
Ginsenoside Rb1 Interfered with Macrophage Activation by Activating PPARγ to Inhibit Insulin Resistance in Obesity


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
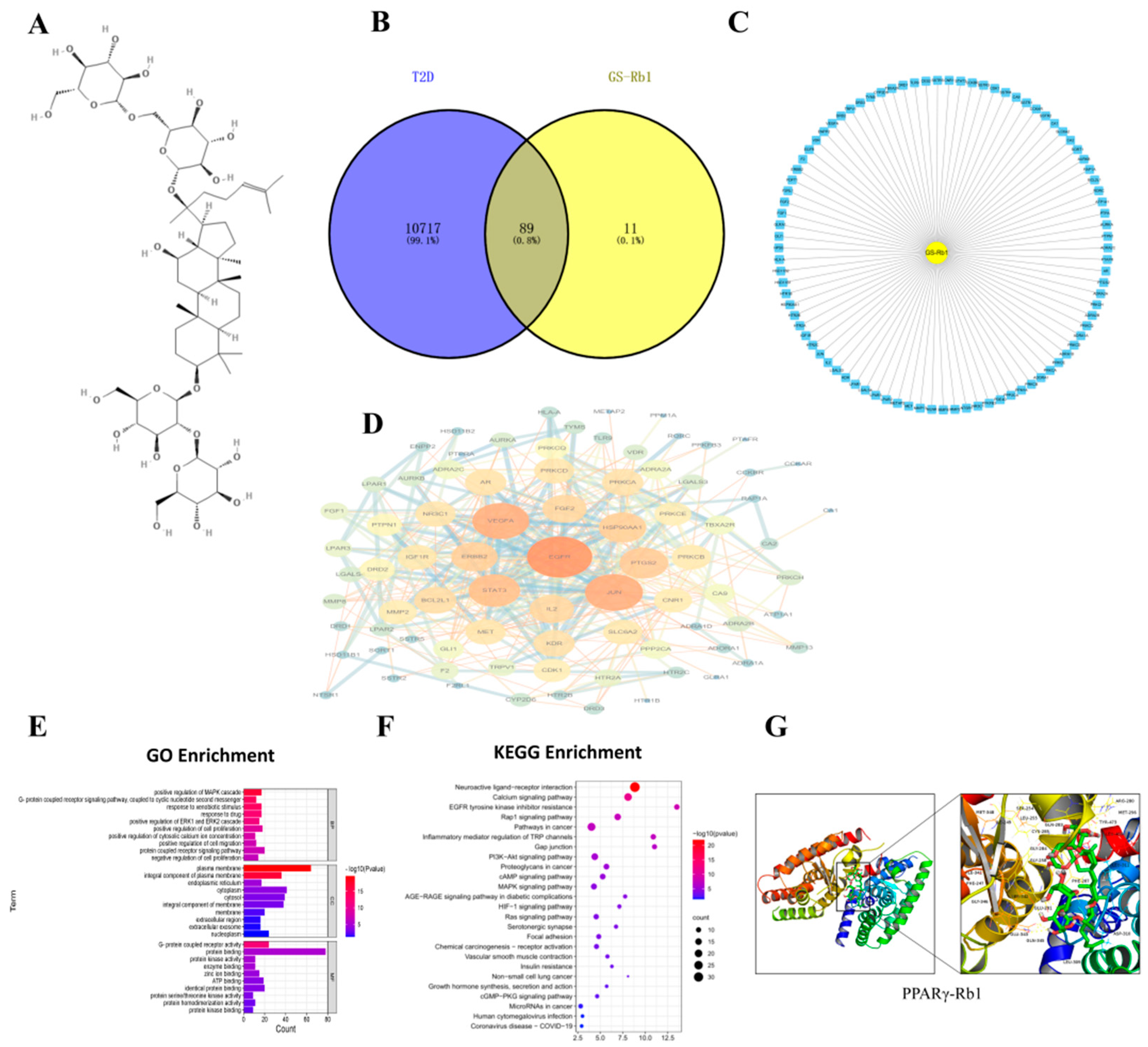
2.1. The Obtainment of Potential Targets
2.2. Acquisition of Common Targets for T2D-Rb1
2.3. PPI Network Analysis and Key Target Acquisition
2.4. GO and KEGG Pathway Enrichment Analysis
2.5. Molecular Modeling of PPARγ-Rb1
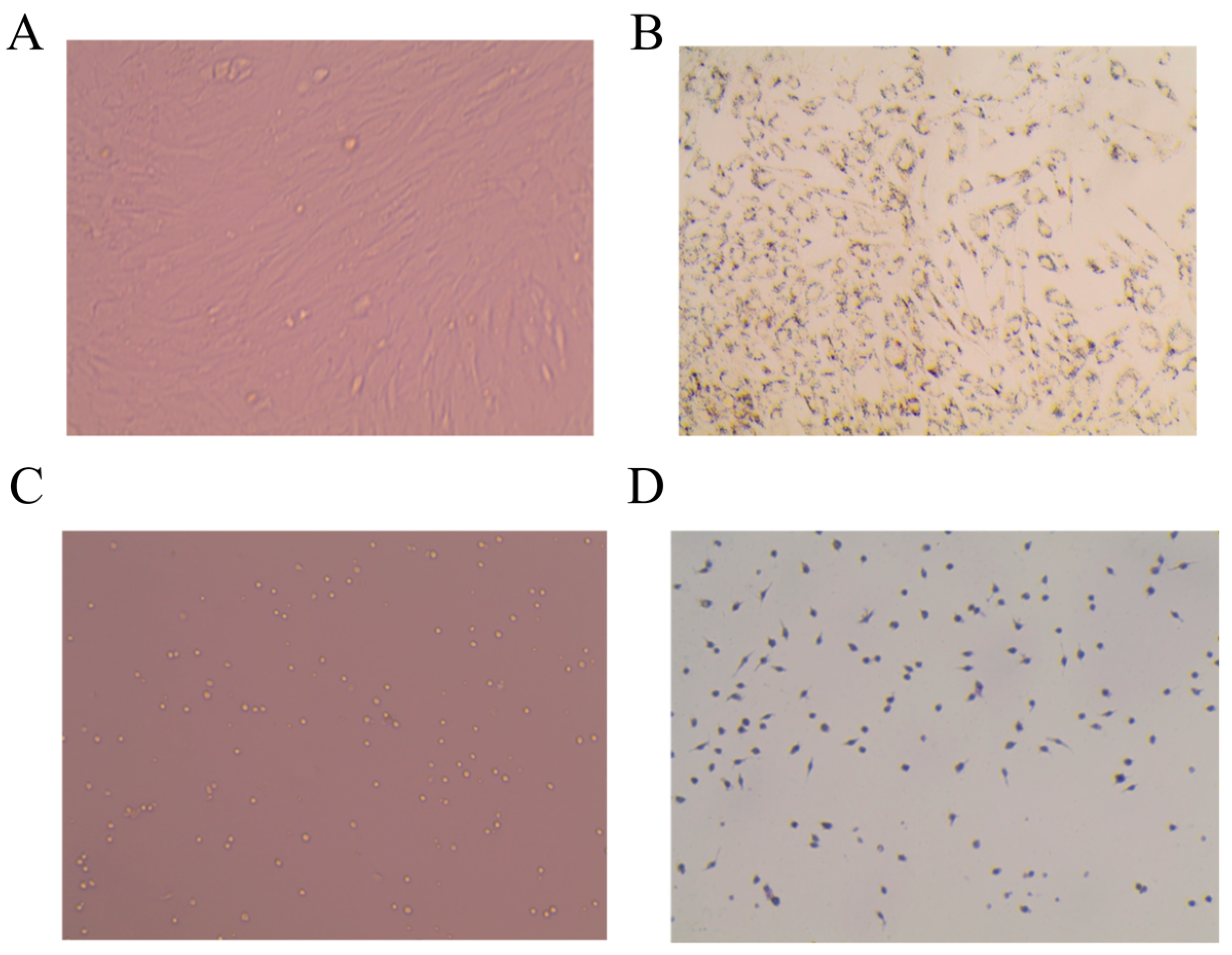
2.6. Identification of ADSC and PM
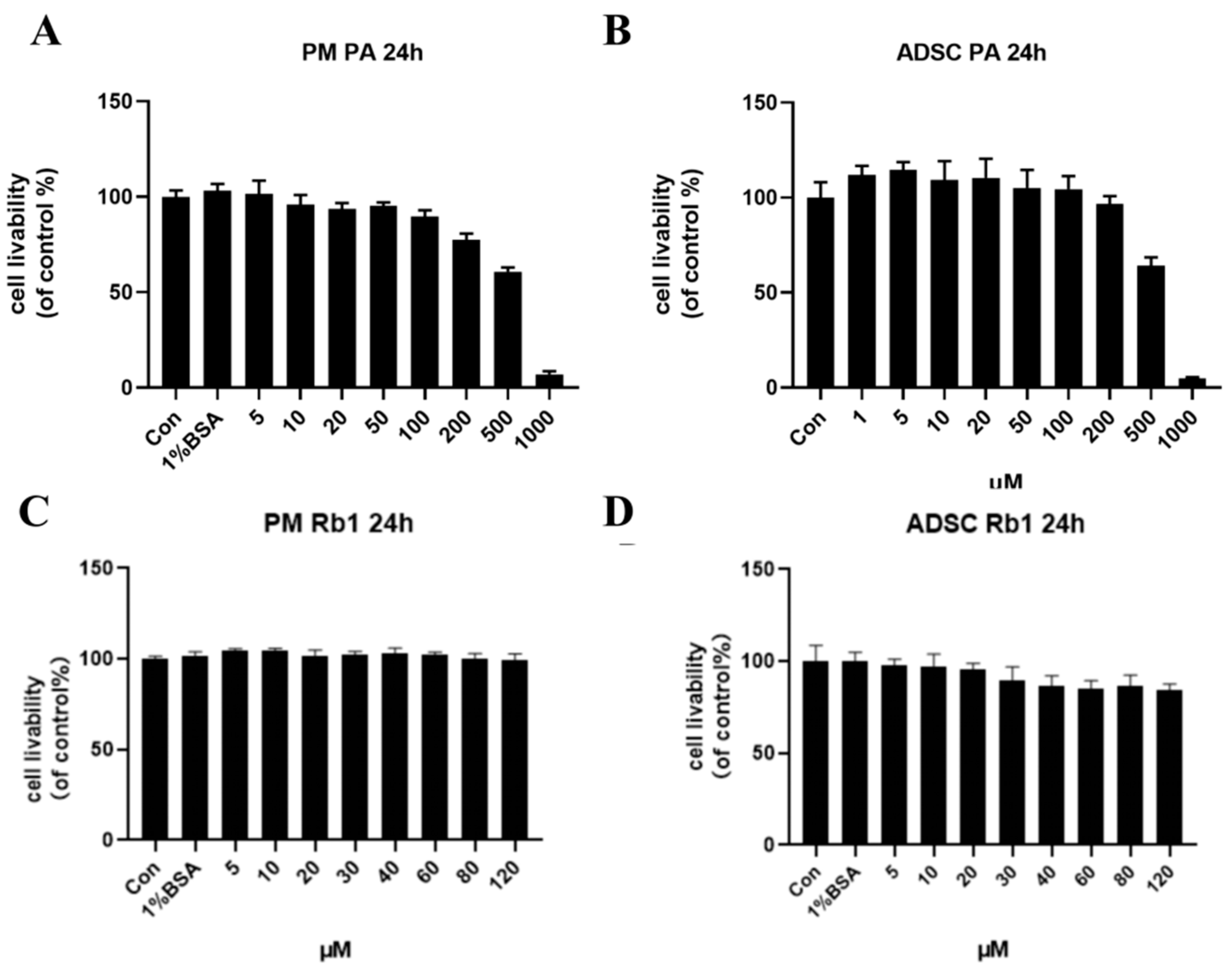
2.7. Cell Viability Assay
2.8. Establishment of Hypertrophy Model of Mature ADSC Cells Stimulated by PA
2.9. Obesity-Induced Insulin Resistance Promotes Macrophage Polarization
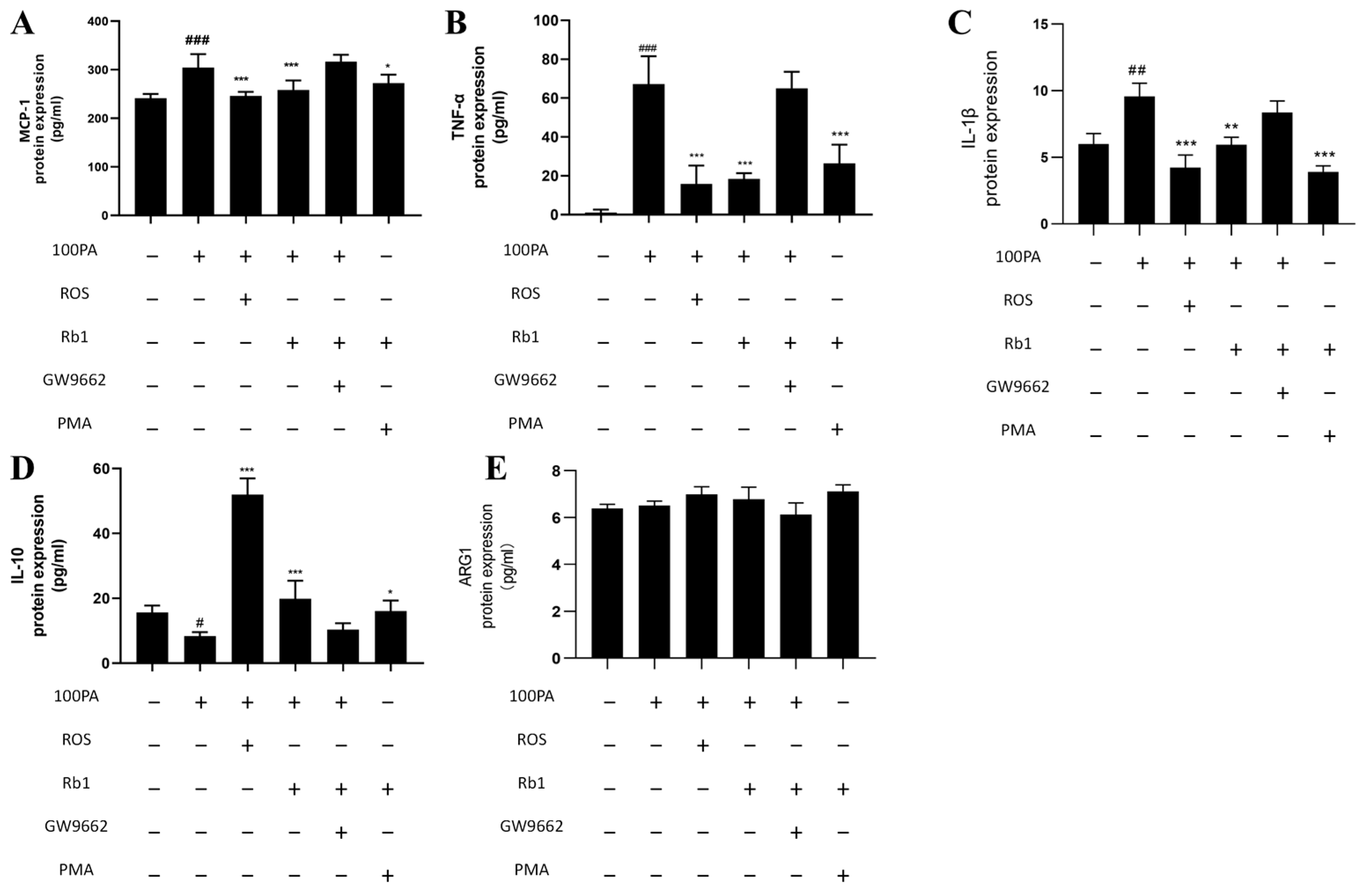
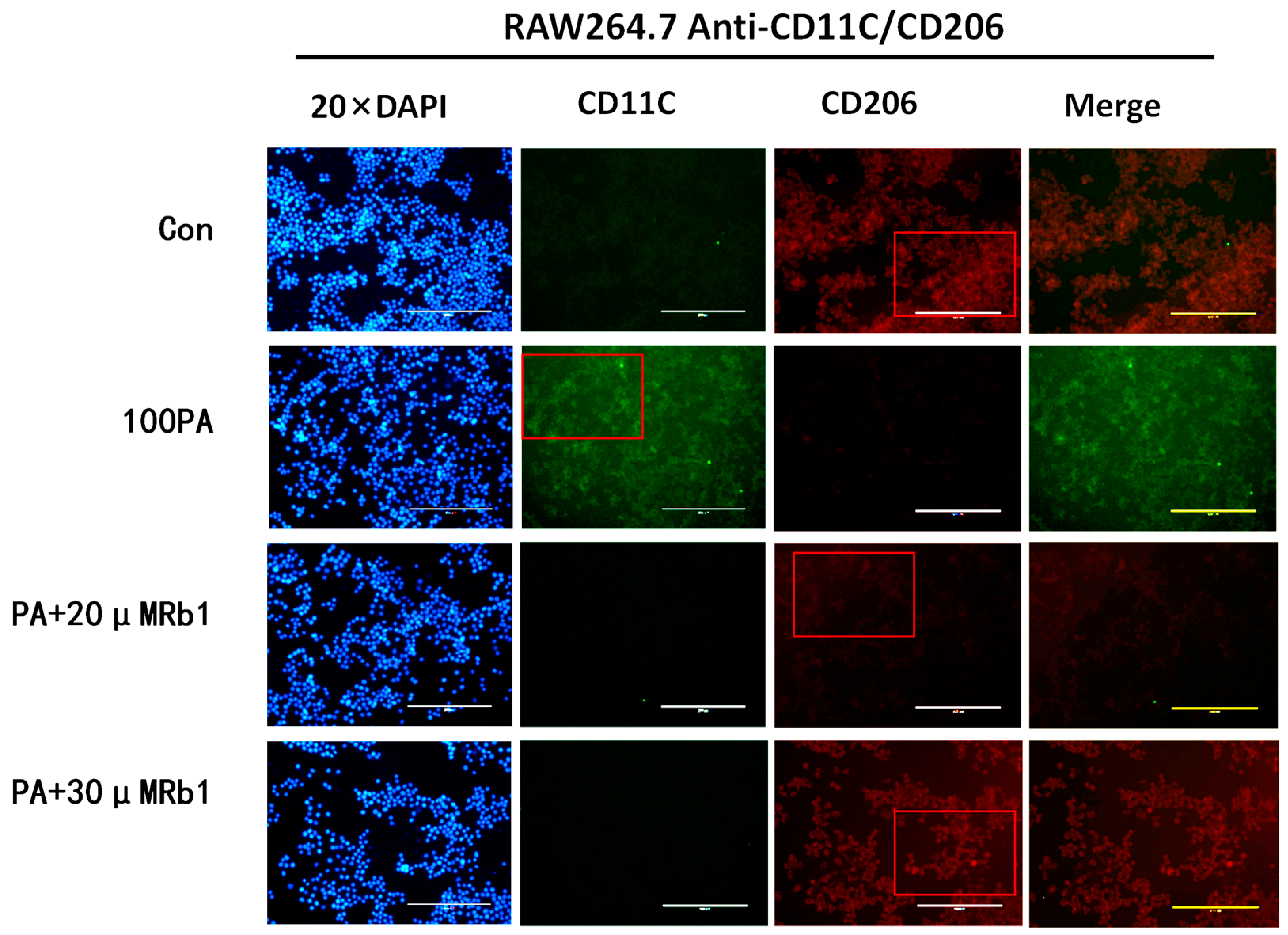
2.10. Effect of Obesity-Induced Insulin Resistance on Macrophage Activation
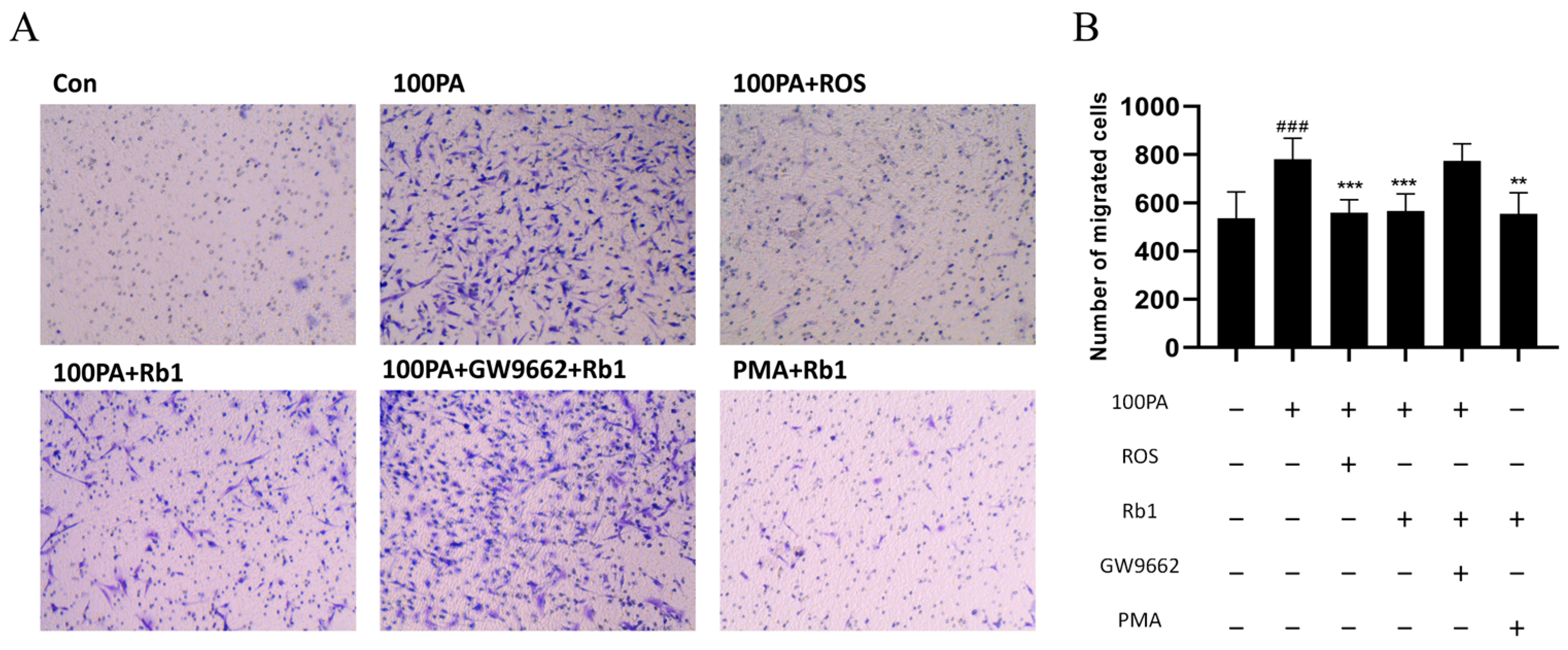
2.11. Obesity-Induced Insulin Resistance Promotes Macrophage Migration
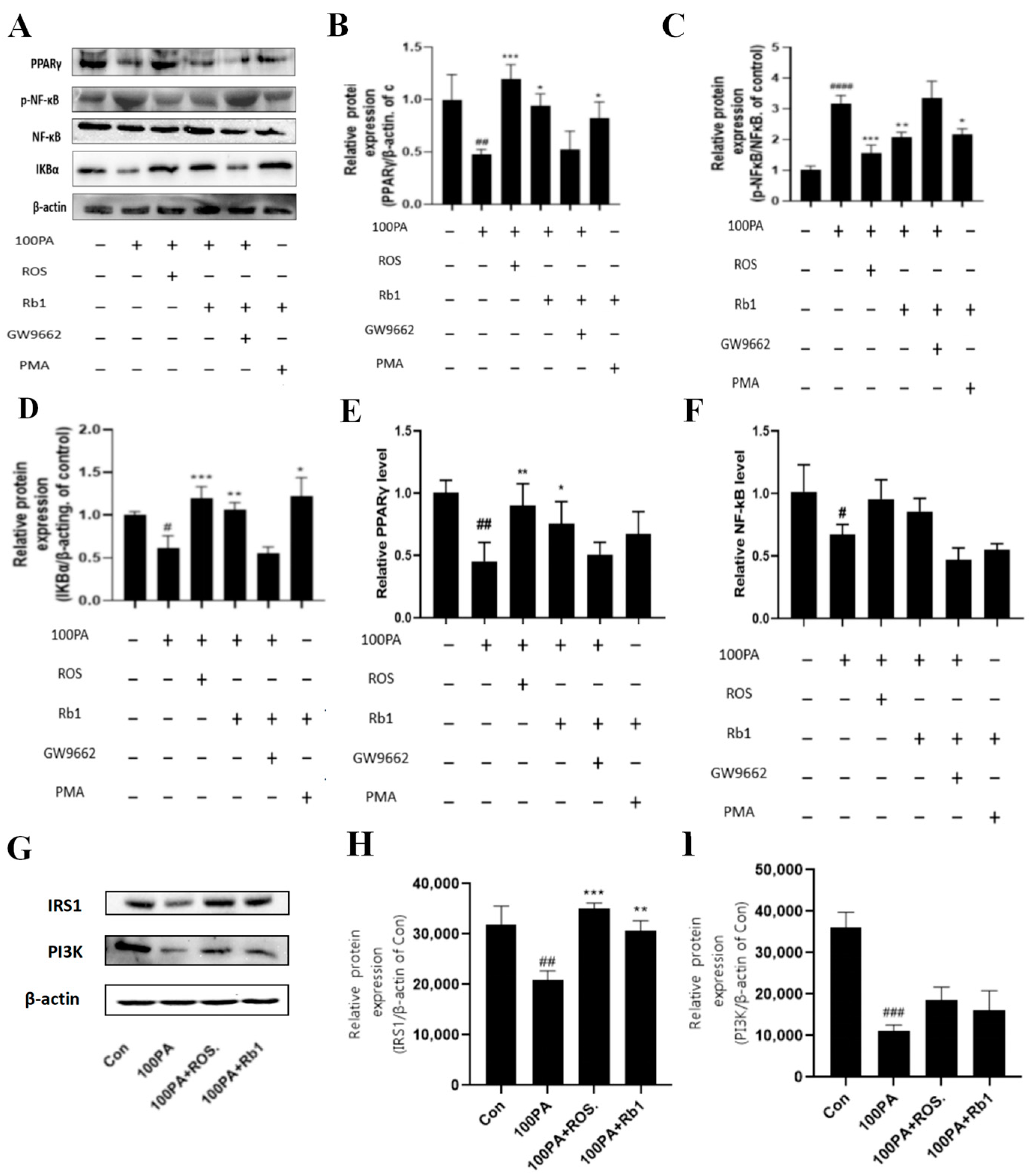
2.12. Signaling Pathway of Rb1 Alleviating Insulin Resistance
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. The Putative Targets of Rb1
4.3. Acquisition of T2D Disease-Related Targets and Drug-Disease Common Targets
4.4. Construction and Visualization of Protein-Protein Interaction (PPI) Network
4.5. GO Biological Function and KEGG Pathway Enrichment Analysis
