

Beneficial Proapoptotic Effect of Heterobasidion Annosum Extract in Colorectal Cancer Xenograft Mouse Model
 , ,
, ,  , and
, and 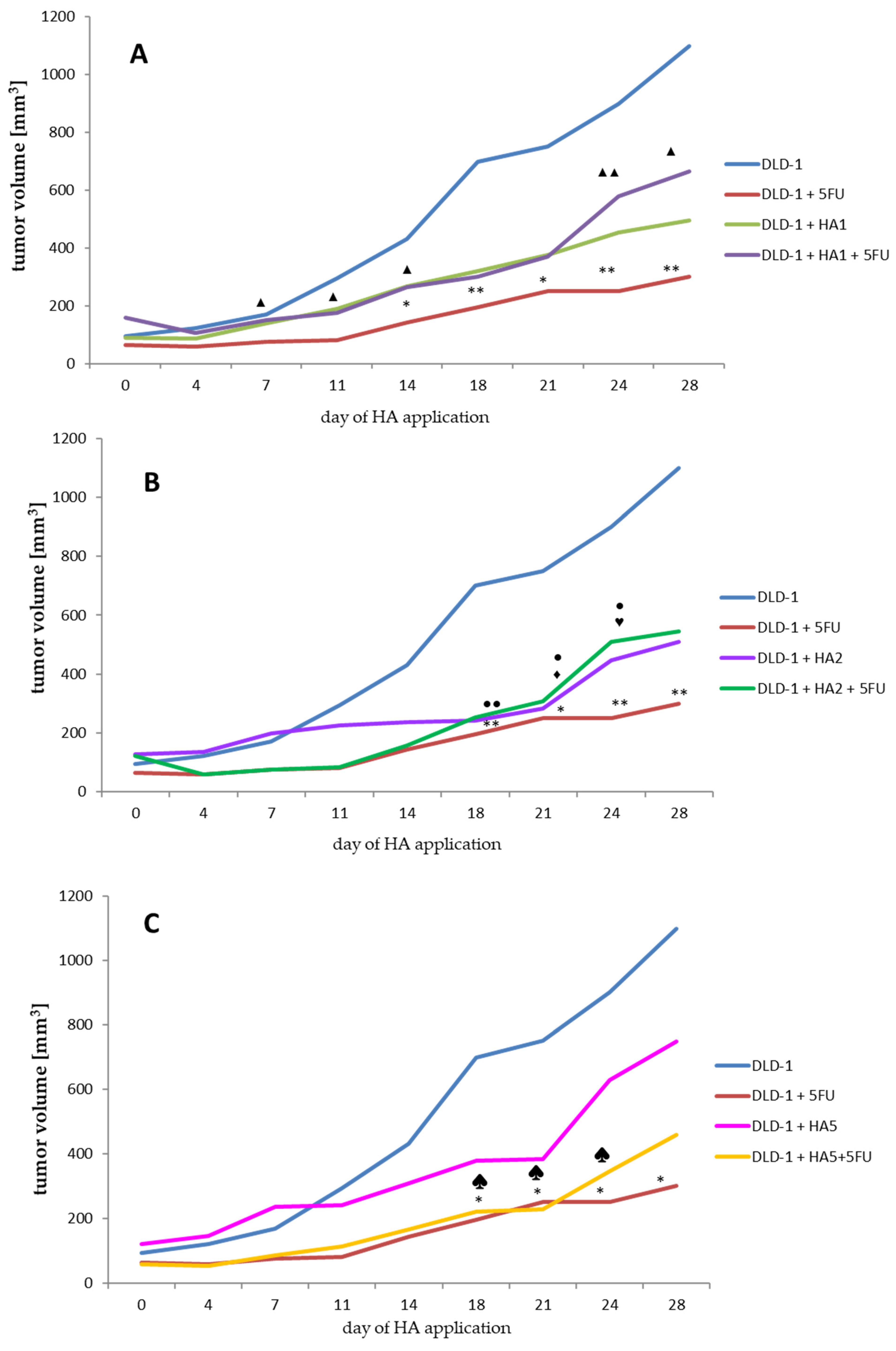{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Evaluation of Tumor Volume
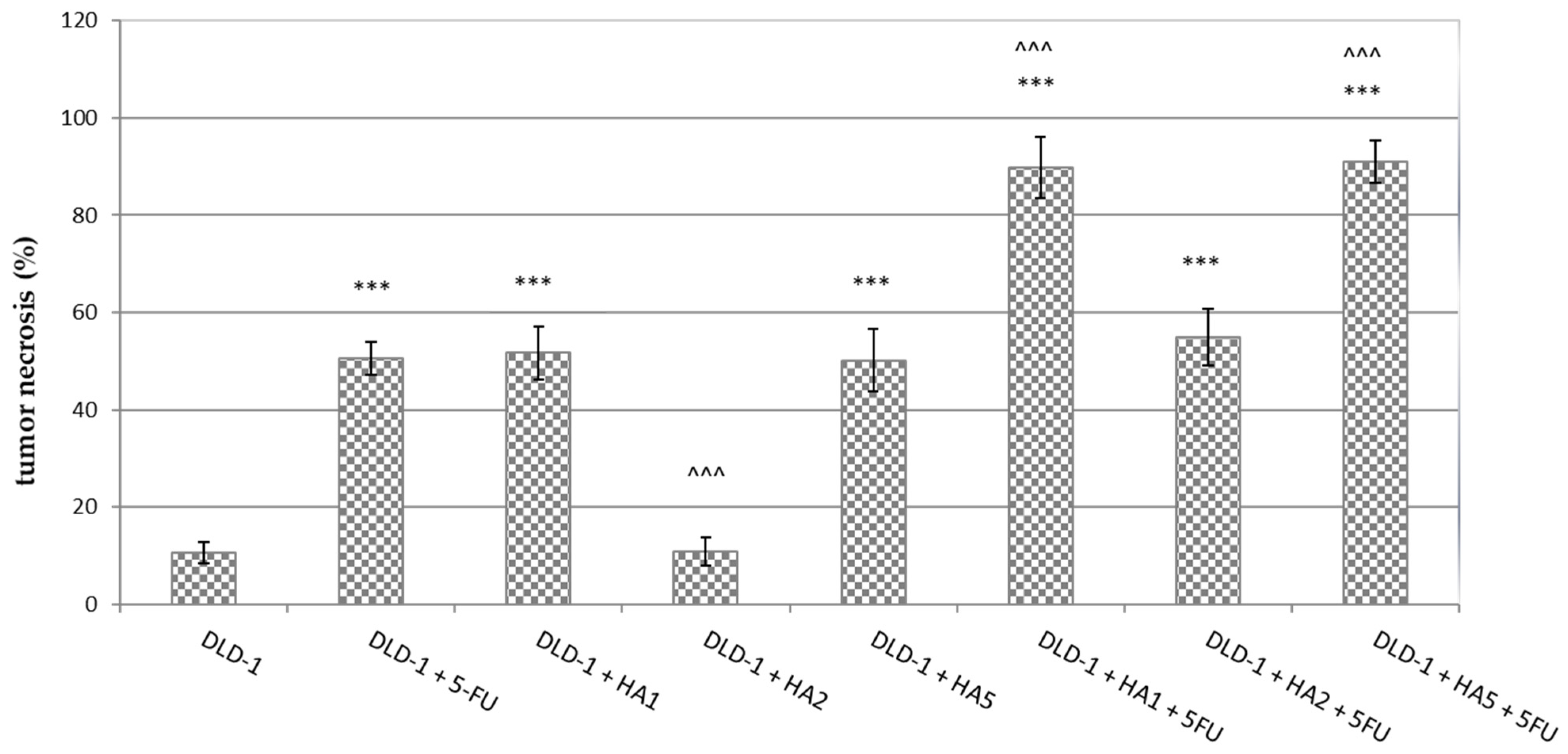
2.2. Histological and Immunohistochemical Examination
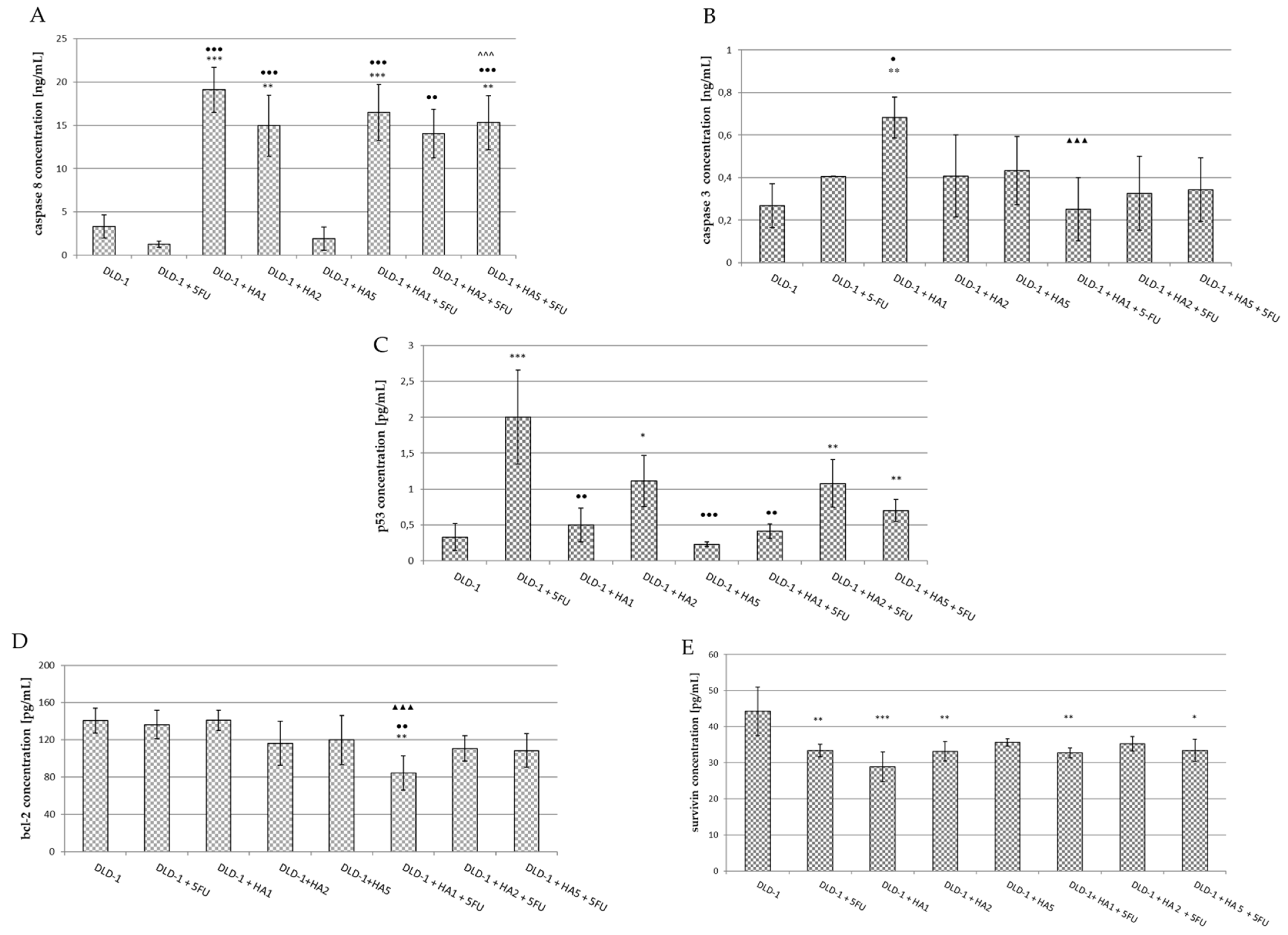
2.3. Parameters of Apoptosis
3. Materials and Methods
3.1. Preparation of HA Extract
3.2. Tissue Culture
3.3. In Vivo Study
Xenograft Studies in Nude Mice
| DLD-1 |
| DLD-1 + 30 mg/kg bw 5-FU (DLD-1 + 5-FU) |
| DLD-1 + 1 mg/kg bw of HA extract (DLD-1 + HA1) |
| DLD-1 + 2 mg/kg bw of HA extract (DLD-1 + HA2) |
| DLD-1 + 5 mg/kg bw of HA extract (DLD-1 + HA5) |
| DLD-1 + 1 mg/kg bw of HA extract + 30 mg/kg bw 5-FU (DLD-1 + HA1 + 5-FU) |
| DLD-1 + 2 mg/kg bw of HA extract + 30 mg/kg bw 5-FU (DLD-1 + HA2 + 5-FU) |
| DLD-1 + 5 mg/kg bw of HA extract + 30 mg/kg bw 5-FU (DLD-1 + HA5 + 5-FU) |
3.4. Immunohistochemistry and H + E Staining
3.5. Immunoenzymatic Assays
3.6. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global Cancer Statistics 2018: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, D.M.; Tooley, K.L.; Butler, R.N.; Smith, C.L.; Geier, M.S.; Howarth, G.S. LyprinolTM Only Partially Improves Indicators of Small Intestinal Integrity in a Rat Model of 5-Fluorouracil-Induced Mucositis. Cancer Biol. Ther. 2008, 7, 295–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Al-Asmari, A.; Al-Zahrani, A.; Khan, A.; Al-Shahrani, H.; Ali Al Amri, M. Taurine Ameliorates 5-Flourouracil-Induced Intestinal Mucositis, Hepatorenal and Reproductive Organ Damage in Wistar Rats: A Biochemical and Histological Study. Hum. Exp. Toxicol. 2016, 35, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Bunz, F.; Hwang, P.M.; Torrance, C.; Waldman, T.; Zhang, Y.; Dillehay, L.; Williams, J.; Lengauer, C.; Kinzler, K.W.; Vogelstein, B. Disruption of P53 in Human Cancer Cells Alters the Responses to Therapeutic Agents. J. Clin. Investig. 1999, 104, 263–269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chaturvedi, V.K.; Agarwal, S.; Gupta, K.K.; Ramteke, P.W.; Singh, M.P. Medicinal Mushroom: Boon for Therapeutic Applications. 3 Biotech 2018, 8, 334. [Google Scholar] [CrossRef]
- Hao, Y.-F.; Jiang, J.-G. Origin and Evolution of China Pharmacopoeia and Its Implication for Traditional Medicines. Mini Rev. Med. Chem. 2015, 15, 595–603. [Google Scholar] [CrossRef]
- Shikov, A.N.; Pozharitskaya, O.N.; Makarov, V.G.; Wagner, H.; Verpoorte, R.; Heinrich, M. Medicinal Plants of the Russian Pharmacopoeia; Their History and Applications. J. Ethnopharmacol. 2014, 154, 481–536. [Google Scholar] [CrossRef] [Green Version]
- Blagodatski, A.; Yatsunskaya, M.; Mikhailova, V.; Tiasto, V.; Kagansky, A.; Katanaev, V.L. Medicinal Mushrooms as an Attractive New Source of Natural Compounds for Future Cancer Therapy. Oncotarget 2018, 9, 29259–29274. [Google Scholar] [CrossRef] [Green Version]
