Abstract
Laccases have been widely used for fruit juice clarification, food modification, and paper pulp delignification. In addition, laccases exhibit remarkable performance in the degradation of toxic substances, including pesticides, organic synthetic dyes, antibiotics, and organic pollutants. Thus, the screening and development of robust laccases has attracted significant attention. In this study, Vibrio sp. LA is a strain capable of producing cold-adapted laccases. The laccase coding gene L01 was cloned from this strain and expressed in Yarrowia lipolytica, a host with good secretion ability. The secreted L01 (approximate MW of 56,000 Da) had the activity and specific activity of 18.6 U/mL and 98.6 U/mg toward ABTS, respectively. The highest activity occurred at 35 °C. At 20 °C, L01 activity was over 70% of the maximum activity in pH conditions ranging from 4.5–10.0. Several synthetic dyes were efficiently degraded by L01. Owing to its robustness, salt tolerance, and pH stability, L01 is a promising catalytic tool for potential industrial applications.
1. Introduction
Laccases (EC 1.10.3.2), a class of polyphenol oxidases, are promising biocatalysts that are independent of cofactors [1]. They can oxidize various phenolic substrates and other aromatic compounds with the concurrent formation of water. Owing to their unique characteristics, laccases have been widely used in fruit juice clarification, food modification, and paper pulp delignification [1,2]. More importantly, they degrade toxic substances, including pesticides, organic synthetic dyes, antibiotics, and organic pollutants [3,4].
Laccases are already used in large scale in the textile industry. Together with low molecular weight redox-mediator compounds, laccases can generate a desired worn appearance on denim by bleaching indigo dye [2,3]. The potential use of laccase for bleaching has been investigated and this has even led to the esoteric suggestion. Recent study displayed the degradation of an azo dye, such as Sudan orange, by bacterial laccases [4].
Laccases from different organisms have been purified and characterized. The sources of these laccases include plants, insects, fungi, and bacteria [5]. The genes of these laccases have also been obtained and heterologously expressed in different hosts [6,7]. Although laccases from fungi are considered the primary choice for industrial processes, the reported fungal laccases fail to meet industrial requirements, which are restricted by low pH stability, a long fermentation period, and a low tolerance to organic solvents [6,8]. Therefore, the screening and development of robust laccases have attracted significant attention.
Laccases from bacteria generally possess a wide and suitable pH range and a large yield. Laccase-producing strains have been found in Bacillus sp., Arthrospira sp., Achromobacter sp., Citrobacter sp., Sinorhizobium sp., and Klebsiella sp. [8]. In this study, a new laccase-producing strain was screened and named Vibrio sp. LA. The laccase coding gene L01 was cloned from genomic DNA and expressed in Yarrowia lipolytica. Recombinant laccase L01 is pH-stable and salt-tolerant; thus, it is an efficient tool for catalysis in industrial production.
2. Results and Discussion
2.1. Laccase-Producing Strain Vibrio sp. LA
Laccases are mainly found in terrestrial bacteria and fungi. To date, few marine bacteria have been shown to produce laccases [6,7,8]. Considering phenol pollution in marine environments, determining whether marine bacteria secrete laccases is of great significance [9]. The marine bacteria were isolated from 2216E plates, and then the strains were purified by streak cultivation. 24 laccase-producing strains sourced from marine bacteria were grown on 2216E medium plates containing 0.4 mM Cu2+ at 25 °C. With 2,2′-azino-bis-(3-ethylbenzothiazoline-6-sulfonic acid) (ABTS) as the substrate, the strain LA was detected with the highest laccase activity of 0.82 ± 0.07 U/mL at flask level (data not shown). For identification, the strain LA was subjected to 16S rDNA sequencing. BLAST alignment showed that the 16S rDNA sequence had the highest similarity (98.05%) to that of an identified strain of Vibrio algivorus, indicating that strain LA is a member of the genus Vibrio. Thus, the assigned designation was Vibrio sp. LA.