4.6. Molecular Modeling
4.7. Extraction and Isolation of ADSCs
4.8. Differentiation Is Induced by ADSC
4.9. PM Extraction and Separation
4.10. Identification of PM
4.11. CCK 8 Experiments
4.12. Establishment of Cell Hypertrophy Model
4.13. Oil Red O Staining
4.14. Determination of TG Content
4.15. Cell Co-Culture and Migration Efficiency Detection
4.16. Detection of Cytokines in Cell Supernatant
4.17. Immunofluorescence
4.18. Western Blot Analysis
4.19. PCR
4.20. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Sood, S.; Feehan, J.; Itsiopoulos, C.; Wilson, K.; Plebanski, M.; Scott, D.; Hebert, J.R.; Shivappa, N.; Mousa, A.; George, E.S.; et al. Higher Adherence to a Mediterranean Diet Is Associated with Improved Insulin Sensitivity and Selected Markers of Inflammation in Individuals Who Are Overweight and Obese without Diabetes. Nutrients 2022, 14, 4437. [Google Scholar] [CrossRef]
- Tee, C.C.L.; Cooke, M.B.; Chong, M.C.; Yeo, W.K.; Camera, D.M. Mechanisms for Combined Hypoxic Conditioning and Divergent Exercise Modes to Regulate Inflammation, Body Composition, Appetite, and Blood Glucose Homeostasis in Overweight and Obese Adults: A Narrative Review. Sports Med. 2023, 53, 327–348. [Google Scholar] [CrossRef] [PubMed]