- Simmons, E.G. The International Mycological Association: Its History in Brief with Summaries of Its International Mycological Congresses and Diverse International Relationships. IMA Fungus 2010, 1, 18–100. [Google Scholar] [CrossRef] [Green Version]
- Nowakowski, P.; Markiewicz-Żukowska, R.; Bielecka, J.; Mielcarek, K.; Grabia, M.; Socha, K. Treasures from the Forest: Evaluation of Mushroom Extracts as Anti-Cancer Agents. Biomed. Pharmacother. 2021, 143, 112106. [Google Scholar] [CrossRef]
- Wong, J.H.; Ng, T.B.; Chan, H.H.L.; Liu, Q.; Man, G.C.W.; Zhang, C.Z.; Guan, S.; Ng, C.C.W.; Fang, E.F.; Wang, H.; et al. Mushroom Extracts and Compounds with Suppressive Action on Breast Cancer: Evidence from Studies Using Cultured Cancer Cells, Tumor-Bearing Animals, and Clinical Trials. Appl. Microbiol. Biotechnol. 2020, 104, 4675–4703. [Google Scholar] [CrossRef] [PubMed]
- Piotrowski, J.; Jędrzejewski, T.; Kozak, W. Immunomodulatory and antitumor properties of polysaccharide peptide (PSP). Adv. Hyg. Exp. Med. 2015, 69, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Piri, T.; Vainio, E.J.; Nuorteva, H.; Hantula, J. High Seedling Mortality of Scots Pine Caused by Heterobasidion annosum s.s. Forests 2021, 12, 1289. [Google Scholar] [CrossRef]
- Asiegbu, F.O.; Adomas, A.; Stenlid, J. Conifer Root and Butt Rot Caused by Heterobasidion annosum (Fr.) Bref. s.l. Mol. Plant Pathol. 2005, 6, 395–409. [Google Scholar] [CrossRef]
- Pratt, J.; Thomsen, I.M. An Experiment in Denmark to Infect Wounded Sitka Spruce with the Rotstop Isolate of Phlebiopsis gigantea, and Its Implications for the Control of Heterobasidion Annosum in Britain. Pathogens 2022, 11, 937. [Google Scholar] [CrossRef]
- Liu, M.; Jaber, E.; Zeng, Z.; Kovalchuk, A.; Asiegbu, F.O. Physiochemical and Molecular Features of the Necrotic Lesion in the Heterobasidion—Norway Spruce Pathosystem. Tree Physiol. 2021, 41, 791–800. [Google Scholar] [CrossRef]
- Choi, J.; Lee, G.-W.; Kim, K.-T.; Jeon, J.; Détry, N.; Kuo, H.-C.; Sun, H.; Asiegbu, F.O.; Lee, Y.-H. Comparative Analysis of Genome Sequences of the Conifer Tree Pathogen, Heterobasidion annosum s.s. Genom. Data 2017, 14, 106–113. [Google Scholar] [CrossRef]
- Hansson, D.; Wubshet, S.; Olson, Å.; Karlsson, M.; Staerk, D.; Broberg, A. Secondary Metabolite Comparison of the Species within the Heterobasidion annosum s.l. Complex. Phytochemistry 2014, 108, 243–251. [Google Scholar] [CrossRef]
- Sadowska, A.; Zapora, E.; Sawicka, D.; Niemirowicz-Laskowska, K.; Surażyński, A.; Sułkowska-Ziaja, K.; Kała, K.; Stocki, M.; Wołkowycki, M.; Bakier, S.; et al. Heterobasidion annosum Induces Apoptosis in DLD-1 Cells and Decreases Colon Cancer Growth in In Vivo Model. Int. J. Mol. Sci. 2020, 21, 3447. [Google Scholar] [CrossRef]
- Swinson, D.E.B.; Jones, J.L.; Richardson, D.; Cox, G.; Edwards, J.G.; O’Byrne, K.J. Tumour Necrosis Is an Independent Prognostic Marker in Non-Small Cell Lung Cancer: Correlation with Biological Variables. Lung Cancer 2002, 37, 235–240. [Google Scholar] [CrossRef]
