences may be more vulnerable to tautomerization (i.e., via mutagenic exposure) in cellular/cell-lysate studies (e.g., by hyperpolarizing one or more nucleotides within a short DNA sequence and exposing it to a tautomerizing agent). Moreover, mismatches in base-pairing could potentially be investigated by unzipping the strand at elevated temperatures, hyperpolarizing the nucleobases, and allowing the strand to anneal upon rapid cooling. Thus, polarization transfer to pairing nucleobases may allow the study of mutagenesis on a molecular level. Indeed, a clear understanding of interactions between SABRE-sensitive and insensitive nucleobases, followed by their various derivatives including nucleotides, may eventually culminate into the hyperpolarization of single- and double-stranded DNA.
Author Contributions
Conceptualization, B.E.K., M.E.G., K.V.K., I.V.K., E.Y.C. and B.M.G.; methodology, B.E.K., M.E.G., R.V.S., K.V.K., E.Y.C. and B.M.G.; software, B.E.K. and M.E.G.; validation, B.E.K., M.E.G., E.Y.C. and B.M.G.; formal analysis, B.E.K., M.E.G., J.A.M., J.L.G., L.B.B., M.N.L. and B.M.G.; investigation, B.E.K., M.E.G., J.A.M., J.L.G., L.B.B., M.N.L., R.V.S. and K.V.K.; resources, R.V.S., K.V.K., I.V.K., E.Y.C. and B.M.G.; data curation, B.E.K., M.E.G., J.L.G., R.V.S., E.Y.C. and B.M.G.; writing—original draft preparation, B.E.K., M.E.G. and B.M.G.; writing—review and editing, B.E.K., M.E.G., M.N.L., R.V.S., I.V.K., E.Y.C. and B.M.G.; visualization, B.E.K., M.E.G. and B.M.G.; supervision, B.E.K., R.V.S., K.V.K., I.V.K., E.Y.C. and B.M.G.; project administration, I.V.K., E.Y.C. and B.M.G.; funding acquisition, R.V.S., I.V.K., E.Y.C. and B.M.G. All authors have read and agreed to the published version of the manuscript.
Funding
This work was funded by NSF (CHE-1905341, CHE-1904780, CHE-1416432, REU DMR-1757954, REU DMR-2150489). IVK thanks the Russian Science Foundation (grant #22-43-04426) for supporting experimental work, and the Russian Ministry of Science and Higher Education for access to NMR equipment. Prof. Roman V. Shchepin is grateful for startup funding from South Dakota School of Mines and Technology (SDSM&T), Department of Chemistry, Biology, and Health Sciences (CBH).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is available upon request to the corresponding author.
Acknowledgments
We thank A. Coffey for assisting with providing access to p-H2 at Vanderbilt.
Conflicts of Interest
BMG, EYC declare stake ownership in XeUS Technologies.
References
- Nikolaou, P.; Goodson, B.M.; Chekmenev, E.Y. NMR Hyperpolarization Techniques for Biomedicine. Chem. Eur. J. 2015, 21, 3156–3166. [Google Scholar] [CrossRef] [PubMed]
- Kovtunov, K.V.; Pokochueva, E.V.; Salnikov, O.G.; Cousin, S.F.; Kurzbach, D.; Vuichoud, B.; Jannin, S.; Chekmenev, E.Y.; Goodson, B.M.; Barskiy, D.A.; et al. Hyperpolarized NMR Spectroscopy: D-DNP, PHIP, and SABRE Techniques. Chem. Asian J. 2018, 13, 1857–1871. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Hilty, C. Applications of dissolution dynamic nuclear polarization in chemistry and biochemistry. Magn. Reson. Chem. 2018, 56, 566–582. [Google Scholar] [CrossRef]