2.2. Analysis of L01 Gene Using Bioinformatics Methods
The laccase from strain LA was named L01. To obtain the L01 gene encoding laccase secreted by Vibrio sp. LA, the genomic DNA of this strain was sequenced and annotated. The putative laccase-encoding gene was confirmed by catalytic region analysis. The coding sequence (CDS) starting from ATG contained 1383 bp, with a translated protein consisting 460 amino acids. It was found that the protein has a theoretical isoelectric point (pI) of 6.14 and a calculated molecular weight (MW) of 51,600 Da. In L01, the first 25 amino acids were inferred to be the signal peptide, which agreed with the extracellular location of L01. NCBI BLAST analysis revealed that L01 has a conserved domain and is a multicopper oxidase.
To further analyze the sequence characteristics of L01, we constructed a phylogenetic tree based on the amino acid sequences of L01 and laccases from other species. Clearly, the L01 groups with bacterial laccases were far from fungal laccases (Figure 1a). The closest laccases to L01 were from Streptomyces ipomoeae (WP 141595389.1) and Ochrobactrum sp. 531 (ADK66266.1) [10,11]. The structure model was constructed using the crystal structure of multicopper oxidase from Campylobacter jejuni (PDB: 3ZX1) as the model (Figure 1b), with a sequence similarity of 35%. L01 has three cupredoxin domains of CumA such as multicopper oxidase, with domain I (residues 36–154), domain II (residues 164–289), and domain III (residues 325–458). To further study the structural features of L01, we performed multiple sequence alignments of L01 and seven well-characterized laccases. The seven laccases were isolated from Fusarium oxysporum (KAG7428168.1), Bacillus amyloliquefaciens (QHT73050.1), Aspergillus niger (GAQ36284.1), Bacillus subtilis (AEK80414.1), Ochrobactrum sp. (ADK66266.1), and Aureobasidium pullulans (AUW14931.1). The results reflect that L01 has typical conserved motifs appearing in laccases, such as “HPXHXHGHXF”, “WYHPH”, “HWHGI”, “HPXHXHGHXF”, and “HCH” (Figure 2), which are verified copper-binding domains [12,13,14,15,16]. Thus, according to bioinformatics analysis, L01 is a typical laccase.

Figure 1.
(a) Phylogenetic tree constructed by the neighbor-joining method with amino acid sequences of L01 and reported laccases. (b) Structure model of L01 by SWISS-MODEL based on the amino sequence.

Figure 2.
Multiple sequence alignments of L01 with other well-characterized bacterial and fungal laccases, with conserved motifs marked in blue boxes.
2.3. Recombinant Expression of L01 in Yarrowia lipolytica
In previous studies, laccases were mostly expressed in E. coli, which is infeasible for their industrial production because of the weak secretory ability of E. coli and the complex post-processing procedures of laccases [6,7,17]. In this study, we expressed L01 gene in Yarrowia lipolytica, a commonly used heterologous host that can produce extraordinary extracellular secretion. Many industrial enzymes have been expressed in this host with high extracellular activities [18,19,20,21]. After 60 h of incubation in the GPPB medium, the laccase secreted by the recombinant strain LA12 had an activity of up to 18.6 ± 0.6 U/mL (Figure 3), over 20-fold higher than that secreted by the wild-type strain. L01 purification was performed on the supernatant, followed by SDS-PAGE analysis. As shown in Figure 3, a clear single band was observed in lanes 3–4. The Mw of L01 was approximately 56,000, which is slightly higher than the theoretical value (52,500) of the cloned amino acids plus the His-tag. The specific activity of purified L01 toward ABTS is 98.6 ± 2.3 U/mg.