- Langner, C.; Hutterer, G.; Chromecki, T.; Leibl, S.; Rehak, P.; Zigeuner, R. Tumor Necrosis as Prognostic Indicator in Transitional Cell Carcinoma of the Upper Urinary Tract. J. Urol. 2006, 176, 910–914. [Google Scholar] [CrossRef] [PubMed]
- Vanselow, B.; Eble, M.; Rudat, V.; Wollensack, P.; Conradt, C.; Dietz, A. Oxygenation of Advanced Head and Neck Cancer Prognostic Marker for the Response to Primary Radiochemotherapy. Otolaryngol. Head Neck Surg. 2000, 122, 856–862. [Google Scholar] [CrossRef]
- Pollheimer, M.J.; Kornprat, P.; Lindtner, R.A.; Harbaum, L.; Schlemmer, A.; Rehak, P.; Langner, C. Tumor Necrosis Is a New Promising Prognostic Factor in Colorectal Cancer. Hum. Pathol. 2010, 41, 1749–1757. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, S.; Lohse, C.M.; Leibovich, B.C.; Frank, I.; Thompson, R.H.; Webster, W.S.; Zincke, H.; Blute, M.L.; Cheville, J.C.; Kwon, E.D. Histologic Coagulative Tumor Necrosis as a Prognostic Indicator of Renal Cell Carcinoma Aggressiveness. Cancer 2005, 104, 511–520. [Google Scholar] [CrossRef]
- Hiraoka, N.; Ino, Y.; Sekine, S.; Tsuda, H.; Shimada, K.; Kosuge, T.; Zavada, J.; Yoshida, M.; Yamada, K.; Koyama, T.; et al. Tumour Necrosis Is a Postoperative Prognostic Marker for Pancreatic Cancer Patients with a High Interobserver Reproducibility in Histological Evaluation. Br. J. Cancer 2010, 103, 1057–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leek, R.D.; Landers, R.J.; Harris, A.L.; Lewis, C.E. Necrosis Correlates with High Vascular Density and Focal Macrophage Infiltration in Invasive Carcinoma of the Breast. Br. J. Cancer 1999, 79, 991–995. [Google Scholar] [CrossRef] [Green Version]
- Homeyer, A.; Schenk, A.; Arlt, J.; Dahmen, U.; Dirsch, O.; Hahn, H.K. Practical Quantification of Necrosis in Histological Whole-Slide Images. Comput. Med. Imaging Graph. 2013, 37, 313–322. [Google Scholar] [CrossRef]
- Zhang, X.; Chen, L. The Recent Progress of the Mechanism and Regulation of Tumor Necrosis in Colorectal Cancer. J. Cancer Res. Clin. Oncol. 2016, 142, 453–463. [Google Scholar] [CrossRef]
- Papucci, L.; Formigli, L.; Schiavone, N.; Tani, A.; Donnini, M.; Lapucci, A.; Perna, F.; Tempestini, A.; Witort, E.; Morganti, M.; et al. Apoptosis Shifts to Necrosis via Intermediate Types of Cell Death by a Mechanism Depending on C-Myc and Bcl-2 Expression. Cell Tissue Res. 2004, 316, 197–209. [Google Scholar] [CrossRef]
- Skulachev, V.P. Bioenergetic Aspects of Apoptosis, Necrosis and Mitoptosis. Apoptosis 2006, 11, 473–485. [Google Scholar] [CrossRef]
- de Azambuja, E.; Cardoso, F.; de Castro, G.; Colozza, M.; Mano, M.S.; Durbecq, V.; Sotiriou, C.; Larsimont, D.; Piccart-Gebhart, M.J.; Paesmans, M. Ki-67 as Prognostic Marker in Early Breast Cancer: A Meta-Analysis of Published Studies Involving 12 155 Patients. Br. J. Cancer 2007, 96, 1504–1513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, R.L.; Salter, J.; A’Hern, R.; Nerurkar, A.; Parton, M.; Reis-Filho, J.S.; Smith, I.E.; Dowsett, M. The Prognostic Significance of Ki67 before and after Neoadjuvant Chemotherapy in Breast Cancer. Breast Cancer Res. Treat. 2009, 116, 53–68. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.-Y.; Xu, J.; Wang, M.; Zhang, Z.-Z.; Tu, L.; Wang, C.-J.; Lin, T.-L.; Shen, Y.-Y.; Liu, Q.; Cao, H. Prognostic Value of Ki67 Index in Gastrointestinal Stromal Tumors. Int. J. Clin. Exp. Pathol. 2014, 7, 2298–2304. [Google Scholar] [PubMed]