- Goodson, B.M. Nuclear magnetic resonance of laser-polarized noble gases in molecules, materials, and organisms. J. Magn. Reson. 2002, 155, 157–216. [Google Scholar] [CrossRef]
- Walker, T.G.; Happer, W. Spin-exchange optical pumping of noble-gas nuclei. Rev. Mod. Phys. 1997, 69, 629–642. [Google Scholar] [CrossRef]
- Hovener, J.B.; Pravdivtsev, A.N.; Kidd, B.; Bowers, C.R.; Gloggler, S.; Kovtunov, K.V.; Plaumann, M.; Katz-Brull, R.; Buckenmaier, K.; Jerschow, A.; et al. Parahydrogen-Based Hyperpolarization for Biomedicine. Angew. Chem. Int. Ed. 2018, 57, 11140–11162. [Google Scholar] [CrossRef]
- Eisenberg, R. Parahydrogen-Induced Polarization-a New Spin on Reactions with H2. Acc. Chem. Res. 1991, 24, 110–116. [Google Scholar] [CrossRef]
- Reineri, F.; Cavallari, E.; Carrera, C.; Aime, S. Hydrogenative-PHIP polarized metabolites for biological studies. Magn. Reson. Mater. Phys. Biol. Med. 2021, 34, 25–47. [Google Scholar] [CrossRef]
- Rayner, P.J.; Duckett, S.B. Signal Amplification by Reversible Exchange (SABRE): From Discovery to Diagnosis. Angew. Chem. Int. Ed. 2018, 57, 6742–6753. [Google Scholar] [CrossRef]
- Adams, R.W.; Aguilar, J.A.; Atkinson, K.D.; Cowley, M.J.; Elliott, P.I.; Duckett, S.B.; Green, G.G.; Khazal, I.G.; Lopez-Serrano, J.; Williamson, D.C. Reversible interactions with para-hydrogen enhance NMR sensitivity by polarization transfer. Science 2009, 323, 1708–1711. [Google Scholar] [CrossRef]
- Truong, M.L.; Theis, T.; Coffey, A.M.; Shchepin, R.V.; Waddell, K.W.; Shi, F.; Goodson, B.M.; Warren, W.S.; Chekmenev, E.Y. (15)N Hyperpolarization by Reversible Exchange Using SABRE-SHEATH. J. Phys. Chem. C 2015, 119, 8786–8797. [Google Scholar] [CrossRef] [PubMed]
- Colell, J.F.; Logan, A.W.; Zhou, Z.; Shchepin, R.V.; Barskiy, D.A.; Ortiz, G.X., Jr.; Wang, Q.; Malcolmson, S.J.; Chekmenev, E.Y.; Warren, W.S.; et al. Generalizing, Extending, and Maximizing Nitrogen-15 Hyperpolarization Induced by Parahydrogen in Reversible Exchange. J. Phys. Chem. C 2017, 121, 6626–6634. [Google Scholar] [CrossRef] [PubMed]
- Fekete, M.; Ahwal, F.; Duckett, S.B. Remarkable Levels of (15)N Polarization Delivered through SABRE into Unlabeled Pyridine, Pyrazine, or Metronidazole Enable Single Scan NMR Quantification at the mM Level. J. Phys. Chem. B 2020, 124, 4573–4580. [Google Scholar] [CrossRef] [PubMed]
- Barskiy, D.A.; Shchepin, R.V.; Coffey, A.M.; Theis, T.; Warren, W.S.; Goodson, B.M.; Chekmenev, E.Y. Over 20% 15N Hyperpolarization in Under One Minute for Metronidazole, an Antibiotic and Hypoxia Probe. J. Am. Chem. Soc. 2016, 138, 8080–8083. [Google Scholar] [CrossRef] [PubMed]
- Theis, T.; Ortiz, G.X.; Logan, A.W.J.; Claytor, K.E.; Feng, Y.; Huhn, W.P.; Blum, V.; Malcolmson, S.J.; Chekmenev, E.Y.; Wang, Q.; et al. Direct and cost-efficient hyperpolarization of long-lived nuclear spin states on universal 15N2-diazirine molecular tags. Sci. Adv. 2016, 2, e1501438. [Google Scholar] [CrossRef] [PubMed]
