
Antibacterial Activity and Mechanism of Madecassic Acid against Staphylococcus aureus
Abstract
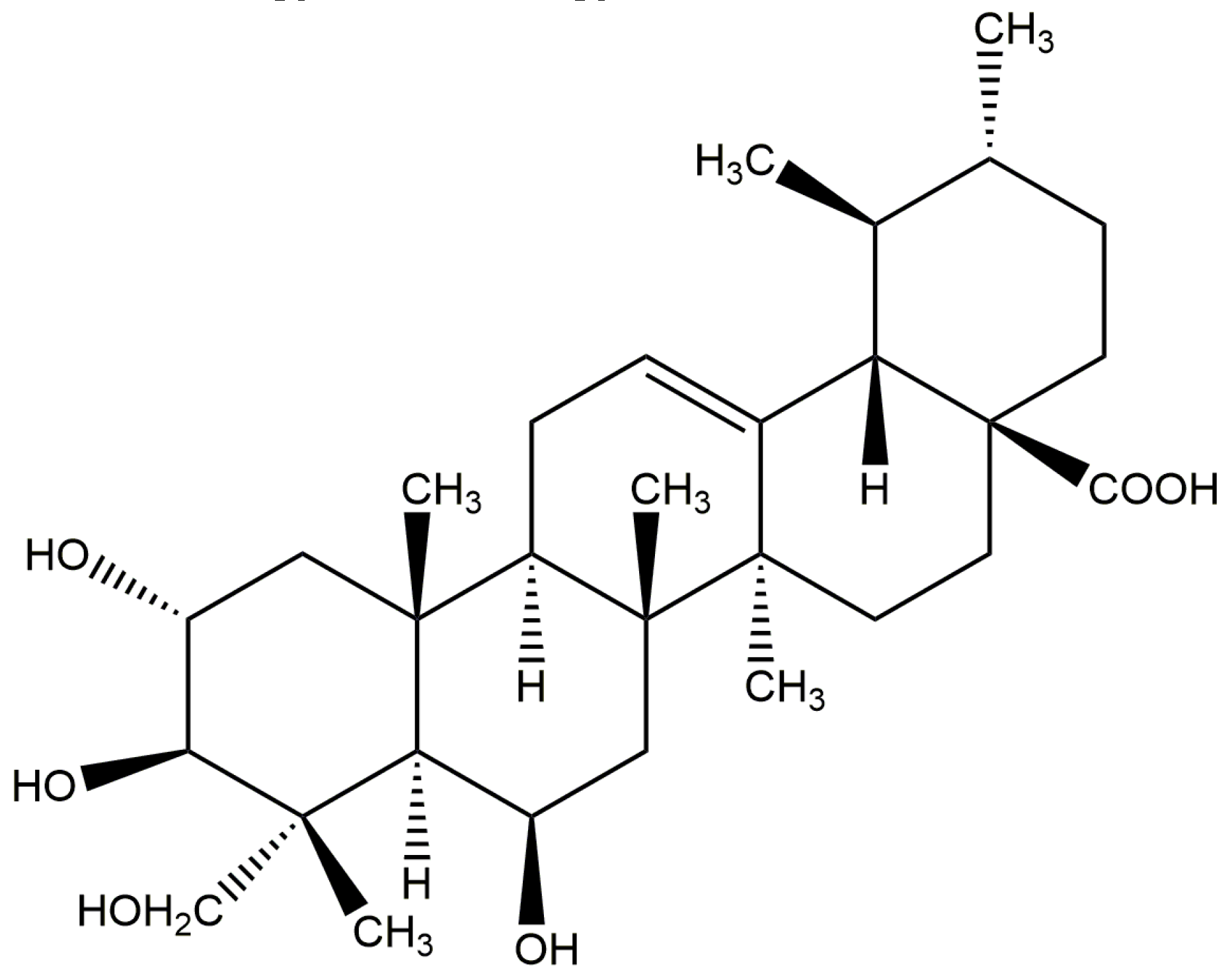
:1. Introduction
2. Results
2.1. Antibacterial Activity of MA
2.1.1. Diameter of Inhibition Zone
2.1.2. Minimum Inhibitory Concentration (MIC)
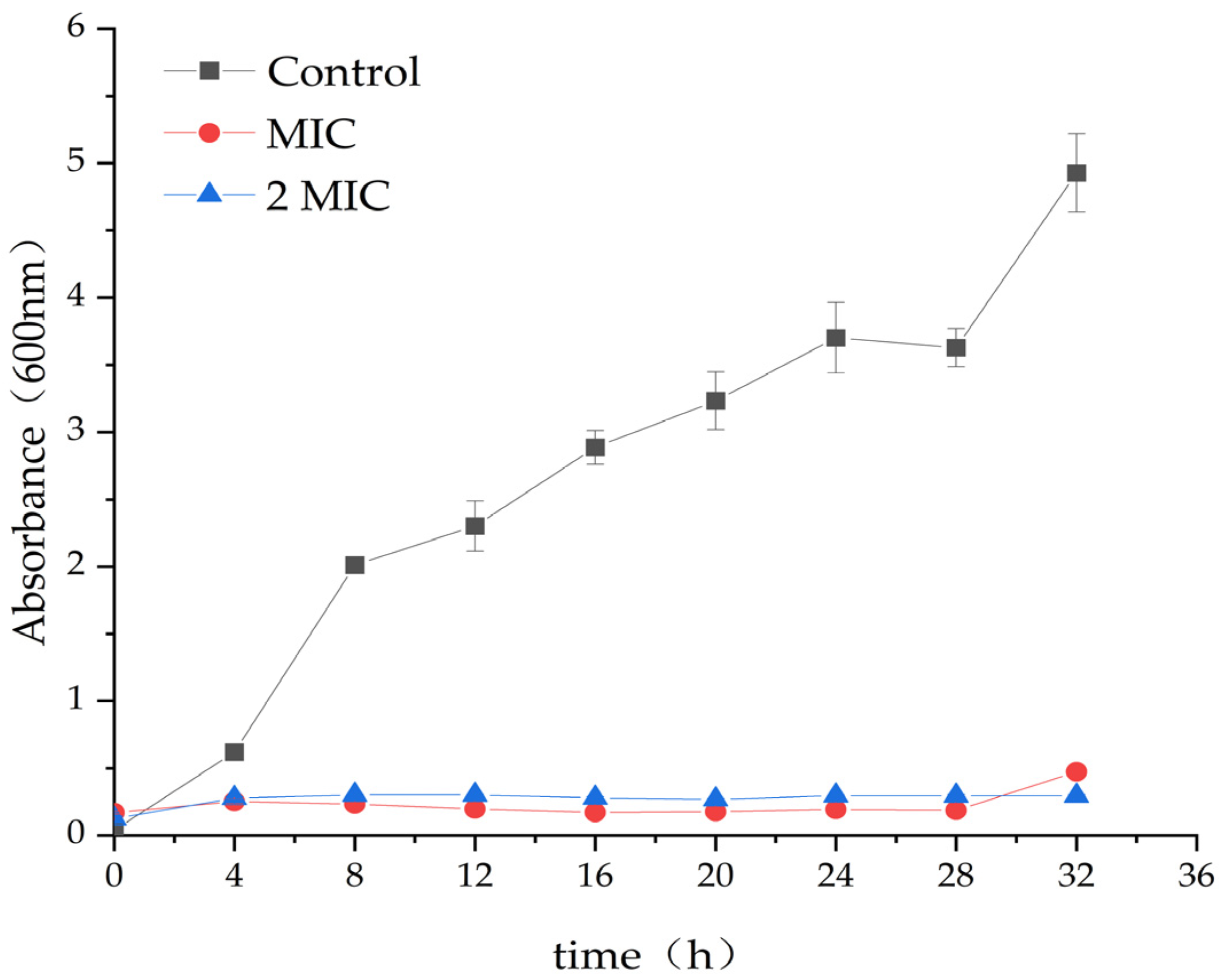
2.2. Antibacterial Effect of MA on S. aureus
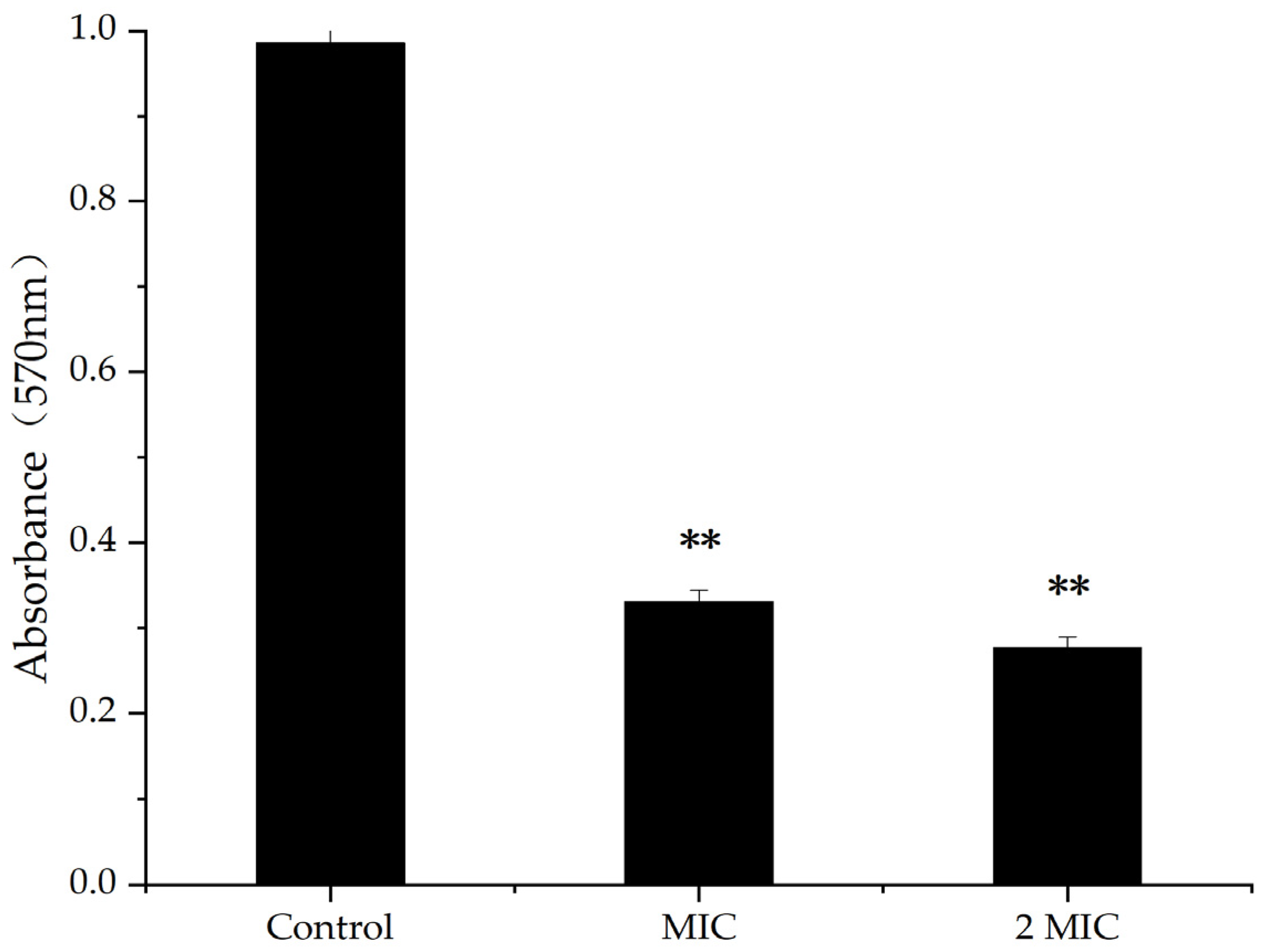
2.3. Effect of MA on Biofilm of S. aureus
2.4. Effect of MA on Cell Membrane and Cell Wall of S. aureus
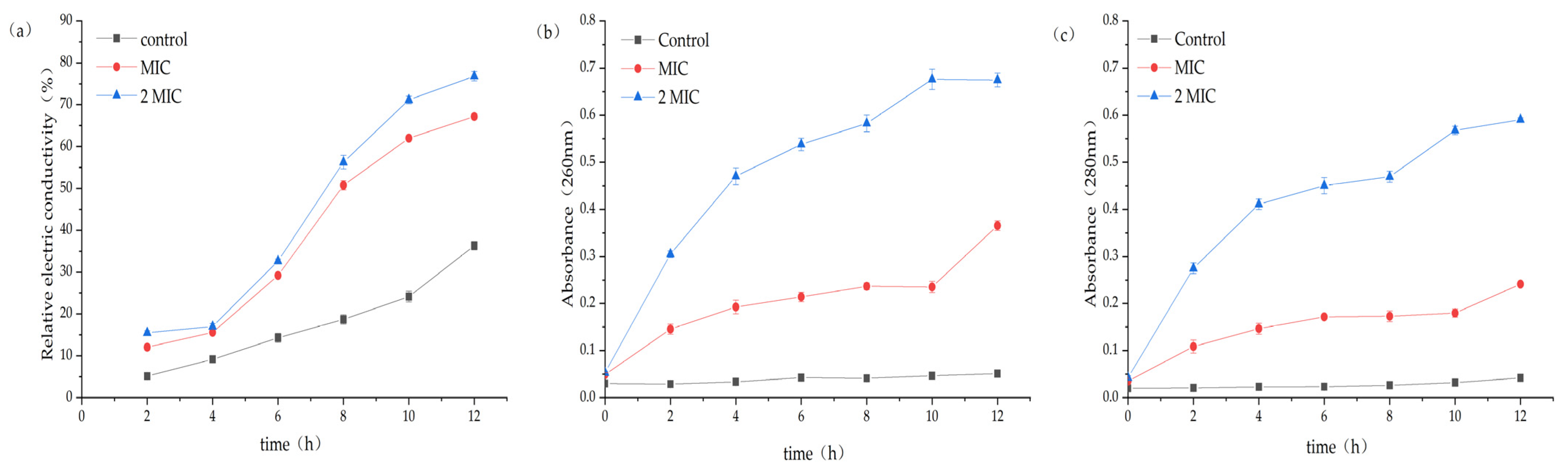
2.4.1. Results of Changes in Conductivity in the Culture Medium after the Intervention of MA on S. aureus
2.4.2. Effect of MA on Macromolecular Substances in Culture Medium of S. aureus
2.4.3. Effect of MA on Cell Wall of S. aureus
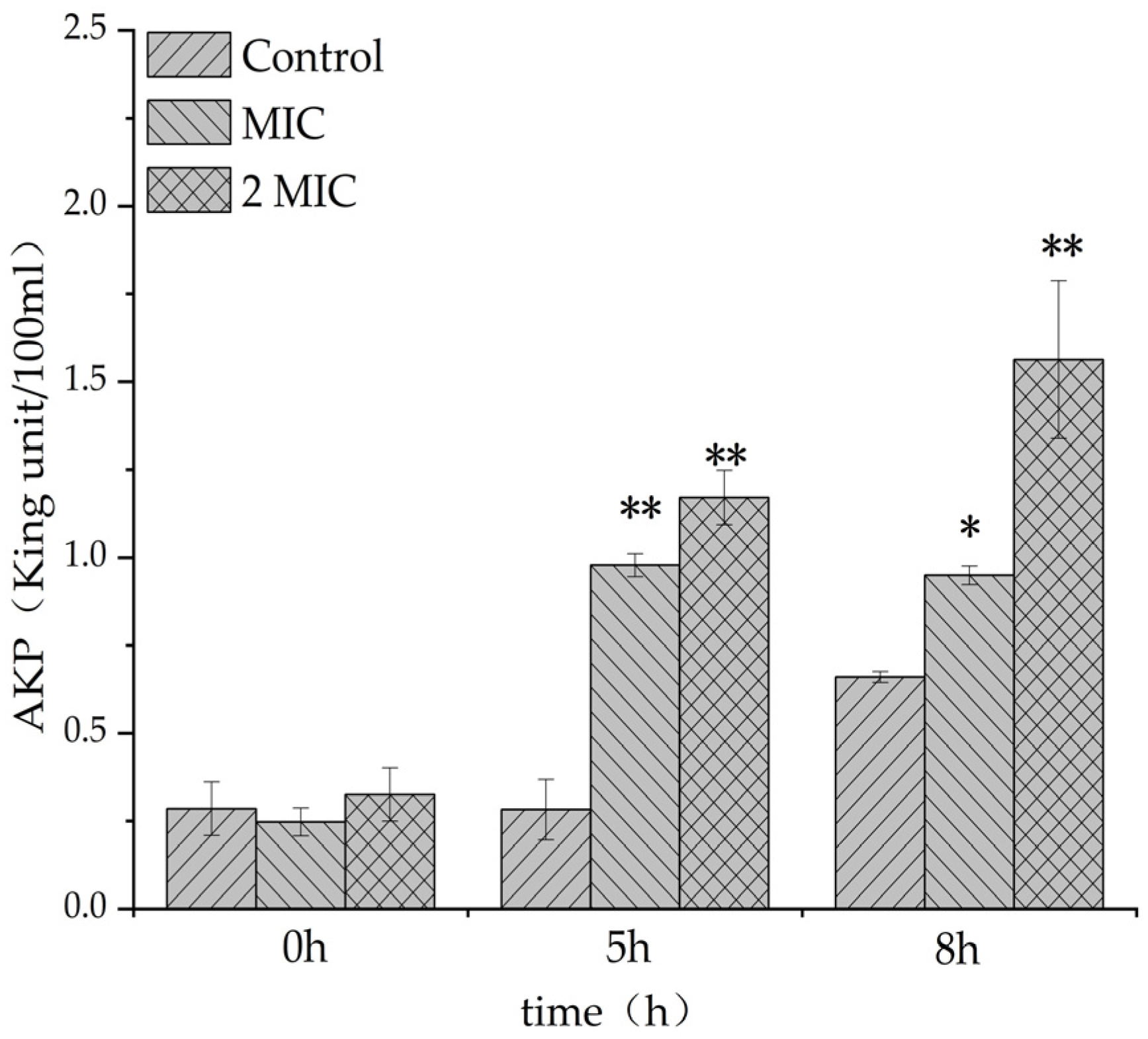
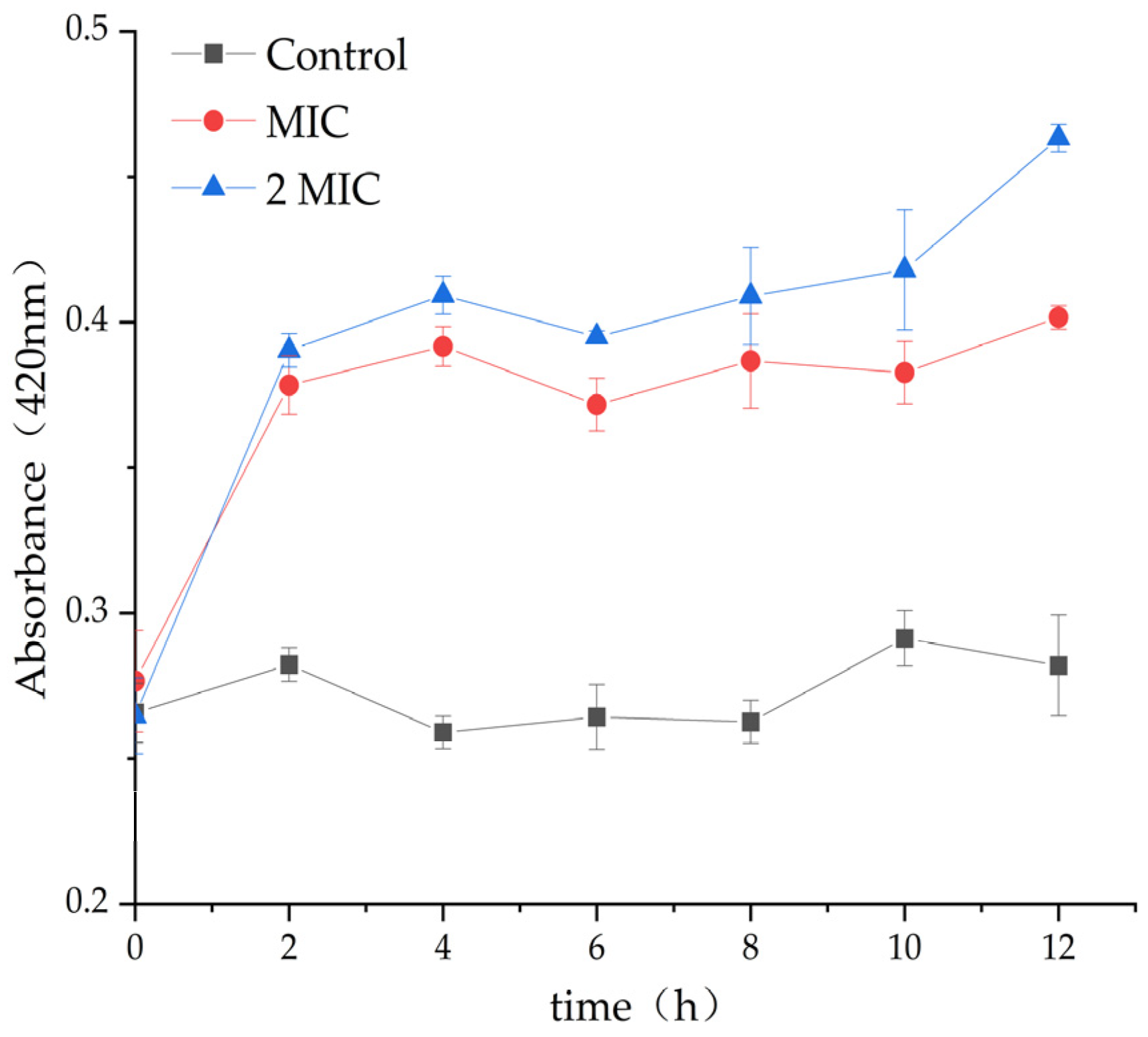
2.4.4. Effect of MA on β-Galactosidase Content
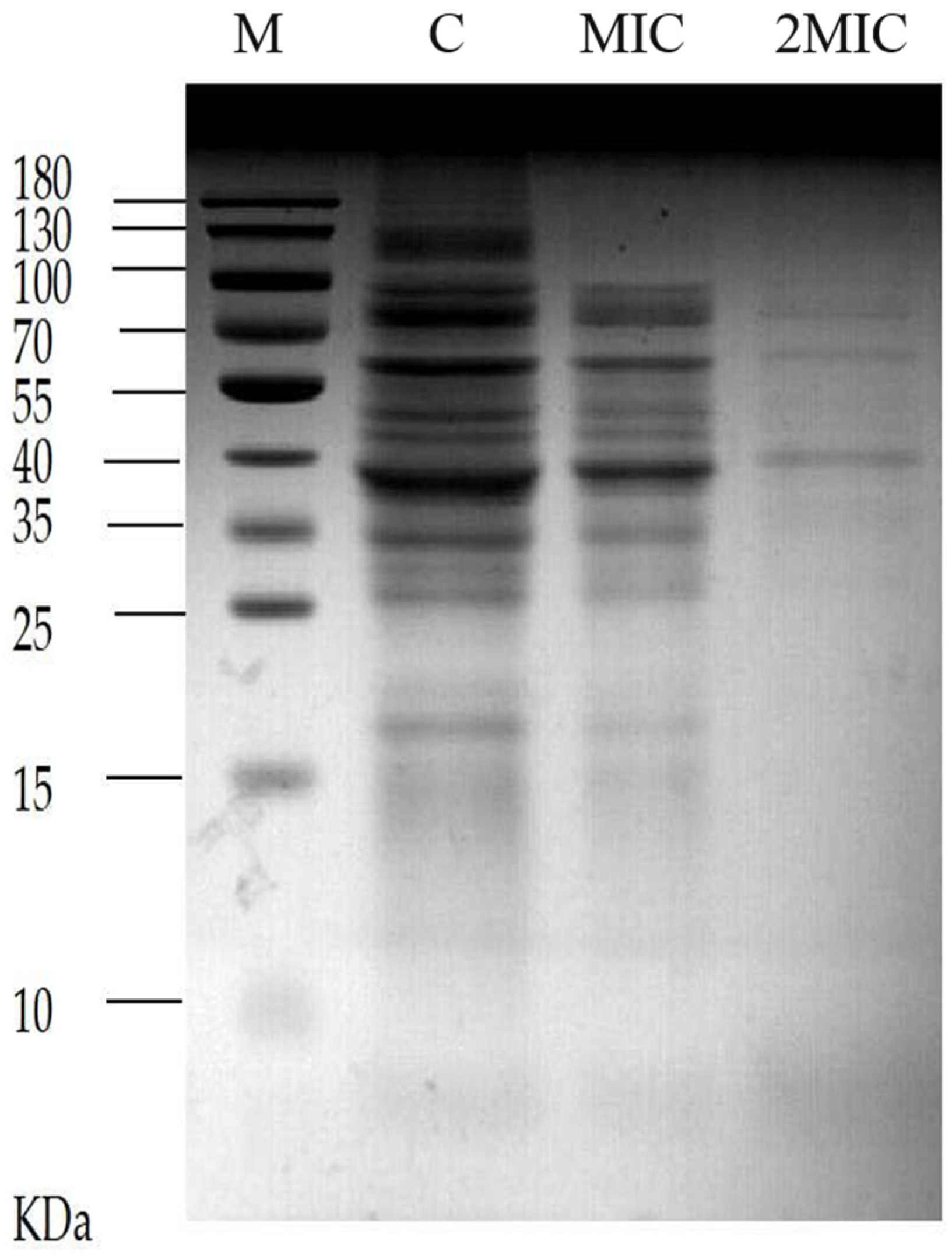
2.5. Effect of MA on Soluble Protein of S. aureus
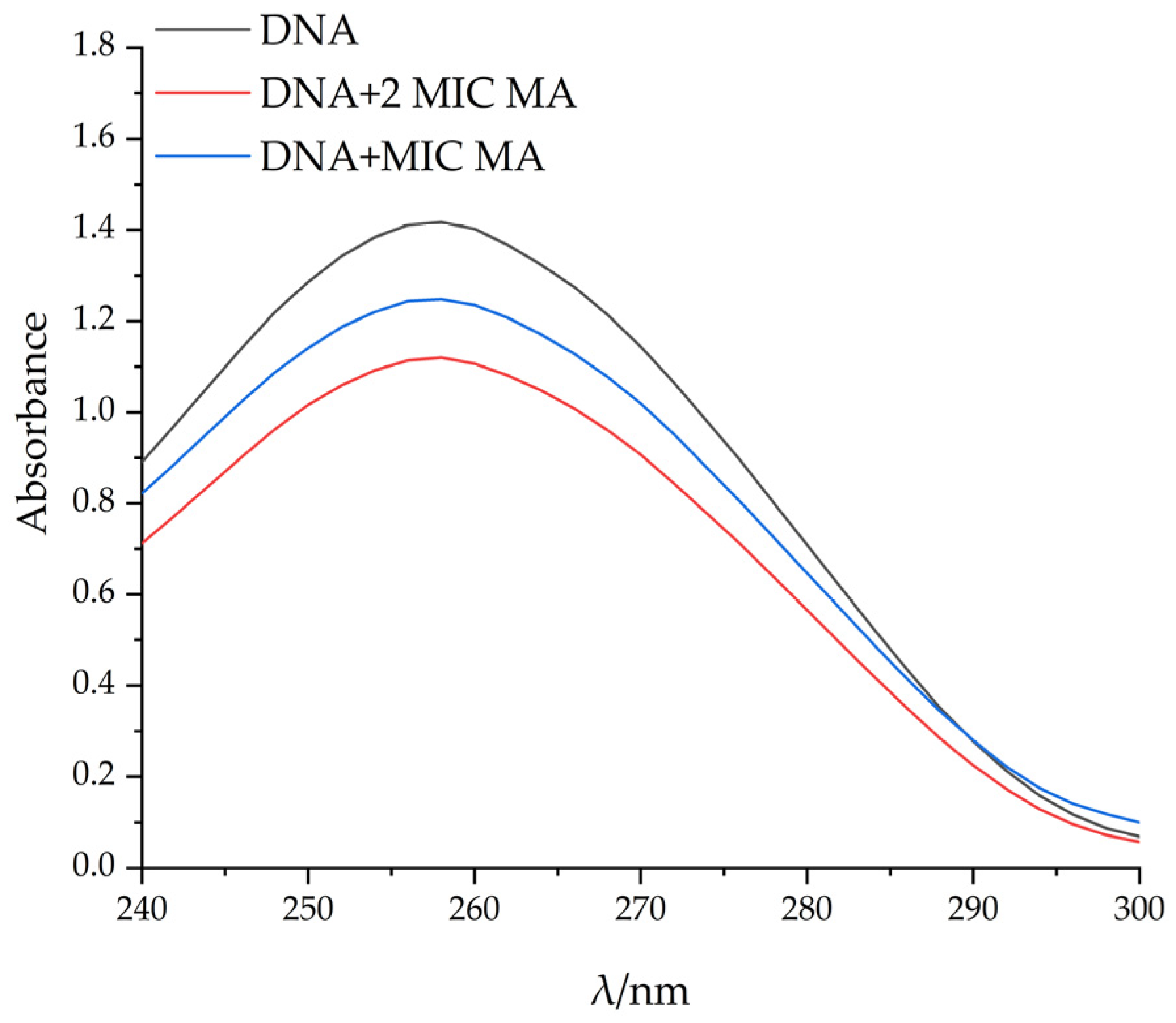
2.6. Study on the Interaction Mode between MA and DNA
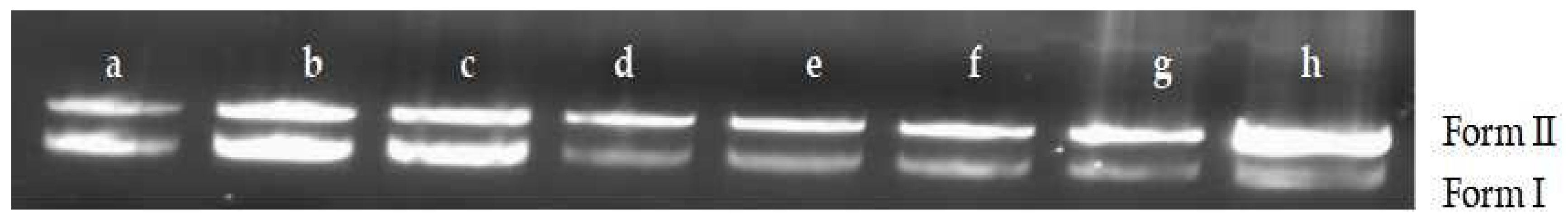
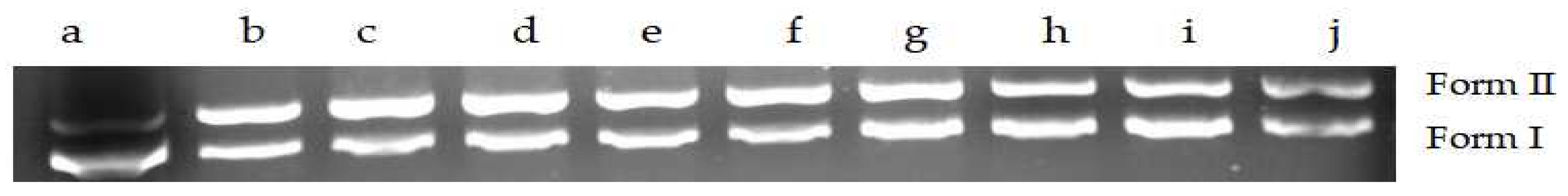
2.6.1. Changes of DNA Structures by Interaction of MA with DNA
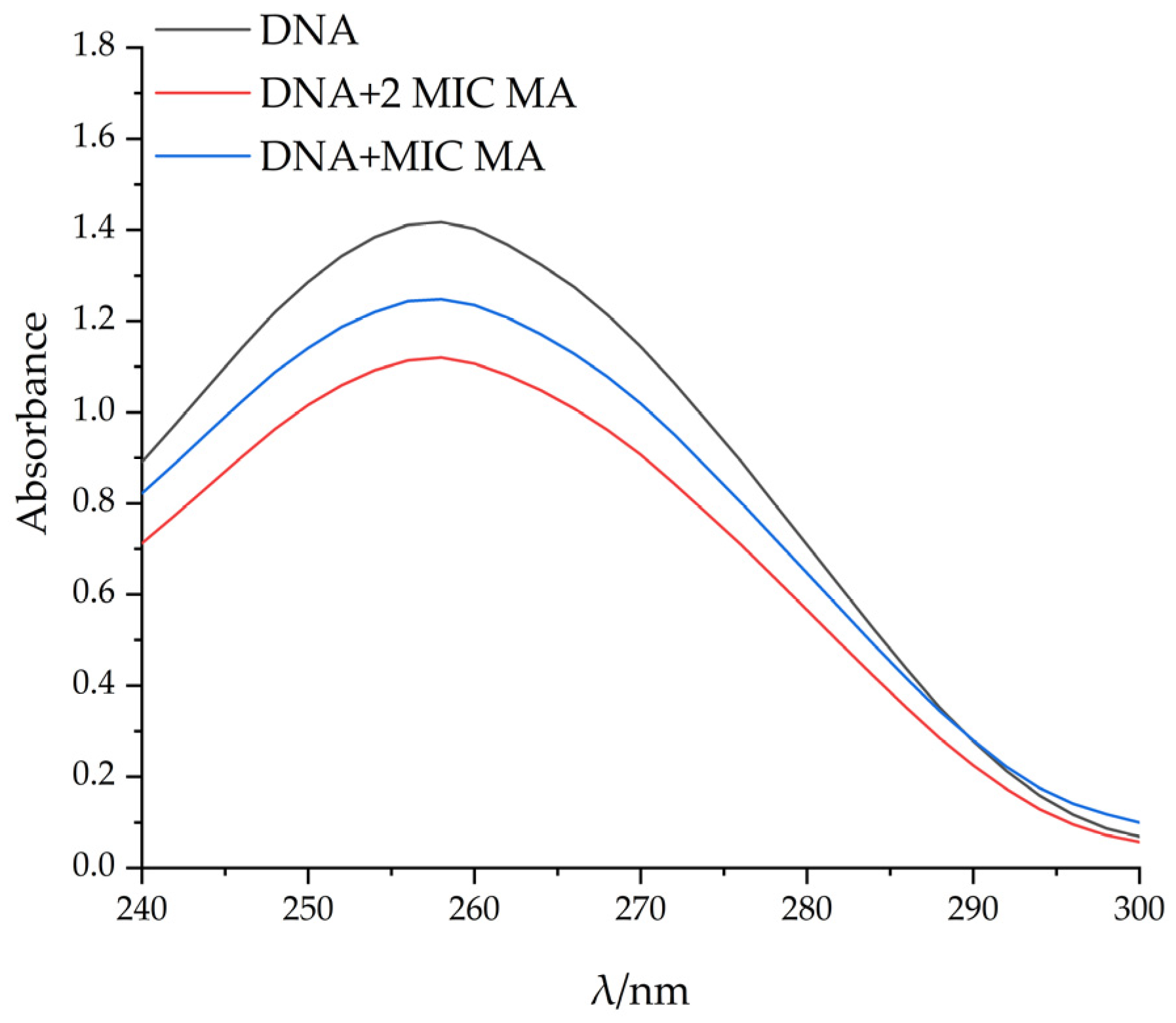
2.6.2. UV Spectral Study on the Binding of MA with DNA
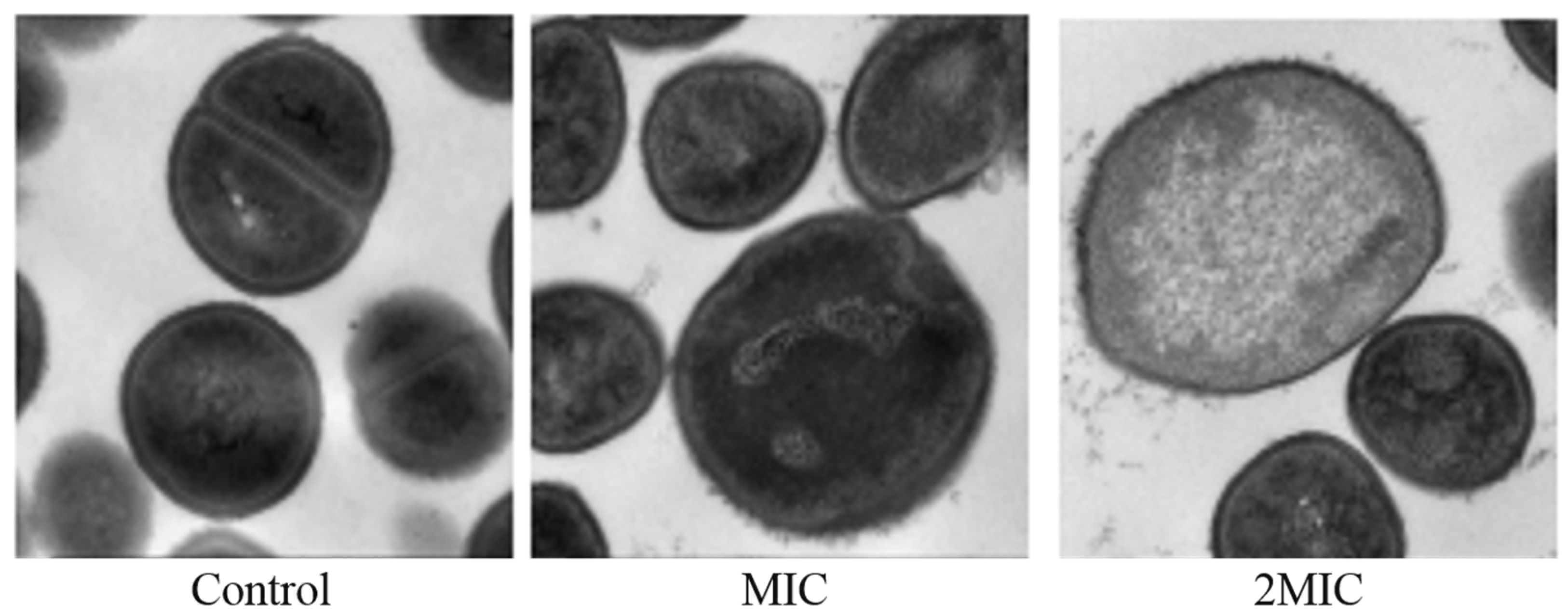
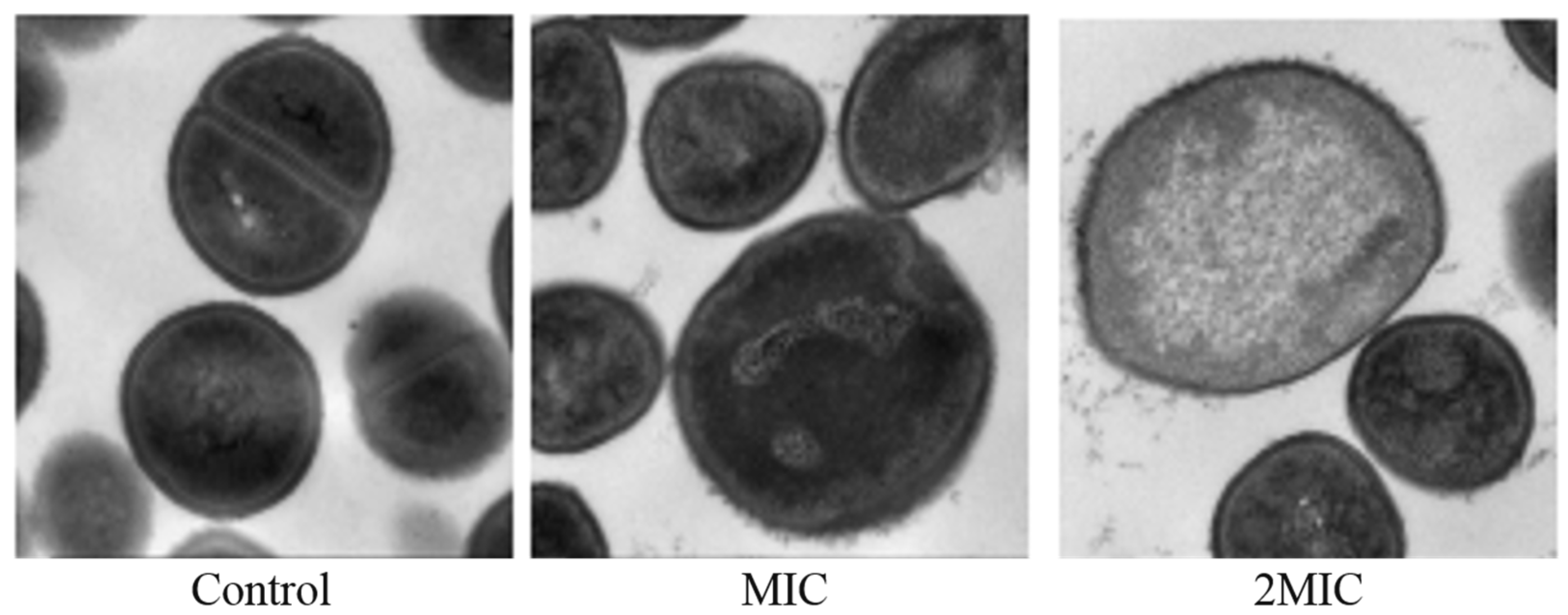
2.7. Effect of MA on the Morphology of S. aureus Cells
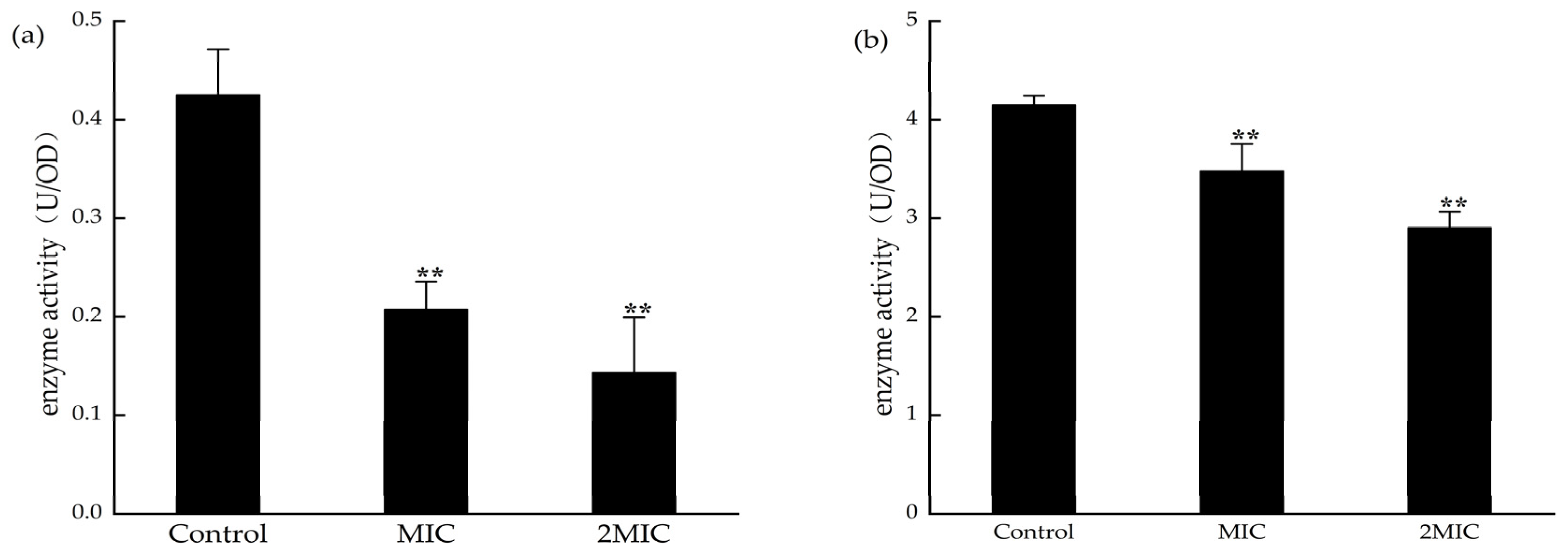
2.8. Effect of MA on the Determination of Malate Dehydrogenase (MDH) and Succinate Dehydrogenase (SDH) Activities
2.9. Effect of MA on DNA Topoisomerase Activity of S. aureus
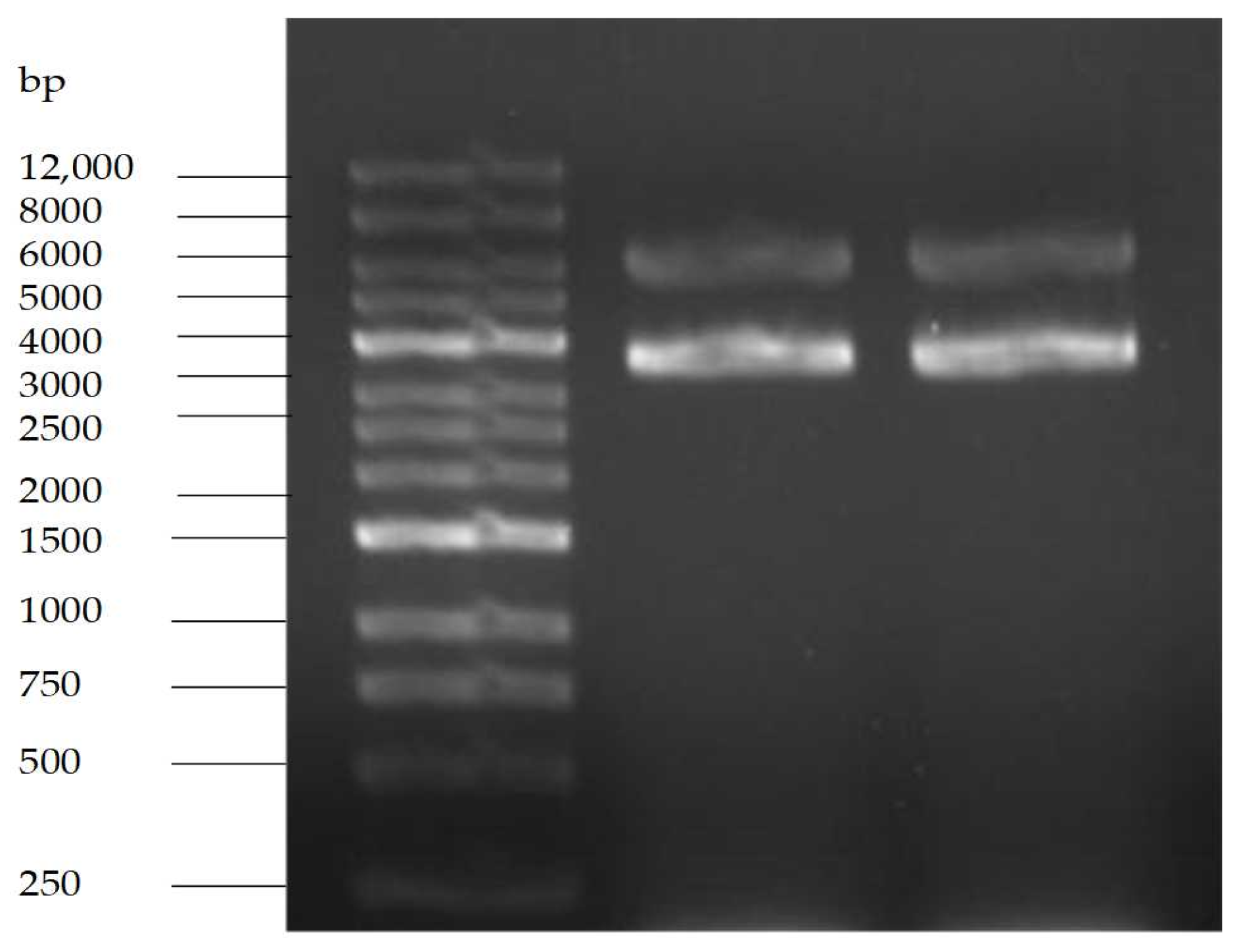
2.9.1. Purity Test Results of pET-28a
2.9.2. Determination of DNA Topoisomerase Activity in Crude Enzyme Extracts of S. aureus