- Yamaguchi, T.; Fujimori, T.; Tomita, S.; Ichikawa, K.; Mitomi, H.; Ohno, K.; Shida, Y.; Kato, H. Clinical Validation of the Gastrointestinal NET Grading System: Ki67 Index Criteria of the WHO 2010 Classification Is Appropriate to Predict Metastasis or Recurrence. Diagn. Pathol. 2013, 8, 65. [Google Scholar] [CrossRef] [Green Version]
- Pollack, A.; DeSilvio, M.; Khor, L.-Y.; Li, R.; Al-Saleem, T.I.; Hammond, M.E.; Venkatesan, V.; Lawton, C.A.; Roach, M.; Shipley, W.U.; et al. Ki-67 Staining Is a Strong Predictor of Distant Metastasis and Mortality for Men with Prostate Cancer Treated with Radiotherapy Plus Androgen Deprivation: Radiation Therapy Oncology Group Trial 92–02. J. Clin. Oncol. 2004, 22, 2133–2140. [Google Scholar] [CrossRef]
- Melling, N.; Kowitz, C.M.; Simon, R.; Bokemeyer, C.; Terracciano, L.; Sauter, G.; Izbicki, J.R.; Marx, A.H. High Ki67 Expression Is an Independent Good Prognostic Marker in Colorectal Cancer. J. Clin. Pathol. 2016, 69, 209–214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Xiao, Z.-T.; Braciak, T.A.; Ou, Q.-J.; Chen, G.; Oduncu, F.S. Association between Ki67 Index and Clinicopathological Features in Colorectal Cancer. Oncol. Res. Treat. 2016, 39, 696–702. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Wang, M.; Zhou, L.; Feng, X.; Cheng, J.; Yu, Y.; Gong, Y.; Zhu, Y.; Li, C.; Tian, L.; et al. Increased HMGB1 and Cleaved Caspase-3 Stimulate the Proliferation of Tumor Cells and Are Correlated with the Poor Prognosis in Colorectal Cancer. J. Exp. Clin. Cancer Res. 2015, 34, 51. [Google Scholar] [CrossRef] [Green Version]
- Salminen, E. Increased Proliferation Activity Measured by Immunoreactive Ki67 Is Associated with Survival Improvement in Rectal/Recto Sigmoid Cancer. World J. Gastroenterol. 2005, 11, 3245. [Google Scholar] [CrossRef]
- Reimers, M.S.; Zeestraten, E.C.M.; van Alphen, T.C.; Dekker, J.W.T.; Putter, H.; Saadatmand, S.; Liefers, G.J.; van de Velde, C.J.H.; Kuppen, P.J.K. Combined Analysis of Biomarkers of Proliferation and Apoptosis in Colon Cancer: An Immunohistochemistry-Based Study Using Tissue Microarray. Int. J. Color. Dis. 2014, 29, 1043–1052. [Google Scholar] [CrossRef]
- Li, K.; Na, K.; Sang, T.; Wu, K.; Wang, Y.; Wang, X. The Ethanol Extracts of Sporoderm-Broken Spores of Ganoderma Lucidum Inhibit Colorectal Cancer in Vitro and in Vivo. Oncol. Rep. 2017, 38, 2803–2813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jedinak, A.; Dudhgaonkar, S.; Jiang, J.; Sandusky, G.; Sliva, D. Pleurotus Ostreatus Inhibits Colitis-Related Colon Carcinogenesis in Mice. Int. J. Mol. Med. 2010, 26, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Sawicka, D.; Hryniewicka, A.; Gohal, S.; Sadowska, A.; Pryczynicz, A.; Guzińska-Ustymowicz, K.; Sokołowska, E.; Morzycki, J.W.; Car, H. Establishment of In Vitro and In Vivo Anticolorectal Cancer Efficacy of Lithocholic Acid-Based Imidazolium Salts. Int. J. Mol. Sci. 2022, 23, 7019. [Google Scholar] [CrossRef]
