
Combination Chemotherapy with Selected Polyphenols in Preclinical and Clinical Studies—An Update Overview



Abstract
:1. Introduction
2. Discussion and Future Perspectives
3. Materials and Methods
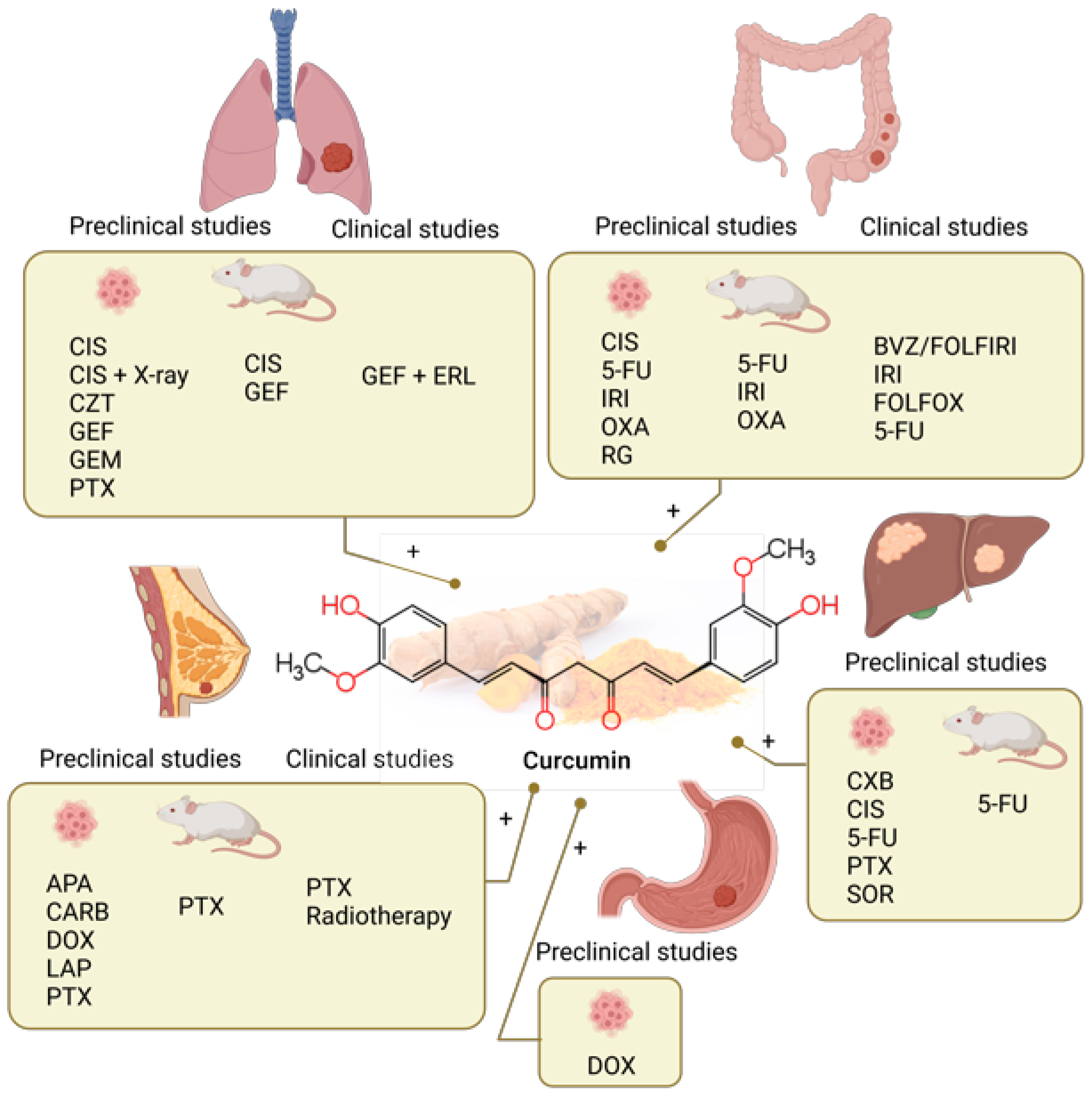
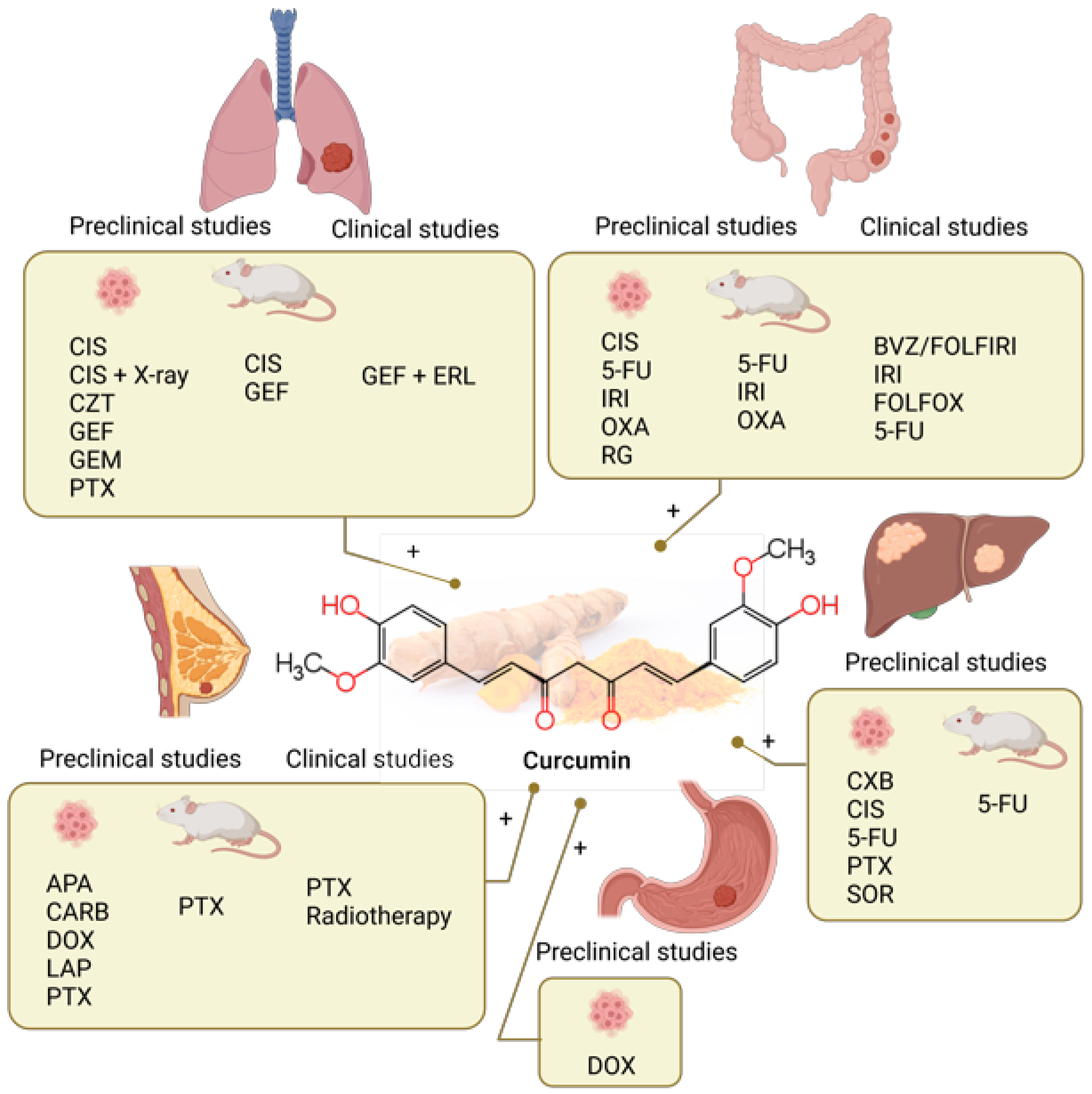
4. Curcumin
4.1. Curcumin Combined with Chemotherapy in Preclinical Studies
4.1.1. Lung Cancer
4.1.2. Colorectal Cancer
4.1.3. Liver Cancer
4.1.4. Stomach Cancer
4.1.5. Breast Cancer
4.2. Clinical Studies of Curcumin Combined with Chemotherapy
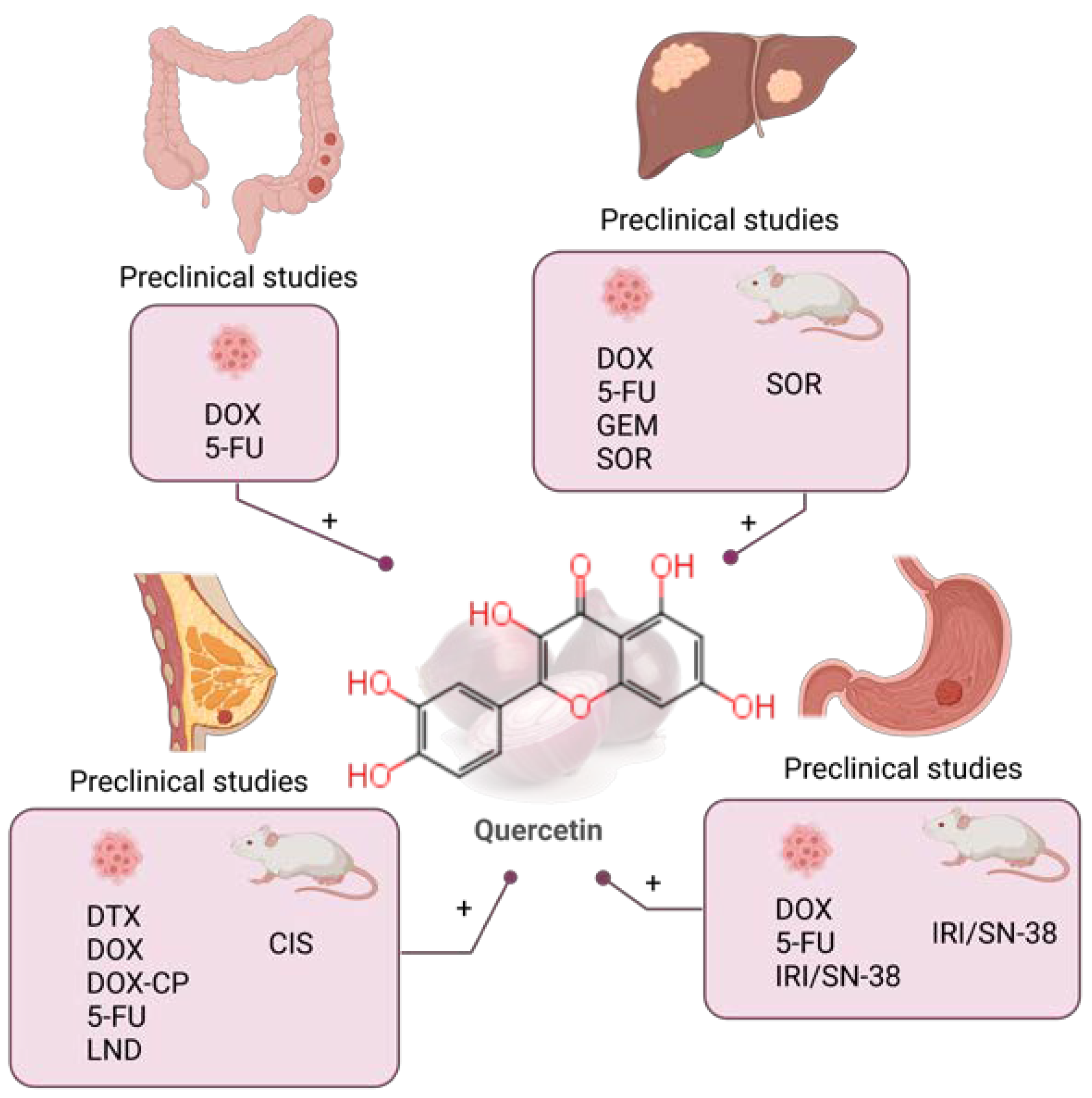
5. Quercetin
5.1. Quercetin Combined with Chemotherapy in Preclinical Studies
5.1.1. Colorectal Cancer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cancer Type | Chemotherapy | Dosage | Assay Type | Molecular Effect | Study Conclusion | Ref. |
|---|---|---|---|---|---|---|
| Colorectal cancer | DOX | 33 µM QUE + 0.5 µM DOX | SW620/DOX drug-resistant cell line and SW620/Ad300 cell line | Reversed P-gp-mediated drug resistance, increased intracellular DOX accumulation; modulates glutamine metabolism in DOX-resistant cells by inhibition of SLC1A5. | Reversed MDR, enhanced sensitivity to DOX. | [130] |
| 5-FU | 180 µg/mL QUE + 110 µg/mL 5-FU | HT-29 cell line | Decreased angiogenesis by inbibition of VEGF. | Synergistically enhanced the anticancer effect of 5-FU. | [131] | |
| 5-FU | 12 µg/mL QUE + 62.5, 125 µg/mL 5-FU | HCT-116 cell line | Enhanced apoptosis; suppression of Wnt/β-catenin signalling. | Enhanced 5-FU sensitivity. | [132] | |
| Liver cancer | DOX, 5-FU | 40–160 µM QUE + 0.2–125 µg/mL DOX/5-FU | BEL-7402 and BEL-7402/5-FU drug-resistant cell lines | Inhibition of FZD7/β-catenin pathway and ABCB1, ABCC1 and ABCC2 efflux pump. | Enhanced DOX and 5-FU sensitivity. | [133] |
| DOX | 0–100 μM QUE + 5–50 μM DOX | HepG2 cell line | Suppresses the efflux activity of MDR1, downregulates HIF-1α; increases apoptosis rate, upregulates p53 and cleaved caspase 3. | Enhances cytotoxic activity of DOX. | [134] | |
| GEM | 100–200 μM QUE + 38 mg/mL GEM | HepG2 cell line | Promotes apoptosis, induces S phase cycle cell arrest by upregulation of p53 and downregulation of cyclin D1. | Increases anticancer effect. | [135] | |
| SOR | In vitro: QUE 20–220 uM; SOR: 5–40 uM); SOR + QUE = 1:1:6.25 uM; 12.5 uM; 25 uM; 50 µM i 100 µM. In vivo: 7.5 mg/kg/day SOR, 2 h later 50 mg/kg/day QUE | HepG2 cell line and chemically induced HCC rat model | Suppressed proliferation, enhanced apoptosis and necrosis. | Synergistically increases anticancer effect and increases liver recovery. | [136] | |
| Gastric cancer | DOX | 100–200 μM QUE + 0.25–1.25 μM DOX | KATO III cell line | Enhanced apoptosis; upregulation of γH2As. | Increases chemotherapeutic effects. | [137] |
| 5-FU; DOX | 50 μM QUE + 25 μM 5-FU; 50 μM QUE + 0.5 μM DOX | AGS-cyr61 cell line | Reverses multidrug resistance; decreased CYR61, MRP1, and p65; induced caspase-dependent apoptosis; suppressed migration and down-regulation of EMT-related proteins; inhibits colony formations. | Strong synergistic effects with 5-FU and DOX. | [138] | |
| IRI/SN-38 | In vitro: 12.5, 50 μM QUE + 5, 25 nM SN-38. In vivo: 20 mg/kg i.p. injection | AGS-cyr61 cell line and AGS xenograft mouse model | In vitro: induces apoptosis, decreases cancer cell metastasis, downregulates β-catenin. In vivo: modulation of angiogenesis-associated and EMT-related factors. | Enhances cytotoxic effects of IRI/SN-38. | [139] | |
| Breast cancer | CIS | 30 mg/kg QUE + 7 mg/kg CIS | Breast tumor-bearing mouse model | Inhibited tumor growth and reduced renal toxicity. | Synergistic effect; inhibits renal toxicity induced by CIS. | [140] |