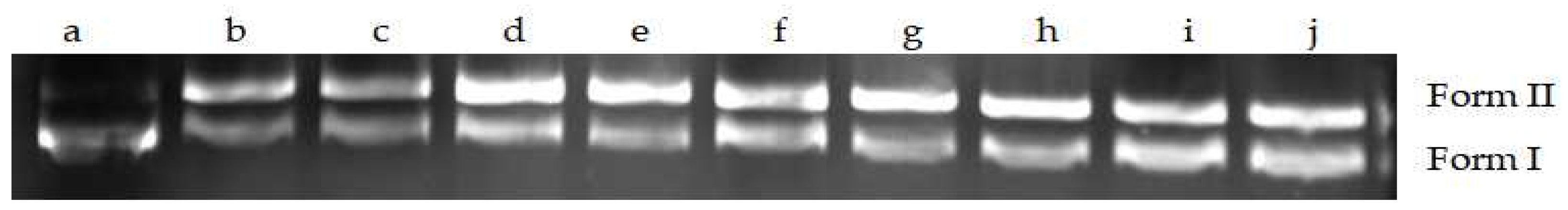
2.9.3. Effects of MA on DNA Topoisomerase I Activity of S. aureus
2.9.4. Effects of MA on DNA Topoisomerase II Activity of S. aureus
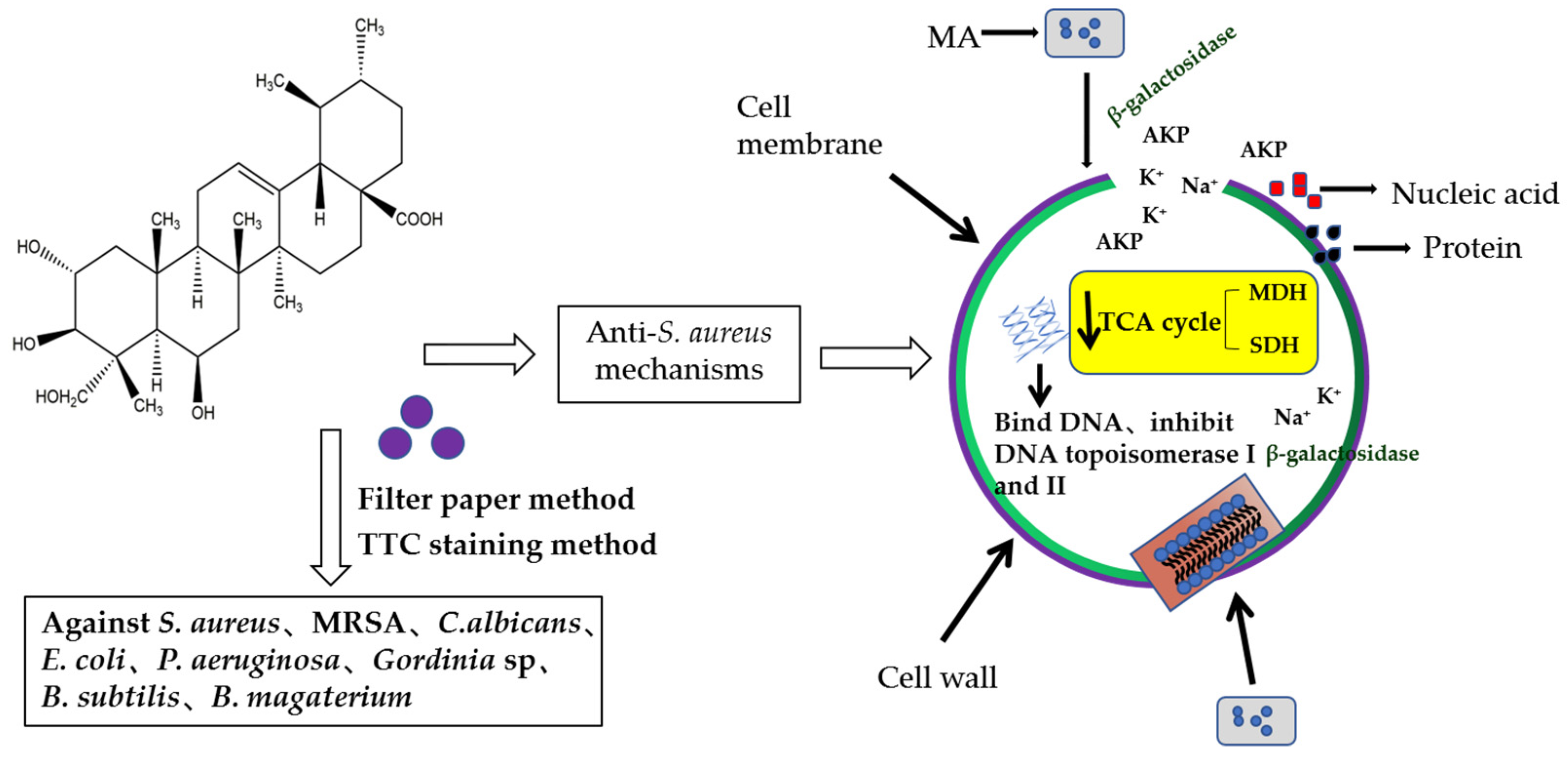
3. Discussion
4. Methods
4.1. Bacterial Strains and Bacterial Culture
4.2. Determination of Diameter of Inhibition Zone of MA
4.3. Determination of Minimum Inhibitory Concentration (MIC)
4.4. Antibacterial Curve of MA against S. aureus
4.5. Inhibitory Effect of MA on the Production of S. aureus Biofilm
4.6. Transmission Electron Microscope (TEM) Analysis of the Effect of MA on the Morphology of S. aureus Cells
4.7. Determination of Electrical Conductivity, DNA, RNA, and Other Macromolecular Substances Extravasation and Alkaline Phosphatase in Bacterial Liquid Phase
4.8. Effect of MA on Bacteria β- Determination of Galactosidase Content
4.9. Effect of MA on Soluble Protein Content of S. aureus Detected by SDS-PAGE
4.10. Determination of SDH and MDH Activities
4.11. Study on the Interaction Mode between MA and DNA
4.11.1. Agarose Gel Electrophoresis Was Used to Determine the Interaction Mechanism between MA and DNA
4.11.2. The Interaction Mode between MA and DNA was Determined by UV Absorption Spectrometry
4.12. Determination Method of the Influence of MA on DNA Topoisomerase Activity
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
Abbreviations
| MA | madecassic acid |
| MIC | minimum inhibitory concentration |
| OXA | oxacillin |
| OD | dimethyl sulfoxide |
| AKP | alkaline phosphatase |
| SDS-PAGE | sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| Form I | supercoiled DNA |
| Form II | relaxation unwinding DNA |
| MDH | malate dehydrogenase |
| SDH | succinate dehydrogenase |
| TTC | 2,3, 5-triphenyltetrazolium chloride |
| M | marker |
| C | control group |