- Theis, T.; Truong, M.L.; Coffey, A.M.; Shchepin, R.V.; Waddell, K.W.; Shi, F.; Goodson, B.M.; Warren, W.S.; Chekmenev, E.Y. Microtesla SABRE enables 10% nitrogen-15 nuclear spin polarization. J. Am. Chem. Soc. 2015, 137, 1404–1407. [Google Scholar] [CrossRef]
- Cowley, M.J.; Adams, R.W.; Atkinson, K.D.; Cockett, M.C.; Duckett, S.B.; Green, G.G.; Lohman, J.A.; Kerssebaum, R.; Kilgour, D.; Mewis, R.E. Iridium N-heterocyclic carbene complexes as efficient catalysts for magnetization transfer from para-hydrogen. J. Am. Chem. Soc. 2011, 133, 6134–6137. [Google Scholar] [CrossRef]
- Iali, W.; Rayner, P.J.; Duckett, S.B. Using parahydrogen to hyperpolarize amines, amides, carboxylic acids, alcohols, phosphates, and carbonates. Sci. Adv. 2018, 4, eaao6250. [Google Scholar] [CrossRef]
- Kidd, B.E.; Mashni, J.A.; Limbach, M.N.; Shi, F.; Chekmenev, E.Y.; Hou, Y.; Goodson, B.M. Toward Cleavable Metabolic/pH Sensing "Double Agents" Hyperpolarized by NMR Signal Amplification by Reversible Exchange. Chem. Eur. J. 2018, 24, 10641–10645. [Google Scholar] [CrossRef]
- Gemeinhardt, M.E.; Limbach, M.N.; Gebhardt, T.R.; Eriksson, C.W.; Eriksson, S.L.; Lindale, J.R.; Goodson, E.A.; Warren, W.S.; Chekmenev, E.Y.; Goodson, B.M. “Direct” 13C Hyperpolarization of 13C-Acetate by MicroTesla NMR Signal Amplification by Reversible Exchange (SABRE). Angew. Chem. Int. Ed. 2020, 59, 418–423. [Google Scholar] [CrossRef]
- Iali, W.; Roy, S.S.; Tickner, B.J.; Ahwal, F.; Kennerley, A.J.; Duckett, S.B. Hyperpolarising Pyruvate through Signal Amplification by Reversible Exchange (SABRE). Angew. Chem. Int. Ed. 2019, 58, 10271–10275. [Google Scholar] [CrossRef] [PubMed]
- Adelabu, I.; TomHon, P.; Kabir, M.S.H.; Nantogma, S.; Abdulmojeed, M.; Mandzhieva, I.; Ettedgui, J.; Swenson, R.E.; Krishna, M.C.; Theis, T.; et al. Order-Unity 13C Nuclear Polarization of [1-13C]Pyruvate in Seconds and the Interplay of Water and SABRE Enhancement. ChemPhysChem 2022, 23, e202100839. [Google Scholar] [CrossRef] [PubMed]
- Rayner, P.J.; Gillions, J.P.; Hannibal, V.D.; John, R.O.; Duckett, S.B. Hyperpolarisation of weakly binding N-heterocycles using signal amplification by reversible exchange. Chem. Sci. 2021, 12, 5910–5917. [Google Scholar] [CrossRef] [PubMed]
- Salnikov, O.G.; Chukanov, N.V.; Svyatova, A.; Trofimov, I.A.; Kabir, M.S.H.; Gelovani, J.G.; Kovtunov, K.V.; Koptyug, I.V.; Chekmenev, E.Y. 15N NMR Hyperpolarization of Radiosensitizing Antibiotic Nimorazole by Reversible Parahydrogen Exchange in Microtesla Magnetic Fields. Angew. Chem. Int. Ed. 2021, 60, 2406–2413. [Google Scholar] [CrossRef] [PubMed]
- Tickner, B.J.; Semenova, O.; Iali, W.; Rayner, P.J.; Whitwood, A.C.; Duckett, S.B. Optimisation of pyruvate hyperpolarisation using SABRE by tuning the active magnetisation transfer catalyst. Catal. Sci. Tech. 2020, 10, 1343–1355. [Google Scholar] [CrossRef]
- Adelabu, I.; Ettedgui, J.; Joshi, S.M.; Nantogma, S.; Chowdhury, M.R.H.; McBride, S.; Theis, T.; Sabbasani, V.R.; Chandrasekhar, M.; Sail, D.; et al. Rapid 13C Hyperpolarization of the TCA Cycle Intermediate α-Ketoglutarate via SABRE-SHEATH. Anal. Chem. 2022, 94, 13422–13431. [Google Scholar] [CrossRef]