| DTX | 95 μM QUE + 7 nM DTX | MDA-MB-231 cell line | Inhibited cancer cell growth, induced apoptosis. | Enhances cytotoxic effects of DTX, decreases toxic effects. | [141] | |
| DOX | 0.7 μM QUE + 2 μg/mL DOX | MCF-10A, MCF-7 and MDA-MB-231 cell lines | Increased intracellular accumulation of DOX in cancer cells by downregulating the expression of P-gp, BCRP and MRP1; decreased cytotoxicity of DOX to non-tumoral MCF-10A mammary cells and myocardial AC16 cells. | Increases chemotherapeutic effects of DOX at a lower concentration; decreases the toxic side effects of DOX. | [142] | |
| 98 μM QUE + 0.35 μM DOX for MCF7 cells; 38 μM QUE + 0.35 μM DOX for MDA-MB-231 cells; 78 μM QUE + 0.35 μM DOX for T47D cells | MCF-7, MDA-MB-231 and T47D cell lines | Modulates vasoconstriction/vasodilatation induced by DOX; inhibited ROS generation; interferes DOX-induced cell cycle arrest; enhances intracellular concentration of doxorubicin in MDA-MB-231 and T47D cells by inhibition of P-gp. | Decreases cardiotoxicity; strong antagonistic interaction in MCF-7 and MDA-MB-231 cells. | [39] | ||
| DOX-CP | 20 μM QUE + (0.5 μg/mL DOX + 40 μg/mL CP); 1–40 μM QUE + (0.5 μg/mL DOX + 40 μg/mL CP) | MDA-MB-231 cell lines | Reduces cardiotoxicity by activating ERK1/2 pathway in cardiomyocytes; enhances the antitumor activity of DOX-CP by inhibiting ERK1/2 pathway in TNBC cells. | Enhances chemotherapeutic effects of DOX-CP; decreases DOX-CP induced cardiotoxicity. | [143] | |
| 5-FU | 150, 300, 446 μM QUE + 100 μM 5-FU | MCF-7 cell line | Enhanced apoptosis by increased expression of Bax and p53 and caspase-9 activity and decreasing the Bcl2 expression; decreased colony formation. | Enhances the sensitivity of breast cancer to 5-FU. | [144] | |
| 50, 200 μM QUE + 1.5, 6.25, 25 μM 5-FU | MDA-MB-231 cell line | Decreased migration rate and MMP-2 and MMP-9 gene expressions. | Synergistic effect. | [145] | ||
| LND | 80 μM QUE + 0.1, 1, 5 μM LND | MCF-7 cell line | Induced cell cycle arrest in the G2/M phase, arrested the cell cycle at S point; induced apoptosis by increased caspase levels, decreased MMP-2/-9 mRNA expression. | Synergistic effect. | [146] |

5.1.2. Liver Cancer
5.1.3. Gastric Cancer
5.1.4. Breast Cancer
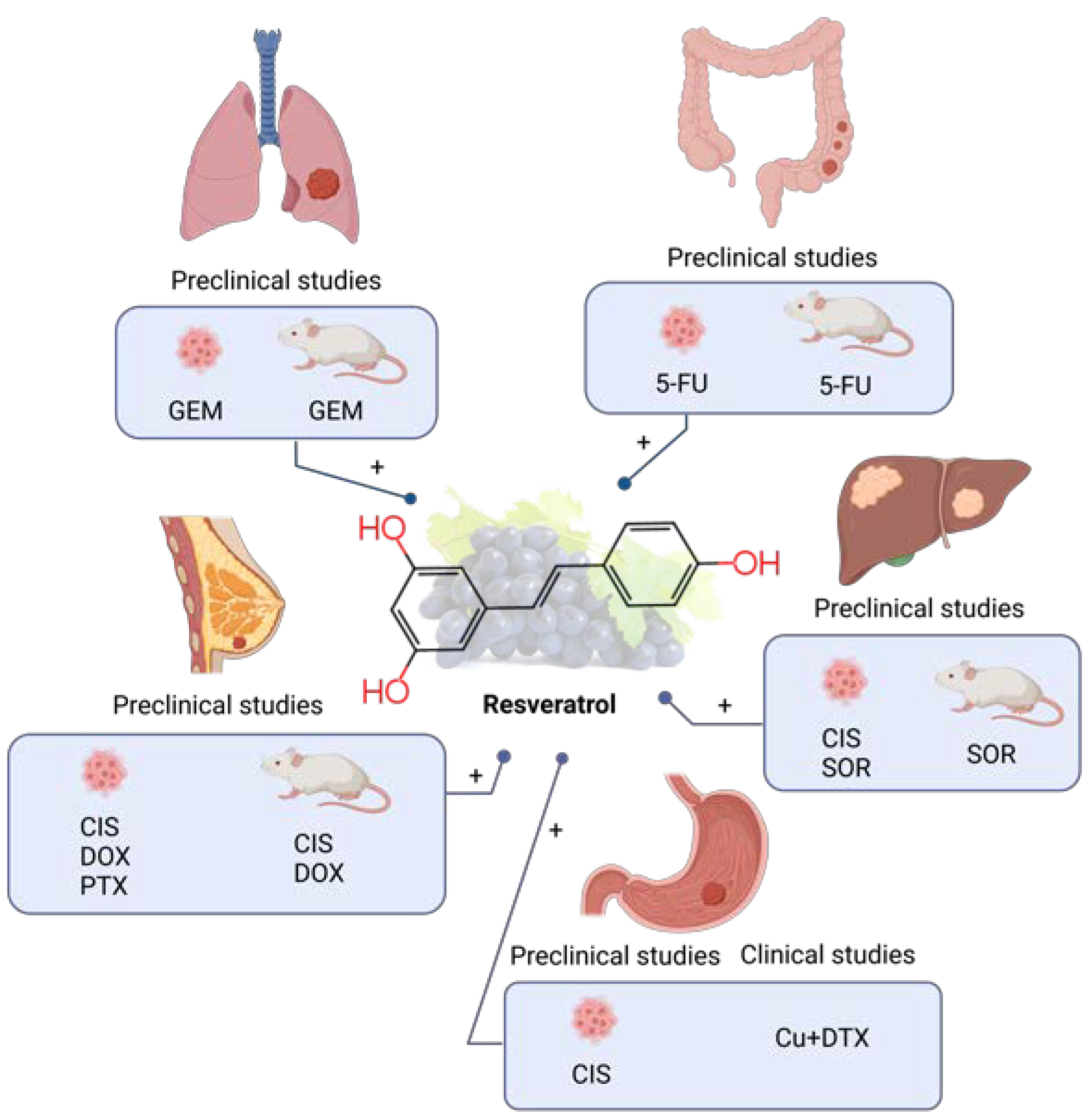
6. Resveratrol
6.1. Resveratrol Combined with Chemotherapy in Preclinical Studies
6.1.1. Lung Cancer
6.1.2. Colorectal Cancer
6.1.3. Liver Cancer
6.1.4. Gastric Cancer
6.1.5. Breast Cancer
6.2. Clinical Studies of Resveratrol Combined with Chemotherapy
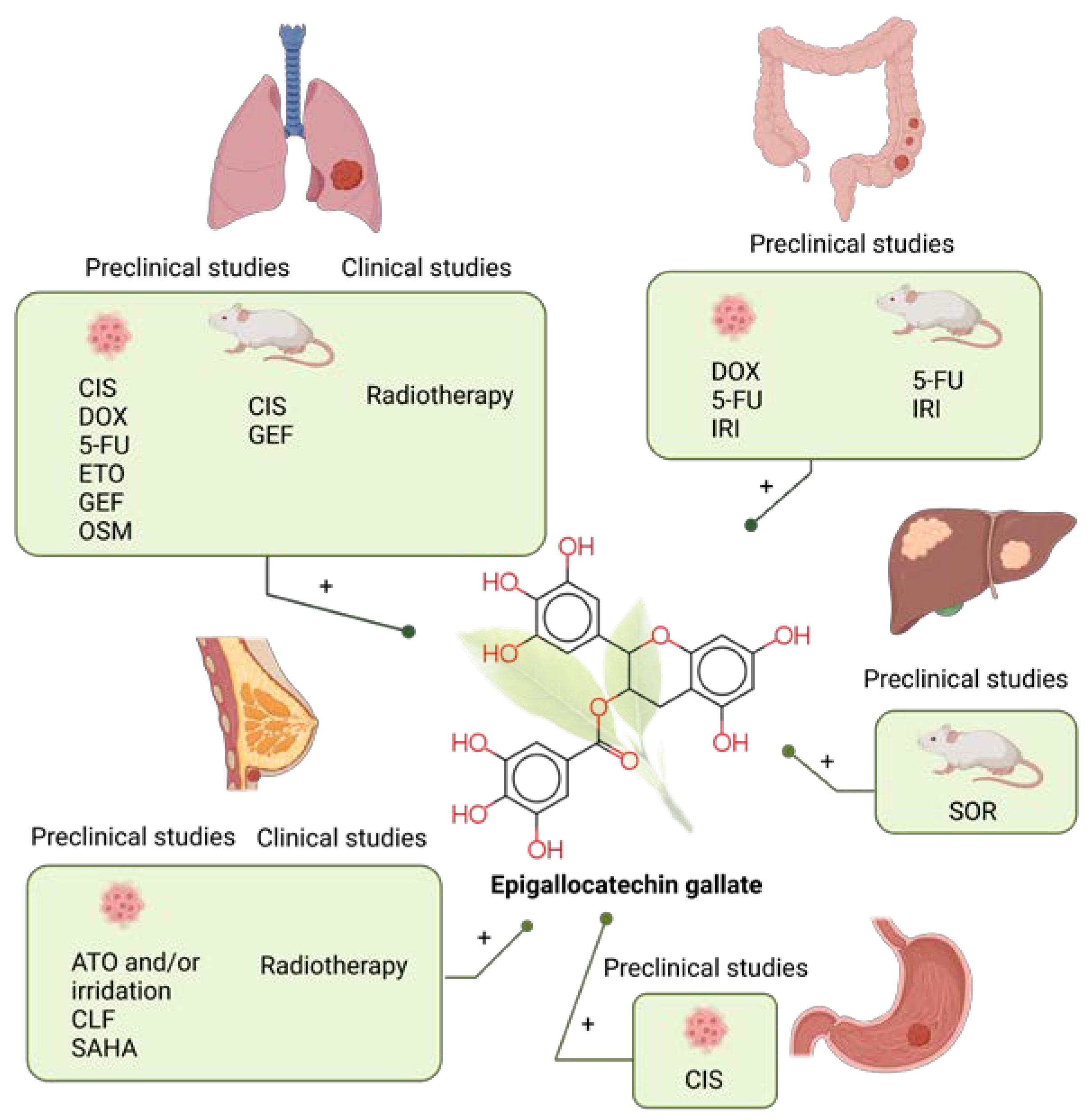
7. Epigallocatechin Gallate
7.1. Epigallocatechin Gallate Combined with Chemotherapy in Preclinical Studies
7.1.1. Lung Cancer
7.1.2. Colorectal Cancer
7.1.3. Liver Cancer
7.1.4. Gastric Cancer
7.1.5. Breast Cancer
7.2. Clinical Studies of Epigallocatechin Gallate Combined with Chemotherapy
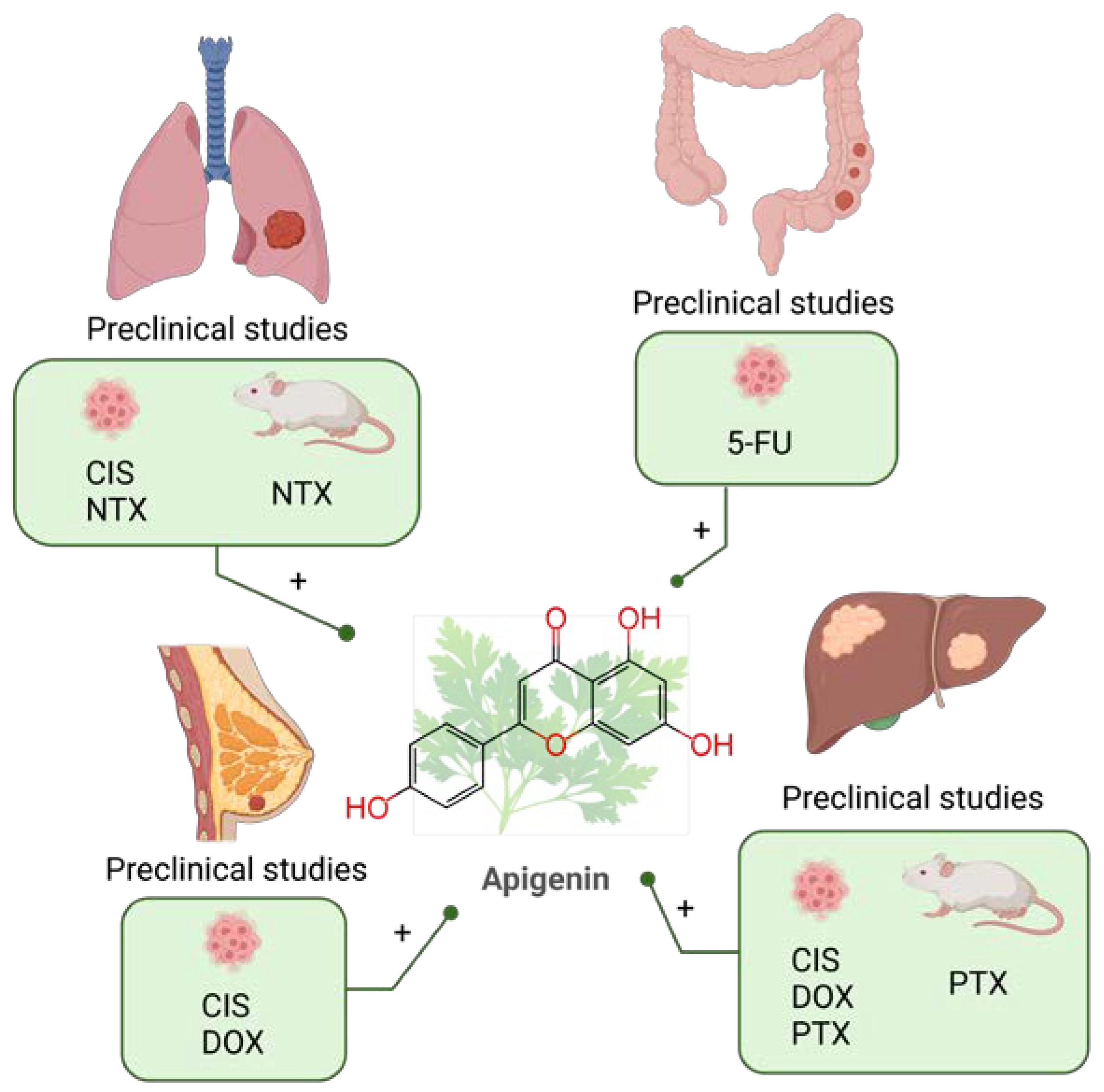
8. Apigenin
8.1. Apigenin Combined with Chemotherapy in Preclinical Studies
8.1.1. Lung Cancer
| Cancer Type | Chemotherapy | Dosage | Assay Type | Molecular Effect | Study Conclusion | Ref. |
|---|---|---|---|---|---|---|
| Lung cancer | CIS | 5–20 µM AP + 10 µM CIS for A549 cells; 10–20 µM AP + 10 µM CIS for A549R cells; 20–30 µM AP + 20 µM CIS for H1299 cells | A549, A549R and H1299 cell lines | Upregulation of p53. | AP enhanced the antitumor effect of CIS in A549, H1299, and A549R cells. | [203] |
| AP:CIS drug ratio 5:1 for A549 cells; 10:1 for H460 cells; 4:1 for H1299 cells | A549, H460 and H1299 cell lines | S phase prolongation and G2/M cell cycle arrest. Inducing p21 and PUMA. | AP enhances the anticancer effect of CIS by inducing apoptosis and arresting the cell cycle. | [18] | ||