| ATP | adenosine triphosphate |
| T | time |
| PBS | phosphate buffered saline |
| TEM | transmission electron microscope |
| ONPG | 2-nitrophenyl β-d-galactopyranoside |
| LB | luria Bertani |
| YPD | yeast extract peptone dextrose |
References
- Ahmad-Mansour, N.; Loubet, P.; Pouget, C.; Dunyach-Remy, C.; Sotto, A.; Lavigne, J.P.; Molle, V. Staphylococcus aureus toxins: An update on their pathogenic properties and potential treatments. Toxins 2021, 13, 677. [Google Scholar] [CrossRef]
- Cheung, G.Y.C.; Bae, J.S.; Otto, M. Pathogenicity and virulence of Staphylococcus aureus. Virulence 2021, 12, 547–569. [Google Scholar] [CrossRef]
- Chinese Pharmacopoeia Commission. Chinese Pharmacopoeia; Chinese Medical Sciences Press: Beijing, China, 2020; Volume I. [Google Scholar]
- Li, M. Study on characteristics and microscopic identification of Centella asiatica. Asia-Paci. Trad. Med. 2020, 16, 65–67. [Google Scholar]
- Zhang, X. Development and utilization of wild plant resources of Centella Asiatica. Chin. Hort. Abs. 2011, 27, 2. [Google Scholar]
- Su, J. Development of low-sugar Centella asiatica beverage. Food Indus. 2017, 45, 143. [Google Scholar]
- Liang, J.; Li, Y.; Liu, Y.; Nie, Y.; Yang, Z.; Zhang, Y.; Qian, Z.; Lin, W.; Song, C.; Zhang, Y. Studies on the neuroprotective effect and chemical constituents of the essential oil from Centella asiatica. West China J. Pharm. Sci. 2022, 37, 53–57. [Google Scholar]
- Mudaliana, S. Antimicrobial activity of Centella asiatica and Gigantochloa apus. J. Basic Clin. Physiol. Pharmacol. 2021, 32, 755–759. [Google Scholar] [CrossRef]
- Lu, J.; Chen, C.; Gai, R.; Qiu, H.; Wu, Y.; He, Q.; Yang, X. Protective effects and possible mechanisms of Centella asiatica (L.) urban extract against acute and chronic liver injury: Evidence from in vivo and in vitro studies. Phytother. Res. 2021, 35, 2785–2796. [Google Scholar] [CrossRef]
