
Parthenolide Phytosomes Attenuated Gentamicin-Induced Nephrotoxicity in Rats via Activation of Sirt-1, Nrf2, OH-1, and NQO1 Axis


Abstract
:
1. Introduction
2. Results
2.1. Preparation and Characterization of PTL-Phytosomes
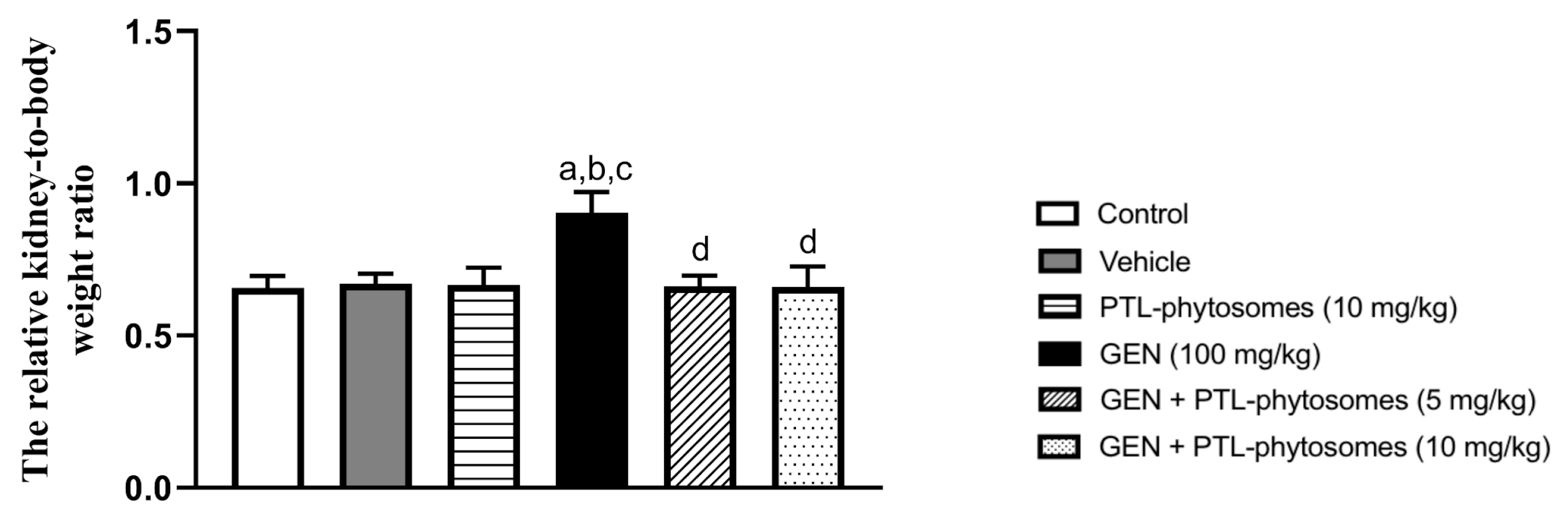
2.2. PTL-Phytosomes Attenuated GEN-Induced Kidney Hypertrophy in Rats
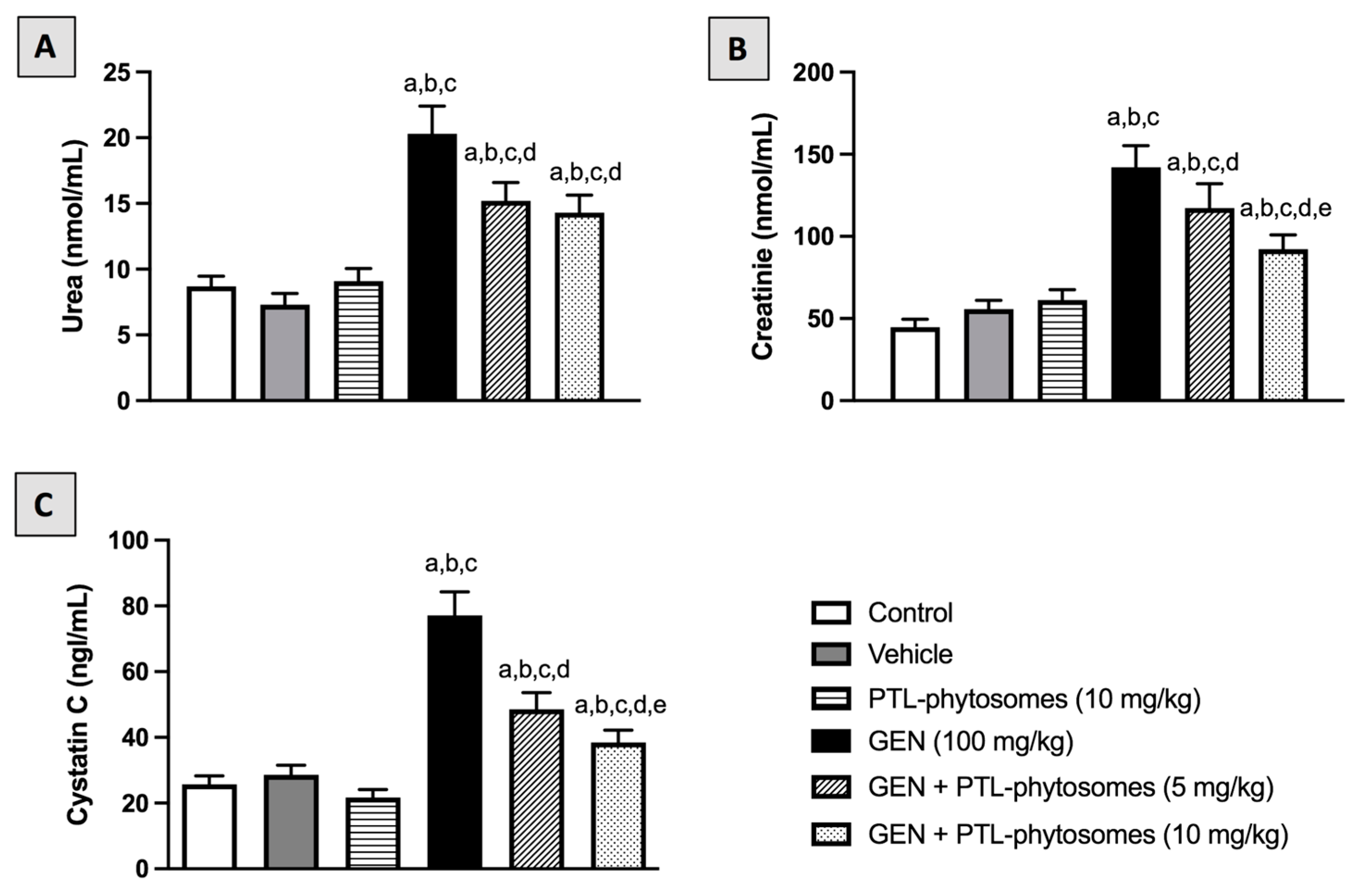
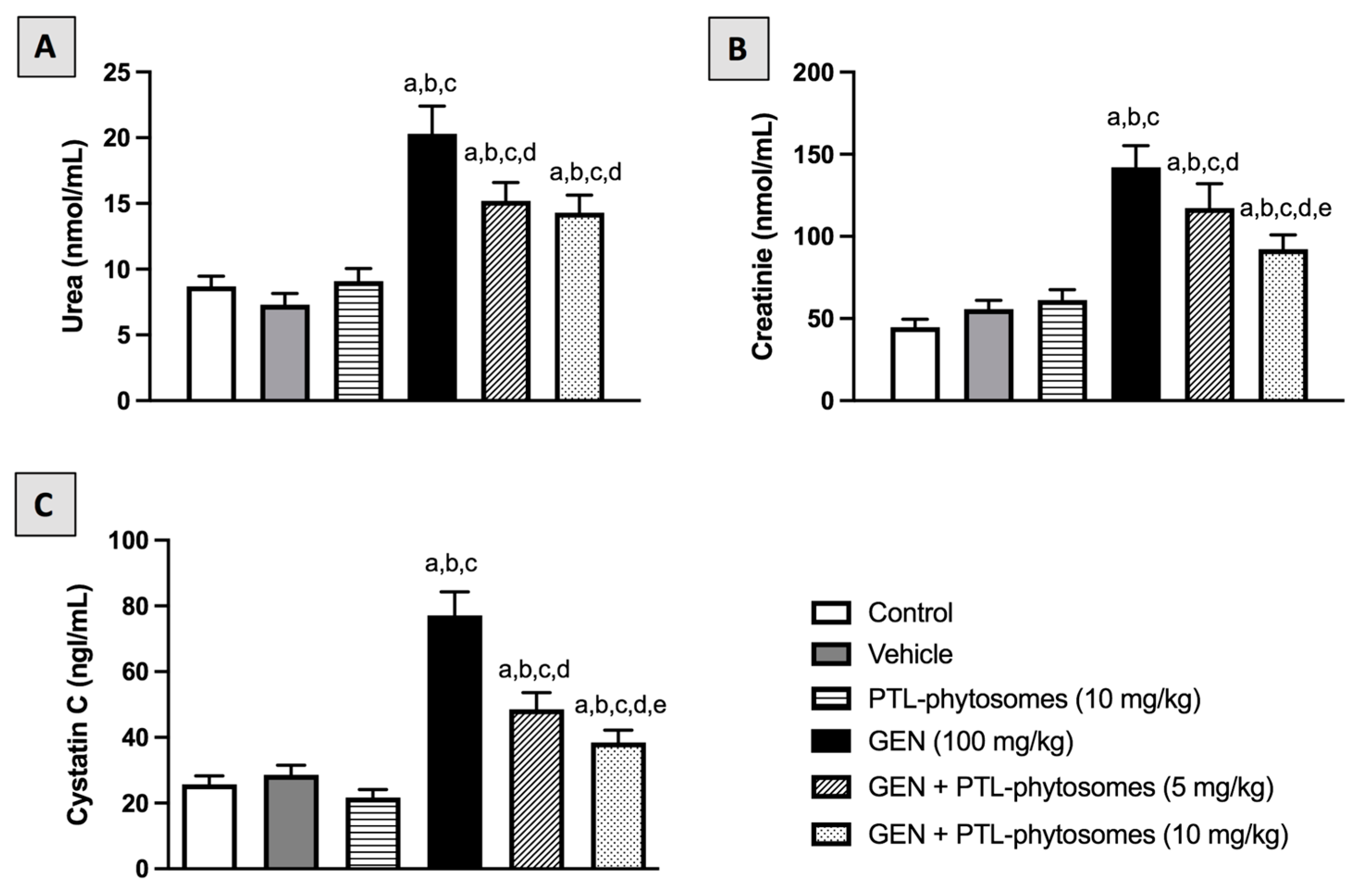
2.3. PTL-Phytosomes Restored the Kidney Function Markers
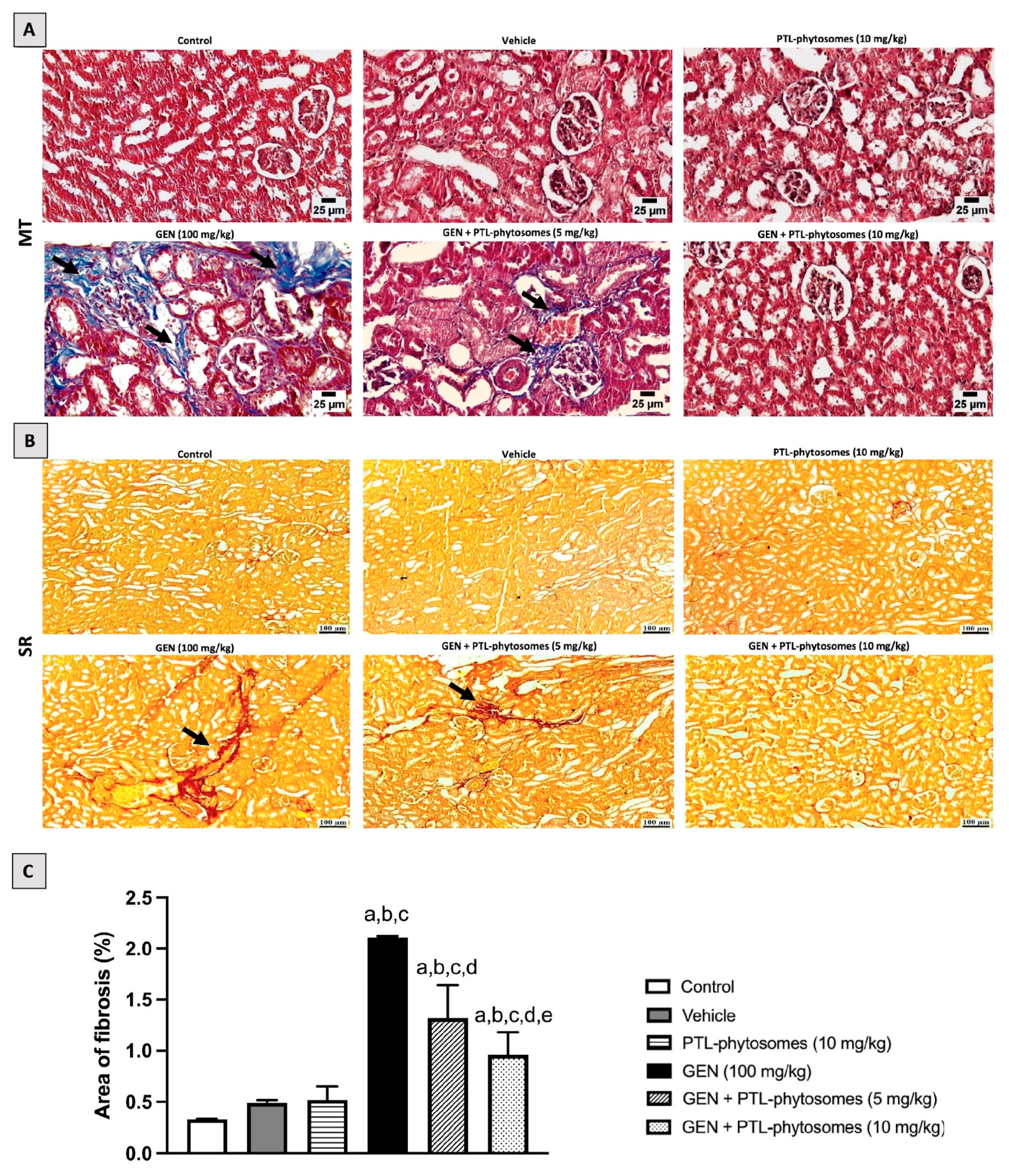
2.4. PTL-Phytosomes Ameliorated, Renal Histopathological Changes, Induced by GEN Treatment
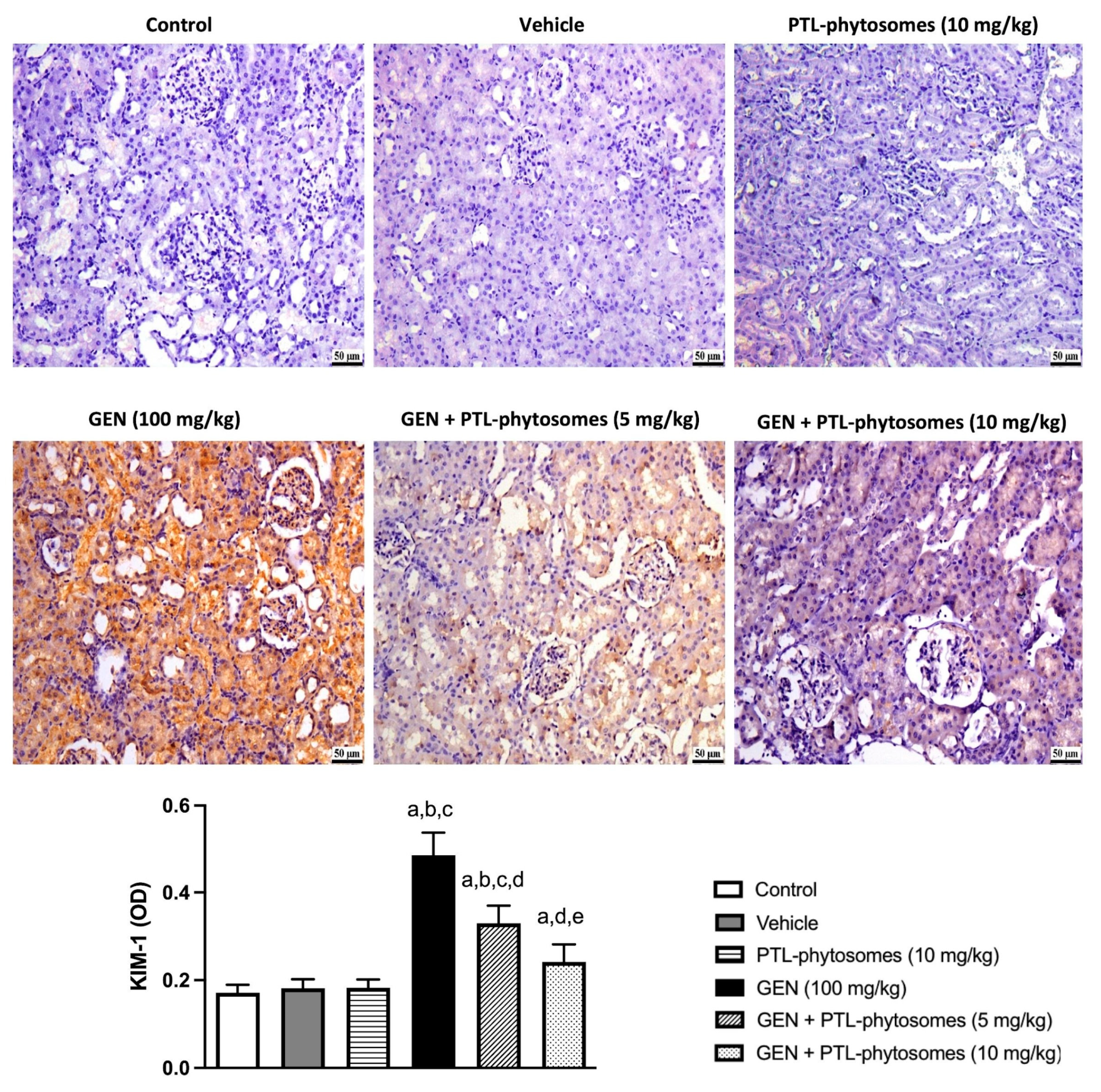
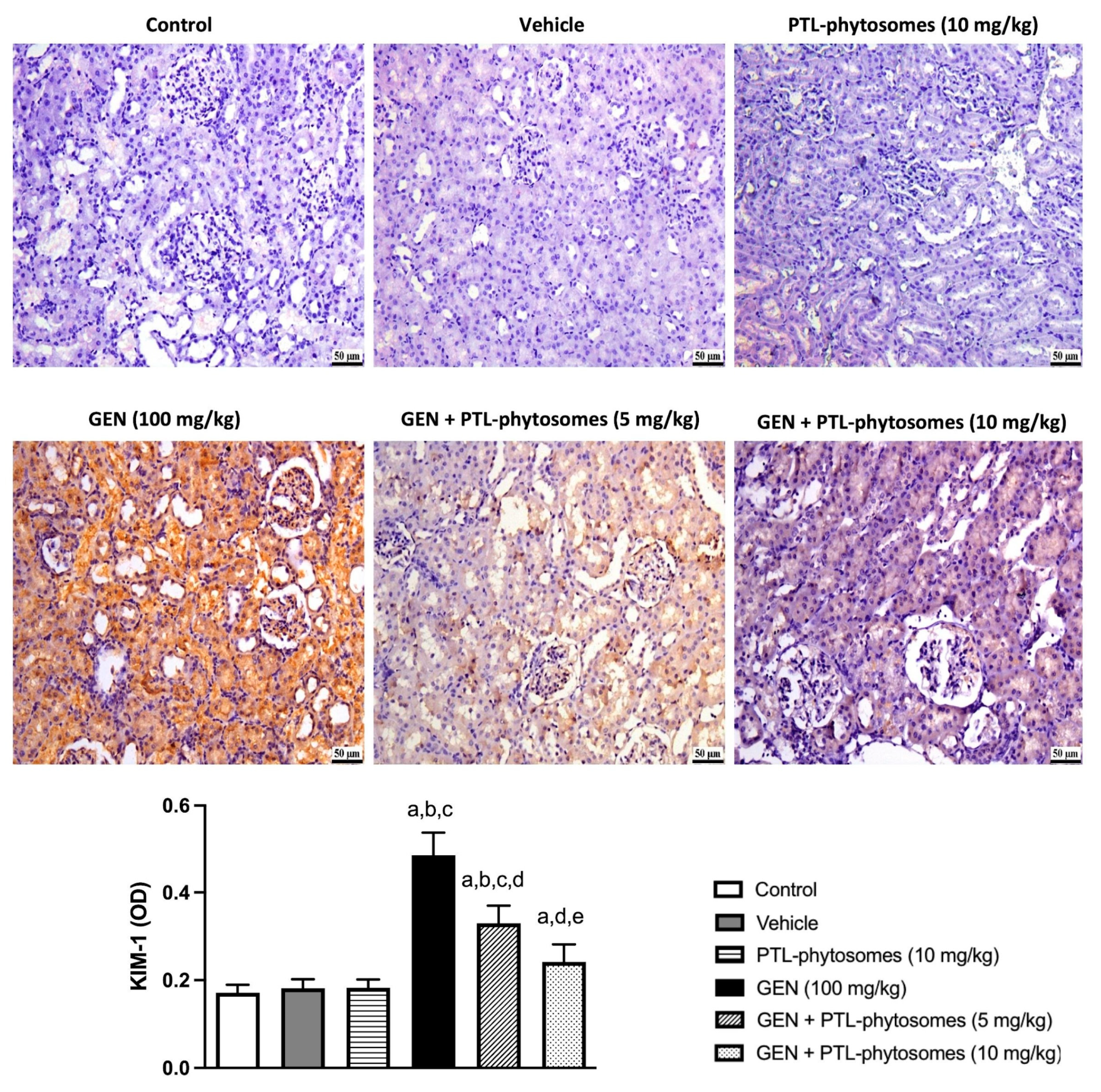
2.5. PTL-Phytosomes Ameliorated GEN-Induced Kidney Injury
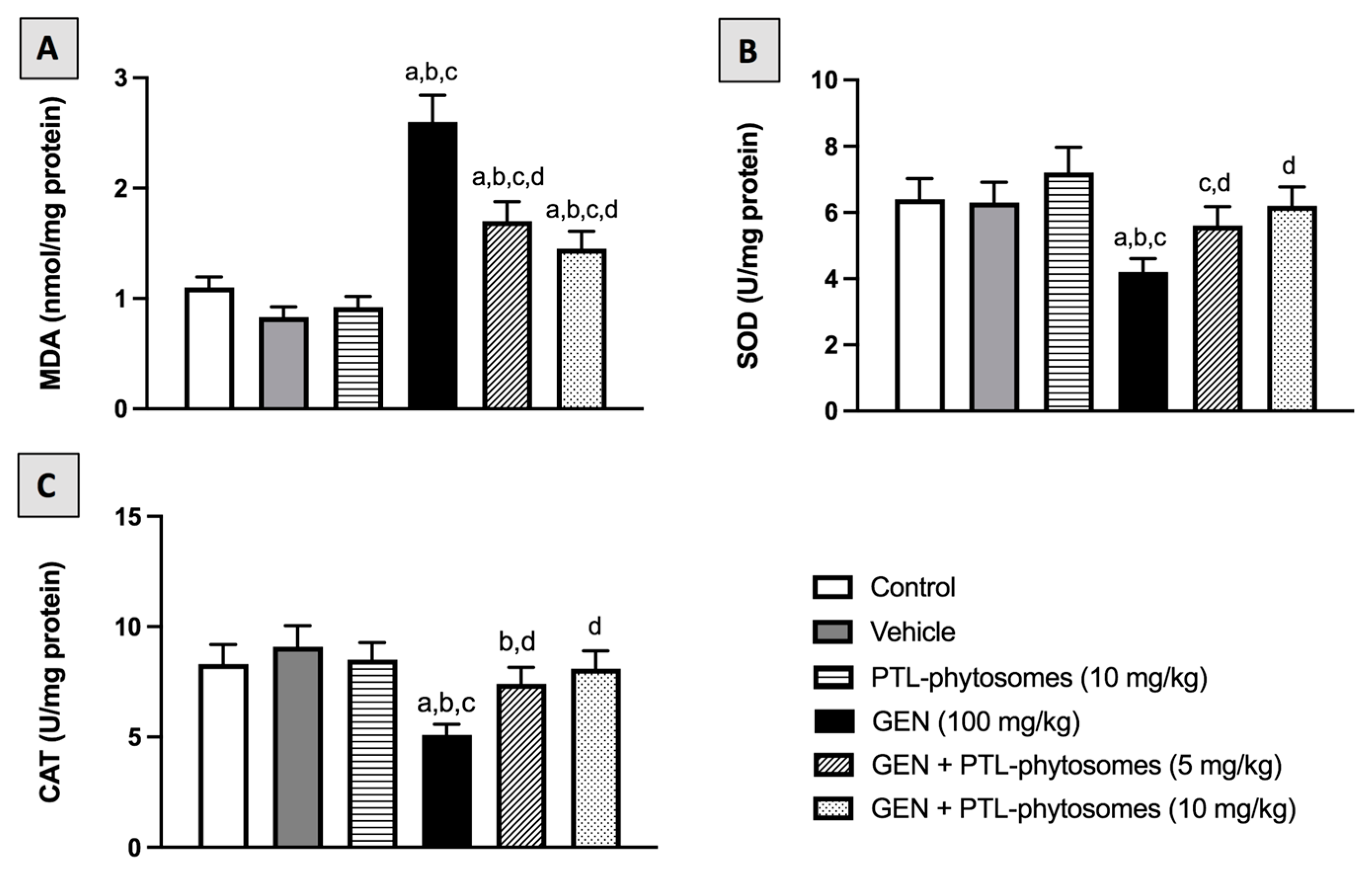
2.6. PTL-Phytosomes Attenuated Oxidative Stress in GEN-Treated Rats
2.7. PTL-Phytosomes Suppressed GEN-Induced Oxidative Stress via Activation of Sirt-1, Nrf2, NQO1, and OH-1 Axis
2.8. PTL-Phytosomes Protected the Kidney against GEN-Induced Inflammation
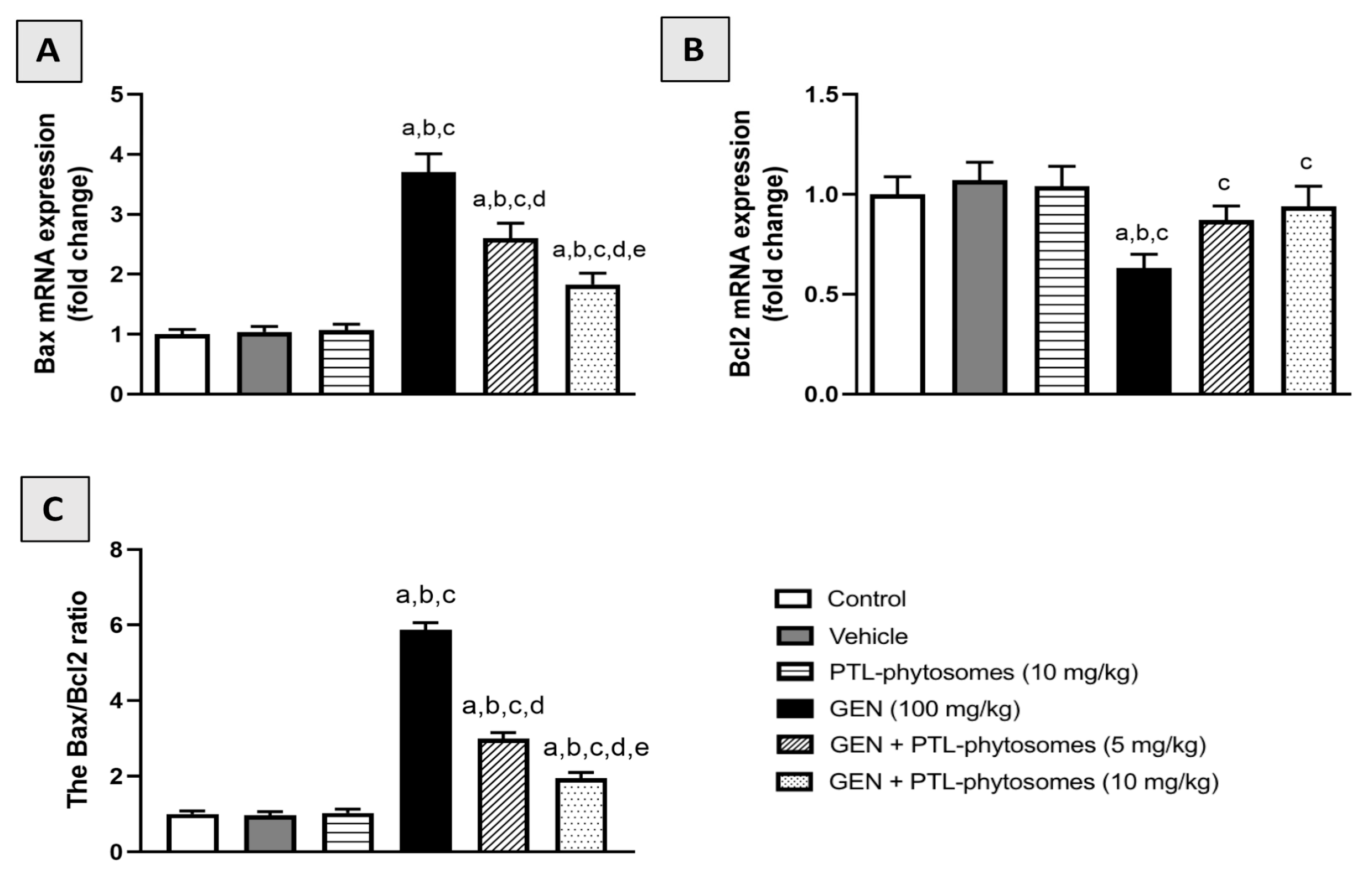
2.9. PTL-Phytosomes Ameliorated GEN-Induced Kidney Apoptosis
3. Discussion
4. Materials and Methods
4.1. Chemicals and Drugs
4.2. Animals
4.3. Preparation of PTL-Phytosomes
4.4. Characterization of PTL-Phytosomes
4.4.1. Particle Size
4.4.2. Transmission Electron Microscopy
4.5. Experimental Design
4.6. Evaluation of Renal Function Markers
4.7. Histopathological Study
4.8. Evaluation of Oxidative Status Markers
4.9. Western Blot Analysis
4.10. Immunohistochemical Analysis
4.11. Real-Time Polymerase Chain Reaction (RT-PCR)
4.12. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
Abbreviations
References
- Downes, K.J.; Hayes, M.; Fitzgerald, J.C.; Pais, G.M.; Liu, J.; Zane, N.R.; Goldstein, S.L.; Scheetz, M.H.; Zuppa, A.F. Mechanisms of Antimicrobial-Induced Nephrotoxicity in Children. J. Antimicrob. Chemother. 2020, 75, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quirós, Y.; Blanco-Gozalo, V.; Sanchez-Gallego, J.I.; López-Hernandez, F.J.; Ruiz, J.; Perez de Obanos, M.P.; López-Novoa, J.M. Cardiotrophin-1 Therapy Prevents Gentamicin-Induced Nephrotoxicity in Rats. Pharmacol. Res. 2016, 107, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Huang, J. Drug-Induced Nephrotoxicity: Pathogenic Mechanisms, Biomarkers and Prevention Strategies. Curr. Drug Metab. 2018, 19, 559–567. [Google Scholar] [CrossRef]
- Juan, S.-H.; Chen, C.-H.; Hsu, Y.-H.; Hou, C.-C.; Chen, T.-H.; Lin, H.; Chu, Y.-L.; Sue, Y.-M. Tetramethylpyrazine Protects Rat Renal Tubular Cell Apoptosis Induced by Gentamicin. Nephrol. Dial. Transplant. 2007, 22, 732–739. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, C.; Lee, H.; Hah, D.-Y.; Heo, J.H.; Kim, C.H.; Kim, E.; Kim, J.S. Protective Effects of Houttuynia cordata Thunb. on Gentamicin-Induced Oxidative Stress and Nephrotoxicity in Rats. Toxicol. Res. 2013, 29, 61–67. [Google Scholar] [CrossRef] [Green Version]
- Bledsoe, G.; Crickman, S.; Mao, J.; Xia, C.-F.; Murakami, H.; Chao, L.; Chao, J. Kallikrein/Kinin Protects against Gentamicin-Induced Nephrotoxicity by Inhibition of Inflammation and Apoptosis. Nephrol. Dial. Transplant. 2006, 21, 624–633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Jin, W.W.; Huang, M.; Ji, H.; Capen, D.E.; Xia, Y.; Yuan, J.; Păunescu, T.G.; Lu, H.A.J. Gentamicin-Induced Acute Kidney Injury in an Animal Model Involves Programmed Necrosis of the Collecting Duct. J. Am. Soc. Nephrol. 2020, 31, 2097–2115. [Google Scholar] [CrossRef]