- Nie, L.J.; Yan, Q.H.; Zhang, S.; Cao, Y.T.; Zhou, X.Q. Duodenal Mucosa: A New Target for the Treatment of Type 2 Diabetes. Endocr. Pract. 2022, 29, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Cadenas-Sanchez, C.; Cabeza, R.; Idoate, F.; Osés, M.; Medrano, M.; Villanueva, A.; Arenaza, L.; Sanz, A.; Ortega, F.B.; Ruiz, J.R.; et al. Effects of a Family-Based Lifestyle Intervention Plus Supervised Exercise Training on Abdominal Fat Depots in Children with Overweight or Obesity: A Secondary Analysis of a Nonrandomized Clinical Trial. JAMA Netw. Open 2022, 5, e2243864. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Yan, X.; Zhao, Y.; Wang, J.; Liu, B.; Yu, S.; Fu, J.; Liu, Y.; Su, J. Macrophage Polarization Mediated by Mitochondrial Dysfunction Induces Adipose Tissue Inflammation in Obesity. Int. J. Mol. Sci. 2022, 23, 9252. [Google Scholar] [CrossRef]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Yan, S.; Wei, H.; Jia, R.; Zhen, M.; Bao, S.; Wang, W.; Liu, F.; Li, J. Wu-Mei-Wan Ameliorates Murine Ulcerative Colitis by Regulating Macrophage Polarization. Front. Pharmacol. 2022, 13, 917. [Google Scholar] [CrossRef]
- Li, Q.; Dai, Z.; Cao, Y.; Wang, L. Caspase-1 inhibition mediates neuroprotection in experimental stroke by polarizing M2 microglia/macrophage and suppressing NF-κB activation. Biochem. Biophys. Res. Commun. 2019, 513, 479–485. [Google Scholar] [CrossRef]