- Wu, Z.-W.; Li, W.-B.; Zhou, J.; Liu, X.; Wang, L.; Chen, B.; Wang, M.-K.; Ji, L.; Hu, W.-C.; Li, F. Oleanane-and ursane-type triterpene saponins from Centella asiatica exhibit neuroprotective effects. J. Agric. Food Chem. 2020, 68, 6977–6986. [Google Scholar] [CrossRef]
- Pan, J. Mechanism of Centella asiatica induced apoptosis in human ovarian cancer cell line A2780. Acta J. Chin. Med. 2018, 33, 521–524. [Google Scholar]
- Zhang, D.; Zhang, H.; Zhao, J.; Shi, X.; Yang, J.; Nie, X. Effect of Centella asiatica glycosides on promoting healing of diabetic skin ulcer in rats. J. Chin. Pharm. 2017, 52, 643–648. [Google Scholar]
- Wojnicz, D.; Tichaczek-Goska, D.; Kicia, M. Pentacyclic triterpenes combined with ciprofloxacin help to eradicate the biofilm formed in vitro by Escherichia coli. Indian. J. Med. Res. 2015, 141, 343. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Liu, T.-C.; Mong, M. Antibacterial effects and action modes of asiatic acid. Biomedicine 2015, 5, 16. [Google Scholar] [CrossRef]
- Sycz, Z.; Tichaczek-Goska, D.; Jezierska-Domaradzka, A.; Wojnicz, D. Are uropathogenic bacteria living in multispecies biofilm susceptible to active plant ingredient—Asiatic acid. Biomolecules 2021, 11, 1754. [Google Scholar] [CrossRef]
- Wojnicz, D.; Tichaczek-Goska, D.; Korzekwa, K.; Kicia, M.; Hendrich, A. Anti-enterococcal activities of pentacyclic triterpenes. Adv. Clin. Exp. Med. 2017, 26, 483–490. [Google Scholar] [CrossRef]
- Valdeira, A.S.C.; Darvishi, E.; Woldemichael, G.M.; Beutler, J.A.; Gustafson, K.R.; Salvador, J.A.R. Madecassic acid derivatives as potential anticancer agents: Synthesis and cytotoxic evaluation. J. Nat. Prod. 2019, 82, 2094–2105. [Google Scholar] [CrossRef]