- Medić, B.; Stojanović, M.; Rovčanin, B.; Kekić, D.; Škodrić, S.R.; Jovanović, G.B.; Vujović, K.S.; Divac, N.; Stojanović, R.; Radenković, M.; et al. Pioglitazone Attenuates Kidney Injury in an Experimental Model of Gentamicin-Induced Nephrotoxicity in Rats. Sci. Rep. 2019, 9, 13689. [Google Scholar] [CrossRef] [Green Version]
- Morales, A.I.; Detaille, D.; Prieto, M.; Puente, A.; Briones, E.; Arévalo, M.; Leverve, X.; López-Novoa, J.M.; El-Mir, M.-Y. Metformin Prevents Experimental Gentamicin-Induced Nephropathy by a Mitochondria-Dependent Pathway. Kidney Int. 2010, 77, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Priyamvada, S.; Farooq, N.; Khan, S.; Khan, M.W.; Yusufi, A.N.K. Protective Effect of Green Tea Extract on Gentamicin-Induced Nephrotoxicity and Oxidative Damage in Rat Kidney. Pharmacol. Res. 2009, 59, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Sundin, D.P.; Sandoval, R.; Molitoris, B.A. Gentamicin Inhibits Renal Protein and Phospholipid Metabolism in Rats: Implications Involving Intracellular Trafficking. J. Am. Soc. Nephrol. 2001, 12, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Vysakh, A.; Abhilash, S.; Kuriakose, J.; Midhun, S.J.; Jyothis, M.; Latha, M.S. Protective Effect of Rotula aquatica Lour against Gentamicin Induced Oxidative Stress and Nephrotoxicity in Wistar Rats. Biomed. Pharmacother. 2018, 106, 1188–1194. [Google Scholar] [CrossRef]
- Fan, X.; Gu, W.; Gao, Y.; Ma, N.; Fan, C.; Ci, X. Daphnetin Ameliorated GM-Induced Renal Injury through the Suppression of Oxidative Stress and Apoptosis in Mice. Int. Immunopharmacol. 2021, 96, 107601. [Google Scholar] [CrossRef]
- Helal, M.G.; Zaki, M.M.A.F.; Said, E. Nephroprotective Effect of Saxagliptin against Gentamicin-Induced Nephrotoxicity, Emphasis on Anti-Oxidant, Anti-Inflammatory and Anti-Apoptic Effects. Life Sci. 2018, 208, 64–71. [Google Scholar] [CrossRef]
- Sharma, V.C.; Kaushik, A.; Dey, Y.N.; Srivastava, B.; Wanjari, M.; Pawar, S.; Chougule, S. Nephroprotective Potential of Anogeissus latifolia Roxb. (Dhava) against Gentamicin-Induced Nephrotoxicity in Rats. J. Ethnopharmacol. 2021, 273, 114001. [Google Scholar] [CrossRef] [PubMed]
- Shoaib, M.; Shah, I.; Ali, N.; Adhikari, A.; Tahir, M.N.; Shah, S.W.A.; Ishtiaq, S.; Khan, J.; Khan, S.; Umer, M.N. Sesquiterpene Lactone! A Promising Antioxidant, Anticancer and Moderate Antinociceptive Agent from Artemisia macrocephala Jacquem. BMC Complement. Altern. Med. 2017, 17, 27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Freund, R.R.A.; Gobrecht, P.; Fischer, D.; Arndt, H.-D. Advances in Chemistry and Bioactivity of Parthenolide. Nat. Prod. Rep. 2020, 37, 541–565. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.Y.; Kang, B.; Hong, J.; Choi, H.-S. Parthenolide Inhibits Lipid Accumulation via Activation of Nrf2/Keap1 Signaling during Adipocyte Differentiation. Food Sci. Biotechnol. 2020, 29, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Huang, Q.; Chen, Y.; Peng, X.; Wang, Y.; Li, S.; Wu, J.; Luo, C.; Gong, W.; Yin, B.; et al. Parthenolide, an NF-κB Inhibitor, Alleviates Peritoneal Fibrosis by Suppressing the TGF-β/Smad Pathway. Int. Immunopharmacol. 2020, 78, 106064. [Google Scholar] [CrossRef]