| NTX | In vitro: 10–20 μM AP + 1–2 μM ABT-263. In vivo: 25 mg/kg AP + 100 mg/kg ABT-263 | H1975, HCC827, H1650 and H3255 cell lines and BALB/c nude mice | Upregulated the expression of Noxa by targeting the AKT-FoxO3a pathway and inhibited ERK. | AP synergized with ABT-263 by suppressing the growth and proliferation of tumor cells in vitro and in vivo. | [205] | |
| Colorectal cancer | 5-FU | 20 µM AP + 20 µM 5-FU | HCT116 and HT29 cell lines | Inhibited the upregulation of TS induced by 5-FU. Increasing reactive oxygen species production, intracellular and intramitochondrial Ca2+ concentrations, and mitochondrial membrane potential. | AP enhanced the efficacy of 5-FU by potentiating HCT116 cell apoptosis and enhancing cell cycle disruption. Acquired resistance to 5-FU was reduced. | [206] |
| 0.1–100 μM AP + 1 μM 5-FU | WiDr cell line | AP and 5-FU exerted synergistic effects in WiDr cells. | [184] | |||
| Liver cancer | CIS | 10 μM–20 μM AP + 0.025–5 μg/mL CIS | HepG2, Hep3B, and Huh7 cell lines | Cell cycle delays during the first mitotic division in Hep3B and Huh7 cells and the second mitotic division in HepG2 cells. | AP enhanced the genotoxic, cytotoxic, anti-invasive and anti-migratory effects of CIS. | [207] |
| DOX | 25–200 μM AP + 1 μM DOX | HepG2 cell line | Inhibition of glycolytic genes expression (hexokinase 2 and lactate dehydrogenase A). | The combination of AP and DOX exhibited a very effective cytotoxic mechanism in HepG2 cells. | [208] | |
| PTX | In vitro: 40 mM AP + 6.25–100 nM PTX. In vivo: 1 mg/kg/day AP + 3.5 mg/kg/day PTX | HepG2 cell line and Balb/c nude mice | Suppressing the intratumoral expression of HIF-1a via inhibiting the AKT/p-AKT pathway and the expression of HSP90 simultaneously. | AP reduced hypoxia-induced PTX resistance in hypoxic tumors. | [209] | |
| Breast cancer | CIS | 5–100 μg/mL AP + 5–100 μg/mL CIS | MDA-MB-231 and HCC1806 cell lines | Inhibition of telomerase activity. Down-regulation of hTERT, Hsp90 and p23 at transcriptional and translational level. | AP and CIS synergistically inhibited telomerase activities by reducing the catalytic subunit of the enzyme. | [210] |
| DOX | 50 μM AP + 1 μM DOX | MCF-7 cell line | Reduced population of cells in G1 phase. Reduced AP site level. Increased phosphorylated H2AX in the nucleus. | AP enhanced the cytotoxic effect of DOX by increasing the formation of DNA damage and decreasing the expression of DNA repair genes. | [211] |

8.1.2. Colorectal Cancer
8.1.3. Liver Cancer
8.1.4. Breast Cancer
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- National Institutes of Health (US). Understanding Cancer—NIH Curriculum Supplement Series. Available online: https://www.ncbi.nlm.nih.gov/books/NBK20362/ (accessed on 27 January 2023).
- World Health Organization (WHO). Cancer. 2023. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 27 January 2023).
- Travis, W.D. Lung Cancer Pathology. Clin. Chest Med. 2020, 41, 67–85. [Google Scholar] [CrossRef] [PubMed]
- Dekker, E.; Tanis, P.J.; Vleugels, J.L.A.; Kasi, P.M.; Wallace, M.B. Colorectal Cancer. Lancet 2019, 394, 1467–1480. [Google Scholar] [CrossRef] [PubMed]
- Mármol, I.; Sánchez-de-Diego, C.; Pradilla Dieste, A.; Cerrada, E.; Rodriguez Yoldi, M. Colorectal Carcinoma: A General Overview and Future Perspectives in Colorectal Cancer. Int. J. Mol. Sci. 2017, 18, 197. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization (WHO). Breast Cancer. 2020. Available online: https://gco.iarc.fr/today/data/factsheets/cancers/20-Breast-fact-sheet.pdf (accessed on 27 January 2023).