- Razali, N.N.M.; Ng, C.T.; Fong, L.Y. Cardiovascular protective effects of Centella asiatica and its triterpenes: A review. Planta Med. 2019, 85, 1203–1215. [Google Scholar] [CrossRef] [Green Version]
- Hsu, Y.-M.; Hung, Y.-C.; Hu, L.; Lee, Y.-J.; Yin, M.-C. Anti-diabetic effects of madecassic acid and rotundic acid. Nutrients 2015, 7, 10065–10075. [Google Scholar] [CrossRef] [Green Version]
- Won, J.-H.; Shin, J.-S.; Park, H.-J.; Jung, H.-J.; Koh, D.-J.; Jo, B.-G.; Lee, J.-Y.; Yun, K.; Lee, K.-T. Anti-inflammatory effects of madecassic acid via the suppression of NF-κB pathway in LPS-induced RAW 264.7 macrophage cells. Planta Med. 2010, 76, 251–257. [Google Scholar] [CrossRef]
- Ngezahayo, J.; Pottier, L.; Ribeiro, S.O.; Delporte, C.; Fontaine, V.; Hari, L.; Stévigny, C.; Duez, P. Plastotoma rotundifolium aerial tissue extracthas antibacterial activities. Ind. Crop. Prod. 2016, 86, 301–310. [Google Scholar] [CrossRef]
- Wang, S.; Liu, X.-Q.; Kang, O.-H.; Kwon, D.-Y. Combination of sanguisorbigenin and conventional antibiotic therapy for Methicillin-resistant Staphylococcus aureus: Inhibition of biofilm formation and alteration of cell membrane permeability. Int. J. Mol. Sci. 2022, 23, 4232. [Google Scholar] [CrossRef]
- Ding, J.; Wang, Y.; Shen, J.; Zhu, J.; Jin, X. Effect of human antimicrobial peptide LL-37 on methicillin-resistant Staphylococcus aureus biofilms. Acad. J. Guangdong. Coll. Pharm. 2016, 32, 498–502. [Google Scholar]
- Shi, C.; Che, M.; Zhang, X.; Liu, Z.; Meng, R.; Bu, X.; Ye, H.; Guo, N. Antibacterial activity and mode of action of totarol against Staphylococcus aureus in carrot juice. J. Food. Sci. Technol. 2018, 55, 924–934. [Google Scholar] [CrossRef]
- Packianathan, S.; Arun, T.; Raman, N. DNA interaction and efficient antimicrobial activities of 4N chelating metal complexes. J. Photochem. Photobiol. B. 2015, 148, 160–167. [Google Scholar] [CrossRef]
