
Antioxidant Activity of Polysaccharides from the Edible Mushroom Pleurotus eryngii

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Proximate Composition
2.2. Glucan Content
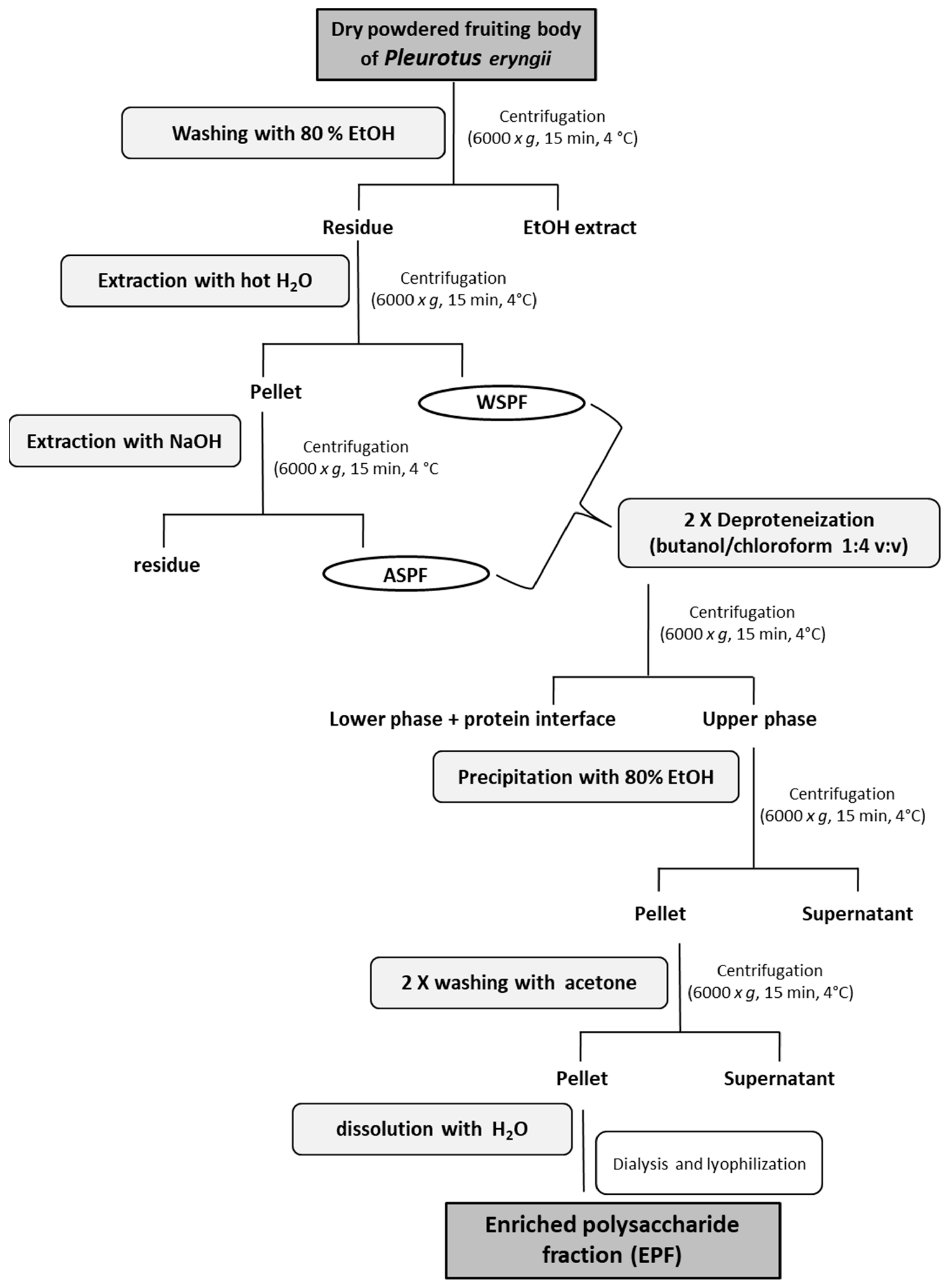
2.3. Extraction of Polysaccharides from the Edible Mushroom P. eryngii
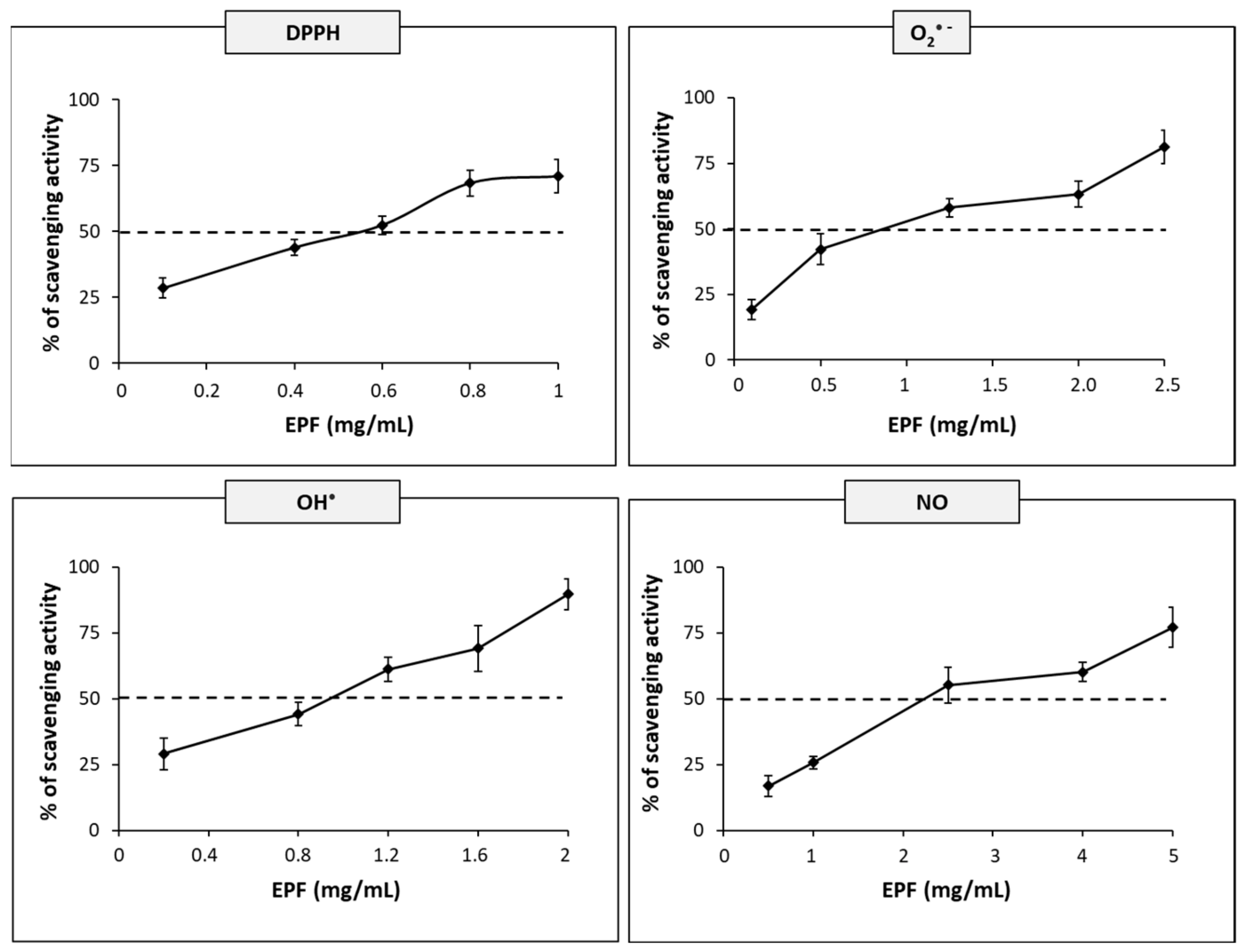
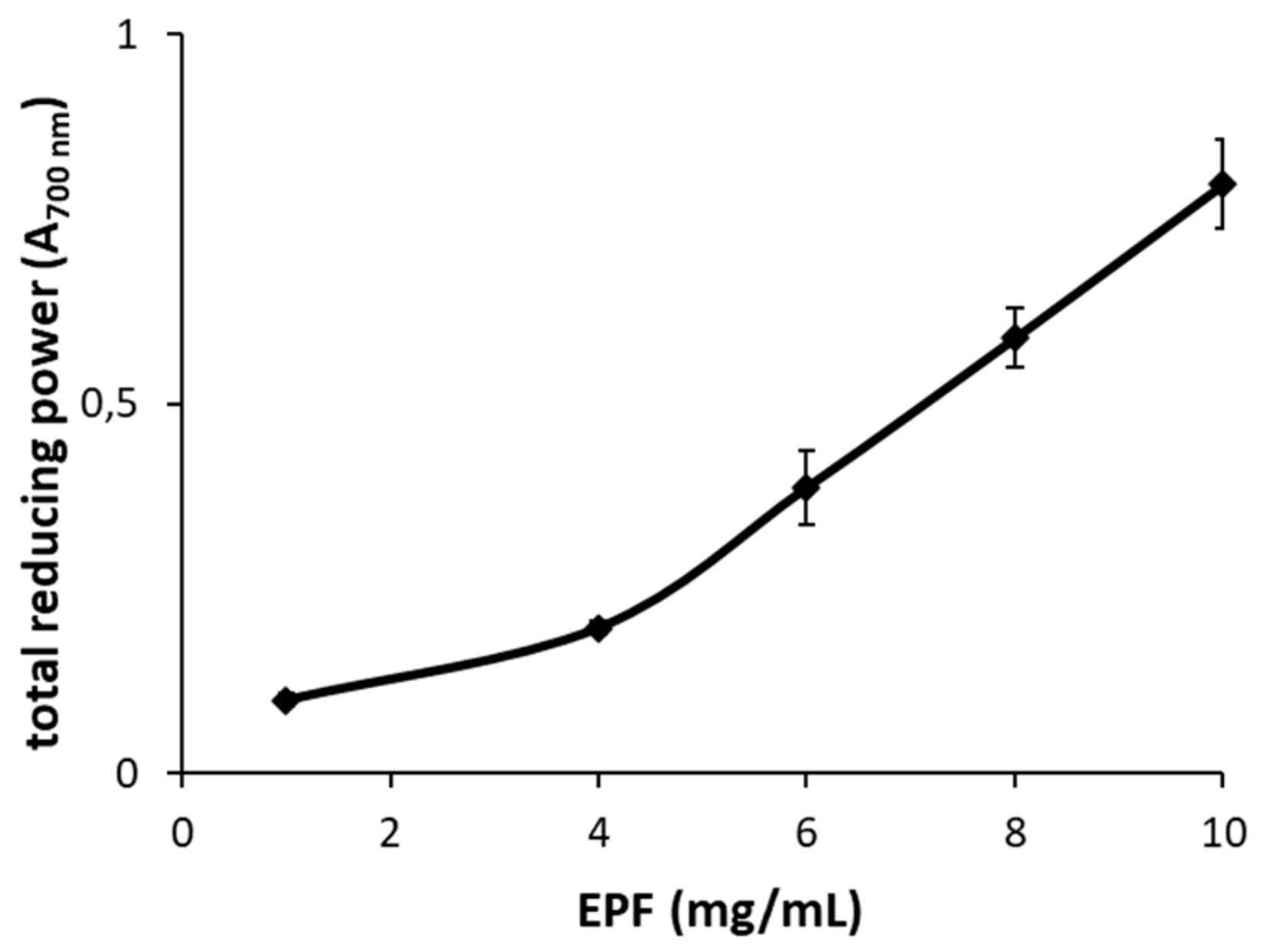
2.4. Antioxidant Activity
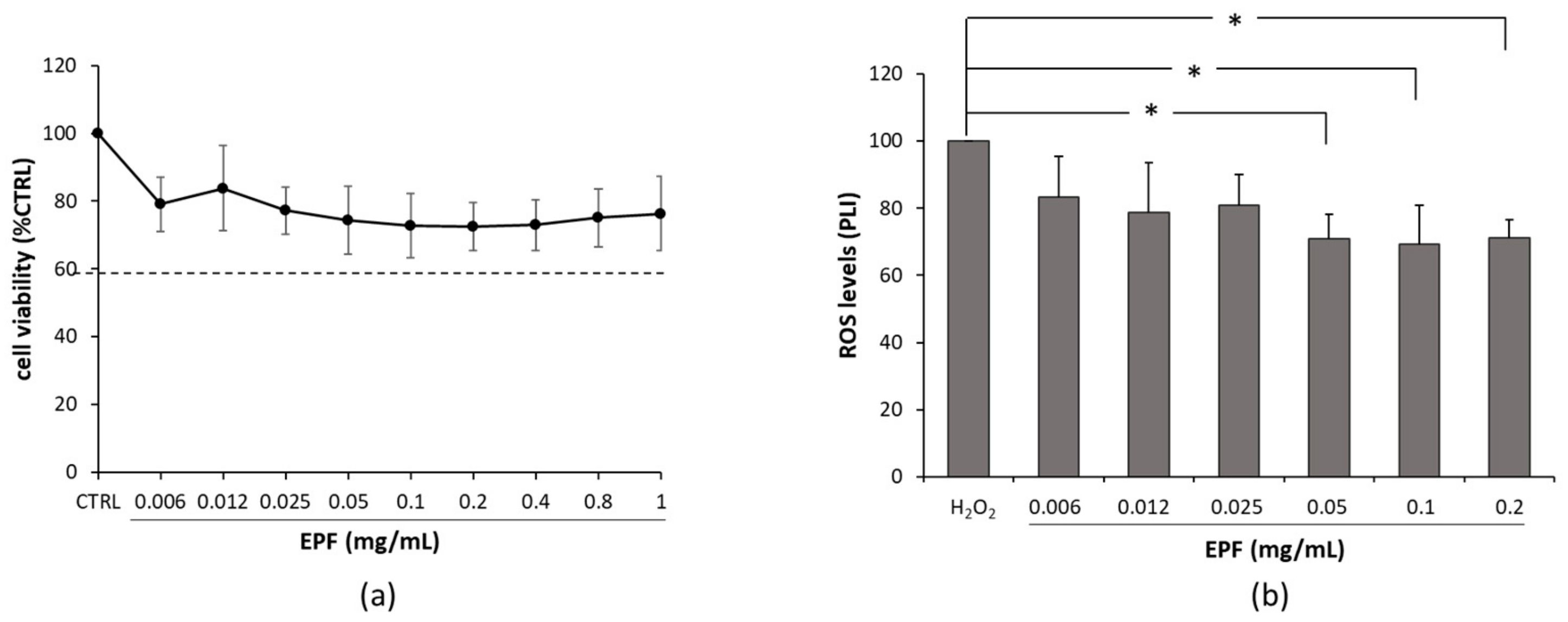
2.5. Effect of EPF on DI-TNC1 Viability and ROS Production
3. Materials and Methods
3.1. Chemicals
3.2. Samples of P. eryngii Fruiting Bodies
3.3. Proximate Composition
3.4. Glucan Measurement
3.5. Preparation of Crude Polysaccharide Fraction
3.6. Determination of Molecular Weight Range of Glucans
3.7. DPPH Radical Scavenging Activity
3.8. Superoxide Radical Scavenging Activity
3.9. Hydroxyl Radical Scavenging Activity
3.10. Nitric Oxide Scavenging Activity
3.11. Total Reducing Power
3.12. MTT Viability Assay
3.13. Intracellular Reactive Oxygen Species Detection
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ferreira, I.C.F.R.; Barros, L.; Abreu, R.M.V. Antioxidants in wild mushrooms. Curr. Med. Chem. 2009, 16, 1543–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindequist, U.; Niedermayer, T.H.J.; Julich, W.D. The pharmacological potential of mushrooms. Evid. Based Complement. Altern. Med. 2005, 2, 285–299. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mishra, K.K.; Pal, R.S.; Arunkumar, R.; Chandrashekara, C.; Jain, S.K.; Bhatt, J.C. Antioxidant properties of different edible mushroom species and increased bioconversion efficiency of Pleurotus eryngii using locally available casing materials. Food Chem. 2013, 138, 1557–1563. [Google Scholar] [CrossRef] [PubMed]
- Patel, Y.; Naraian, R.; Singh, V.K. Medicinal properties of Pleurotus species (oyster mushroom): A review. World J. Fungal Plant. Biol. 2012, 3, 1–12. [Google Scholar]
- Kozarski, M.; Klaus, A.; Jakovljevic, D.; Todorovic, N.; Vunduk, J.; Petrovic, P.; Niksic, M.; Vrvic, M.M.; Van Griensven, L. Antioxidants of edible mushrooms. Molecules 2015, 20, 19489–19525. [Google Scholar] [CrossRef] [Green Version]
- Wasser, S.P. Medicinal mushroom science: History, current status, future trends, and unsolved problems. Int. J Med. Mushrooms 2010, 12, 1–16. [Google Scholar] [CrossRef]
- Singhal, S.; Rasane, P.; Kaur, S.; Garba, U.; Singh, J.; Raj, N.; Gupta, N. Mushroom cultivation, processing and value-added products: A patent-based review. Recent Pat. Food Nutr. Agric. 2019, 10, 3–19. [Google Scholar] [CrossRef]