- Richards, C.H.; Roxburgh, C.S.D.; Anderson, J.H.; McKee, R.F.; Foulis, A.K.; Horgan, P.G.; McMillan, D.C. Prognostic Value of Tumour Necrosis and Host Inflammatory Responses in Colorectal Cancer. Br. J. Surg. 2012, 99, 287–294. [Google Scholar] [CrossRef] [PubMed]
- Kruidering, M.; Evan, G. Caspase-8 in Apoptosis: The Beginning of “The End”? IUBMB Life 2000, 50, 85–90. [Google Scholar] [CrossRef]
- Mishra, N.C.; Kumar, S. Apoptosis: A Mitochondrial Perspective on Cell Death. Indian J. Exp. Biol. 2005, 43, 25–34. [Google Scholar]
- Wong, R.S. Apoptosis in Cancer: From Pathogenesis to Treatment. J. Exp. Clin. Cancer Res. 2011, 30, 87. [Google Scholar] [CrossRef] [Green Version]
- Hector, S.; Prehn, J.H.M. Apoptosis Signaling Proteins as Prognostic Biomarkers in Colorectal Cancer: A Review. Biochim. Biophys. Acta Rev. Cancer 2009, 1795, 117–129. [Google Scholar] [CrossRef]
- Igney, F.H.; Krammer, P.H. Death and Anti-Death: Tumour Resistance to Apoptosis. Nat. Rev. Cancer 2002, 2, 277–288. [Google Scholar] [CrossRef]
- Locksley, R.M.; Killeen, N.; Lenardo, M.J. The TNF and TNF Receptor Superfamilies. Cell 2001, 104, 487–501. [Google Scholar] [CrossRef] [Green Version]
- Bijnsdorp, I.V.; Peters, G.J.; Temmink, O.H.; Fukushima, M.; Kruyt, F.A. Differential Activation of Cell Death and Autophagy Results in an Increased Cytotoxic Potential for Trifluorothymidine Compared to 5-Fluorouracil in Colon Cancer Cells. Int. J. Cancer 2010, 126, 2457–2468. [Google Scholar] [CrossRef] [PubMed]
- Saelens, X.; Festjens, N.; Walle, L.V.; van Gurp, M.; van Loo, G.; Vandenabeele, P. Toxic Proteins Released from Mitochondria in Cell Death. Oncogene 2004, 23, 2861–2874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambrosini, G.; Adida, C.; Altieri, D.C. A Novel Anti-Apoptosis Gene, Survivin, Expressed in Cancer and Lymphoma. Nat. Med. 1997, 3, 917–921. [Google Scholar] [CrossRef] [PubMed]
- Salvesen, G.S.; Walsh, C.M. Functions of Caspase 8: The Identified and the Mysterious. Semin. Immunol. 2014, 26, 246–252. [Google Scholar] [CrossRef] [Green Version]
- Putt, K.S.; Chen, G.W.; Pearson, J.M.; Sandhorst, J.S.; Hoagland, M.S.; Kwon, J.-T.; Hwang, S.-K.; Jin, H.; Churchwell, M.I.; Cho, M.-H.; et al. Small-Molecule Activation of Procaspase-3 to Caspase-3 as a Personalized Anticancer Strategy. Nat. Chem. Biol. 2006, 2, 543–550. [Google Scholar] [CrossRef]
- Peterson, Q.P.; Hsu, D.C.; Novotny, C.J.; West, D.C.; Kim, D.; Schmit, J.M.; Dirikolu, L.; Hergenrother, P.J.; Fan, T.M. Discovery and Canine Preclinical Assessment of a Nontoxic Procaspase-3–Activating Compound. Cancer Res. 2010, 70, 7232–7241. [Google Scholar] [CrossRef] [Green Version]