- Duan, F.; Li, X.; Cai, S.; Xin, G.; Wang, Y.; Du, D.; He, S.; Huang, B.; Guo, X.; Zhao, H.; et al. Haloemodin as novel antibacterial agent inhibiting DNA gyrase and bacterial topoisomerase I. J. Med. Chem. 2014, 57, 3707–3714. [Google Scholar] [CrossRef]
- Zhang, J.-J.; Qu, L.-B.; Bi, Y.-F.; Pan, C.-X.; Yang, R.; Zeng, H.-J. Antibacterial activity and mechanism of chloroform fraction from aqueous extract of mugwort leaves (Artemisia argyi L.) against Staphylococcus aureus. Lett. Appl. Microbiol. 2022, 74, 893–900. [Google Scholar] [CrossRef]
- Zhou, D.; Liu, Z.-H.; Wang, D.-M.; Li, D.-W.; Yang, L.-N.; Wang, W. Chemical composition, antibacterial activity and related mechanism of valonia and shell from Quercus variabilis Blume (Fagaceae) against Salmonella paratyphi a and Staphylococcus aureus. BMC Complement. Altern. Med. 2019, 19, 271. [Google Scholar] [CrossRef] [Green Version]
- Ning, Y.; Hou, L.; Ma, M.; Li, M.; Zhao, Z.; Zhang, D.; Wang, Z.; Jia, Y. Synergistic antibacterial mechanism of sucrose laurate combined with nisin against Staphylococcus aureus and its application in milk beverage. LWT 2022, 158, 113145. [Google Scholar] [CrossRef]
- Hillman, T. Reducing bacterial antibiotic resistance by targeting bacterial metabolic pathways and disrupting RND efflux pump activity. Iberoam J. Med. 2022, 4, 60–74. [Google Scholar] [CrossRef]
- Wang, X.; Shen, Y.; Thakur, K.; Han, J.; Zhang, J.-G.; Hu, F.; Wei, Z.-J. Antibacterial activity and mechanism of ginger essential oil against Escherichia coli and Staphylococcus aureus. Molecules 2020, 25, 3955. [Google Scholar] [CrossRef]
- Huang, M.; Luo, J.; Shen, J. Effects of dihydroartemisinin and cefuroxim on Escherichia coli and its mechanism. Chin. J. Chin. Mater. Med. 2020, 45, 2975–2981. [Google Scholar]