- Carrasco-González, J.A.; Serna-Saldívar, S.O.; Gutiérrez-Uribe, J.A. Nutritional composition and nutraceutical properties of the Pleurotus fruiting bodies: Potencial use as food ingredient. J. Food Compos. Anal. 2017, 58, 69–81. [Google Scholar] [CrossRef]
- Ma, G.; Yang, W.; Mariga, A.M.; Fang, Y.; Ma, N.; Pei, F.; Hu, Q. Purification, characterization and antitumor activity of polysaccharides from Pleurotus eryngii residue. Carbohydr. Polym. 2014, 114, 297–305. [Google Scholar] [CrossRef]
- Hu, Q.; Du, H.; Ma, G.; Pei, F.; Ma, N.; Yuan, B.; Nakata, P.A.; Yang, W. Purification, identification and functional characterization of an immunomodulatory protein from Pleurotus eryngii. Food Funct. 2018, 9, 3764–3775. [Google Scholar] [CrossRef]
- Zhiming, F.; Yi, L.; Qiang, Z. A potent pharmacological mushroom: Pleurotus eryngii. Fungal Genom. Biol. 2016, 6, 1. [Google Scholar]
- Petraglia, T.; Latronico, T.; Liuzzi, G.M.; Fanigliulo, A.; Crescenzi, A.; Rossano, R. Edible mushrooms as source of fibrin(ogen)olytic enzymes: Comparison between four cultivated species. Molecules 2022, 27, 8145. [Google Scholar] [CrossRef] [PubMed]
- Gong, P.; Long, H.; Guo, Y.; Wang, S.; Chen, F.; Chen, X. Isolation, structural characterization, and hypoglycemic activities in vitro of polysaccharides from Pleurotus eryngii. Molecules 2022, 27, 7140. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Mao, D.; Yong, Y.; Li, J.; Wei, H.; Lu, L. Hepatoprotective and hypolipidemic effects of water-soluble polysaccharidic extract of Pleurotus eryngii. Food Chem. 2012, 130, 687–694. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhang, Z.; Wang, L.; Li, W.; Du, J.; Zhang, S.; Chen, X. Hypolipidemic mechanism of Pleurotus eryngii polysaccharides in high-fat diet-induced obese mice based on metabolomics. Front. Nutr. 2023, 10, 1118923. [Google Scholar] [CrossRef] [PubMed]
- Manzi, P.; Marconi, S.; Aguzzi, A.; Pizzoferrato, L. Commercial mushrooms: Nutritional quality and effect of cooking. Food Chem. 2004, 84, 201–206. [Google Scholar] [CrossRef]
- Zhang, B.; Li, Y.; Zhang, F.; Linhardt, R.J.; Zeng, G.; Zhang, A. Extraction, structure and bioactivities of the polysaccharides from Pleurotus eryngii: A review. Int. J. Biol. Macromol. 2020, 150, 1342–1347. [Google Scholar] [CrossRef] [PubMed]