- Wang, J.; Tong, M.; Zhao, B.; Zhu, G.; Xi, D.; Yang, J. Parthenolide Ameliorates Intracerebral Hemorrhage-induced Brain Injury in Rats. Phytother. Res. 2020, 34, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Francescato, H.D.C.; Costa, R.S.; Scavone, C.; Coimbra, T.M. Parthenolide Reduces Cisplatin-Induced Renal Damage. Toxicology 2007, 230, 64–75. [Google Scholar] [CrossRef]
- Kansanen, E.; Kuosmanen, S.M.; Leinonen, H.; Levonen, A.-L. The Keap1-Nrf2 Pathway: Mechanisms of Activation and Dysregulation in Cancer. Redox Biol. 2013, 1, 45–49. [Google Scholar] [CrossRef] [Green Version]
- Zoja, C.; Benigni, A.; Remuzzi, G. The Nrf2 Pathway in the Progression of Renal Disease. Nephrol. Dial. Transplant. 2014, 29, i19–i24. [Google Scholar] [CrossRef] [Green Version]
- Huang, K.; Gao, X.; Wei, W. The Crosstalk between Sirt1 and Keap1/Nrf2/ARE Anti-Oxidative Pathway Forms a Positive Feedback Loop to Inhibit FN and TGF-β1 Expressions in Rat Glomerular Mesangial Cells. Exp. Cell Res. 2017, 361, 63–72. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Peng, X.; Zhu, J.; Liu, G.; Chen, X.; Tang, C.; Liu, H.; Liu, F.; Peng, Y. Protective Effects of Curcumin on Acute Gentamicin-Induced Nephrotoxicity in Rats. Can. J. Physiol. Pharmacol. 2015, 93, 275–282. [Google Scholar] [CrossRef] [PubMed]
- Nadeem, R.I.; Aboutaleb, A.S.; Younis, N.S.; Ahmed, H.I. Diosmin Mitigates Gentamicin-Induced Nephrotoxicity in Rats: Insights on MiR-21 and -155 Expression, Nrf2/HO-1 and P38-MAPK/NF-κB Pathways. Toxics 2023, 11, 48. [Google Scholar] [CrossRef]
- Subramanian, P.; Anandan, R.; Jayapalan, J.J.; Hashim, O.H. Hesperidin Protects Gentamicin-Induced Nephrotoxicity via Nrf2/HO-1 Signaling and Inhibits Inflammation Mediated by NF-κB in Rats. J. Funct. Foods 2015, 13, 89–99. [Google Scholar] [CrossRef]
- Magni, P.; Ruscica, M.; Dozio, E.; Rizzi, E.; Beretta, G.; Facino, R.M. Parthenolide Inhibits the LPS-Induced Secretion of IL-6 and TNF-α and NF-κB Nuclear Translocation in BV-2 Microglia: Parthenolide inhibition of BV-2 microglia. Phytother. Res. 2012, 26, 1405–1409. [Google Scholar] [CrossRef]
- Zhang, H.; Sun, S.-C. NF-κB in Inflammation and Renal Diseases. Cell Biosci. 2015, 5, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.-C. NF-κB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li-Weber, M.; Giaisi, M.; Baumann, S.; Treiber, M.K.; Krammer, P.H. The Anti-Inflammatory Sesquiterpene Lactone Parthenolide Suppresses CD95-Mediated Activation-Induced-Cell-Death in T-Cells. Cell Death Differ. 2002, 9, 1256–1265. [Google Scholar] [CrossRef] [PubMed]
- Yao, H.; Tang, X.; Shao, X.; Feng, L.; Wu, N.; Yao, K. Parthenolide Protects Human Lens Epithelial Cells from Oxidative Stress-Induced Apoptosis via Inhibition of Activation of Caspase-3 and Caspase-9. Cell Res. 2007, 17, 565–571. [Google Scholar] [CrossRef] [PubMed]
- Taleghani, A.; Nasseri, M.A.; Iranshahi, M. Synthesis of Dual-Action Parthenolide Prodrugs as Potent Anticancer Agents. Bioorganic Chem. 2017, 71, 128–134. [Google Scholar] [CrossRef] [PubMed]