- Lan, W.; Zhang, N.; Liu, S.; Chen, M.; Xie, J. ε-Polylysine inhibits Shewanella putrefaciens with membrane disruption and cell damage. Molecules 2019, 24, 3727. [Google Scholar] [CrossRef] [Green Version]
- Kang, S.; Kong, F.; Shi, X.; Han, H.; Li, M.; Guan, B.; Yang, M.; Cao, X.; Tao, D.; Zheng, Y. Activity and mechanism of lactobionic acid against Pseudomonas fluorescens and Methicillin-resistant Staphylococcus aureus and its application on whole milk. Food Control 2020, 108, 106876. [Google Scholar] [CrossRef]
- Zhang, J.; Ye, K.-P.; Zhang, X.; Pan, D.-D.; Sun, Y.-Y.; Cao, J.-X. Antibacterial activity and mechanism of action of black pepper essential oil on meat-borne Escherichia coli. Front. Microbiol. 2017, 7, 2094. [Google Scholar] [CrossRef] [Green Version]
- Wang, N.; Liu, X.; Li, J.; Zhang, Q.; Li, X.; An, Q.; Ye, X.; Zhao, Z.; Cai, L.; Han, Y.; et al. Antibacterial mechanism of the synergistic combination between streptomycin and alcohol extracts from the Chimonanthus salicifolius SY Hu. leaves. J. Ethnopharmacol. 2020, 250, 112467. [Google Scholar] [CrossRef]
- Li, X.; He, C.; Song, L.; Li, T.; Cui, S.; Zhang, L.; Jia, Y. Antimicrobial activity and mechanism of larch bark procyanidins against Staphylococcus aureus. Acta Biochim. Biophys. Sin. 2017, 49, 1058–1066. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Chen, Z.; Zhang, S.; Shao, X.; Zhou, Z. Antibacterial activity of the structurally novel ocotillol-type lactone and its analogues. Fitoterapia 2020, 144, 104597. [Google Scholar] [CrossRef]
- Guo, Y.; Liu, Y.; Zhang, Z.; Chen, M.; Zhang, D.; Tian, C.; Liu, M.; Jiang, G. The antibacterial activity and mechanism of action of luteolin against Trueperella pyogenes. Infect. Drug. Resist. 2020, 13, 1697. [Google Scholar] [CrossRef]
- Karia, P.S.; Vekariya, P.A.; Patidar, A.P.; Kanthecha, D.N.; Bhatt, B.S.; Patel, M.N. DNA interaction, in vitro antibacterial and cytotoxic activities of Ru(III) heterochelates. Acta. Chim. Slov. 2019, 66, 944–949. [Google Scholar] [CrossRef]