- Friedman, M. Mushroom polysaccharides: Chemistry and antiobesity, antidiabetes, anticancer, and antibiotic properties in cells, rodents, and humans. Foods 2016, 5, 80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giavasis, I. Bioactive fungal polysaccharides as potential functional ingredients in food and nutraceuticals. Curr. Opin. Biotechol. 2014, 26, 162–173. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Qu, Y.; Wang, Y.; Wang, X.; Xu, J.; Zhao, H.; Zheng, D.; Sun, L.; Tai, G.; Zhou, Y.; et al. β-1,6-Glucan from Pleurotus eryngii modulates the immunity and gut microbiota. Front. Immunol. 2022, 13, 859923. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chen, X.; Zhao, Y.; Jia, W.; Chang, X.; Liu, H.; Liu, N. Optimization of extraction parameters of Pleurotus eryngii polysaccharides and evaluation of the hypolipidemic effect. RSC Adv. 2020, 10, 11918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, A.; Li, X.; Xing, C.; Yang, J.; Sun, P. Antioxidant activity of polysaccharide extracted from Pleurotus eryngii using response surface methodology. Int. J. Biol. Macromol. 2014, 65, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Mickova, K.; Jablonsky, I.; Slukoya, M.; Copikova, J. Mushrooms of genus Pleurotus as a source of dietary fibers and glucans for food supplements. Czech. J. Food Sci. 2008, 26, 441–446. [Google Scholar] [CrossRef] [Green Version]
- Synytsya, A.; Novak, M. Structural analysis of glucans. Ann. Transl. Med. 2014, 2, 17. [Google Scholar] [PubMed]
- Manzi, P.; Pizzoferrato, L. Beta-glucans in edible mushrooms. Food Chem. 2000, 68, 315–318. [Google Scholar] [CrossRef]
- Wasser, S.P. Medicinal mushrooms as a source of antitumor and immunomodulating polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar]
- Li, S.; Xiong, Q.; Lai, X.; Li, X.; Wan, M.; Zhang, J.; Li, Y. Molecular modification of polysaccharides and resulting bioactivities. Compr. Rev. Food Sci. Food Saf. 2015, 15, 237–250. [Google Scholar] [CrossRef] [Green Version]
- Yan, J.; Zhu, L.; Qu, Y.; Qu, X.; Mu, M.; Zhang, M.; Muneer, G.; Zhou, Y.; Sun, L. Analyses of active antioxidant polysaccharides from four edible mushrooms. Int. J. Biol. Macromol. 2019, 123, 945–956. [Google Scholar] [CrossRef]
- Cateni, F.; Zacchigna, M.; Bavisotto, C.C.; Procida, G.; Bonaventura, G.; Saporita, P.; Calvo, R.; Venturella, G.; Gargano, M.L. Structural characterization of polysaccharides of a productive strain of the culinary-medicinal king Oyster mushroom, Pleurotus eryngii (Agaricomycetes), from Italy. Int. J. Med. Mushrooms 2018, 20, 717–726. [Google Scholar] [CrossRef]