- Kent, E.E.; Forsythe, L.P.; Yabroff, K.R.; Weaver, K.E.; de Moor, J.S.; Rodriguez, J.L.; Rowland, J.H. Are Survivors Who Report Cancer-Related Financial Problems More Likely to Forgo or Delay Medical Care? Cancer 2013, 119, 3710–3717. [Google Scholar] [CrossRef] [PubMed]
- Alfarouk, K.O.; Stock, C.-M.; Taylor, S.; Walsh, M.; Muddathir, A.K.; Verduzco, D.; Bashir, A.H.H.; Mohammed, O.Y.; Elhassan, G.O.; Harguindey, S.; et al. Resistance to Cancer Chemotherapy: Failure in Drug Response from ADME to P-Gp. Cancer Cell Int. 2015, 15, 71. [Google Scholar] [CrossRef]
- Oun, R.; Moussa, Y.E.; Wheate, N.J. The side effects of platinum-based chemotherapy drugs: A review for chemists. Dalton Trans. 2018, 47, 6645–6653. [Google Scholar] [CrossRef]
- Oyenihi, A.B.; Smith, C. Are Polyphenol Antioxidants at the Root of Medicinal Plant Anti-Cancer Success? J. Ethnopharmacol. 2019, 229, 54–72. [Google Scholar] [CrossRef]
- Patra, S.; Mishra, S.R.; Behera, B.P.; Mahapatra, K.K.; Panigrahi, D.P.; Bhol, C.S.; Praharaj, P.P.; Sethi, G.; Patra, S.K.; Bhutia, S.K. Autophagy-Modulating Phytochemicals in Cancer Therapeutics: Current Evidences and Future Perspectives. Semin. Cancer Biol. 2022, 80, 205–217. [Google Scholar] [CrossRef]
- Gao, X.; Xu, Z.; Liu, G.; Wu, J. Polyphenols as a Versatile Component in Tissue Engineering. Acta Biomater. 2021, 119, 57–74. [Google Scholar] [CrossRef]
- Durazzo, A.; Lucarini, M.; Souto, E.B.; Cicala, C.; Caiazzo, E.; Izzo, A.A.; Novellino, E.; Santini, A. Polyphenols: A Concise Overview on the Chemistry, Occurrence, and Human Health. Phytother. Res. 2019, 33, 2221–2243. [Google Scholar] [CrossRef]
- Mithul Aravind, S.; Wichienchot, S.; Tsao, R.; Ramakrishnan, S.; Chakkaravarthi, S. Role of Dietary Polyphenols on Gut Microbiota, Their Metabolites and Health Benefits. Food Res. Internat. 2021, 142, 110189. [Google Scholar] [CrossRef]
- Rasouli, H.; Dehghan, N.F.; Khodarahmi, R. May phytophenolics alleviate aflatoxins-induced health challenges? A holistic insight on current landscape and future prospects. Front. Nutr. 2022, 9, 981984. [Google Scholar] [CrossRef]
- Weisel, K.C.; Morgner-Miehlke, A.; Petersen, C.; Fiedler, W.; Block, A.; Schafhausen, P.; Knobloch, J.K.; Bokemeyer, C. Implications of SARS-CoV-2 Infection and COVID-19 Crisis on Clinical Cancer Care: Report of the University Cancer Center Hamburg. Oncol. Res. Treat. 2020, 43, 307–313. [Google Scholar] [CrossRef]
- Ivanyi, P.; Park-Simon, T.; Christiansen, H.; Gutzmer, R.; Vogel, A.; Heuser, M.; Golpon, H.; Hillemanns, P.; Haier, J. Protective Measures for Patients with Advanced Cancer during the SARS-CoV-2 Pandemic: Quo Vadis? Clin. Exp. Metastasis 2021, 38, 257–261. [Google Scholar] [CrossRef]
- Nurgali, K.; Jagoe, R.T.; Abalo, R. Editorial: Adverse Effects of Cancer Chemotherapy: Anything New to Improve Tolerance and Reduce Sequelae? Front. Pharmacol. 2018, 9, 245. [Google Scholar] [CrossRef]
- Glass, C.K.; Mitchell, R.N. Winning the Battle, but Losing the War: Mechanisms and Morphology of Cancer-Therapy-Associated Cardiovascular Toxicity. Cardiovasc. Pathol. 2017, 30, 55–63. [Google Scholar] [CrossRef]
- Garcia-Oliveira, P.; Otero, P.; Pereira, A.G.; Chamorro, F.; Carpena, M.; Echave, J.; Fraga-Corral, M.; Simal-Gandara, J.; Prieto, M.A. Status and Challenges of Plant-Anticancer Compounds in Cancer Treatment. Pharmaceuticals 2021, 14, 157. [Google Scholar] [CrossRef]
- Bouyahya, A.; Omari, N.E.; Bakrim, S.; Hachlafi, N.E.; Balahbib, A.; Wilairatana, P.; Mubarak, M.S. Advances in Dietary Phenolic Compounds to Improve Chemosensitivity of Anticancer Drugs. Cancers 2022, 14, 4573. [Google Scholar] [CrossRef]
- Abotaleb, M.; Liskova, A.; Kubatka, P.; Büsselberg, D. Therapeutic Potential of Plant Phenolic Acids in the Treatment of Cancer. Biomolecules 2020, 10, 221. [Google Scholar] [CrossRef]
- Anantharaju, P.G.; Gowda, P.C.; Vimalambike, M.G.; Madhunapantula, S.V. An Overview on the Role of Dietary Phenolics for the Treatment of Cancers. Nutr. J. 2016, 15, 99. [Google Scholar] [CrossRef]
- Liu, Y.-Q.; Wang, X.-L.; He, D.-H.; Cheng, Y.-X. Protection against Chemotherapy- and Radiotherapy-Induced Side Effects: A Review Based on the Mechanisms and Therapeutic Opportunities of Phytochemicals. Phytomedicine 2021, 80, 153402. [Google Scholar] [CrossRef] [PubMed]
- Khatoon, E.; Banik, K.; Harsha, C.; Sailo, B.L.; Thakur, K.K.; Khwairakpam, A.D.; Vikkurthi, R.; Devi, T.B.; Gupta, S.C.; Kunnumakkara, A.B. Phytochemicals in Cancer Cell Chemosensitization: Current Knowledge and Future Perspectives. Semin. Cancer Biol. 2022, 80, 306–339. [Google Scholar] [CrossRef]
- Arrigoni, R.; Ballini, A.; Santacroce, L.; Cantore, S.; Inchingolo, A.M.; Inchingolo, F.; Di Domenico, M.; Quagliuolo, L.; Boccellino, M. Another Look at Dietary Polyphenols: Challenges in Cancer Prevention and Treatment. Cur. Med. Chem. 2022, 29, 1061–1082. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.-P.; Li, S.; Chen, Y.-M.; Li, H.-B. Natural Polyphenols for Prevention and Treatment of Cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef] [PubMed]
- Rudrapal, M.; Khairnar, S.J.; Khan, J.; Dukhyil, A.B.; Ansari, M.A.; Alomary, M.N.; Alshabrmi, F.M.; Palai, S.; Deb, P.K.; Devi, R. Dietary Polyphenols and Their Role in Oxidative Stress-Induced Human Diseases: Insights into Protective Effects, Antioxidant Potentials and Mechanism(s) of Action. Front. Pharmacol. 2022, 13, 806470. [Google Scholar] [CrossRef] [PubMed]
- Rahman, M.; Rahaman, S.; Islam, R.; Rahman, F.; Mithi, F.M.; Alqahtani, T.; Almikhlafi, M.A.; Alghamdi, S.Q.; Alruwaili, A.S.; Hossain, S.; et al. Role of Phenolic Compounds in Human Disease: Current Knowledge and Future Prospects. Molecules 2021, 27, 233. [Google Scholar] [CrossRef]
- Dini, I.; Grumetto, L. Recent Advances in Natural Polyphenol Research. Molecules 2022, 27, 8777. [Google Scholar] [CrossRef]
- Almatroodi, S.A.; Alsahli, M.A.; Aljohani, A.S.M.; Aljohani, A.; Alhumaydhi, F.A.; Babiker, A.Y.; Khan, A.A.; Rahmani, A.H. Potential Therapeutic Targets of Resveratrol, a Plant Polyphenol, and Its Role in the Therapy of Various Types of Cancer. Molecules 2022, 27, 2665. [Google Scholar] [CrossRef]
- Mitra, S.; Tareq, A.M.; Das, R.; Emran, T.B.; Nainu, F.; Chakraborty, A.J.; Ahmad, I.; Tallei, T.E.; Idris, A.M.; Simal-Gandara, J. Polyphenols: A First Evidence in the Synergism and Bioactivities. Food Rev. Int. 2022, 1–23. [Google Scholar] [CrossRef]
- Vladu, A.F.; Ficai, D.; Ene, A.G.; Ficai, A. Combination Therapy Using Polyphenols: An Efficient Way to Improve Antitumoral Activity and Reduce Resistance. Int. J. Mol. Sci. 2022, 23, 10244. [Google Scholar] [CrossRef]
- Moutabian, H.; Majdaeen, M.; Ghahramani-Asl, R.; Yadollahi, M.; Gharepapagh, E.; Ataei, G.; Falahatpour, Z.; Bagheri, H.; Farhood, B. A Systematic Review of the Therapeutic Effects of Resveratrol in Combination with 5-Fluorouracil during Colorectal Cancer Treatment: With a Special Focus on the Oxidant, Apoptotic, and Anti-Inflammatory Activities. Cancer Cell Int. 2022, 22, 142. [Google Scholar] [CrossRef]
- Yan, X.H.; Qi, M.; Li, P.F.; Zhan, Y.H.; Shao, H.J. Apigenin in cancer therapy: Anti-cancer effects and mechanisms of action. Cell Biosci. 2017, 7, 50. [Google Scholar] [CrossRef]
- Kong, W.-Y.; Ngai, S.C.; Goh, B.-H.; Lee, L.-H.; Htar, T.-T.; Chuah, L.-H. Is CUR the Answer to Future Chemotherapy Cocktail? Molecules 2021, 26, 4329. [Google Scholar] [CrossRef]
- Lai, H.; Chien, S.; Kuo, S.; Tseng, L.; Lin, H.; Chi, C.; Chen, D. The Potential Utility of Curcumin in the Treatment of HER-2-Overexpressed Breast Cancer: An In Vitro and In Vivo Comparison Study with Herceptin. Evid Based Complement. Altern. Med. 2012, 2012, 486568. [Google Scholar] [CrossRef]
- Abdallah, F.M.; Helmy, M.W.; Katary, M.A.; Ghoneim, A.I. Synergistic Antiproliferative Effects of CUR and Celecoxib in Hepatocellular Carcinoma HepG2 Cells. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2018, 391, 1399–1410. [Google Scholar] [CrossRef]
- Henidi, H.A.; Al-Abbasi, F.A.; El-Moselhy, M.A.; El-Bassossy, H.M.; Al-Abd, A.M. Despite Blocking Doxorubicin-Induced Vascular Damage, Quercetin Ameliorates Its Antibreast Cancer Activity. Oxid. Med. Cell. Longev. 2020, 2020, 8157640. [Google Scholar] [CrossRef]
- Boretti, A. Quercetin as a Cancer Chemopreventive or Chemotherapeutic Agent: Where We Stand. Phytother. Res. 2022, 37, ptr.7699. [Google Scholar] [CrossRef]
- Younes, M.; Mardirossian, R.; Rizk, L.; Fazlian, T.; Khairallah, J.P.; Sleiman, C.; Naim, H.Y.; Rizk, S. The Synergistic Effects of CUR and Chemotherapeutic Drugs in Inhibiting Metastatic, Invasive and Proliferative Pathways. Plants 2022, 11, 2137. [Google Scholar] [CrossRef]
- Shaito, A.; Posadino, A.M.; Younes, N.; Hasan, H.; Halabi, S.; Alhababi, D.; Al-Mohannadi, A.; Abdel-Rahman, W.M.; Eid, A.H.; Nasrallah, G.K.; et al. Potential Adverse Effects of Resveratrol: A Literature Review. Int. J. Mol. Sci. 2020, 21, 2084. [Google Scholar] [CrossRef]
- Morón, E.B.; Calderón-Montaño, J.M.; Salvador, J.; Robles, A.; López-Lázaro, M. The dark side of CUR. Int. J. Cancer 2010, 126, 1771–1775. [Google Scholar] [CrossRef]
- Reyes-Farias, M.; Carrasco-Pozo, C. The Anti-Cancer Effect of Quercetin: Molecular Implications in Cancer Metabolism. Int. J. Mol. Sci. 2019, 20, 3177. [Google Scholar] [CrossRef] [PubMed]
- Eghbaliferiz, S.; Iranshahi, M. Prooxidant Activity of Polyphenols, Flavonoids, Anthocyanins and Carotenoids: Updated Review of Mechanisms and Catalyzing Metals: Prooxidant Activity of Polyphenols and Carotenoids. Phytother. Res. 2016, 30, 1379–1391. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Jia, L.; Zhou, H.-M.; Liu, Y.; Zhong, L.-F. Mitochondrial and Nuclear DNA Damage Induced by CUR in Human Hepatoma G2 Cells. Toxicol. Sci. 2006, 91, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Ingold, K.U.; Pratt, D.A. Advances in Radical-Trapping Antioxidant Chemistry in the 21st Century: A Kinetics and Mechanisms Perspective. Chem. Rev. 2014, 114, 9022–9046. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Yu, Y.; Lv, H.; Zhang, H.; Liang, T.; Zhou, G.; Huang, L.; Tian, Y.; Liang, W. Apigenin in Cancer Therapy: From Mechanism of Action to Nano-Therapeutic Agent. Food Chem. Toxicol. 2022, 168, 113385. [Google Scholar] [CrossRef] [PubMed]
- Rudrapal, M.; Mishra, A.K.; Rani, L.; Sarwa, K.K.; Zothantluanga, J.H.; Khan, J.; Kamal, M.; Palai, S.; Bendale, A.R.; Talele, S.G.; et al. Nanodelivery of Dietary Polyphenols for Therapeutic Applications. Molecules 2022, 27, 8706. [Google Scholar] [CrossRef]
- Gavas, S.; Quazi, S.; Karpiński, T.M. Nanoparticles for Cancer Therapy: Current Progress and Challenges. Nanoscale. Res. Lett. 2021, 16, 173. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lin, Z.; Ju, Y.; Rahim, A.; Richardson, J.J.; Caruso, F. Polyphenol-Mediated Assembly for Particle Engineering. Acc. Chem. Res. 2020, 53, 1269–1278. [Google Scholar] [CrossRef] [PubMed]
- Davatgaran-Taghipour, Y.; Masoomzadeh, S.; Farzaei, M.H.; Bahramsoltani, R.; Karimi-Soureh, Z.; Rahimi, R.; Abdollahi, M. Polyphenol Nanoformulations for Cancer Therapy: Experimental Evidence and Clinical Perspective. Int. J. Nanomed. 2017, 12, 2689–2702. [Google Scholar] [CrossRef]
- Vieira, I.R.S.; Conte-Junior, C.A. Nano-Delivery Systems for Food Bioactive Compounds in Cancer: Prevention, Therapy, and Clinical Applications. Crit. Rev. Food Sci. Nutr. 2022. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. 2023. Available online: https://clinicaltrials.gov/ (accessed on 27 January 2023).