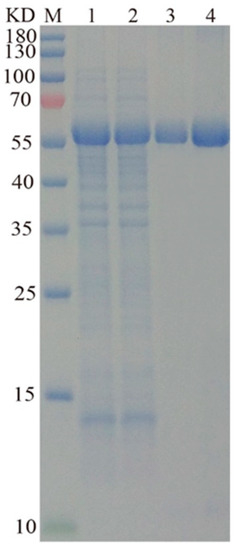
Figure 3.
SDS-PAGE analysis of L01. Samples were loaded in different lanes. Lane M, prestained protein ladder; Lanes 1–2, the concentrated fermentation broth; Lanes 3–4, purified L01.
2.4. Effects of Temperature and pH on Activity of L01
We investigated the enzymatic properties of purified L01 and found that its catalytic activity peaked at 35 °C toward ABTS, decreased dramatically above 50 °C, and was 51.2 ± 2.1% and 74.5 ± 5.2% of the maximum activity at 10 °C and 20 °C, respectively (Figure 4a). L01 was relatively stable at temperatures below 35 °C, and 63.1 ± 4.2% of its highest activity remained after processing at 35 °C for 2 h. However, at temperatures above 45 °C, the activity was quickly lost (Figure 4b).
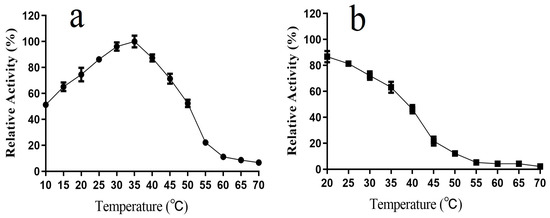
Figure 4.
(a) Effect of temperature on L01 catalytic activity; (b) effect of temperature on L01 stability after 2 h of incubation (n = 3).
Most bacterial laccases exhibit their highest activity below 40 °C. Most laccases have an activity of less than half of the highest level below 30 °C [12,22,23,24,25,26,27]. In contrast, L01 exhibits higher activity at 10 °C and 20 °C and relatively better thermostability compared to that of other laccases, which indicates that L01 is a typical cold-adapted laccase. Reactions catalyzed by cold-adapted laccases are characterized by reduced energy consumption and a low contamination risk. Thus, L01 has promising prospects as a biocatalyst candidate for industrial applications [2,6,21].
The optimal pH for the activity of L01 toward ABTS was 8.0, and its activity in the range of pH 4.5–10.0 was higher than 60% of the maximum (Figure 5a). After 6 h of incubation, over 40% of the highest activity was retained at pH 4.0–10.0. Notably, in the test pH range (5.5–9.5) in this study, the activity of L01 exceeded 65% of the highest activity after incubation.
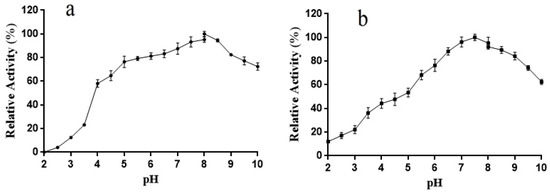
Figure 5.
(a) L01 catalytic activity at different pH; (b) L01 stability at different pH after 4 h of incubation (n = 3).
Generally, fungal laccases play a catalytic role in oxidation reactions within a narrow acidic pH range, whereas bacterial laccases prefer a relatively neutral pH [2,12,13,14,15]. Compared with the laccases reported in Table 1, L01 is stable over a much broader pH range, and pH can play a role in its catalytic activity. For example, laccases from Aureobasidium melanogenum and Pseudomonas extremorientalis are stable in narrow pH ranges of 2.8–3.6 and 7.0–10.0, respectively [12,24]. It is well recognized that pH-stable and mesophilic laccases can be generated by Aquisalibacillus elongatus [23]. L01 outperformed A. elongatus in terms of pH stability, which can contribute to the biotransformation of various phenolic substrates, especially in cases with a significant pH change.

Table 1.
Comparison of properties between L01 and reported laccases.