- Hu, Q.; Yao, J.; Wu, X.; Li, J.; Li, G.; Tang, W.; Liu, J.; Wan, M. Emodin attenuates severe acute pancreatitis-associated acute lung injury by suppressing pancreatic exosome-mediated alveolar macrophage activation. Acta Pharm. Sin. B 2021, 12, 3986–4003. [Google Scholar] [CrossRef]
- Zhao, D.; Xue, C.; Yang, Y.; Li, J.; Wang, X.; Chen, Y.; Zhang, S.; Chen, Y.; Duan, Y.; Yang, X.; et al. Lack of Nogo-B expression ameliorates PPARγ deficiency-aggravated liver fibrosis by regulating TLR4-NF-κB-TNF-α axis and macrophage polarization. Biomed. Pharmacother. 2022, 153, 113444. [Google Scholar] [CrossRef]
- Bauer, D.; Hennig, M.; Wasmuth, S.; Baehler, H.; Busch, M.; Steuhl, K.-P.; Thanos, S.; Heiligenhaus, A. Amniotic Membrane Induces Peroxisome Proliferator-Activated Receptor-γ Positive Alternatively Activated Macrophages. Investig. Opthalmol. Vis. Sci. 2012, 53, 799–810. [Google Scholar] [CrossRef] [PubMed]
- Guo, R.; Wang, L.; Zeng, X.; Liu, M.; Zhou, P.; Lu, H.; Lin, H.; Dong, M. Aquaporin 7 involved in GINSENOSIDE-RB1-mediated anti-obesity via peroxisome proliferator-activated receptor gamma pathway. Nutr. Metab. 2020, 17, 69. [Google Scholar] [CrossRef] [PubMed]
- You, L.; Cha, S.; Kim, M.-Y.; Cho, J.Y. Ginsenosides are active ingredients in Panax ginseng with immunomodulatory properties from cellular to organismal levels. J. Ginseng Res. 2021, 46, 711–721. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, S.; Zhu, Z.; Zhou, R.; Xu, P.; Zhou, L.; Kan, Y.; Li, J.; Zhao, J.; Fang, P.; et al. Upregulation of adiponectin by Ginsenoside Rb1 contributes to amelioration of hepatic steatosis induced by high fat diet. J. Ginseng Res. 2021, 46, 561–571. [Google Scholar] [CrossRef]