- Chapman, B.; Joalland, B.; Meersman, C.; Ettedgui, J.; Swenson, R.E.; Krishna, M.C.; Nikolaou, P.; Kovtunov, K.V.; Salnikov, O.G.; Koptyug, I.V.; et al. Low-Cost High-Pressure Clinical-Scale 50% Parahydrogen Generator Using Liquid Nitrogen at 77 K. Anal. Chem. 2021, 93, 8476–8483. [Google Scholar] [CrossRef]
- TomHon, P.; Abdulmojeed, M.; Adelabu, I.; Nantogma, S.; Kabir, M.S.H.; Lehmkuhl, S.; Chekmenev, E.Y.; Theis, T. Temperature Cycling Enables Efficient 13C SABRE-SHEATH Hyperpolarization and Imaging of [1-13C]-Pyruvate. J. Am. Chem. Soc. 2022, 144, 282–287. [Google Scholar] [CrossRef]
- Hyde, J.E. Targeting purine and pyrimidine metabolism in human apicomplexan parasites. Curr. Drug Targets 2007, 8, 31–47. [Google Scholar] [CrossRef]
- Díaz-Muñoz, M.; Hernández-Muñoz, R.; Butanda-Ochoa, A. Structure-activity features of purines and their receptors: Implications in cell physiopathology. Mol. Biomed. 2022, 3, 5. [Google Scholar] [CrossRef]
- Moffatt, B.A.; Ashihara, H. Purine and pyrimidine nucleotide synthesis and metabolism. Arab. Book 2002, 1, e0018. [Google Scholar] [CrossRef] [PubMed]
- Watson, J.D.; Crick, F.H.C. Molecular Structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid. Nature 1953, 171, 737–738. [Google Scholar] [CrossRef] [PubMed]
- Irobalieva, R.N.; Fogg, J.M.; Catanese, D.J., Jr.; Sutthibutpong, T.; Chen, M.; Barker, A.K.; Ludtke, S.J.; Harris, S.A.; Schmid, M.F.; Chiu, W.; et al. Structural diversity of supercoiled DNA. Nat. Commun. 2015, 6, 8851. [Google Scholar] [CrossRef]
- Bannister, A.J.; Kouzarides, T. Regulation of chromatin by histone modifications. Cell Res. 2011, 21, 381–395. [Google Scholar] [CrossRef] [PubMed]
- Jaenisch, R.; Bird, A. Epigenetic regulation of gene expression: How the genome integrates intrinsic and environmental signals. Nat. Genet. 2003, 33, 245. [Google Scholar] [CrossRef]
- Handy, D.E.; Castro, R.; Loscalzo, J. Epigenetic Modifications: Basic Mechanisms and Role in Cardiovascular Disease. Circulation 2011, 123, 2145–2156. [Google Scholar] [CrossRef]
- Li, G.-M. Mechanisms and functions of DNA mismatch repair. Cell Res. 2008, 18, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Gibney, E.R.; Nolan, C.M. Epigenetics and gene expression. Heredity 2010, 105, 4–13. [Google Scholar] [CrossRef] [PubMed]
- Walter, N.G.; Engelke, D.R. Ribozymes: Catalytic RNAs that cut things, make things, and do odd and useful jobs. Biol. (Lond.) 2002, 49, 199–203. [Google Scholar]
- Giuliani, A.L.; Sarti, A.C.; Di Virgilio, F. Extracellular nucleotides and nucleosides as signalling molecules. Immunol. Lett. 2019, 205, 16–24. [Google Scholar] [CrossRef]
- Li, D.; Fedeles, B.I.; Singh, V.; Peng, C.S.; Silvestre, K.J.; Simi, A.K.; Simpson, J.H.; Tokmakoff, A.; Essigmann, J.M. Tautomerism provides a molecular explanation for the mutagenic properties of the anti-HIV nucleoside 5-aza-5,6-dihydro-2′-deoxycytidine. Proc. Natl. Acad. Sci. USA 2014, 111, E3252–E3259. [Google Scholar] [CrossRef] [PubMed]