2.5. L01 Activity with the Existence of Various Metal Ions and Some Compounds
Figure 6 shows L01 activities affected by metal ions with different concentrations.
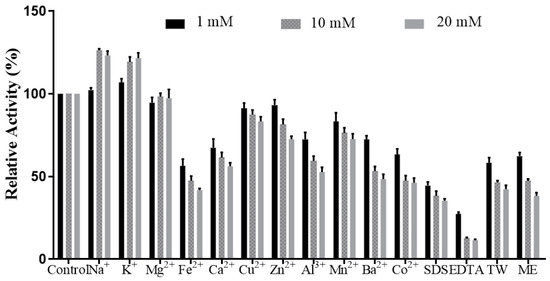
Figure 6.
Effects of metal ions, SDS, TW, ME, and EDTA on L01 activity (n = 3). TW: Tween 80, ME: mercaptoethanol.
When the concentrations of K+ were 1 mM, 10 mM, and 20 mM, the relative activities were 106.8 ± 2.3%, 119.2 ± 3.1%, and 121.5 ± 3.2%, respectively, compared to that of the control. With the presence of Na+ at 10 mM or 20 mM, the relative activities were also elevated to 126.3 ± 0.9% and 123.2 ± 2.6%, respectively. Among the tested metal ions, Fe2+ and Ca2+ were detrimental to L01 activity, reducing L01 activity to 41.7 ± 1.0% and 56.1 ± 2.1% at 20 mM, respectively (Figure 6). The activity of L01 declined to 44.5 ± 2.1% and 27.2 ± 1.2% in the presence of 1 mM SDS and EDTA, respectively. With the concentrations of SDS and EDTA increased, the activities continued to decrease. 10 mM and 20 mM EDAT can reduce the activities to 12.3 ± 0.8% and 11.4 ± 0.5%. When in the presence of all metal ions at three concentrations, the activities were all above 40%. The activities were higher than those reduced by Fe2+ and Ca2+ at 10 mM and 20 mM. The above results demonstrate that L01 has a good tolerance to metal ions, and K+ and Na+ at 10 mM and 20 mM are good choices for activating agents [2,6,21]. Because it is salt-tolerant, L01 is suitable for various specific processing requirements.
2.6. Decolorization of Synthetic Dyes by L01
In this study, we used L01 to decolorize various synthetic dyes and found that it was effective for decolorizing several dyes. Compared with the commercial laccase from Aspergillus oryzae, L01 was a preferred choice for decolorizing eriochrome black T, methyl orange, and bromophenol blue. The decolorization rates for eriochrome black T, methyl orange, and bromophenol blue were 62.3 ± 3.1%, 32.1 ± 1.3%, and 46.3 ± 2.2%, respectively (Table 2). However, L01 was less satisfactory in decolorizing malachite green, crystal violet, and safranine 0, and the decolorization rates were only in the range of 10.2–15.2% within 24 h. The recombinant laccase from Trametes hirsute exhibited a more efficient decolorization ability for remazol brilliant blue R, whereas laccase from the basidiomycete Coprinopsis was efficient for indigo dye decolorization [28,29].
Table 2.
Decolorization rates (%) of synthetic dyes by L01 and commercial laccase (n = 3).
3. Materials and Methods
3.1. Materials, Strains, and Media
2216E plates were used to isolate bacteria from seawater samples [30]. The auxotrophic mutant yeast strain of Y. lipolytica and the vector pINA1312 were used for gene expression analysis. Marine sample-derived strains were cultivated in 2216E medium. A YNB plate was used to screen for the URA transformants of Y. lipolytica. GPPB medium (pH 6.8) was used to produce the recombinant enzyme [31].