- Zou, H.; Zhang, M.; Zhu, X.; Zhu, L.; Chen, S.; Luo, M.; Xie, Q.; Chen, Y.; Zhang, K.; Bu, Q.; et al. Ginsenoside Rb1 Improves Metabolic Disorder in High-Fat Diet-Induced Obese Mice Associated with Modulation of Gut Microbiota. Front. Microbiol. 2022, 13, 826487. [Google Scholar] [CrossRef]
- Li, H.S.; Kuang, J.Y.; Liu, G.J.; Wu, W.J.; Yin, X.L.; Li, H.D.; Wang, L.; Qin, T.; Zhang, W.C.; Sun, Y.Y. Myostain is involved in ginsenoside Rb1-mediated anti-obesity. Pharm. Biol. 2022, 60, 1106–1115. [Google Scholar] [CrossRef]
- Croasdell, A.; Duffney, P.F.; Kim, N.; Lacy, S.H.; Sime, P.J.; Phipps, R.P. PPARγ and the Innate Immune System Mediate the Resolution of Inflammation. PPAR Res. 2015, 2015, 549691. [Google Scholar] [CrossRef] [Green Version]
- Juge-Aubry, C.E.; Gorla-Bajszczak, A.; Pernin, A.; Lemberger, T.; Wahli, W.; Burger, A.G.; Meier, C.A. Peroxisome proliferator-activated receptor mediates cross-talk with thyroid hormone receptor by competition for retinoid X receptor. Possible role of a leucine zipper-like heptad repeat. J. Biol. Chem. 1995, 270, 18117–18122. [Google Scholar] [CrossRef] [Green Version]