- Wang, J.; Wan, Y.; Lang, W.; Li, L.; Li, S.; Yi, T. Spectral study on the interaction of antimicrobial peptide AN5-2 with Escherichia coli genomic DNA. Chin-Fore. Women’s. Health 2016, 14, 220–221. [Google Scholar]
- Kwon, J.; Kang, H.Y.; Yang, H. Permeabilization-free β-galactosidase-induction-based electrochemical detection of Escherichia coli. Sens. Actuators B-Chem. 2021, 337, 129768. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strains | Inhibition Zone (mm) | MIC (µg/mL) | ||
|---|---|---|---|---|
| MA | MA | OXA | ||
| Gram(−) | E. coli | 10.0 | 250.0 | / |
| Gram(+) | S. aureus | 13.0 | 31.2 | 0.0 |
| Gram(+) | MRSA | 14.5 | 62.5 | 7.8 |
| Gram(+) | Gordinia sp. | 13.5 | / | / |
| Gram(−) | P. aeruginosa | 11.5 | 125.0 | / |
| Fungus | C.albicans | 14.0 | / | / |
| Gram(+) | B. subtilis | 10.5 | 62.5 | 3.9 |
| Gram(+) | B. magaterium | 10.5 | 62.5 | 1.9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wei, C.; Cui, P.; Liu, X. Antibacterial Activity and Mechanism of Madecassic Acid against Staphylococcus aureus. Molecules 2023, 28, 1895. https://doi.org/10.3390/molecules28041895
Wei C, Cui P, Liu X. Antibacterial Activity and Mechanism of Madecassic Acid against Staphylococcus aureus. Molecules. 2023; 28(4):1895. https://doi.org/10.3390/molecules28041895
Chicago/Turabian StyleWei, Chunling, Peiwu Cui, and Xiangqian Liu. 2023. "Antibacterial Activity and Mechanism of Madecassic Acid against Staphylococcus aureus" Molecules 28, no. 4: 1895. https://doi.org/10.3390/molecules28041895
APA StyleWei, C., Cui, P., & Liu, X. (2023). Antibacterial Activity and Mechanism of Madecassic Acid against Staphylococcus aureus. Molecules, 28(4), 1895. https://doi.org/10.3390/molecules28041895