3.2. Screening Laccase-Producing Strains at a Low Temperature
The isolated strains were cultivated on 2216E plates in the presence of 0.4 mM Cu2+ at 25 °C for 48 h to screen for potential laccase-producing strains [30]. A total of 24 laccase-producing strains sourced from marine bacteria were grown on 2216E medium plates containing 0.4 mM Cu2+ at 25 °C. PCR amplification was performed on the 16S rDNA of strain LA using universal primers 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and 1492R (5′-TACCTTGTTACGACTT-3′). 16S rDNA was then subjected to sequencing and BLAST alignment. After centrifugation of the culture at 5000× g, laccase activity of the supernatant was determined using 190 μL of 1 mM ABTS solution (in 10 mM sodium acetate buffer, pH 4.5) [32]. The reaction was conducted at 25 °C for 10 min and terminated by adding 300 μL of 10% trichloroacetic acid. One enzyme unit (U) in this study was defined as the enzyme amount required to produce an absorbance increase of one unit at 405 nm per milliliter of the mixture per minute under assay conditions. All treatments were performed in triplicate.
3.3. Bioinformatics Analysis of L01
To obtain the L01 gene, the genomic DNA of LA was sent to Novogene (China) for sequencing and annotation. A putative laccase-encoding gene was identified. We analyzed the signal peptide using the online server (http://www.cbs.dtu.dk/services/SignalP-4.1/ (accessed on 11 December 2022)) and performed conserved domain analysis (https://www.ncbi.nlm.nih.gov/cdd (accessed on 11 December 2022)). The theoretical pI and Mw values were also calculated (http://web.expasy.org/compute_pi/ (accessed on 11 December 2022)). We also constructed a phylogenetic tree based on the amino acid sequences of L01 and other laccases, using MEGA version 7.0. Multiple sequence alignments were performed on L01, and laccases were characterized using DNAMAN 6.0.
3.4. Recombinant Expression of L01 in Yeast Host and Purification of the Recombinant Enzyme
We entrusted Synbio Technologies (Suzhou, China) to synthesize the gene L01 with the XPR2 signal peptide after codons were optimized, and then transformed the obtained DNA fragment into a URA-strain [30]. The transformants were cultivated at 30 °C for 60 h in GPPB liquid medium and laccase activity was determined. The recombinant strain LA12 presented the highest laccase activity. After the pH of the supernatant was adjusted to 7.5, purification was conducted using a Ni–NTA column. L01 was adsorbed onto the gel, which was then washed off after loading with the imidazole solution. The purity and the Mw of L01 were estimated by SDS-PAGE analysis.
3.5. Impacts of Temperatures and Different pH on Activity and Stability of L01
To clarify the influence of temperature on L01 activity, we investigated a hydrolysis reaction catalyzed by L01 in glycine-NaOH buffer (10 mM, pH 8.0) at different temperatures (10–60 °C). The activity of L01 was used to determine the optimal reaction temperature. To clarify its thermal stability, we incubated purified L01 for 2 h at 10–60 °C and then detected the remaining activity at 35 °C. ABTS solutions with substrates were prepared with different pH buffers (glycine-NaOH (10 mM, pH 8.5–11.0) and Na2HPO4-citric acid (10 mM, pH 3.0–8.0)), which were used in enzymatic activity assays to determine the optimal pH for the reaction. The pH stability was estimated by detecting enzymatic activity after 4 h of incubation at 4 °C in pH buffers. The reactions were performed in triplicate.
3.6. L01 Activity with the Existence of Various Metal Ions and Some Compounds
High-concentration metal ions and compounds were diluted with ABTS solution to a concentration of 1 mM. L01 activity was detected in the presence of various metal ions and some compounds through reactions catalyzed by L01 at 35 °C. The reactions were performed in triplicate.