- Recinella, L.; Gorica, E.; Chiavaroli, A.; Fraschetti, C.; Filippi, A.; Cesa, S.; Cairone, F.; Martelli, A.; Calderone, V.; Veschi, S.; et al. Anti-Inflammatory and Antioxidant Effects Induced by Allium sativum L. Extracts on an Ex Vivo Experimental Model of Ulcerative Colitis. Foods 2022, 11, 3559. [Google Scholar] [CrossRef]
- Baskaran, R.; Poornima, P.; Huang, C.Y.; Padma, V.V. Neferine prevents NF-κB translocation and protects muscle cells from oxidative stress and apoptosis induced by hypoxia. Biofactors 2016, 42, 407–417. [Google Scholar] [CrossRef]
- Le, Y.Z.; Xu, B.; Chucair-Elliott, A.J.; Zhang, H.; Zhu, M. VEGF Mediates Retinal Müller Cell Viability and Neuroprotection through BDNF in Diabetes. Biomolecules 2021, 11, 712. [Google Scholar] [CrossRef] [PubMed]
- Gharbia, S.; Nazarie, S.-R.; Dinescu, S.; Balta, C.; Herman, H.; Peteu, V.E.; Gherghiceanu, M.; Hermenean, A.; Costache, M. Adipose-Derived Stem Cells (ADSCs) Supplemented with Hepatocyte Growth Factor (HGF) Attenuate Hepatic Stellate Cell Activation and Liver Fibrosis by Inhibiting the TGF-β/Smad Signaling Pathway in Chemical-Induced Liver Fibrosis Associated with Diabetes. Cells 2022, 11, 3338. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Dong, J.; Ding, H.; Wang, B.; Wang, Y.; Qiu, Z.; Yao, F. Ginsenoside compound K inhibits obesity-induced insulin resistance by regulation of macrophage recruitment and polarization via activating PPARγ. Food Funct. 