- Zong, H.; Sen, S.; Zhang, G.; Mu, C.; Albayati, Z.F.; Gorenstein, D.G.; Liu, X.; Ferrari, M.; Crooks, P.A.; Roboz, G.J.; et al. In Vivo Targeting of Leukemia Stem Cells by Directing Parthenolide-Loaded Nanoparticles to the Bone Marrow Niche. Leukemia 2016, 30, 1582–1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, W.; Li, L.; Zhang, X.; Luo, L.; He, Y.; Cong, C.; Gao, D. Nanomagnetic Liposome-Encapsulated Parthenolide and Indocyanine Green for Targeting and Chemo-Photothermal Antitumor Therapy. Nanomedicine 2020, 15, 871–890. [Google Scholar] [CrossRef]
- Darwish, N.H.E.; Sudha, T.; Godugu, K.; Bharali, D.J.; Elbaz, O.; El-ghaffar, H.A.A.; Azmy, E.; Anber, N.; Mousa, S.A. Novel Targeted Nano-Parthenolide Molecule against NF-KB in Acute Myeloid Leukemia. Molecules 2019, 24, 2103. [Google Scholar] [CrossRef] [Green Version]
- Liang, P.; Wu, H.; Zhang, Z.; Jiang, S.; Lv, H. Preparation and Characterization of Parthenolide Nanocrystals for Enhancing Therapeutic Effects of Sorafenib against Advanced Hepatocellular Carcinoma. Int. J. Pharm. 2020, 583, 119375. [Google Scholar] [CrossRef]
- Liu, Y.; Lu, W.-L.; Guo, J.; Du, J.; Li, T.; Wu, J.-W.; Wang, G.-L.; Wang, J.-C.; Zhang, X.; Zhang, Q. A Potential Target Associated with Both Cancer and Cancer Stem Cells: A Combination Therapy for Eradication of Breast Cancer Using Vinorelbine Stealthy Liposomes plus Parthenolide Stealthy Liposomes. J. Control. Release 2008, 129, 18–25. [Google Scholar] [CrossRef] [PubMed]
- Anjana, R.; Kumar, S.; Sharma, H.; Khar, R. Phytosome Drug Delivery of Natural Products: A Promising Technique for Enhancing Bioavailability. Int. J. Drug Deliv. Technol. 2017, 7, 157–165. [Google Scholar] [CrossRef]
- Lu, M.; Qiu, Q.; Luo, X.; Liu, X.; Sun, J.; Wang, C.; Lin, X.; Deng, Y.; Song, Y. Phyto-Phospholipid Complexes (Phytosomes): A Novel Strategy to Improve the Bioavailability of Active Constituents. Asian J. Pharm. Sci. 2019, 14, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Park, S.; Kang, M.; Park, M.; Choi, Y.W. Folic Acid-Tethered Pep-1 Peptide-Conjugated Liposomal Nanocarrier for Enhanced Intracellular Drug Delivery to Cancer Cells: Conformational Characterization and in Vitro Cellular Uptake Evaluation. Int. J. Nanomed. 2013, 2013, 1155–1165. [Google Scholar] [CrossRef] [Green Version]
- Montenegro, L.; Santagati, L.M.; Sarpietro, M.G.; Castelli, F.; Panico, A.; Siciliano, E.A.; Lai, F.; Valenti, D.; Sinico, C. In Vitro Skin Permeation of Idebenone from Lipid Nanoparticles Containing Chemical Penetration Enhancers. Pharmaceutics 2021, 13, 1027. [Google Scholar] [CrossRef] [PubMed]
- Mudalige, T.; Qu, H.; Van Haute, D.; Ansar, S.M.; Paredes, A.; Ingle, T. Characterization of Nanomaterials. In Nanomaterials for Food Applications; Elsevier: Amsterdam, The Netherlands, 2019; pp. 313–353. ISBN 978-0-12-814130-4. [Google Scholar]
- Barani, M.; Sangiovanni, E.; Angarano, M.; Rajizadeh, M.A.; Mehrabani, M.; Piazza, S.; Gangadharappa, H.V.; Pardakhty, A.; Mehrbani, M.; Dell’Agli, M.; et al. Phytosomes as Innovative Delivery Systems for Phytochemicals: A Comprehensive Review of Literature. Int. J. Nanomed. 2021, 16, 6983–7022. [Google Scholar] [CrossRef]