- Kriukiene, E.; Liutkeviciute, Z.; Klimasauskas, S. 5-Hydroxymethylcytosine-the elusive epigenetic mark in mammalian DNA. Chem. Soc. Rev. 2012, 41, 6916–6930. [Google Scholar] [CrossRef] [PubMed]
- Emran, M.Y.; El-Safty, S.A.; Selim, M.M.; Reda, A.; Morita, H.; Shenashen, M.A. Electrochemical sensors-based phosphorus-doped carbon for determination of adenine DNA-nucleobases in living cells. Carbon 2021, 173, 1093–1104. [Google Scholar] [CrossRef]
- Soflaee, M.H.; Kesavan, R.; Sahu, U.; Tasdogan, A.; Villa, E.; Djabari, Z.; Cai, F.; Tran, D.H.; Vu, H.S.; Ali, E.S.; et al. Purine nucleotide depletion prompts cell migration by stimulating the serine synthesis pathway. Nat. Commun. 2022, 13, 2698. [Google Scholar] [CrossRef] [PubMed]
- Linden, J.; Koch-Nolte, F.; Dahl, G. Purine Release, Metabolism, and Signaling in the Inflammatory Response. Ann. Rev. Immunol. 2019, 37, 325–347. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, H. Extracellular ATP and other nucleotides-ubiquitous triggers of intercellular messenger release. Purinergic Signal. 2016, 12, 25–57. [Google Scholar] [CrossRef] [PubMed]
- Hövelmann, F.; Seitz, O. DNA Stains as Surrogate Nucleobases in Fluorogenic Hybridization Probes. Acc. Chem. Res. 2016, 49, 714–723. [Google Scholar] [CrossRef]
- Hermkens, N.K.; Eshuis, N.; van Weerdenburg, B.J.; Feiters, M.C.; Rutjes, F.P.; Wijmenga, S.S.; Tessari, M. NMR-Based Chemosensing via p-H2 Hyperpolarization: Application to Natural Extracts. Anal. Chem. 2016, 88, 3406–3412. [Google Scholar] [CrossRef]
- Hermkens, N.K.J.; Aspers, R.L.E.G.; Feiters, M.C.; Rutjes, F.P.J.T.; Tessari, M. Trace analysis in water-alcohol mixtures by continuous p-H2 hyperpolarization at high magnetic field. Magn. Reson. Chem. 2018, 56, 633–640. [Google Scholar] [CrossRef]
- Sellies, L.; Reile, I.; Aspers, R.L.E.G.; Feiters, M.C.; Rutjes, F.P.J.T.; Tessari, M. Parahydrogen induced hyperpolarization provides a tool for NMR metabolomics at nanomolar concentrations. Chem. Commun. 2019, 55, 7235–7238. [Google Scholar] [CrossRef]
- Ausmees, K.; Reimets, N.; Reile, I. Understanding Parahydrogen Hyperpolarized Urine Spectra: The Case of Adenosine Derivatives. Molecules 2022, 27, 802. [Google Scholar] [CrossRef] [PubMed]
- Wood, N.J.; Brannigan, J.A.; Duckett, S.B.; Heath, S.L.; Wagstaff, J. Detection of Picomole Amounts of Biological Substrates by para-Hydrogen-Enhanced NMR Methods in Conjunction with a Suitable Receptor Complex. J. Am. Chem. Soc. 2007, 129, 11012–11013. [Google Scholar] [CrossRef] [PubMed]
- Hövener, J.-B.; Schwaderlapp, N.; Lickert, T.; Duckett, S.B.; Mewis, R.E.; Highton, L.A.R.; Kenny, S.M.; Green, G.G.R.; Leibfritz, D.; Korvink, J.G.; et al. A hyperpolarized equilibrium for magnetic resonance. Nat. Commun. 2013, 4, 2946. [Google Scholar] [CrossRef] [PubMed]
- Voet, D.; Rich, A. The Crystal Structures of Purines, Pyrimidines and Their Intermolecular Complexes. In Progress in Nucleic Acid Research and Molecular Biology Davidson; Davidson, J.N., Cohn, W.E., Eds.; Academic Press: Cambridge, MA, USA, 1970; Volume 10, pp. 183–265. [Google Scholar]