2022, 13, 3561–3571. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Dong, J.; Xu, J.; Qiu, Z.; Yao, F. Ginsenoside CK inhibits obese insulin resistance by activating PPARγ to interfere with macrophage activation. Microb. Pathog. 2021, 157, 105002. [Google Scholar] [CrossRef] [PubMed]
- Yao, F.; Yu, Y.; Feng, L.; Li, J.; Zhang, M.; Lan, X.; Yan, X.; Liu, Y.; Guan, F.; Zhang, M.; et al. Adipogenic miR-27a in adipose tissue upregulates macrophage activation via inhibiting PPARγ of insulin resistance induced by high-fat diet-associated obesity. Exp. Cell Res. 2017, 355, 105–112. [Google Scholar] [CrossRef]
- Yang, X.; Dong, B.; An, L.; Zhang, Q.; Chen, Y.; Wang, H.; Song, Z. Ginsenoside Rb1 ameliorates Glycemic Disorder in Mice with High Fat Diet-Induced Obesity via Regulating Gut Microbiota and Amino Acid Metabolism. Front. Pharmacol. 2021, 12, 756491. [Google Scholar] [CrossRef]
- Samuel, V.T.; Shulman, G.I. Mechanisms for insulin resistance: Common threads and missing links. Cell 2012, 148, 852–871. [Google Scholar] [CrossRef] [Green Version]
- Bosi, E.; Marselli, L.; Suleiman, M.; Tesi, M.; De Luca, C.; Del Guerra, S.; Cnop, M.; Eizirik, D.L.; Marchetti, P. A single-cell human islet interactome atlas identifies disrupted autocrine and paracrine communications in type 2 diabetes. NAR Genom. Bioinform. 2022, 4, lqac084. [Google Scholar] [CrossRef]