- Ledda, A.; Belcaro, G.; Feragalli, B.; Hosoi, M.; Cacchio, M.; Luzzi, R.; Dugall, M.; Cotellese, R. Temporary Kidney Dysfunction: Supplementation with Meriva® in Initial, Transient Kidney Micro-Macro Albuminuria. Panminerva Med. 2020, 61, 444–448. [Google Scholar] [CrossRef]
- Kumar, A.B.; Habbu, P.; Hullatti, P.; Kumar, R.S. Phytosomes as Novel Drug Delivery System for Herbal Medicine—A Review. Syst. Rev. Pharm. 2016, 8, 5–7. [Google Scholar] [CrossRef]
- Amin, T.; Bhat, S.V. A Review on Phytosome Technology as a Novel Approach to Improve the Bioavailability of Nutraceuticals. Int. J. Adv. Res. Technol. 2012, 1, 43–57. [Google Scholar]
- Ma, Y.; Luo, X.; Wu, Y.; Zhang, T.; Zhang, F.; Zhang, G.; Wu, X. Alteration of Renal Excretion Pathways in Gentamicin-Induced Renal Injury in Rats: Renal Excretion Pathway Alterations in GEN-Induced Renal Injury Rats. J. Appl. Toxicol. 2018, 38, 968–977. [Google Scholar] [CrossRef] [PubMed]
- Jaikumkao, K.; Pongchaidecha, A.; Thongnak, L.; Wanchai, K.; Arjinajarn, P.; Chatsudthipong, V.; Chattipakorn, N.; Lungkaphin, A. Amelioration of Renal Inflammation, Endoplasmic Reticulum Stress and Apoptosis Underlies the Protective Effect of Low Dosage of Atorvastatin in Gentamicin-Induced Nephrotoxicity. PLoS ONE 2016, 11, e0164528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hao, Q.F.; Wang, B.B.; Zhang, W.; Qiu, W.; Liu, Q.L.; Li, X.M. NF-κB Inhibitor Parthenolide Promotes Renal Tubules Albumin Uptake in Type 2 Diabetic Nephropathy. Chin. Med. Sci. J. Chung-Kuo Hsueh Ko Hsueh Tsa Chih 2020, 35, 31–42. [Google Scholar] [CrossRef]
- Zhou, Y.; Vaidya, V.S.; Brown, R.P.; Zhang, J.; Rosenzweig, B.A.; Thompson, K.L.; Miller, T.J.; Bonventre, J.V.; Goering, P.L. Comparison of Kidney Injury Molecule-1 and Other Nephrotoxicity Biomarkers in Urine and Kidney Following Acute Exposure to Gentamicin, Mercury, and Chromium. Toxicol. Sci. 2008, 101, 159–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waanders, F.; van Timmeren, M.M.; Stegeman, C.A.; Bakker, S.J.; van Goor, H. Kidney Injury Molecule-1 in Renal Disease: Kidney Injury Molecule-1 in Renal Disease. J. Pathol. 2010, 220, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.; Chen, M.; Sun, F.; Chen, Z.; Li, M.; Zeng, W.; Gong, L.; Cheng, A.; Peng, X.; Fang, J.; et al. KIM-1 and NGAL as Biomarkers of Nephrotoxicity Induced by Gentamicin in Rats. Mol. Cell. Biochem. 2014, 397, 53–60. [Google Scholar] [CrossRef]
- Adil, M.; Kandhare, A.D.; Dalvi, G.; Ghosh, P.; Venkata, S.; Raygude, K.S.; Bodhankar, S.L. Ameliorative Effect of Berberine against Gentamicin-Induced Nephrotoxicity in Rats via Attenuation of Oxidative Stress, Inflammation, Apoptosis and Mitochondrial Dysfunction. Ren. Fail. 2016, 38, 996–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.; Yang, H.; Ruan, H.; Li, W.; He, X.; Wang, L.; Liu, F.; Zhang, J. The Protective Effect of Sika Deer Antler Protein on Gentamicin-Induced Nephrotoxicity in Vitro and in Vivo. Cell. Physiol. Biochem. 2018, 50, 841–850. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Novoa, J.M.; Quiros, Y.; Vicente, L.; Morales, A.I.; Lopez-Hernandez, F.J. New Insights into the Mechanism of Aminoglycoside Nephrotoxicity: An Integrative Point of View. Kidney Int. 2011, 79, 33–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales, A.I.; Buitrago, J.M.; Santiago, J.M.; Fernández-Tagarro, M.; López-Novoa, J.M.; Pérez-Barriocanal, F. Protective Effect of trans-Resveratrol on Gentamicin-Induced Nephrotoxicity. Antioxid. Redox Signal. 2002, 4, 893–898. [Google Scholar] [CrossRef]