- Flanagan, L.; Meyer, M.; Fay, J.; Curry, S.; Bacon, O.; Duessmann, H.; John, K.; Boland, K.C.; McNamara, D.A.; Kay, E.W.; et al. Low Levels of Caspase-3 Predict Favourable Response to 5FU-Based Chemotherapy in Advanced Colorectal Cancer: Caspase-3 Inhibition as a Therapeutic Approach. Cell Death Dis. 2016, 7, e2087. [Google Scholar] [CrossRef] [Green Version]
- Mhaidat, N.M.; Bouklihacene, M.; Thorne, R.F. 5-Fluorouracil-Induced Apoptosis in Colorectal Cancer Cells Is Caspase-9-Dependent and Mediated by Activation of Protein Kinase C-δ. Oncol. Lett. 2014, 8, 699–704. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wu, D.; Luo, K.; Wu, S.; Wu, P. Andrographolide Enhances 5-Fluorouracil-Induced Apoptosis via Caspase-8-Dependent Mitochondrial Pathway Involving P53 Participation in Hepatocellular Carcinoma (SMMC-7721) Cells. Cancer Lett. 2009, 276, 180–188. [Google Scholar] [CrossRef]
- Vousden, K.H. P53. Cell 2000, 103, 691–694. [Google Scholar] [CrossRef] [Green Version]
- Lane, D.P. P53, Guardian of the Genome. Nature 1992, 358, 15–16. [Google Scholar] [CrossRef] [PubMed]
- Longley, D.B.; Latif, T.; Boyer, J.; Allen, W.L.; Maxwell, P.J.; Johnston, P.G. The Interaction of Thymidylate Synthase Expression with P53-Regulated Signaling Pathways in Tumor Cells. Semin. Oncol. 2003, 30, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Hafner, A.; Bulyk, M.L.; Jambhekar, A.; Lahav, G. The Multiple Mechanisms That Regulate P53 Activity and Cell Fate. Nat. Rev. Mol. Cell Biol. 2019, 20, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.-H.; Ro, E.J.; Yoon, J.-S.; Mizutani, T.; Kang, D.-W.; Park, J.-C.; Il Kim, T.; Clevers, H.; Choi, K.-Y. 5-FU Promotes Stemness of Colorectal Cancer via P53-Mediated WNT/β-Catenin Pathway Activation. Nat. Commun. 2020, 11, 5321. [Google Scholar] [CrossRef] [PubMed]
- Russo, A.; Esposito, D.; Catillo, M.; Pietropaolo, C.; Crescenzi, E.; Russo, G. Human RpL3 Induces G₁/S Arrest or Apoptosis by Modulating P21 (Waf1/Cip1) Levels in a P53-Independent Manner. Cell Cycle 2013, 12, 76–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, A.; Pagliara, V.; Albano, F.; Esposito, D.; Sagar, V.; Loreni, F.; Irace, C.; Santamaria, R.; Russo, G. Regulatory Role of RpL3 in Cell Response to Nucleolar Stress Induced by Act D in Tumor Cells Lacking Functional P53. Cell Cycle 2016, 15, 41–51. [Google Scholar] [CrossRef] [Green Version]
- Balogh, G.A.; Mailo, D.; Nardi, H.; Corte, M.M.; Vincent, E.; Barutta, E.; Lizarraga, G.; Lizarraga, P.; Montero, H.; Gentili, R. Serological Levels of Mutated P53 Protein Are Highly Detected at Early Stages in Breast Cancer Patients. Exp. Ther. Med. 2010, 1, 357–361. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.H.; Oh, J.Y.; Ryu, S.K.; Kim, S.J.; Lee, N.Y.; Kim, Y.S.; Yi, S.Y.; Shim, K.S.; Han, W.S. Detection of Epidermal Growth Factor Receptor in the Serum of Gastric Carcinoma Patients. Cancer 1997, 79, 1879–1883. [Google Scholar] [CrossRef]
- Oh, M.J.; Choi, J.H.; Kim, I.H.; Lee, Y.H.; Huh, J.Y.; Park, Y.K.; Lee, K.W.; Chough, S.Y.; Joo, K.S.; Ku, B.S.; et al. Detection of Epidermal Growth Factor Receptor in the Serum of Patients with Cervical Carcinoma. Clin. Cancer Res. 2000, 6, 4760–4763. [Google Scholar]