3.7. Decolorization of Synthetic Dyes by L01
Safranine 0 (60 mg/L), methylene blue (10 mg/L), bromophenol blue (300 mg/L), eriochrome black T (300 mg/L), crystal violet (10 mg/L), malachite green (10 mg/L), and methyl orange (10 mg/L) were prepared in a pH 5.0 solution. L01 solution and commercial laccase from Aspergillus oryzae (Sunson Biotech, China), with an activity of 5 U/mL was added to the dye solution. The resulting mixtures were incubated for 6 h and 24 h at 28 °C. The ODs at the maximum absorbance wavelengths were recorded using a spectrophotometer. Dyes mixed with inactivated L01 were used as controls, and their ODs at maximal absorbance wavelengths were determined using the same method [12,32]. The remaining dye was used to calculate the decolorization rate. All reactions were performed in triplicates.
4. Conclusions
In this study, a new cold-adapted laccase was cloned, extracellularly expressed, and characterized. Recombinant laccase L01 presented its highest activity at 35 °C and retained over 50% of its maximum activity at 10 °C and 20 °C. In addition, over 60% of the activity was maintained at pH 4.5–10.0. In addition to being pH-stable, L01 was salt-tolerant and NaCl-independent. Several synthetic dyes were efficiently degraded. L01 is an attractive biocatalyst for industrial applications owing to its robustness and unique pH stability.
Author Contributions
J.J.: Conceptualization, writing—original draft, writing—review and editing, supervision, funding acquisition. J.-L.D.: Methodology, investigation, and writing—original draft. Z.-G.W.: Investigation, writing—review and editing. X.-Y.C.: Data curation. S.-J.W.: Methodology. Y.-C.W.: Methodology. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Natural Science Foundation of China (42007131), Natural Science Research Project of Higher Education in Jiangsu Province (20KJB610004), and National and Local Joint Engineering Laboratory for Municipal Sewage Resource Utilization Technology, Suzhou University of Science and Technology (2021KF02).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Morsi, R.; Bilal, M.; Iqbal, H.M.N.; Ashraf, S.S. Laccases and peroxidases: The smart, greener and futuristic biocatalytic tools to mitigate recalcitrant emerging pollutants. Sci. Total Environ. 2020, 714, 136572. [Google Scholar] [CrossRef] [PubMed]
- Senthivelan, T.; Kanagaraj, J.; Panda, R.C. Recent trends in fungal laccase for various industrial applications: A eco-friendly approach: A review. Biotechnol. Bioprocess Eng. 2016, 21, 19–38. [Google Scholar] [CrossRef]
- Yadav, A.; Yadav, P.; Singh, A.K.; Kumar, V.; Sonawane, V.C.; Markandeya; Bharagava, R.N.; Raj, A. Decolourisation of textile dye by laccase: Process evaluation and assessment of its degradation bioproducts. Bioresour. Technol. 2021, 340, 125591. [Google Scholar] [CrossRef]
- Sousa, A.C.; Martins, L.O.; Robalo, M.P. Laccases: Versatile Biocatalysts for the Synthesis of Heterocyclic Cores. Molecules 2021, 26, 3719. [Google Scholar] [CrossRef] [PubMed]
- Su, J.; Fu, J.; Wang, Q.; Silva, C.; Cavaco-Paulo, A. Laccase: A green catalyst for the biosynthesis of poly-phenols. Crit. Rev. Biotechnol. 2018, 38, 294–307. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.D.; Tiwari, A.; Anisha, G.S.; Chen, C.W.; Singh, A.; Haldar, D.; Patel, A.K.; Singhania, R.R. Laccase: A potential biocatalyst for pollutant degradation. Environ. Pollut. 2023, 319, 120999. [Google Scholar] [CrossRef]