- Randjelović, P.; Veljković, S.; Stojiljković, N.; Sokolović, D.; Ilić, I. Gentamicin Nephrotoxicity in Animals: Current Knowledge and Future Perspectives. EXCLI J. 2017, 16, 388–399. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Chen, T.; Hao, Z.; Yang, X.; Wang, M.; Zhang, Z.; Hao, S.; Lang, F.; Hao, H. Oxymatrine Alleviates Gentamicin-Induced Renal Injury in Rats. Molecules 2022, 27, 6209. [Google Scholar] [CrossRef] [PubMed]
- Sharawy, M.H.; Serrya, M.S. Pirfenidone Attenuates Gentamicin-Induced Acute Kidney Injury by Inhibiting Inflammasome-Dependent NLRP3 Pathway in Rats. Life Sci. 2020, 260, 118454. [Google Scholar] [CrossRef]
- Li-Weber, M.; Palfi, K.; Giaisi, M.; Krammer, P.H. Dual Role of the Anti-Inflammatory Sesquiterpene Lactone: Regulation of Life and Death by Parthenolide. Cell Death Differ. 2005, 12, 408–409. [Google Scholar] [CrossRef]
- Herrera, F.; Martin, V.; Rodriguez-Blanco, J.; García-Santos, G.; Antolín, I.; Rodriguez, C. Intracellular Redox State Regulation by Parthenolide. Biochem. Biophys. Res. Commun. 2005, 332, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Mao, W.; Zhu, Z. Parthenolide Inhibits Hydrogen Peroxide-induced Osteoblast Apoptosis. Mol. Med. Rep. 2018, 17, 8369–8376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, K.; Huang, J.; Xie, X.; Wang, S.; Chen, C.; Shen, X.; Liu, P.; Huang, H. Sirt1 Resists Advanced Glycation End Products-Induced Expressions of Fibronectin and TGF-β1 by Activating the Nrf2/ARE Pathway in Glomerular Mesangial Cells. Free Radic. Biol. Med. 2013, 65, 528–540. [Google Scholar] [CrossRef] [PubMed]
- Zaky, H.S.; Abdel-Sattar, S.A.; Allam, A.; Ahmed, H.I. Further Insights into the Impact of Rebamipide on Gentamicin-Induced Nephrotoxicity in Rats: Modulation of SIRT1 and β-Catenin/Cyclin D1 Pathways. Drug Chem. Toxicol. 2022, 45, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Sue, Y.-M.; Cheng, C.-F.; Chang, C.-C.; Chou, Y.; Chen, C.-H.; Juan, S.-H. Antioxidation and Anti-Inflammation by Haem Oxygenase-1 Contribute to Protection by Tetramethylpyrazine against Gentamicin-Induced Apoptosis in Murine Renal Tubular Cells. Nephrol. Dial. Transplant. 2008, 24, 769–777. [Google Scholar] [CrossRef] [Green Version]
- Kalayarasan, S.; Prabhu, P.N.; Sriram, N.; Manikandan, R.; Arumugam, M.; Sudhandiran, G. Diallyl Sulfide Enhances Antioxidants and Inhibits Inflammation through the Activation of Nrf2 against Gentamicin-Induced Nephrotoxicity in Wistar Rats. Eur. J. Pharmacol. 2009, 606, 162–171. [Google Scholar] [CrossRef]
- Tugcu, V.; Ozbek, E.; Tasci, A.I.; Kemahli, E.; Somay, A.; Bas, M.; Karaca, C.; Altug, T.; Cekmen, M.B.; Ozdogan, H.K. Selective Nuclear Factor κ-B Inhibitors, Pyrolidium Dithiocarbamate and Sulfasalazine, Prevent the Nephrotoxicity Induced by Gentamicin. BJU Int. 2006, 98, 680–686. [Google Scholar] [CrossRef]
- Amigó, M.; Payá, M.; Braza-Boïls, A.; De Rosa, S.; Terencio, M.C. Avarol Inhibits TNF-α Generation and NF-κB Activation in Human Cells and in Animal Models. Life Sci. 2008, 82, 256–264. [Google Scholar] [CrossRef]