- Condezo-Hoyos, L.; Gazi, C.; Perez-Jimenez, J. Design of polyphenol-rich diets in clinical trials: A systematic review. Food Res. Int. 2021, 149, 110655. [Google Scholar] [CrossRef] [PubMed]
- Neveu, V.; Perez-Jiménez, J.; Vos, F.; Crespy, V.; du Chaffaut, L.; Mennen, L.; Knox, C.; Eisner, R.; Cruz, J.; Wishart, D.; et al. Phenol-Explorer: An online comprehensive database on polyphenol contents in foods. Database 2010, 2010, bap024. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.A.; Urpi-Sarda, M.; Boto-Ordoñez, M.; Knox, C.; Llorach, R.; Eisner, R.; Cruz, J.; Neveu, V.; Wishart, D.; Manach, C.; et al. Phenol-Explorer 2.0: A major update of the Phenol-Explorer database integrating data on polyphenol metabolism and pharmacokinetics in humans and experimental animals. Database 2012, 2012, bas031. [Google Scholar] [CrossRef] [PubMed]
- Rothwell, J.A.; Pérez-Jiménez, J.; Neveu, V.; Medina-Ramon, A.; M’Hiri, N.; Garcia Lobato, P.; Manach, C.; Knox, K.; Eisner, R.; Wishart, D.; et al. Phenol-Explorer 3.0: A major update of the Phenol-Explorer database to incorporate data on the effects of food processing on polyphenol content. Database 2013, 2013, bat070. [Google Scholar] [CrossRef] [PubMed]
- Web of Science. Clarivate. Available online: https://www.webofscience.com/wos/woscc/basic-search (accessed on 27 January 2023).
- Urošević, M.; Nikolić, L.; Gajić, I.; Nikolić, V.; Dinić, A.; Miljković, V. CUR: Biological Activities and Modern Pharmaceutical Forms. Antibiotics 2022, 11, 135. [Google Scholar] [CrossRef]
- Tang, C.; Liu, J.; Yang, C.; Ma, J.; Chen, X.; Liu, D.; Zhou, Y.; Zhou, W.; Lin, Y.; Yuan, X. CUR and Its Analogs in Non-Small Cell Lung Cancer Treatment: Challenges and Expectations. Biomolecules 2022, 12, 1636. [Google Scholar] [CrossRef]
- Kharat, M.; Du, Z.; Zhang, G.; McClements, D.J. Physical and Chemical Stability of CUR in Aqueous Solutions and Emulsions: Impact of pH, Temperature, and Molecular Environment. J. Agric. Food Chem. 2017, 65, 1525–1532. [Google Scholar] [CrossRef]
- Ghoran, S.H.; Calcaterra, A.; Abbasi, M.; Taktaz, F.; Nieselt, K.; Babaei, E. CUR-Based Nanoformulations: A Promising Adjuvant towards Cancer Treatment. Molecules 2022, 27, 5236. [Google Scholar] [CrossRef]
- Menon, L.G.; Kuttan, R.; Kuttan, G. Inhibition of Lung Metastasis in Mice Induced by B16F10 Melanoma Cells by Polyphenolic Compounds. Cancer Lett. 1995, 95, 221–225. [Google Scholar] [CrossRef]
- Zhang, W.; Jiang, H.; Chen, Y.; Ren, F. Resveratrol Chemosensitizes Adriamycin-Resistant Breast Cancer Cells by Modulating miR-122-5p. J. Cell. Biochem. 2019, 120, 16283–16292. [Google Scholar] [CrossRef]
- Wu, S.-H.; Hang, L.-W.; Yang, J.-S.; Chen, H.-Y.; Lin, H.-Y.; Chiang, J.-H.; Lu, C.-C.; Yang, J.-L.; Lai, T.-Y.; Ko, Y.-C.; et al. CUR Induces Apoptosis in Human Non-Small Cell Lung Cancer NCI-H460 Cells through ER Stress and Caspase Cascade- and Mitochondria-Dependent Pathways. Anticancer Res. 2010, 30, 2125–2133. [Google Scholar] [PubMed]
- Wang, M.; Jiang, S.; Zhou, L.; Yu, F.; Ding, H.; Li, P.; Zhou, M.; Wang, K. Potential Mechanisms of Action of CUR for Cancer Prevention: Focus on Cellular Signaling Pathways and MiRNAs. Int. J. Biol. Sci. 2019, 15, 1200–1214. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.-F.; Hsieh, T.-H.; Lee, J.-N.; Hsu, C.-Y.; Wang, Y.-C.; Kuo, K.-K.; Wu, H.-L.; Chiu, C.-C.; Tsai, E.-M.; Kuo, P.-L. CUR Suppresses Phthalate-Induced Metastasis and the Proportion of Cancer Stem Cell (CSC)-like Cells via the Inhibition of AhR/ERK/SK1 Signaling in Hepatocellular Carcinoma. J. Agric. Food Chem. 2015, 63, 10388–10398. [Google Scholar] [CrossRef] [PubMed]
- Chen, P.; Li, J.; Jiang, H.-G.; Lan, T.; Chen, Y.-C. CUR Reverses Cisplatin Resistance in Cisplatin-Resistant Lung Caner Cells by Inhibiting FA/BRCA Pathway. Tumor. Biol. 2015, 36, 3591–3599. [Google Scholar] [CrossRef] [PubMed]
- Ye, M.; Zhang, J.; Zhang, J.; Miao, Q.; Yao, L.; Zhang, J. CUR Promotes Apoptosis by Activating the P53-MiR-192-5p/215-XIAP Pathway in Non-Small Cell Lung Cancer. Cancer Lett. 2015, 357, 196–205. [Google Scholar] [CrossRef]
- Pongrakhananon, V.; Nimmannit, U.; Luanpitpong, S.; Rojanasakul, Y.; Chanvorachote, P. CUR Sensitizes Non-Small Cell Lung Cancer Cell Anoikis through Reactive Oxygen Species-Mediated Bcl-2 Downregulation. Apoptosis 2010, 15, 574–585. [Google Scholar] [CrossRef]
- Tang, X.; Ding, H.; Liang, M.; Chen, X.; Yan, Y.; Wan, N.; Chen, Q.; Zhang, J.; Cao, J. CUR Induces Ferroptosis in Non-Small-Cell Lung Cancer via Activating Autophagy. Thorac. Cancer 2021, 12, 1219–1230. [Google Scholar] [CrossRef]
- Mirza, S.; Vasaiya, A.; Vora, H.; Jain, N.; Rawal, R. CUR Targets Circulating Cancer Stem Cells by Inhibiting Self-Renewal Efficacy in Non-Small Cell Lung Carcinoma. Anti-Cancer Agents Med. Chem. 2017, 17, 859–864. [Google Scholar] [CrossRef]
- Li, F.; Chen, X.; Xu, B.; Zhou, H. CUR Induces P53-Independent Necrosis in H1299 Cells via a Mitochondria-Associated Pathway. Mol. Med. Rep. 2015, 12, 7806–7814. [Google Scholar] [CrossRef]
- Su, C.-C.; Lin, J.-G.; Li, T.-M.; Chung, J.-G.; Yang, J.-S.; Ip, S.-W.; Lin, W.-C.; Chen, G.-W. CUR-Induced Apoptosis of Human Colon Cancer Colo 205 Cells through the Production of ROS, Ca2+ and the Activation of Caspase-3. Anticancer Res. 2006, 26, 4379–4390. [Google Scholar] [PubMed]
- Mosieniak, G.; Adamowicz, M.; Alster, O.; Jaskowiak, H.; Szczepankiewicz, A.A.; Wilczynski, G.M.; Ciechomska, I.A.; Sikora, E. CUR Induces Permanent Growth Arrest of Human Colon Cancer Cells: Link between Senescence and Autophagy. Mech. Ageing Dev. 2012, 133, 444–455. [Google Scholar] [CrossRef]
- Chang, B.-D.; Broude, E.V.; Fang, J.; Kalinichenko, T.V.; Abdryashitov, R.; Poole, J.C.; Roninson, I.B. P21Waf1/Cip1/Sdi1-Induced Growth Arrest Is Associated with Depletion of Mitosis-Control Proteins and Leads to Abnormal Mitosis and Endoreduplication in Recovering Cells. Oncogene 2000, 19, 2165–2170. [Google Scholar] [CrossRef] [PubMed]
- Watson, J.L.; Hill, R.; Yaffe, P.B.; Greenshields, A.; Walsh, M.; Lee, P.W.; Giacomantonio, C.A.; Hoskin, D.W. CUR Causes Superoxide Anion Production and P53-Independent Apoptosis in Human Colon Cancer Cells. Cancer Lett. 2010, 297, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Darvesh, A.S.; Aggarwal, B.B.; Bishayee, A. CUR and Liver Cancer: A Review. Curr. Pharm. Biotechnol. 2012, 13, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Pan, Z.; Zhuang, J.; Ji, C.; Cai, Z.; Liao, W.; Huang, Z. CUR Inhibits Hepatocellular Carcinoma Growth by Targeting VEGF Expression. Oncol. Lett. 2018, 15, 4821–4826. [Google Scholar] [CrossRef]
- Amoodizaj, F.F.; Baghaeifar, S.; Taheri, E.; Jadid, M.F.S.; Safi, M.; Sani, N.S.; Hajazimian, S.; Isazadeh, A.; Shanehbandi, D. Enhanced Anticancer Potency of Doxorubicin in Combination with CUR in Gastric Adenocarcinoma. J. Biochem. Mol. Toxicol. 2020, 34, e22486. [Google Scholar] [CrossRef]
- Farghadani, R.; Naidu, R. CUR as an Enhancer of Therapeutic Efficiency of Chemotherapy Drugs in Breast Cancer. Int. J. Mol. Sci. 2022, 23, 2144. [Google Scholar] [CrossRef]
- Abdul Satar, N.; Ismail, M.N.; Yahaya, B.H. Synergistic Roles of CUR in Sensitising the Cisplatin Effect on a Cancer Stem Cell-Like Population Derived from Non-Small Cell Lung Cancer Cell Lines. Molecules 2021, 26, 1056. [Google Scholar] [CrossRef]
- Cai, Y.; Sheng, Z.; Liang, S. Radiosensitization Effects of CUR plus Cisplatin on Non-small Cell Lung Cancer A549 Cells. Oncol. Lett. 2019, 18, 529–534. [Google Scholar] [CrossRef]
- Zhang, W.; Shi, H.; Chen, C.; Ren, K.; Xu, Y.; Liu, X.; He, L. CUR Enhances Cisplatin Sensitivity of Human NSCLC Cell Lines through Influencing Cu-Sp1-CTR1 Regulatory Loop. Phytomedicine 2018, 48, 51–61. [Google Scholar] [CrossRef]
- He, Y.Z.; Yu, S.L.; Li, X.N.; Bai, X.H.; Li, H.T.; Liu, Y.C.; Lv, B.L.; Zhao, X.M.; Wei, D.; Zhang, H.L.; et al. CUR increases crizotinib sensitivity through the inactivation of autophagy via epigenetic modulation of the miR-142-5p/Ulk1 axis in non-small cell lung cancer. Cancer Biomark. 2022, 34, 297–307. [Google Scholar] [CrossRef]
- Chen, P.; Huang, H.-P.; Wang, Y.; Jin, J.; Long, W.-G.; Chen, K.; Zhao, X.-H.; Chen, C.-G.; Li, J. CUR Overcome Primary Gefitinib Resistance in Non-Small-Cell Lung Cancer Cells through Inducing Autophagy-Related Cell Death. J. Exp. Clin. Cancer Res. 2019, 38, 254. [Google Scholar] [CrossRef]
- Dong, Z.; Feng, Q.; Zhang, H.; Liu, Q.; Gong, J. CUR enhances drug sensitivity of gemcitabine-resistant lung cancer cells and inhibits metastasis. Die Pharm. 2021, 76, 538–543. [Google Scholar] [CrossRef]
- Lee, W.-H.; Loo, C.-Y.; Traini, D.; Young, P.M. Development and Evaluation of Paclitaxel and CUR Dry Powder for Inhalation Lung Cancer Treatment. Pharmaceutics 2021, 13, 9. [Google Scholar] [CrossRef]