- Meulendijks, D.; Henricks, L.M.; Jacobs, B.A.W.; Aliev, A.; Deenen, M.J.; de Vries, N.; Rosing, H.; van Werkhoven, E.; de Boer, A.; Beijnen, J.H.; et al. Pretreatment Serum Uracil Concentration as a Predictor of Severe and Fatal Fluoropyrimidine-Associated Toxicity. Br. J. Cancer 2017, 116, 1415–1424. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, Y.; Liu, F. Influence of Pholiota Adiposa on Gut Microbiota and Promote Tumor Cell Apoptosis Properties in H22 Tumor-Bearing Mice. Sci. Rep. 2022, 12, 8589. [Google Scholar] [CrossRef] [PubMed]
- Osaki, M.; Osaki, M.; Tatebe, S.; Goto, A.; Hayashi, H.; Oshimura, M.; Ito, H. 5-Fluorouracil (5-FU) Induced Apoptosis in Gastric Cancer Cell Lines: Role of the P53 Gene. Apoptosis 1997, 2, 221–226. [Google Scholar] [CrossRef] [PubMed]
- Giannoulis, K.; Fountzilas, G.; Angouridakis, N.; Angouridaki, C.; Giannoulis, E.; Gamvros, O. Serum Levels of Bcl-2 in Patients with Colorectal Cancer. Tech. Coloproctol. 2004, 8, s56–s58. [Google Scholar] [CrossRef] [PubMed]
- Dohi, T.; Beltrami, E.; Wall, N.R.; Plescia, J.; Altieri, D.C. Mitochondrial Survivin Inhibits Apoptosis and Promotes Tumorigenesis. J. Clin. Investig. 2004, 114, 1117–1127. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Bennit, H.F.; Wall, N.R. The Emerging Role of Exosomes in Survivin Secretion. Histol. Histopathol. 2015, 30, 43–50. [Google Scholar] [CrossRef]
- Chang, J.-Y.; Cheung, C.H.A.; Huang, C.-C.; Tsai, F.-Y.; Lee, J.Y.-C.; Cheng, S.M.; Chang, Y.-C.; Huang, Y.-C.; Chen, S.-H. Survivin – Biology and Potential as a Therapeutic Target in Oncology. OncoTargets Ther. 2013, 6, 1453–1462. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.E.; Howerth, E.W. Survivin: A Bifunctional Inhibitor of Apoptosis Protein. Vet. Pathol. 2004, 41, 599–607. [Google Scholar] [CrossRef]
- Fernández, J.G.; Rodríguez, D.A.; Valenzuela, M.; Calderon, C.; Urzúa, U.; Munroe, D.; Rosas, C.; Lemus, D.; Díaz, N.; Wright, M.C.; et al. Survivin Expression Promotes VEGF-Induced Tumor Angiogenesis via PI3K/Akt Enhanced β-Catenin/Tcf-Lef Dependent Transcription. Mol. Cancer 2014, 13, 209. [Google Scholar] [CrossRef] [Green Version]
- Gunaldi, M.; Isiksacan, N.; Kocoglu, H.; Okuturlar, Y.; Gunaldi, O.; Topcu, T.; Karabulut, M. The Value of Serum Survivin Level in Early Diagnosis of Cancer. J. Cancer Res. Ther. 2018, 14, 570. [Google Scholar] [CrossRef]
- Dobrzycka, B.; Mackowiak-Matejczyk, B.; Terlikowska, K.M.; Kulesza-Bronczyk, B.; Kinalski, M.; Terlikowski, S.J. Prognostic Significance of Pretreatment VEGF, Survivin, and Smac/DIABLO Serum Levels in Patients with Serous Ovarian Carcinoma. Tumor Biol. 2015, 36, 4157–4165. [Google Scholar] [CrossRef] [Green Version]
- Ahmed, M.B.; Shehata, H.H.; Moussa, M.; Ibrahim, T.M. Prognostic Significance of Survivin and Tumor Necrosis Factor-Alpha in Adult Acute Lymphoblastic Leukemia. Clin. Biochem. 2012, 45, 112–116. [Google Scholar] [CrossRef] [Green Version]