- Zhao, Y.; Chen, X.; Jia, W.; Gong, G.; Zhao, Y.; Li, G.; Zhou, J.; Li, X.; Zhao, Y.; Ma, W. Extraction, isolation, characterisation, antioxidant and anti-fatigue activities of Pleurotus eryngii polysaccharides. Int. J. Food Sci. Technol. 2020, 55, 2492–2508. [Google Scholar] [CrossRef]
- Sun, X.; Hao, L.; Ma, H.; Li, T.; Zheng, L.; Ma, Z.; Zhai, G.; Wang, L.; Gao, S.; Liu, X.; et al. Extraction and in vitro antioxidant activity of exopolysaccharide by Pleurotus eryngii SI-02. Braz. J. Microbiol. 2013, 44, 1081–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozarski, M.S.; Klaus, A.S.; Niksic, M.P.; van Griensven, L.J.L.D.; Vrvic, M.M.; Jakovljevic, D.M. Polysaccharides of higher fungi: Biological role, structure and antioxidative activity. Chem. Ind. 2014, 68, 305–320. [Google Scholar] [CrossRef]
- Ping, C.X.; Yan, C.; Bing, L.S.; Guo, C.Y.; Yun, L.J.; Ping, L.L. Free radical scavenging of Ganoderma lucidum polysaccharides and its effect on antioxidant enzymes and immunity activities in cervical carcinoma rats. Carbohydr. Polym. 2009, 77, 389–393. [Google Scholar]
- Kishk, Y.F.M.; Al-Sayed, H.M.A. Free-radical scavenging and antioxidative activities of some polysaccharides in emulsions. LWT Food Sci. Technol. 2007, 40, 270–277. [Google Scholar] [CrossRef]
- Mingyi, Y.; Belwal, T.; Devkota, H.P.; Li, L.; Luo, Z. Trends of utilizing mushroom polysaccharides (MPs) as potent nutraceutical components in food and medicine: A comprehensive review. Trends Food Sci. Technol. 2019, 92, 94–110. [Google Scholar] [CrossRef]
- Garcia, J.; Rodrigues, F.; Saavedra, M.J.; Nunes, F.M.; Marques, G. Bioactive polysaccharides from medicinal mushrooms: A review on their isolation, structural characteristics and antitumor activity. Food Biosci. 2022, 49, 101955. [Google Scholar] [CrossRef]
- Zhang, C.J.; Guo, J.Y.; Cheng, H.; Lin, L.; Liu, Y.; Shi, Y.; Xu, J.; Yu, H.T. Protective effects of the king oyster culinary-medicinal mushroom, Pleurotus eryngii (Agaricomycetes), polysaccharides on β-amyloid-induced neurotoxicity in PC12 cells and aging rats, in vitro and in vivo studies. Int. J. Med. Mushrooms 2020, 22, 325–333. [Google Scholar] [CrossRef]
- Kushairi, N.; Phan, C.W.; Sabaratnam, V.; Vidyadaran, S.; Naidu, M.; David, P. Comparative neuroprotective, anti-Inflammatory and neurite outgrowth activities of extracts of king oyster mushroom, Pleurotus eryngii (Agaricomycetes). Int. J. Med. Mushrooms 2020, 22, 1171–1181. [Google Scholar] [CrossRef]