- Li, Y.; Chen, D.; Sun, L.; Wu, Y.; Zou, Y.; Liang, C.; Bao, Y.; Yi, J.; Zhang, Y.; Hou, J.; et al. Induced Expression of VEGFC, ANGPT, and EFNB2 and Their Receptors Characterizes Neovascularization in Proliferative Diabetic Retinopathy. Investig. Ophthalmol. Vis. Sci. 2019, 60, 4084–4096. [Google Scholar] [CrossRef] [Green Version]
- Dorjsembe, B.; Joo, H.; Nho, C.; Ham, J.; Kim, J.C. Aruncus dioicus var. kamtschaticus Extract Ameliorates Psoriasis-like Skin Inflammation via Akt/mTOR and JAK2/STAT3 Signaling Pathways in a Murine Model. Nutrients 2022, 14, 5094. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, H.; Dong, J.; Wang, Y.; Huang, Q.; Xu, J.; Qiu, Z.; Yao, F. Ginsenoside Rb1 Interfered with Macrophage Activation by Activating PPARγ to Inhibit Insulin Resistance in Obesity. Molecules 2023, 28, 3083. https://doi.org/10.3390/molecules28073083
Ding H, Dong J, Wang Y, Huang Q, Xu J, Qiu Z, Yao F. Ginsenoside Rb1 Interfered with Macrophage Activation by Activating PPARγ to Inhibit Insulin Resistance in Obesity. Molecules. 2023; 28(7):3083. https://doi.org/10.3390/molecules28073083
Chicago/Turabian StyleDing, Hongyue, Jinxiang Dong, Yuqi Wang, Qiang Huang, Jie Xu, Zhidong Qiu, and Fan Yao. 2023. "Ginsenoside Rb1 Interfered with Macrophage Activation by Activating PPARγ to Inhibit Insulin Resistance in Obesity" Molecules 28, no. 7: 3083. https://doi.org/10.3390/molecules28073083