- Moiseenko, K.V.; Savinova, O.S.; Vasina, D.V.; Kononikhin, A.S.; Tyazhelova, T.V.; Fedorova, T.V. Laccase isoenzymes of Trametes hirsute LE-BIN072: Degradation of industrial dyes and secretion under the different induction conditions. Appl. Biochem. Microbiol. 2018, 54, 834–841. [Google Scholar] [CrossRef]
- Chauhan, P.S.; Goradia, B.; Saxena, A. Bacterial laccase: Recent update on production, properties and industrial applications. 3 Biotech 2017, 7, 323. [Google Scholar] [CrossRef]
- Li, H.; Meng, F.; Duan, W.; Lin, Y.; Zheng, Y. Biodegradation of phenol in saline or hypersaline environments by bacteria: A review. Ecotoxicol. Environ. Saf. 2019, 184, 109658. [Google Scholar] [CrossRef]
- Molina-Guijarro, J.M.; Pérez, J.; Muñoz-Dorado, J.; Guillén, F.; Moya, R.; Hernández, M.; Arias, M.E. Detoxification of azo dyes by a novel pH-versatile, salt-resistant laccase from Streptomyces ipomoea. Int. Microbiol. 2009, 12, 13–21. [Google Scholar]
- Li, Y.; Zuo, W.F.; Li, Y.D.; Wang, X.G. Cloning of multicopper oxidase gene from Ochrobactrum sp. 531 and characterization of its alkaline laccase activity towards phenolic substrates. Adv. Biol. Chem. 2012, 2, 248–255. [Google Scholar] [CrossRef]
- Aung, T.; Jiang, H.; Chen, C.C.; Liu, G.L.; Hu, Z.; Chi, Z.M.; Chi, Z. Production, Gene Cloning, and Overexpression of a Laccase in the Marine-Derived Yeast Aureobasidium melanogenum Strain 11-1 and Characterization of the Recombinant Laccase. Mar. Biotechnol. 2019, 21, 76–87. [Google Scholar] [CrossRef] [PubMed]
- Silva, C.S.; Durão, P.; Fillat, A.; Lindley, P.F.; Martins, L.O.; Bento, I. Crystal structure of the multicopper oxidase from the pathogenic bacterium Campylobacter jejuni CGUG11284: Characterization of a metallo-oxidase. Metallomics 2012, 4, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.Y.; Lu, L.; Feng, F.J. Combined strategies for improving production of a thermo-alkali stable laccase in Pichia pastoris. Electron. J. Biotechnol. 2017, 28, 7–13. [Google Scholar] [CrossRef]
- Wang, S.S.; Ning, Y.J.; Wang, S.N.; Zhang, J.; Zhang, G.Q.; Chen, Q.J. Purification, characterization, and cloning of an extracellular laccase with potent dye decolorizing ability from white rot fungus Cerrena unicolor GSM-01. Int. J. Biol. Macromol. 2017, 95, 920–927. [Google Scholar] [CrossRef]
- Campos, P.A.; Levin, L.N.; Wirth, S.A. Heterologous production, characterization and dye decolorization ability of a novel thermostable laccase isoenzyme from Trametes trogii bafc 463. Process Biochem. 2016, 51, 895–903. [Google Scholar] [CrossRef]
- Miyamoto, T.; Okano, S.; Kasai, N. Inactivation of Escherichia coli endotoxin by soft hydrothermal processing. Appl. Environ. Microbiol. 2009, 75, 5058–5063. [Google Scholar] [CrossRef]
- Madzak, C. Yarrowia lipolytica: Recent achievements in heterologous protein expression and pathway engineering. Appl. Microbiol. Biotechnol. 2015, 99, 4559–4577. [Google Scholar] [CrossRef]