- Yamashina, K.; Yamamoto, S.; Matsumoto, M.; Iwasa, K.; Takeda, N.; Haruta, C.; Maruyama, K.; Shimizu, K.; Yoshikawa, K. Suppressive effect of fruiting bodies of medicinal mushrooms on demyelination and motor dysfunction in a Cuprizone-induced multiple sclerosis mouse model. Int. J. Med. Mushrooms 2022, 24, 15–24. [Google Scholar] [CrossRef]
- Roncero-Ramos, I.; Mendiola-Lanao, M.; Perez-Clavijo, M.; Delgado-Andrade, C. Effect of different cooking methods on nutritional value and antioxidant activity of cultivated mushrooms. Int. J. Food Sci. Nutr. 2017, 68, 287–297. [Google Scholar] [CrossRef]
- Sari, M.; Prange, A.; Lelley, J.I.; Hambitzer, R. Screening of beta-glucan contents in commercially cultivated and wild growing mushrooms. Food Chem. 2017, 216, 45–51. [Google Scholar] [CrossRef] [PubMed]
- McCleary, B.V.; Draga, A. Measurement of β-glucan in mushrooms and mycelial products. J. AOAC Int. 2016, 99, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bradford, M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Synytsya, A.; Míčková, K.; Synytsya, A.; Jablonský, I.; Spěváček, J.; Erban, V.; Kováříková, E.; Čopíková, J. Glucans from fruit bodies of cultivated mushrooms Pleurotus ostreatus and Pleurotus eryngii: Structure and potential prebiotic activity. Carbohydr. Polym. 2009, 76, 548–556. [Google Scholar] [CrossRef]
- Ellefsen, C.F.; Winther Wold, C.; Wilkins, A.L.; Rise, F.; Samuelsen, A.B.C. Water-soluble polysaccharides from Pleurotus eryngii fruiting bodies, their activity and affinity for Toll-like receptor 2 and dectin-1. Carbohydr. Polym. 2021, 264, 117991. [Google Scholar] [CrossRef]
- Rodrigues Barbosa, J.; Dos Santos Freitas, M.M.; da Silva Martins, L.H.; de Carvalho, R.N.J. Polysaccharides of mushroom Pleurotus spp.: New extraction techniques, biological activities and development of new technologies. Carbohydr. Polym. 2020, 229, 115550. [Google Scholar] [CrossRef]
- Ruthes, A.C.; Smiderle, F.R.; Iacomini, M. D-Glucans from edible mushrooms: A review on the extraction, purificatio and chemical characterization approaches. Carbohydr. Polym. 2015, 117, 753–761. [Google Scholar] [CrossRef]
- Xu, D.; Wang, H.; Zheng, W.; Gao, Y.; Wang, M.; Zhang, Y.; Gao, Q. Charaterization and immunomodulatory activities of polysaccharide isolated from Pleurotus eryngii. Int. J. Biol. Macromol. 2016, 92, 30–36. [Google Scholar] [CrossRef]