- Truong, M.L.; Shi, F.; He, P.; Yuan, B.; Plunkett, K.N.; Coffey, A.M.; Shchepin, R.V.; Barskiy, D.A.; Kovtunov, K.V.; Koptyug, I.V.; et al. Irreversible catalyst activation enables hyperpolarization and water solubility for NMR signal amplification by reversible exchange. J. Phys. Chem. B 2014, 118, 13882–13889. [Google Scholar] [CrossRef]
- Joalland, B.; Nantogma, S.; Chowdhury, M.R.H.; Nikolaou, P.; Chekmenev, E.Y. Magnetic shielding of parahydrogen hyperpolarization experiments for the masses. Magn. Reson. Chem. 2021, 59, 1180–1186. [Google Scholar] [CrossRef]
- Shchepin, R.V.; Jaigirdar, L.; Chekmenev, E.Y. Spin–Lattice Relaxation of Hyperpolarized Metronidazole in Signal Amplification by Reversible Exchange in Micro-Tesla Fields. J. Phys. Chem. C 2018, 122, 4984–4996. [Google Scholar] [CrossRef]
- Birchall, J.R.; Kabir, M.S.H.; Salnikov, O.G.; Chukanov, N.V.; Svyatova, A.; Kovtunov, K.V.; Koptyug, I.V.; Gelovani, J.G.; Goodson, B.M.; Pham, W.; et al. Quantifying the effects of quadrupolar sinks via 15N relaxation dynamics in metronidazoles hyperpolarized via SABRE-SHEATH. Chem. Commun. 2020, 56, 9098–9101. [Google Scholar] [CrossRef]
- Kidd, B.E.; Gesiorski, J.L.; Gemeinhardt, M.E.; Shchepin, R.V.; Kovtunov, K.V.; Koptyug, I.V.; Chekmenev, E.Y.; Goodson, B.M. Facile Removal of Homogeneous SABRE Catalysts for Purifying Hyperpolarized Metronidazole, a Potential Hypoxia Sensor. J. Phys. Chem. C 2018, 122, 16848–16852. [Google Scholar] [CrossRef]
- Shchepin, R.V.; Truong, M.L.; Theis, T.; Coffey, A.M.; Shi, F.; Waddell, K.W.; Warren, W.S.; Goodson, B.M.; Chekmenev, E.Y. Hyperpolarization of “Neat” Liquids by NMR Signal Amplification by Reversible Exchange. J. Phys. Chem. Lett. 2015, 6, 1961–1967. [Google Scholar] [CrossRef]
- Moreno, K.X.; Nasr, K.; Milne, M.; Sherry, A.D.; Goux, W.J. Nuclear spin hyperpolarization of the solvent using signal amplification by reversible exchange (SABRE). J. Magn. Reson. 2015, 257, 15–23. [Google Scholar] [CrossRef]
- Shi, F.; He, P.; Best, Q.A.; Groome, K.; Truong, M.L.; Coffey, A.M.; Zimay, G.; Shchepin, R.V.; Waddell, K.W.; Chekmenev, E.Y.; et al. Aqueous NMR Signal Enhancement by Reversible Exchange in a Single Step Using Water-Soluble Catalysts. J. Phys. Chem. C 2016, 120, 12149–12156. [Google Scholar] [CrossRef] [PubMed]
- Iali, W.; Olaru, A.M.; Green, G.G.R.; Duckett, S.B. Achieving High Levels of NMR-Hyperpolarization in Aqueous Media With Minimal Catalyst Contamination Using SABRE. Chem. Eur. J. 2017, 23, 10491–10495. [Google Scholar] [CrossRef] [PubMed]
- Hövener, J.-B.; Schwaderlapp, N.; Borowiak, R.; Lickert, T.; Duckett, S.B.; Mewis, R.E.; Adams, R.W.; Burns, M.J.; Highton, L.A.R.; Green, G.G.R.; et al. Toward Biocompatible Nuclear Hyperpolarization Using Signal Amplification by Reversible Exchange: Quantitative in Situ Spectroscopy and High-Field Imaging. Anal. Chem. 2014, 86, 1767–1774. [Google Scholar] [CrossRef]
- Colell, J.F.P.; Emondts, M.; Logan, A.W.J.; Shen, K.; Bae, J.; Shchepin, R.V.; Ortiz, G.X.; Spannring, P.; Wang, Q.; Malcolmson, S.J.; et al. Direct Hyperpolarization of Nitrogen-15 in Aqueous Media with Parahydrogen in Reversible Exchange. J. Am. Chem. Soc. 2017, 139, 7761–7767. [Google Scholar] [CrossRef]