- Li, S.Y.; Wang, Z.P.; Wang, L.N.; Peng, J.X.; Wang, Y.N.; Han, Y.T.; Zhao, S.F. Combined enzymatic hydrolysis and selective fermentation for green production of alginate oligosaccharides from Laminaria japonica. Bioresour. Technol. 2019, 281, 84–89. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Li, B.; Huang, B.C.; Wang, F.B.; Zhang, Y.Q.; Zhao, S.G.; Li, M.; Wang, H.Y.; Yu, X.J.; Liu, X.Y.; et al. Production, Biosynthesis, and Commercial Applications of Fatty Acids from Oleaginous Fungi. Front. Nutr. 2022, 9, 873657. [Google Scholar] [CrossRef]
- Antosova, Z.; Sychrova, H. Yeast hosts for the production of recombinant laccases: A review. Mol. Biotechnol. 2016, 58, 93–116. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Deng, W.; Yang, Y. Characterization of a Novel Laccase LAC-Yang1 from White-Rot Fungus Pleurotus ostreatus Strain Yang1 with a Strong Ability to Degrade and Detoxify Chlorophenols. Molecules 2021, 26, 473. [Google Scholar] [CrossRef] [PubMed]
- Rezaei, S.; Shahverdi, A.R.; Faramarzi, M.A. Isolation, one-step affinity purification, and characterization of a polyextremotolerant laccase from the halophilic bacterium Aquisalibacillus elongatus and its application in the delignification of sugar beet pulp. Bioresour. Technol. 2017, 230, 67–75. [Google Scholar] [CrossRef]
- Neifar, M.; Chouchane, H.; Mahjoubi, M.; Jaouani, A.; Cherif, A. Pseudomonas extremorientalis BU118: A new salt-tolerant laccase-secreting bacterium with biotechnological potential in textile azo dye decolourization. 3 Biotech 2016, 6, 107. [Google Scholar] [CrossRef]
- Wang, C.L.; Zhao, M.; Lu, L.; Wei, X.D.; Li, T.L. Characterization of spore laccase from Bacillus subtilis wd23 and its use in dye decolorization. Afr. J. Biotehnol. 2011, 10, 2186–2192. [Google Scholar]
- Singh, G.; Bhalla, A.; Capalash, N.; Sharma, P. Characterization of immobilized Laccase from γ-proteobacterium JB: Approach towards the development of biosensor for the detection of phenolic compounds. Indian J. Sci. Technol. 2010, 3, 48–53. [Google Scholar] [CrossRef]
- Mcmahon, A.M.; Doyle, E.M.; Brooks, S.; O’ Connor, K.E. Biochemical characterisation of the coexisting tyrosinase and laccase in the soil bacterium pseudomonas putida f6. Enzym. Microb. Technol. 2007, 40, 1435–1441. [Google Scholar] [CrossRef]
- Jia, Y.; Huang, Q.; Zhu, L.; Pan, C. Characterization of a Recombinant Laccase B from Trametes hirsuta MX2 and Its Application for Decolorization of Dyes. Molecules 2022, 27, 1581. [Google Scholar] [CrossRef]
- Yin, Q.; Zhou, G.; Peng, C.; Zhang, Y.; Kues, U.; Liu, J.; Xiao, Y.; Fang, Z. The first fungal laccases with an alkaline pH optimum obtained by directed evolution and its application in indigo dye decolorization. AMB Express 2019, 9, 151. [Google Scholar] [CrossRef]
- Wang, Z.-P.; Cao, M.; Li, B.; Ji, X.-F.; Zhang, X.-Y.; Zhang, Y.-Q.; Wang, H.-Y. Cloning, Secretory Expression and Characterization of a Unique pH-Stable and Cold-Adapted Alginate Lyase. Mar. Drugs 2020, 18, 189. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, Z.P.; Sheng, J.; Zheng, Y.; Ji, X.F.; Zhou, H.X.; Liu, X.Y.; Chi, Z.M. High and efficient isomaltulose production using an engineered Yarrowia lipolytica strain. Bioresour. Technol. 2018, 265, 577–580. [Google Scholar] [CrossRef] [PubMed]
- Ning, Y.J.; Wang, S.S.; Chen, Q.J.; Ling, Z.R.; Wang, S.N.; Wang, W.P.; Zhang, G.Q.; Zhu, M.J. An extracellular yellow laccase with potent dye decolorizing ability from the fungus Leucoagaricus naucinus LAC-04. Int. J. Biol. Macromol. 2016, 93 Pt A, 837–842. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).