- Li, S.; Shah, N.P. Characterization, antioxidative and bifidogenic effects of polysaccharides from Pleurotus eryngii after heat treatments. Food Chem. 2016, 197, 240–249. [Google Scholar] [CrossRef]
- He, P.; Zhang, A.; Zhou, S.; Zhang, F.; Linhardt, R.J.; Sun, P. Structural elucidation of polysaccharide containing 3-O-methyl galactose from fruiting bodies of Pleurotus citrinopileatus. Carbohydr. Res. 2016, 434, 72–76. [Google Scholar] [CrossRef]
- Valko, M.; Leibfritz, D.; Moncola, J.; Cronin, M.T.D.; Mazur, M.; Telser, J. Free radicals and antioxidants in normal physiological functions and human disease. Int. J. Biochem. Cell. Biol. 2007, 39, 44–84. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Zhou, B.; Lin, R.; Jia, L.; Deng, P.; Fan, K.; Wang, G.; Wang, L.; Zhang, J. Extraction and antioxidant activities of intracellular polysaccharide from Pleurotus sp. mycelium. Int. J. Biol. Macromol. 2010, 47, 116–119. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; You, Q.; Zhou, X. Complex enzyme-assisted extraction, purification, and antioxidant activity of polysaccharides from the Button Mushroom, Agaricus bisporus (Higher Basidiomycetes). Int. J. Med. Mushrooms 2015, 17, 987–996. [Google Scholar] [CrossRef] [PubMed]
- Di Meo, S.; Reed, T.T.; Venditti, P.; Victor, V.M. Role of ROS and RNS sources in physiological and pathological conditions. Oxid. Med. Cell. Longev. 2016, 2016, 1245049. [Google Scholar] [CrossRef]
- Ghasemi, M.; Mayasi, Y.; Hannoun, A.; Eslami, S.M.; Carandang, R. Nitric oxide and mitochondrial function in neurological diseases. Neuroscience 2018, 376, 48–71. [Google Scholar] [CrossRef]
- Xu, X.L.; Li, S.; Zhang, R.; Le, W.D. Neuroprotective effects of naturally sourced bioactive polysaccharides: An update. Neural Regen. Res. 2022, 17, 1907–1912. [Google Scholar]
- Radany, E.H.; Brenner, M.; Besnard, F.; Bigornia, V.; Bishop, J.M.; Deschepper, C.F. Directed establishment of rat brain cell lines with the phenotypic characteristics of type 1 astrocytes. Proc. Natl. Acad. Sci. USA 1992, 89, 6467–6471. [Google Scholar] [CrossRef] [Green Version]
- Picca, A.; Calvani, R.; Coelho-Junior, H.J.; Landi, F.; Bernabei, R.; Marzetti, E. Mitochondrial dysfunction, oxidative stress, and neuroinflammation: Intertwined roads to neurodegeneration. Antioxidants 2020, 9, 647. [Google Scholar] [CrossRef]
- Teleanu, D.M.; Niculescu, A.G.; Lungu, I.I.; Radu, C.I.; Vladâcenco, O.; Roza, E.; Costăchescu, B.; Grumezescu, A.M.; Teleanu, R.I. An Overview of oxidative stress, neuroinflammation, and neurodegenerative diseases. Int. J. Mol. Sci. 2022, 23, 5938. [Google Scholar] [CrossRef]