- Fan, W.; Wang, F.; Jin, Z.; Zhu, L.; Zhang, J. CUR Synergizes with Cisplatin to Inhibit Colon Cancer through Targeting the MicroRNA-137-Glutaminase Axis. Curr. Med. Sci. 2022, 42, 108–117. [Google Scholar] [CrossRef]
- Li, G.; Fang, S.; Shao, X.; Li, Y.; Tong, Q.; Kong, B.; Chen, L.; Wang, Y.; Yang, J.; Yu, H.; et al. CUR Reverses NNMT-Induced 5-Fluorouracil Resistance via Increasing ROS and Cell Cycle Arrest in Colorectal Cancer Cells. Biomolecules 2021, 11, 1295. [Google Scholar] [CrossRef]
- Zheng, X.; Yang, X.; Lin, J.; Song, F.; Shao, Y. Low CUR Concentration Enhances the Anticancer Effect of 5-Fluorouracil against Colorectal Cancer. Phytomedicine 2021, 85, 153547. [Google Scholar] [CrossRef]
- Lue, Y.; Zhangd, R.; Zhangd, X.; Zhanga, B.; Yaoa, Q. CUR May Reverse 5-Fluorouracil Resistance on Colonic Cancer Cells by Regulating TET1-NKD-Wnt Signal Pathway to Inhibit the EMT Progress. Biomed. Phar. 2020, 129, 110381. [Google Scholar] [CrossRef]
- He, W.; Zhu, Y.; Zhang, T.; Abulimiti, P.; Zeng, F.; Zhang, L.; Luo, L.; Xie, X.; Zhang, H. CUR Reverses 5-Fluorouracil Resistance by Promoting Human Colon Cancer HCT-8/5-FU Cell Apoptosis and Down-Regulating Heat Shock Protein 27 and P-Glycoprotein. Chin. J. Integr. Med. 2019, 25, 416–424. [Google Scholar] [CrossRef]
- Zhu, C.; Fang, Z.; Peng, L.; Gao, F.; Peng, W.; Song, F. CUR Suppresses the Progression of Colorectal Cancer by Improving Immunogenic Cell Death Caused by Irinotecan. Chemotherapy 2022, 67, 211–222. [Google Scholar] [CrossRef]
- Ouyang, M.; Luo, Z.; Zhang, W.; Zhu, D.; Lu, Y.; Wu, J.; Yao, X. Protective Effect of CUR against Irinotecan-induced Intestinal Mucosal Injury via Attenuation of NF-κB Activation, Oxidative Stress and Endoplasmic Reticulum Stress. Int. J. Oncol. 2019, 54, 1376–1386. [Google Scholar] [CrossRef] [PubMed]
- Su, P.; Yang, Y.; Wang, G.; Chen, X.; Ju, Y. CUR Attenuates Resistance to Irinotecan via Induction of Apoptosis of Cancer Stem Cells in Chemoresistant Colon Cancer Cells. Internat. J. Oncol. 2018, 53, 1343–1353. [Google Scholar] [CrossRef]
- Zhang, L.; Tao, X.; Fu, Q.; Ge, C.; Li, R.; Li, Z.; Zhu, Y.; Tian, H.; Li, Q.; Liu, M.; et al. CUR Inhibits Cell Proliferation and Migration in NSCLC through a Synergistic Effect on the TLR4/MyD88 and EGFR Pathways. Oncol. Rep. 2019, 42, 1843–1855. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Wang, L.; Wang, Y.; Shen, H.; Wang, X.; Wu, L. CUR Reverses Oxaliplatin Resistance in Human Colorectal Cancer via Regulation of TGF-β/Smad2/3 Signaling Pathway. OncoTargets Ther. 2019, 12, 3893–3903. [Google Scholar] [CrossRef]
- Wu, C.-S.; Wu, S.-Y.; Chen, H.-C.; Chu, C.-A.; Tang, H.-H.; Liu, H.-S.; Hong, Y.-R.; Huang, C.-Y.F.; Huang, G.-C.; Su, C.-L. CUR Functions as a MEK Inhibitor to Induce a Synthetic Lethal Effect on KRAS Mutant Colorectal Cancer Cells Receiving Targeted Drug Regorafenib. J. Nutr. Biochem. 2019, 74, 108227. [Google Scholar] [CrossRef]
- Becit, M.; Dilsiz, S.A.; Basaran, N. Interaction of CUR on Cisplatin Cytotoxicity in HeLa and HepG2 Carcinoma Cells. Istanbul J. Pharm. 2020, 50, 202–210. [Google Scholar] [CrossRef]
- Xu, T.; Guo, P.; Pi, C.; He, Y.; Yang, H.; Hou, Y.; Feng, X.; Jiang, Q.; Wei, Y.; Zhao, L. Synergistic Effects of CUR and 5-Fluorouracil on the Hepatocellular Carcinoma In Vivo and Vitro through Regulating the Expression of COX-2 and NF-ΚB. J. Cancer 2020, 11, 3955–3964. [Google Scholar] [CrossRef]
- Tian, N.; Shangguan, W.; Zhou, Z.; Yao, Y.; Fan, C.; Cai, L. Lin28b Is Involved in CUR-Reversed Paclitaxel Chemoresistance and Associated with Poor Prognosis in Hepatocellular Carcinoma. J. Cancer 2019, 10, 6074–6087. [Google Scholar] [CrossRef]
- Bahman, A.; Abaza, M.; Khoushiash, S.; AlAttiyah, R. Sequencedependent Effect of Sorafenib in Combination with Natural Phenolic Compounds on Hepatic Cancer Cells and the Possible Mechanism of Action. Int. J. Mol. Med. 2018, 42, 1695–1715. [Google Scholar] [CrossRef]
- Jadid, M.F.S.; Jahangirzadehd, G.; Behroozi, J. Anti-Proliferation Effects of Apatinib in Combination with CUR in Breast Cancer Cells. Horm. Mol. Biol. Clin. Investig. 2022, 44, 27–32. [Google Scholar] [CrossRef]
- Wang, G.; Duan, P.; Wei, Z.; Liu, F. CUR Sensitizes Carboplatin Treatment in Triple Negative Breast Cancer through Reactive Oxygen Species Induced DNA Repair Pathway. Mol. Biol. Rep. 2022, 49, 3259–3270. [Google Scholar] [CrossRef]
- Biswas, S.; Mahapatra, E.; Ghosh, A.; Das, S.; Roy, M.; Mukherjee, S. CUR Rescues Doxorubicin Responsiveness via Regulating Aurora a Signaling Network in Breast Cancer Cells. Asian Pac. J. Cancer Prev. 2021, 22, 957–970. [Google Scholar] [CrossRef]
- Wen, C.; Fu, L.; Huang, J.; Dai, Y.; Wang, B.; Xu, G.; Wu, L.; Zhou, H. CUR Reverses Doxorubicin Resistance via Inhibition the Efflux Function of ABCB4 in Doxorubicin-Resistant Breast Cancer Cells. Mol. Med. Rep. 2019, 19, 5162–5168. [Google Scholar] [CrossRef]
- Saxena, A.R.; Ilic, Z.; Sripada, V.; Crawford, D.R. Lower Concentrations of CUR Inhibit Her2-Akt Pathway Components in Human Breast Cancer Cells, and Other Dietary Botanicals Potentiate This and Lapatinib Inhibition. Nutr. Res. 2020, 78, 93–104. [Google Scholar] [CrossRef]
- Attia, Y.M.; El-Kersh, D.M.; Ammar, R.A.; Adel, A.; Khalil, A.; Walid, H.; Eskander, K.; Hamdy, M.; Reda, N.; Mohsen, N.E.; et al. Inhibition of Aldehyde Dehydrogenase-1 and p-Glycoprotein-Mediated Multidrug Resistance by CUR and Vitamin D3 Increases Sensitivity to Paclitaxel in Breast Cancer. Chem. Biol. Interact. 2020, 315, 108865. [Google Scholar] [CrossRef]
- Calaf, G.M.; Ponce-Cusi, R.; Carrión, F. CUR and Paclitaxel Induce Cell Death in Breast Cancer Cell Lines. Oncol. Reports 2018, 40, 2381–2388. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. A Open-Label Prospective Cohort Trial of CUR Plus Tyrosine Kinase Inhibitors (TKI) for EGFR-Mutant Advanced NSCLC (CUR). Available online: https://clinicaltrials.gov/ct2/show/NCT02321293?id=NCT02321293&draw=2&rank=1 (accessed on 27 January 2023).
- Jeon, Y.; Sym, S.J.; Yoo, B.K.; Baek, J.-H. Long-Term Survival, Tolerability, and Safety of First-Line Bevacizumab and FOLFIRI in Combination with Ginsenoside-Modified Nanostructured Lipid Carrier Containing CUR in Patients with Unresectable Metastatic Colorectal Cancer. Integr. Cancer Ther. 2022, 21, 153473542211054. [Google Scholar] [CrossRef]
- Gbolahan, O.B.; O’Neil, B.H.; McRee, A.J.; Sanoff, H.K.; Fallon, J.K.; Smith, P.C.; Ivanova, A.; Moore, D.T.; Dumond, J.; Asher, G.N. A Phase I Evaluation of the Effect of CUR on Dose-limiting Toxicity and Pharmacokinetics of Irinotecan in Participants with Solid Tumors. Clin. Transl. Sci. 2022, 15, 1304–1315. [Google Scholar] [CrossRef]
- Howells, L.M.; Iwuji, C.O.O.; Irving, G.R.B.; Barber, S.; Walter, H.; Sidat, Z.; Griffin-Teall, N.; Singh, R.; Foreman, N.; Patel, S.R.; et al. CUR Combined with FOLFOX Chemotherapy Is Safe and Tolerable in Patients with Metastatic Colorectal Cancer in a Randomized Phase IIa Trial. J. Nutr. 2019, 149, 1133–1139. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Avastin/FOLFIRI in Combination with CUR in Colorectal Cancer Patients with Unresectable Metastasis. Available online: https://clinicaltrials.gov/ct2/show/study/NCT02439385 (accessed on 27 January 2023).
- ClinicalTrials.gov. Effect of CUR on Dose Limiting Toxicity and Pharmacokinetics of Irinotecan in Patients with Solid Tumors. Available online: https://clinicaltrials.gov/ct2/show/NCT01859858 (accessed on 27 January 2023).
- ClinicalTrials.gov. Combining CUR with FOLFOX Chemotherapy in Patients with Inoperable Colorectal Cancer (CUFOX). Available online: https://clinicaltrials.gov/ct2/show/NCT01490996?id=NCT01490996&draw=2&rank=1 (accessed on 27 January 2023).
- ClinicalTrials.gov. CUR in Combination with 5FU for Colon Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT02724202 (accessed on 27 January 2023).
- Saghatelyan, T.; Tananyan, A.; Janoyan, N.; Tadevosyan, A.; Petrosyan, H.; Hovhannisyan, A.; Hayrapetyan, L.; Arustamyan, M.; Arnhold, J.; Rotmann, A.-R.; et al. Efficacy and Safety of CUR in Combination with Paclitaxel in Patients with Advanced, Metastatic Breast Cancer: A Comparative, Randomized, Double-Blind, Placebo-Controlled Clinical Trial. Phytomedicine 2020, 70, 153218. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. CUR in Combination with Chemotherapy in Advanced Breast Cancer. Available online: https://clinicaltrials.gov/ct2/show/NCT03072992?id=NCT03072992&draw=2&rank=1 (accessed on 27 January 2023).
- ClinicalTrials.gov. Phase II Study of CUR vs Placebo for Chemotherapy-Treated Breast Cancer Patients Undergoing Radiotherapy. Available online: https://clinicaltrials.gov/ct2/show/record/NCT01740323?view=record (accessed on 27 January 2023).