- Abdelrahman, R.S.; Abdelmageed, M.E. Renoprotective Effect of Celecoxib against Gentamicin-Induced Nephrotoxicity through Suppressing NFκB and Caspase-3 Signaling Pathways in Rats. Chem. Biol. Interact. 2020, 315, 108863. [Google Scholar] [CrossRef]
- Liu, Q.; Zhang, L.; Zhang, W.; Hao, Q.; Qiu, W.; Wen, Y.; Wang, H.; Li, X. Inhibition of NF-κB Reduces Renal Inflammation and Expression of PEPCK in Type 2 Diabetic Mice. Inflammation 2018, 41, 2018–2029. [Google Scholar] [CrossRef] [Green Version]
- López-Franco, O.; Hernández-Vargas, P.; Ortiz-Muñoz, G.; Sanjuán, G.; Suzuki, Y.; Ortega, L.; Blanco, J.; Egido, J.; Gómez-Guerrero, C. Parthenolide Modulates the NF-κB–Mediated Inflammatory Responses in Experimental Atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2006, 26, 1864–1870. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.J.; Xiang, J.Y.; Liu, L.; Huang, X.L.; Gan, H.T. Parthenolide, an Inhibitor of the Nuclear Factor-κB Pathway, Ameliorates Dextran Sulfate Sodium-Induced Colitis in Mice. Int. Immunopharmacol. 2012, 12, 169–174. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.L.; Liu, Y.C.; Seo, S.Y.; Kim, S.H.; Kim, I.H.; Lee, S.O.; Lee, S.T.; Kim, D.-G.; Kim, S.W. Parthenolide Induces Apoptosis in Colitis-Associated Colon Cancer, Inhibiting NF-κB Signaling. Oncol. Lett. 2015, 9, 2135–2142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Xiao, T.; Yang, J.; Qin, Y.; Gao, J.; Liu, H.; Zhou, H. Parthenolide Attenuated Bleomycin-Induced Pulmonary Fibrosis via the NF-κB/Snail Signaling Pathway. Respir. Res. 2018, 19, 111. [Google Scholar] [CrossRef] [Green Version]
- Zingarelli, B.; Hake, P.W.; Denenberg, A.; Wong, H.R. Sesquiterpene Lactone Parthenolide, an Inhibitor of IκB Kinase Complex and Nuclear Factor-κB, Exerts Beneficial Effects in Myocardial Reperfusion Injury. Shock 2002, 17, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Rummel, C.; Gerstberger, R.; Roth, J.; Hübschle, T. Parthenolide Attenuates LPS-Induced Fever, Circulating Cytokines and Markers of Brain Inflammation in Rats. Cytokine 2011, 56, 739–748. [Google Scholar] [CrossRef]
- Zhang, M.; Liu, R.-T.; Zhang, P.; Zhang, N.; Yang, C.-L.; Yue, L.-T.; Li, X.-L.; Liu, Y.; Li, H.; Du, J.; et al. Parthenolide Inhibits the Initiation of Experimental Autoimmune Neuritis. J. Neuroimmunol. 2017, 305, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Sobota, R.; Szwed, M.; Kasza, A.; Bugno, M.; Kordula, T. Parthenolide Inhibits Activation of Signal Transducers and Activators of Transcription (STATs) Induced by Cytokines of the IL-6 Family. Biochem. Biophys. Res. Commun. 2000, 267, 329–333. [Google Scholar] [CrossRef]
- Kwok, B.H.B.; Koh, B.; Ndubuisi, M.I.; Elofsson, M.; Crews, C.M. The Anti-Inflammatory Natural Product Parthenolide from the Medicinal Herb Feverfew Directly Binds to and Inhibits IκB Kinase. Chem. Biol. 2001, 8, 759–766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.-C.; Chen, C.-H.; Hsu, Y.-H.; Chen, T.-H.; Sue, Y.-M.; Cheng, C.-Y.; Chen, T.-W. Leptin Reduces Gentamicin-Induced Apoptosis in Rat Renal Tubular Cells via the PI3K-Akt Signaling Pathway. Eur. J. Pharmacol. 