- Yang, L.; Wang, Y.; Zheng, H.; Zhang, D.; Wu, X.; Sun, G.; Yang, T. Low-Dose 5-Fluorouracil Sensitizes HepG2 Cells to TRAIL through TRAIL Receptor DR5 and Survivin-Dependent Mechanisms. J. Chemother. 2017, 29, 179–188. [Google Scholar] [CrossRef]
- Zhang, Y.; Sun, D.; Meng, Q.; Guo, W.; Chen, Q.; Zhang, Y. Grifola Frondosa Polysaccharides Induce Breast Cancer Cell Apoptosis via the Mitochondrial-Dependent Apoptotic Pathway. Int. J. Mol. Med. 2017, 40, 1089–1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, S.-L.; Yu, V.C. Proteins of the BCL-2 Family in Apoptosis Signalling: From Mechanistic Insights to Therapeutic Opportunities. Clin. Exp. Pharm. Physiol. 2004, 31, 119–128. [Google Scholar] [CrossRef]
- Lin, C.-H.; Chang, C.-Y.; Lee, K.-R.; Lin, H.-J.; Lin, W.-C.; Chen, T.-H.; Wan, L. Cold-Water Extracts of Grifola Frondosa and Its Purified Active Fraction Inhibit Hepatocellular Carcinoma in Vitro and in Vivo. Exp. Biol. Med. 2016, 241, 1374–1385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, D.; Zhang, Y.; Lu, J.; Wang, Y.; Wang, J.; Meng, Q.; Lee, R.J.; Wang, D.; Teng, L. Cordycepin, a Natural Antineoplastic Agent, Induces Apoptosis of Breast Cancer Cells via Caspase-Dependent Pathways. Nat. Prod. Commun. 2016, 11, 63–68. [Google Scholar] [CrossRef] [Green Version]
- Tsai, C.-C.; Li, Y.-S.; Lin, P.-P. Inonotus Obliquus Extract Induces Apoptosis in the Human Colorectal Carcinoma’s HCT-116 Cell Line. Biomed. Pharmacother. 2017, 96, 1119–1126. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-C.; Hsieh, Y.-Y.; Lo, H.-L.; Li, A.; Chou, C.-J.; Yang, P.-M. In Vitro and In Silico Mechanistic Insights into MiR-21-5p-Mediated Topoisomerase Drug Resistance in Human Colorectal Cancer Cells. Biomolecules 2019, 9, 467. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadowska, A.; Sawicka, D.; Godlewska, K.; Guzińska-Ustymowicz, K.; Zapora, E.; Sokołowska, E.; Car, H. Beneficial Proapoptotic Effect of Heterobasidion Annosum Extract in Colorectal Cancer Xenograft Mouse Model. Molecules 2023, 28, 1352. https://doi.org/10.3390/molecules28031352
Sadowska A, Sawicka D, Godlewska K, Guzińska-Ustymowicz K, Zapora E, Sokołowska E, Car H. Beneficial Proapoptotic Effect of Heterobasidion Annosum Extract in Colorectal Cancer Xenograft Mouse Model. Molecules. 2023; 28(3):1352. https://doi.org/10.3390/molecules28031352
Chicago/Turabian StyleSadowska, Anna, Diana Sawicka, Katarzyna Godlewska, Katarzyna Guzińska-Ustymowicz, Ewa Zapora, Emilia Sokołowska, and Halina Car. 2023. "Beneficial Proapoptotic Effect of Heterobasidion Annosum Extract in Colorectal Cancer Xenograft Mouse Model" Molecules 28, no. 3: 1352. https://doi.org/10.3390/molecules28031352
APA StyleSadowska, A., Sawicka, D., Godlewska, K., Guzińska-Ustymowicz, K., Zapora, E., Sokołowska, E., & Car, H. (2023). Beneficial Proapoptotic Effect of Heterobasidion Annosum Extract in Colorectal Cancer Xenograft Mouse Model. Molecules, 28(3), 1352. https://doi.org/10.3390/molecules28031352