- Tan, B.L.; Norhaizan, M.E. CUR Combination Chemotherapy: The Implication and Efficacy in Cancer. Molecules 2019, 24, 2527. [Google Scholar] [CrossRef] [PubMed]
- Haritha, N.H.; Nawab, A.; Vijayakurup, V.; Anto, N.P.; Liju, V.B.; Alex, V.V.; Amrutha, A.N.; Aiswarya, S.U.; Swetha, M.; Vinod, B.S.; et al. Targeting Thymidylate Synthase Enhances the Chemosensitivity of Triple-Negative Breast Cancer Towards 5-FU-Based Combinatorial Therapy. Front. Oncol. 2021, 11, 656804. [Google Scholar] [CrossRef] [PubMed]
- Najafi, M.; Tavakol, S.; Zarrabi, A.; Ashrafizadeh, M. Dual Role of Quercetin in Enhancing the Efficacy of Cisplatin in Chemotherapy and Protection against Its Side Effects: A Review. Arch. Physiol. Biochem. 2022, 128, 1438–1452. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Khan, H.; Ullah, H.; Hassan, S.T.S.; Šmejkal, K.; Efferth, T.; Mahomoodally, M.F.; Xu, S.; Habtemariam, S.; Filosa, R.; et al. MicroRNA Targeting by Quercetin in Cancer Treatment and Chemoprotection. Pharmacol. Res. 2019, 147, 104346. [Google Scholar] [CrossRef] [PubMed]
- Ghafouri-Fard, S.; Shoorei, H.; Khanbabapour Sasi, A.; Taheri, M.; Ayatollahi, S.A. The Impact of the Phytotherapeutic Agent Quercetin on Expression of Genes and Activity of Signaling Pathways. Biomed. Pharmacother. 2021, 141, 111847. [Google Scholar] [CrossRef]
- Varghese, E.; Samuel, S.M.; Abotaleb, M.; Cheema, S.; Mamtani, R.; Buesselberg, D. The “Yin and Yang” of Natural Compounds in Anticancer Therapy of Triple-Negative Breast Cancers. Cancers 2018, 10, 346. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, K.; Yan, C.; Yin, Y.; He, S.; Qiu, L.; Li, G. Natural Polyphenols for Treatment of Colorectal Cancer. Molecules 2022, 27, 8810. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhang, J.; Wang, K.; Han, W.; Wang, X.; Gao, M.; Wang, Z.; Sun, Y.; Yan, H.; Zhang, H.; et al. Quercetin Overcomes Colon Cancer Cells Resistance to Chemotherapy by Inhibiting Solute Carrier Family 1, Member 5 Transporter. Eur. J. Pharmacol. 2020, 881, 173185. [Google Scholar] [CrossRef]
- Erdoğan, M.K.; Ağca, C.A.; Aşkın, H. Quercetin and Luteolin Improve the Anticancer Effects of 5-Fluorouracil in Human Colorectal Adenocarcinoma In Vitro Model: A Mechanistic Insight. Nutr. Cancer 2022, 74, 660–676. [Google Scholar] [CrossRef]
- Terana, G.; Abd-Alhaseeb, M.; Omran, G.; Okda, T.M. Quercetin Potentiates 5-Fluorouracil Effects in Human Colon Cancer Cells through Targeting the Wnt/β-Catenin Signalling Pathway: The Role of MiR-27a. Contemp. Oncol. 2022, 26, 229–238. [Google Scholar] [CrossRef]
- Chen, Z.; Huang, C.; Ma, T.; Jiang, L.; Tang, L.; Shi, T.; Zhang, S.; Zhang, L.; Zhu, P.; Li, J.; et al. Reversal Effect of Quercetin on Multidrug Resistance via FZD7/β-Catenin Pathway in Hepatocellular Carcinoma Cells. Phytomedicine 2018, 43, 37–45. [Google Scholar] [CrossRef]
- Hassan, S.; Peluso, J.; Chalhoub, S.; Idoux Gillet, Y.; Benkirane-Jessel, N.; Rochel, N.; Fuhrmann, G.; Ubeaud-Sequier, G. Quercetin Potentializes the Respective Cytotoxic Activity of Gemcitabine or Doxorubicin on 3D Culture of AsPC-1 or HepG2 Cells, through the Inhibition of HIF-1α and MDR1. PLoS ONE 2020, 15, e0240676. [Google Scholar] [CrossRef]
- Liu, Z.; Xu, W.; Han, J.; Liu, Q.; Gao, L.; Wang, X.; Li, X. Quercetin Induces Apoptosis and Enhances Gemcitabine Therapeutic Efficacy against Gemcitabine-Resistant Cancer Cells. Anti-Cancer Drug 2020, 31, 684–692. [Google Scholar] [CrossRef]
- Abdu, S.; Juaid, N.; Amin, A.; Moulay, M.; Miled, N. Effects of Sorafenib and Quercetin Alone or in Combination in Treating Hepatocellular Carcinoma: In Vitro and In Vivo Approaches. Molecules 2022, 27, 8082. [Google Scholar] [CrossRef]
- Chen, M.; Duan, C.; Pan, L. Quercetin Increases Doxorubicin-Induced Apoptosis Through Oxidative DNA Damage in KATO III Gastric Cancer Cells. Iran. Red. Crescent. Med. J. 2021, 23. [Google Scholar] [CrossRef]
- Hyun, H.B.; Moon, J.Y.; Cho, S.K. Quercetin Suppresses CYR61-Mediated Multidrug Resistance in Human Gastric Adenocarcinoma AGS Cells. Molecules 2018, 23, 209. [Google Scholar] [CrossRef]
- Lei, C.-S.; Hou, Y.-C.; Pai, M.-H.; Lin, M.-T.; Yeh, S.-L. Effects of Quercetin Combined with Anticancer Drugs on Metastasis-Associated Factors of Gastric Cancer Cells: In Vitro and In Vivo Studies. J. Nutr. Biochem. 2018, 51, 105–113. [Google Scholar] [CrossRef]
- Liu, H.; Lee, J.I.; Ahn, T.-G. Effect of Quercetin on the Anti-Tumor Activity of Cisplatin in EMT6 Breast Tumor-Bearing Mice. Obstet. Gynecol. Sci. 2019, 62, 242. [Google Scholar] [CrossRef]
- Safi, A.; Heidarian, E.; Ahmadi, R. Quercetin Synergistically Enhances the Anticancer Efficacy of Docetaxel through Induction of Apoptosis and Modulation of PI3K/AKT, MAPK/ERK, and JAK/STAT3 Signaling Pathways in MDA-MB-231 Breast Cancer Cell Line. Int. J. Mol. Cell. Med. 2021, 10, 11–22. [Google Scholar] [CrossRef]
- Li, S.; Yuan, S.; Zhao, Q.; Wang, B.; Wang, X.; Li, K. Quercetin Enhances Chemotherapeutic Effect of Doxorubicin against Human Breast Cancer Cells While Reducing Toxic Side Effects of It. Biomed. Pharmacother. 2018, 100, 441–447. [Google Scholar] [CrossRef]
- Zhang, P.; Zhang, J.; Zhao, L.; Li, S.; Li, K. Quercetin Attenuates the Cardiotoxicity of Doxorubicin–Cyclophosphamide Regimen and Potentiates Its Chemotherapeutic Effect against Triple-negative Breast Cancer. Phytother. Res. 2022, 36, 551–561. [Google Scholar] [CrossRef] [PubMed]
- Mawalizadeh, F.; Mohammadzadeh, G.; Khedri, A.; Rashidi, M. Quercetin Potentiates the Chemosensitivity of MCF-7 Breast Cancer Cells to 5-Fluorouracil. Mol. Biol. Rep. 2021, 48, 7733–7742. [Google Scholar] [CrossRef] [PubMed]
- Roshanazadeh, M.; Rezaei, H.B.; Rashidi, M. Quercetin Synergistically Potentiates the Anti-Metastatic Effect of 5-Fluorouracil on the MDA-MB-231 Breast Cancer Cell Line. Iran. J. Basic Med. Sci. 2021, 24, 928–934. [Google Scholar] [CrossRef] [PubMed]
- Ozkan, E.; Bakar-Ates, F. Potentiation of the Effect of Lonidamine by Quercetin in MCF-7 Human Breast Cancer Cells through Downregulation of MMP-2/9 MRNA Expression. An. Acad. Bras. Ciênc. 2020, 92, e20200548. [Google Scholar] [CrossRef]
- Zhang, L.-X.; Li, C.-X.; Kakar, M.U.; Khan, M.S.; Wu, P.-F.; Amir, R.M.; Dai, D.-F.; Naveed, M.; Li, Q.-Y.; Saeed, M.; et al. Resveratrol (RV): A Pharmacological Review and Call for Further Research. Biomed. Pharmacother. 2021, 143, 112164. [Google Scholar] [CrossRef]
- Riccio, B.V.F.; Spósito, L.; Carvalho, G.C.; Ferrari, P.C.; Chorilli, M. Resveratrol Isoforms and Conjugates: A Review from Biosynthesis in Plants to Elimination from the Human Body. Arch. Pharm. 2020, 353, 2000146. [Google Scholar] [CrossRef]
- Ren, B.; Kwah, M.X.-Y.; Liu, C.; Ma, Z.; Shanmugam, M.K.; Ding, L.; Xiang, X.; Ho, P.C.-L.; Wang, L.; Ong, P.S.; et al. Resveratrol for Cancer Therapy: Challenges and Future Perspectives. Cancer Lett. 2021, 515, 63–72. [Google Scholar] [CrossRef]
- Roshani, M.; Jafari, A.; Loghman, A.; Sheida, A.H.; Taghavi, T.; Tamehri Zadeh, S.S.; Hamblin, M.R.; Homayounfal, M.; Mirzaei, H. Applications of Resveratrol in the Treatment of Gastrointestinal Cancer. Biomed. Pharmacother. 2022, 153, 113274. [Google Scholar] [CrossRef]
- Ren, M.; Zhou, X.; Gu, M.; Jiao, W.; Yu, M.; Wang, Y.; Liu, S.; Yang, J.; Ji, F. Resveratrol Synergizes with Cisplatin in Antineoplastic Effects against AGS Gastric Cancer Cells by Inducing Endoplasmic Reticulum Stressmediated Apoptosis and G2/M Phase Arrest. Oncol. Rep. 2020, 44, 1605–1615. [Google Scholar] [CrossRef]
- Ghafouri-Fard, S.; Bahroudi, Z.; Shoorei, H.; Hussen, B.M.; Talebi, S.F.; Baig, S.G.; Taheri, M.; Ayatollahi, S.A. Disease-Associated Regulation of Gene Expression by Resveratrol: Special Focus on the PI3K/AKT Signaling Pathway. Cancer Cell Int. 2022, 22, 298. [Google Scholar] [CrossRef]
- Fu, X.; Li, M.; Tang, C.; Huang, Z.; Najafi, M. Targeting of Cancer Cell Death Mechanisms by Resveratrol: A Review. Apoptosis 2021, 26, 561–573. [Google Scholar] [CrossRef]
- Chen, L.; Musa, A.E. Boosting Immune System against Cancer by Resveratrol. Phytother. Res. 2021, 35, 5514–5526. [Google Scholar] [CrossRef]
- Choi, C.Y.; Lim, S.-C.; Lee, T.-B.; Han, S.I. Molecular Basis of Resveratrol-Induced Resensitization of Acquired Drug-Resistant Cancer Cells. Nutrients 2022, 14, 699. [Google Scholar] [CrossRef]
- Cocetta, V.; Quagliariello, V.; Fiorica, F.; Berretta, M.; Montopoli, M. Resveratrol as Chemosensitizer Agent: State of Art and Future Perspectives. Int. J. Mol. Sci. 2021, 22, 2049. [Google Scholar] [CrossRef]
- Qin, S.-H.; Lau, A.T.Y.; Liang, Z.-L.; Tan, H.W.; Ji, Y.-C.; Zhong, Q.-H.; Zhao, X.-Y.; Xu, Y.-M. Resveratrol Promotes Tumor Microvessel Growth via Endoglin and Extracellular Signal-Regulated Kinase Signaling Pathway and Enhances the Anticancer Efficacy of Gemcitabine against Lung Cancer. Cancers 2020, 12, 974. [Google Scholar] [CrossRef]
- Liu, Z.; Peng, Q.; Li, Y.; Gao, Y. Resveratrol Enhances Cisplatin-Induced Apoptosis in Human Hepatoma Cells via Glutamine Metabolism Inhibition. BMB Rep. 2018, 51, 474–479. [Google Scholar] [CrossRef]
- Gao, M.; Deng, C.; Dang, F. Synergistic Antitumor Effect of Resveratrol and Sorafenib on Hepatocellular Carcinoma through PKA/AMPK/EEF2K Pathway. Food Nutr. Res. 2021, 65, 3602. [Google Scholar] [CrossRef]
- Yang, M.-D.; Sun, Y.; Zhou, W.-J.; Xie, X.-Z.; Zhou, Q.-M.; Lu, Y.-Y.; Su, S.-B. Resveratrol Enhances Inhibition Effects of Cisplatin on Cell Migration and Invasion and Tumor Growth in Breast Cancer MDA-MB-231 Cell Models In Vivo and In Vitro. Molecules 2021, 26, 2204. [Google Scholar] [CrossRef]
- Ozdemir, F.; Sever, A.; Öğünç Keçeci, Y.; Incesu, Z. Resveratrol Increases the Sensitivity of Breast Cancer MDA-MB-231 Cell Line to Cisplatin by Regulating Intrinsic Apoptosis. Iran J. Basic Med. Sci. 2021, 24, 66–72. [Google Scholar] [CrossRef]
- LeonGalicia, I.; DiazChavez, J.; AlbinoSanchez, M.E.; GarciaVilla, E.; BermudezCruz, R.; GarciaMena, J.; Herrera, L.A.; GarcíaCarrancá, A.; Gariglio, P. Resveratrol Decreases Rad51 Expression and Sensitizes Cisplatinresistant MCF7 Breast Cancer Cells. Oncol. Rep. 2018, 39, 3025–3033. [Google Scholar] [CrossRef]
- Vargas, J.E.; Puga, R.; Lenz, G.; Trindade, C.; Filippi-Chiela, E. Cellular Mechanisms Triggered by the Cotreatment of Resveratrol and Doxorubicin in Breast Cancer: A Translational In Vitro–In Silico Model. Oxid. Med. Cell. Longev. 2020, 2020, 5432651. [Google Scholar] [CrossRef] [PubMed]