- Zeng, H.; Xu, J.; McMahon, M.T.; Lohman, J.A.; van Zijl, P.C. Achieving 1% NMR polarization in water in less than 1min using SABRE. J. Magn. Reson. 2014, 246, 119–121. [Google Scholar] [CrossRef]
- Spannring, P.; Reile, I.; Emondts, M.; Schleker, P.P.; Hermkens, N.K.; van der Zwaluw, N.G.; van Weerdenburg, B.J.; Tinnemans, P.; Tessari, M.; Blümich, B.; et al. A New Ir-NHC Catalyst for Signal Amplification by Reversible Exchange in D2O. Chem. Eur. J. 2016, 22, 9277–9282. [Google Scholar] [CrossRef] [PubMed]
- Lin, K.; TomHon, P.; Lehmkuhl, S.; Laasner, R.; Theis, T.; Blum, V. Density Functional Theory Study of Reaction Equilibria in Signal Amplification by Reversible Exchange. Chemphyschem 2021, 22, 1937–1938. [Google Scholar] [CrossRef] [PubMed]
- Olaru, A.M.; Burns, M.J.; Green, G.G.R.; Duckett, S.B. SABRE hyperpolarisation of vitamin B3 as a function of pH. Chem. Sci. 2017, 8, 2257–2266. [Google Scholar] [CrossRef] [PubMed]
- Wong, Y.P.; Wong, K.L.; Kearns, D.R. On the tautomeric states of guanine and cytosine. Biochem. Biophys. Res. Commun. 1972, 49, 1580–1587. [Google Scholar] [CrossRef]
- MacPhail, R.A.; Williams, L.D.; Jones, D.A.; Shaw, B.R. Variable temperature infrared spectroscopy of cytosine-guanine base pairs: Tautomerism versus polarization. J. Biomol. Struct. Dyn. 1992, 9, 881–898. [Google Scholar] [CrossRef]
- Laxer, A.; Major, D.T.; Gottlieb, H.E.; Fischer, B. (15N5)-Labeled Adenine Derivatives: Synthesis and Studies of Tautomerism by 15N NMR Spectroscopy and Theoretical Calculations. J. Org. Chem. 2001, 66, 5463–5481. [Google Scholar] [CrossRef] [PubMed]
- Gonnella, N.C.; Nakanishi, H.; Holtwick, J.B.; Horowitz, D.S.; Kanamori, K.; Leonard, N.J.; Roberts, J.D. Studies of Tautomers and Protonation of Adenine and Its Derivatives by Nitrogen-15 Nuclear Magnetic Resonance Spectroscopy. J. Am. Chem. Soc. 1983, 105, 2050–2055. [Google Scholar] [CrossRef]
- Wishart, D.S.; Bigam, C.G.; Yao, J.; Abildgaard, F.; Dyson, H.J.; Oldfield, E.; Markley, J.L.; Sykes, B.D. 1H, 13C and 15N chemical shift referencing in biomolecular NMR. J. Biomolec. NMR 1995, 6, 135–140. [Google Scholar] [CrossRef] [PubMed]
- Shchepin, R.V.; Barskiy, D.A.; Coffey, A.M.; Feldman, M.A.; Kovtunova, L.M.; Bukhtiyarov, V.I.; Kovtunov, K.V.; Goodson, B.M.; Koptyug, I.V.; Chekmenev, E.Y. Robust Imidazole-15N2 Synthesis for High-Resolution Low-Field (0.05 T) 15N Hyperpolarized NMR Spectroscopy. ChemistrySelect 2017, 2, 4478–4483. [Google Scholar] [CrossRef]
- Barskiy, D.A.; Shchepin, R.V.; Tanner, C.P.N.; Colell, J.F.P.; Goodson, B.M.; Theis, T.; Warren, W.S.; Chekmenev, E.Y. The absence of quadrupolar nuclei facilitates efficient 13C hyperpolarization via reversible exchange with parahydrogen. ChemPhysChem 2017, 18, 1493–1498. [Google Scholar] [CrossRef]
- Li, X.; Lindale, J.R.; Eriksson, S.L.; Warren, W.S. SABRE enhancement with oscillating pulse sequences. Phys. Chem. Chem. Phys. 2022, 24, 16462–16470. [Google Scholar] [CrossRef]









Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).