- Tobore, T.O. Oxidative/nitroxidative stress and multiple sclerosis. J. Mol. Neurosci. 2021, 71, 506–514. [Google Scholar] [CrossRef]
- Riccio, P.; Rossano, R.; Liuzzi, G.M. May diet and dietary supplements improve the wellness of multiple sclerosis patients? A molecular approach. Autoimmune Dis. 2011, 2010, 249842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larocca, M.; Di Marsico, M.; Riccio, P.; Rossano, R. The in vitro antioxidant properties of Muscari comosum bulbs and their inhibitory activity on enzymes involved in inflammation, post-prandial hyperglycemia, and cognitive/neuromuscular functions. J. Food Biochem. 2018, 2018, e12580. [Google Scholar] [CrossRef]
- McGrattan, A.M.; McGuinness, B.; McKinley, M.C.; Kee, F.; Passmore, P.; Woodside, J.V.; McEvoy, C.T. Diet and inflammation in cognitive ageing and Alzheimer’s disease. Curr. Nutr. Rep. 2019, 8, 53–65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aryal, S.; Skinner, T.; Bridges, B.; Weber, J.T. The Pathology of Parkinson’s disease and potential benefit of dietary polyphenols. Molecules 2020, 25, 4382. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis, 16th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1995. [Google Scholar]
- Staub, A.M. Removal of proteins: Sevag method. Methods Carbohydr. Chem. 1965, 5, 5–6. [Google Scholar]
- Blumenkrantz, N.; Asboe-Hansen, G. New method for quantitative determination of uronic acids. Anal. Biochem. 1973, 54, 484–489. [Google Scholar] [CrossRef] [PubMed]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Meth. Enzymol. 1999, 299, 152–178. [Google Scholar]
- Larocca, M.; Perna, A.M.; Simonetti, A.; Gambacorta, E.; Iannuzzi, A.; Perucatti, A.; Rossano, R. Antioxidant and anti-inflammatory effects of leaves cauliflower powder-enriched diet against LPS induced toxicity in rabbits. Food Funct. 2017, 8, 3288–3296. [Google Scholar] [CrossRef]
- Winterbourn, C.C.; Sutton, H.C. Hydroxyl radical productionfrom hydrogen peroxide and enzymatically generated paraquat radicals: Catalytic requirements and oxygen dependence. Arch. Biochem. Biophys. 1984, 235, 116–126. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reactions: Antioxidant activities of products of browning reaction prepared from glucose amine. Jpn. J. Nutr. 1986, 44, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Latronico, T.; Pati, I.; Ciavarella, R.; Fasano, A.; Mengoni, F.; Lichtner, M.; Vullo, V.; Mastroianni, C.M.; Liuzzi, G.M. In vitro effect of antiretroviral drugs on cultured primary astrocytes: Analysis of neurotoxicity and matrix metalloproteinase inhibition. J. Neurochem. 2018, 144, 271–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Latronico, T.; Larocca, M.; Milella, S.; Fasano, A.; Rossano, R.; Liuzzi, G.M. Neuroprotective potential of isothiocyanates in an in vitro model of neuroinflammation. Inflammopharmacology 2021, 29, 561–571. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Moisture | Proteins | Fat | Ash | Carbohydrates | Energy | |
|---|---|---|---|---|---|---|
| (g/100 g fw) (g/100 g dm) | 89.08 ± 0.98 | 1.36 ± 0.02 12.45 ± 0.18 | 0.17 ± 0.01 1.55 ± 0.09 | 0.71 ± 0.05 6.50 ± 0.46 | 8.68 ± 0.21 79.49 ± 1.92 | * 41.69 ± 1.34 ** 381.78 ± 12.27 |
| Total Glucans | α-Glucans | β-Glucans |
|---|---|---|
| (g/100 g Dry Mass) | (g/100 g Dry Mass) | (g/100 g Dry Mass) |
| 28.37 ± 0.92 | 2.15 ± 0.18 | 26.22 ± 0.73 |
| (100%) | (7.58 ± 0.63%) | (92.42 ± 2.57%) |
| a Yield | b Glucan (%) | b Protein (%) | b Polyphenol (%) | b Uronic Acid (%) |
|---|---|---|---|---|
| 26.26 ± 0.93 | 89.59 ± 2.55 | 3.55 ± 0.41 | n.d. | 2.24 ± 0.23 |
| Glucans < 10 kDa | 10 kDa < Glucans < 50 kDa | Glucans > 50 kDa |
|---|---|---|
| (n.d.) | (79.61 ± 5.08%) | (17.54 ± 1.46%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petraglia, T.; Latronico, T.; Fanigliulo, A.; Crescenzi, A.; Liuzzi, G.M.; Rossano, R. Antioxidant Activity of Polysaccharides from the Edible Mushroom Pleurotus eryngii. Molecules 2023, 28, 2176. https://doi.org/10.3390/molecules28052176
Petraglia T, Latronico T, Fanigliulo A, Crescenzi A, Liuzzi GM, Rossano R. Antioxidant Activity of Polysaccharides from the Edible Mushroom Pleurotus eryngii. Molecules. 2023; 28(5):2176. https://doi.org/10.3390/molecules28052176
Chicago/Turabian StylePetraglia, Tania, Tiziana Latronico, Angela Fanigliulo, Aniello Crescenzi, Grazia Maria Liuzzi, and Rocco Rossano. 2023. "Antioxidant Activity of Polysaccharides from the Edible Mushroom Pleurotus eryngii" Molecules 28, no. 5: 2176. https://doi.org/10.3390/molecules28052176
APA StylePetraglia, T., Latronico, T., Fanigliulo, A., Crescenzi, A., Liuzzi, G. M., & Rossano, R. (2023). Antioxidant Activity of Polysaccharides from the Edible Mushroom Pleurotus eryngii. Molecules, 28(5), 2176. https://doi.org/10.3390/molecules28052176