2011, 658, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Sahu, B.D.; Tatireddy, S.; Koneru, M.; Borkar, R.M.; Kumar, J.M.; Kuncha, M.; R., S.; R., S.S.; Sistla, R. Naringin Ameliorates Gentamicin-Induced Nephrotoxicity and Associated Mitochondrial Dysfunction, Apoptosis and Inflammation in Rats: Possible Mechanism of Nephroprotection. Toxicol. Appl. Pharmacol. 2014, 277, 8–20. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Salgado, C.; Eleno, N.; Morales, A.I.; Pérez-Barriocanal, F.; Arévalo, M.; López-Novoa, J.M. Gentamicin Treatment Induces Simultaneous Mesangial Proliferation and Apoptosis in Rats. Kidney Int. 2004, 65, 2161–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kalkan, Y.; Kapakin, K.A.T.; Kara, A.; Atabay, T.; Karadeniz, A.; Simsek, N.; Karakus, E.; Can, I.; Yildirim, S.; Ozkanlar, S.; et al. Protective Effect of Panax ginseng against Serum Biochemical Changes and Apoptosis in Kidney of Rats Treated with Gentamicin Sulphate. J. Mol. Histol. 2012, 43, 603–613. [Google Scholar] [CrossRef]
- Jado, J.C.; Humanes, B.; González-Nicolás, M.Á.; Camaño, S.; Lara, J.M.; López, B.; Cercenado, E.; García-Bordas, J.; Tejedor, A.; Lázaro, A. Nephroprotective Effect of Cilastatin against Gentamicin-Induced Renal Injury In Vitro and In Vivo without Altering Its Bactericidal Efficiency. Antioxidants 2020, 9, 821. [Google Scholar] [CrossRef] [PubMed]
- Alhakamy, N.A.; Fahmy, U.A.; Badr-Eldin, S.M.; Ahmed, O.A.A.; Asfour, H.Z.; Aldawsari, H.M.; Algandaby, M.M.; Eid, B.G.; Abdel-Naim, A.B.; Awan, Z.A.; et al. Optimized Icariin Phytosomes Exhibit Enhanced Cytotoxicity and Apoptosis-Inducing Activities in Ovarian Cancer Cells. Pharmaceutics 2020, 12, 346. [Google Scholar] [CrossRef] [PubMed]
- Chomczynski, P.; Sacchi, N. Single-Step Method of RNA Isolation by Acid Guanidinium Thiocyanate-Phenol-Chloroform Extraction. Anal. Biochem. 1987, 162, 156–159. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Scores | Severity of Lesions |
|---|---|
| 0 | No pathological change |
| +1 | Mild change |
| +2 | Mild to moderate change |
| +3 | Moderate change |
| +4 | Moderate to severe change |
| +5 | Severe pathological change |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Albalawi, R.S.; Binmahfouz, L.S.; Hareeri, R.H.; Shaik, R.A.; Bagher, A.M. Parthenolide Phytosomes Attenuated Gentamicin-Induced Nephrotoxicity in Rats via Activation of Sirt-1, Nrf2, OH-1, and NQO1 Axis. Molecules 2023, 28, 2741. https://doi.org/10.3390/molecules28062741
Albalawi RS, Binmahfouz LS, Hareeri RH, Shaik RA, Bagher AM. Parthenolide Phytosomes Attenuated Gentamicin-Induced Nephrotoxicity in Rats via Activation of Sirt-1, Nrf2, OH-1, and NQO1 Axis. Molecules. 2023; 28(6):2741. https://doi.org/10.3390/molecules28062741
Chicago/Turabian StyleAlbalawi, Rawan S., Lenah S. Binmahfouz, Rawan H. Hareeri, Rasheed A. Shaik, and Amina M. Bagher. 2023. "Parthenolide Phytosomes Attenuated Gentamicin-Induced Nephrotoxicity in Rats via Activation of Sirt-1, Nrf2, OH-1, and NQO1 Axis" Molecules 28, no. 6: 2741. https://doi.org/10.3390/molecules28062741
APA StyleAlbalawi, R. S., Binmahfouz, L. S., Hareeri, R. H., Shaik, R. A., & Bagher, A. M. (2023). Parthenolide Phytosomes Attenuated Gentamicin-Induced Nephrotoxicity in Rats via Activation of Sirt-1, Nrf2, OH-1, and NQO1 Axis. Molecules, 28(6), 2741. https://doi.org/10.3390/molecules28062741