- Jin, X.; Wei, Y.; Liu, Y.; Lu, X.; Ding, F.; Wang, J.; Yang, S. Resveratrol Promotes Sensitization to Doxorubicin by Inhibiting Epithelial-Mesenchymal Transition and Modulating SIRT1/βcatenin Signaling Pathway in Breast Cancer. Cancer Med. 2019, 8, 1246–1257. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.-M.; Bai, J.-Y.; Yang, K.-X. Effect of Resveratrol on Doxorubicin Resistance in Breast Neoplasm Cells by Modulating PI3K/Akt Signaling Pathway. IUBMB Life 2018, 70, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Hallajian, F.; Ghasmi, M.; Abedi, S.M.; Behzadi, R.; Hayati, E.; Sadeghzadeh, N.; Rezazadeh, F.; Karimi, H. Evaluation of the Effect of Resveratrol and Doxorubicin on 99m Tc-MIBI Uptake in Breast Cancer Cell Xenografts in Mice. Cancer Biother. Radio. 2018, 33, 403–410. [Google Scholar] [CrossRef]
- Cipolletti, M.; Montalesi, E.; Nuzzo, M.T.; Fiocchetti, M.; Ascenzi, P.; Marino, M. Potentiation of Paclitaxel Effect by Resveratrol in Human Breast Cancer Cells by Counteracting the 17β-estradiol/Estrogen Receptor α/Neuroglobin Pathway. J. Cell. Physiol. 2019, 234, 3147–3157. [Google Scholar] [CrossRef]
- Ostwal, V.; Ramaswamy, A.; Bhargava, P.; Srinivas, S.; Mandavkar, S.; Chaugule, D.; Peelay, Z.; Baheti, A.; Tandel, H.; Jadhav, V.K.; et al. A Pro-Oxidant Combination of Resveratrol and Copper Reduces Chemotherapy-Related Non-Haematological Toxicities in Advanced Gastric Cancer: Results of a Prospective Open Label Phase II Single-Arm Study (RESCU III Study). Med. Oncol. 2023, 40, 17. [Google Scholar] [CrossRef]
- Abdel Latif, Y.; El-Bana, M.; Hussein, J.; El-Khayat, Z.; Farrag, A.R. Effects of Resveratrol in Combination with 5-Fluorouracil on N-Methylnitrosourea-Induced Colon Cancer in Rats. Comp. Clin. Pathol. 2019, 28, 1351–1362. [Google Scholar] [CrossRef]
- Chung, S.S.; Dutta, P.; Austin, D.; Wang, P.; Awad, A.; Vadgama, J.V. Combination of Resveratrol and 5-Flurouracil Enhanced Anti-Telomerase Activity and Apoptosis by Inhibiting STAT3 and Akt Signaling Pathways in Human Colorectal Cancer Cells. Oncotarget 2018, 9, 32943–32957. [Google Scholar] [CrossRef]
- Buhrmann, C.; Yazdi, M.; Popper, B.; Shayan, P.; Goel, A.; Aggarwal, B.; Shakibaei, M. Resveratrol Chemosensitizes TNF-β-Induced Survival of 5-FU-Treated Colorectal Cancer Cells. Nutrients 2018, 10, 888. [Google Scholar] [CrossRef]
- Mokra, D.; Joskova, M.; Mokry, J. Therapeutic Effects of Green Tea Polyphenol (−)-Epigallocatechin-3-Gallate (EGCG) in Relation to Molecular Pathways Controlling Inflammation, Oxidative Stress, and Apoptosis. Int. J. Mol. Sci. 2023, 24, 340. [Google Scholar] [CrossRef]
- Mitra, T.; Bhattacharya, R. Phytochemicals Modulate Cancer Aggressiveness: A Review Depicting the Anticancer Efficacy of Dietary Polyphenols and Their Combinations. J. Cell. Physiol. 2020, 235, 7696–7708. [Google Scholar] [CrossRef]
- Gan, R.-Y.; Li, H.-B.; Sui, Z.-Q.; Corke, H. Absorption, Metabolism, Anti-Cancer Effect and Molecular Targets of Epigallocatechin Gallate (EGCG): An Updated Review. Crit. Rev. Food Sci. Nutr. 2018, 58, 924–941. [Google Scholar] [CrossRef]
- Deng, P.; Hu, C.; Xiong, Z.; Li, Y.; Jiang, J.; Yang, H.; Tang, Y.; Cao, L.; Lu, R. Epigallocatechin-3-gallate-induced vascular normalization in A549-cell xenograft-bearing nude mice: Therapeutic efficacy in combination with chemotherapy. Cancer Manag. Res. 2019, 11, 2425–2439. [Google Scholar] [CrossRef]
- Heyza, J.R.; Arora, S.; Zhang, H.; Conner, K.L.; Lei, W.; Floyd, A.M.; Deshmukh, R.R.; Sarver, J.; Trabbic, C.J.; Erhardt, P.; et al. Targeting the DNA Repair Endonuclease ERCC1-XPF with Green Tea Polyphenol Epigallocatechin-3-Gallate (EGCG) and Its Prodrug to Enhance Cisplatin Efficacy in Human Cancer Cells. Nutrients 2018, 10, 1644. [Google Scholar] [CrossRef]
- Datta, S.; Sinha, D. Low Dose Epigallocatechin-3-Gallate Revives Doxorubicin Responsiveness by a Redox-Sensitive Pathway in A549 Lung Adenocarcinoma Cells. J. Biochem. Mol. Toxicol. 2022, 36, e22999. [Google Scholar] [CrossRef]
- Datta, S.; Sinha, D. EGCG Maintained Nrf2-Mediated Redox Homeostasis and Minimized Etoposide Resistance in Lung Cancer Cells. J. Funct. Foods 2019, 62, 103553. [Google Scholar] [CrossRef]
- Polonio-Alcalá, E.; Palomeras, S.; Torres-Oteros, D.; Relat, J.; Planas, M.; Feliu, L.; Ciurana, J.; Ruiz-Martínez, S.; Puig, T. Fatty Acid Synthase Inhibitor G28 Shows Anticancer Activity in EGFR Tyrosine Kinase Inhibitor Resistant Lung Adenocarcinoma Models. Cancers 2020, 12, 1283. [Google Scholar] [CrossRef]
- Meng, J.; Chang, C.; Chen, Y.; Bi, F.; Ji, C.; Liu, W. EGCG Overcomes Gefitinib Resistance by Inhibiting Autophagy and Augmenting Cell Death through Targeting ERK Phosphorylation in NSCLC. OncoTargets Ther. 2019, 12, 6033–6043. [Google Scholar] [CrossRef]
- Wei, R.; Wirkus, J.; Yang, Z.; Machuca, J.; Esparza, Y.; Mackenzie, G.G. EGCG sensitizes chemotherapeutic-induced cytotoxicity by targeting the ERK pathway in multiple cancer cell lines. Arch. Biochem. Biophys. 2020, 692, 108546. [Google Scholar] [CrossRef]
- Li, H.; Krstin, S.; Wink, M. Modulation of multidrug resistant in cancer cells by EGCG, tannic acid and curcumin. Phytomedicine 2018, 50, 213–222. [Google Scholar] [CrossRef]
- Moracci, L.; Sensi, F.; Biccari, A.; Crotti, S.; Gaio, E.; Benetti, F.; Traldi, P.; Pucciarelli, S.; Agostini, M. An Investigation on [5 Fluorouracil and Epigallocatechin-3-Gallate] Complex Activity on HT-29 Cell Death and Its Stability in Gastrointestinal Fluid. Oncotarget 2022, 13, 476–489. [Google Scholar] [CrossRef] [PubMed]
- Kucukkaraduman, B.; Cicek, E.G.; Akbar, M.W.; Canli, S.D.; Vural, B.; Gure, A.O. Epithelial-to-Mesenchymal Transition Is Not a Major Modulating Factor in the Cytotoxic Response to Natural Products in Cancer Cell Lines. Molecules 2021, 26, 5858. [Google Scholar] [CrossRef] [PubMed]
- La, X.; Zhang, L.; Li, Z.; Li, H.; Yang, Y. (−)-Epigallocatechin Gallate (EGCG) Enhances the Sensitivity of Colorectal Cancer Cells to 5-FU by Inhibiting GRP78/NF-κB/miR-155-5p/MDR1 Pathway. J. Agric. Food Chem. 2019, 67, 2510–2518. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Gou, H.; Xiang, B.; Geng, R.; Dong, J.; Yang, X.; Chen, D.; Dai, R.; Chen, L.; Liu, J. EGCG Enhances the Chemosensitivity of Colorectal Cancer to Irinotecan through GRP78-Mediated Endoplasmic Reticulum Stress. J. Oncol. 2022, 2022, 7099589. [Google Scholar] [CrossRef]
- Wu, W.; Dong, J.; Gou, H.; Geng, R.; Yang, X.; Chen, D.; Xiang, B.; Zhang, Z.; Ren, S.; Chen, L.; et al. EGCG Synergizes the Therapeutic Effect of Irinotecan through Enhanced DNA Damage in Human Colorectal Cancer Cells. J. Cell Mol. Med. 2021, 25, 7913–7921. [Google Scholar] [CrossRef]
- Neelamegam, U.; Muthuvel, R.; Suganthi, V. Hepatoprotective Effect of Epigallocatechin-Gallate (Egcg) And Sorafenib Against Den Induced Hepato Cellular Carcinoma In Experimental Animals. J. Pharm. Negat. Results 2022, 13, 921–929. [Google Scholar] [CrossRef]
- Xue, M.; Liu, X.; Cheng, B.; Rui, X.; Wu, M.; Lv, J. Epigallocatechin Gallate Enhances Inhibition Effect of DDP on the Proliferation of Gastric Cancer BGC-823 Cells by Regulating p19Arf-p53-p21Cip1 Signaling Pathway. Asian Pac. J. Cancer Prev. 2021, 22, 1263–1270. [Google Scholar] [CrossRef]
- Changizi, V.; Azariasl, S.; Motevaseli, E.; Jafari Nodooshan, S. Assessment Synergistic Effects of Integrated Therapy with Epigallocatechin-3-Gallate (EGCG) & Arsenic Trioxide and Irradiation on Breast Cancer Cell Line. Iran J. Public Health 2020, 49, 1555–1563. [Google Scholar]
- Lubecka, K.; Kaufman-Szymczyk, A.; Cebula-Obrzut, B.; Smolewski, P.; Szemraj, J.; Fabianowska-Majewska, K. Novel Clofarabine-Based Combinations with Polyphenols Epigenetically Reactivate Retinoic Acid Receptor Beta, Inhibit Cell Growth, and Induce Apoptosis of Breast Cancer Cells. Int. J. Mol. Sci. 2018, 19, 3970. [Google Scholar] [CrossRef]
- Steed, K.L.; Jordan, H.R.; Tollefsbol, T.O. SAHA and EGCG Promote Apoptosis in Triple-Negative Breast Cancer Cells, Possibly Through the Modulation of cIAP2. Anticancer Res. 2020, 40, 9–26. [Google Scholar] [CrossRef]
- Lewis, K.A.; Jordan, H.R.; Tollefsbol, T.O. Effects of SAHA and EGCG on Growth Potentiation of Triple-Negative Breast Cancer Cells. Cancers 2019, 11, 23. [Google Scholar] [CrossRef]
- Zhao, H.; Jia, L.; Chen, G.; Li, X.; Meng, X.; Zhao, X.; Xing, L.; Zhu, W. A Prospective, Three-Arm, Randomized Trial of EGCG for Preventing Radiation-Induced Esophagitis in Lung Cancer Patients Receiving Radiotherapy. Radiother. Oncol. 2019, 137, 186–191. [Google Scholar] [CrossRef]
- Zhu, W.; Zhao, Y.; Zhang, S.; Li, X.; Xing, L.; Zhao, H.; Yu, J. Evaluation of Epigallocatechin-3-Gallate as a Radioprotective Agent During Radiotherapy of Lung Cancer Patients: A 5-Year Survival Analysis of a Phase 2 Study. Front. Oncol. 2021, 11, 686950. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Study of Epigallocatechin-3-gallate (EGCG) for Esophagus Protection in Patients with Lung Cancer Receiving Radial Radiotherapy. Available online: https://clinicaltrials.gov/ct2/show/study/NCT02577393 (accessed on 12 March 2023).
- Zhao, H.; Zhu, W.; Zhao, X.; Li, X.; Zhou, Z.; Zheng, M.; Meng, X.; Kong, L.; Zhang, S.; He, D.; et al. Efficacy of Epigallocatechin-3-Gallate in Preventing Dermatitis in Patients with Breast Cancer Receiving Postoperative Radiotherapy: A Double-Blind, Placebo-Controlled, Phase 2 Randomized Clinical Trial. JAMA Dermatol. 2022, 158, 779–786. [Google Scholar] [CrossRef]
- ClinicalTrials.gov. Study of Epigallocatechin-3-gallate (EGCG) for Skin Prevention in Patients with Breast Cancer Receiving Adjuvant Radiotherapy. Available online: https://clinicaltrials.gov/ct2/show/NCT02580279 (accessed on 12 March 2023).
- Nozhat, Z.; Heydarzadeh, S.; Memariani, Z.; Ahmadi, A. Chemoprotective and Chemosensitizing Effects of Apigenin on Cancer Therapy. Cancer Cell Int. 2021, 21, 574. [Google Scholar] [CrossRef]
- Imran, M.; Gondal, T.A.; Atif, M.; Shahbaz, M.; Qaisarani, T.B.; Mughal, M.H.; Salehi, B.; Martorell, M.; Sharifi-Rad, J. Apigenin as an Anticancer Agent. Phytother. Res. 2020, 34, 1812–1828. [Google Scholar] [CrossRef]
- Xu, Y.; Li, X.; Wang, H. Protective Roles of Apigenin Against Cardiometabolic Diseases: A Systematic Review. Front. Nutr. 2022, 9, 875826. [Google Scholar] [CrossRef]
- Rahmani, A.H.; Alsahli, M.A.; Almatroudi, A.; Almogbel, M.A.; Khan, A.A.; Anwar, S.; Almatroodi, S.A. The Potential Role of Apigenin in Cancer Prevention and Treatment. Molecules 2022, 27, 6051. [Google Scholar] [CrossRef]
- Li, Y.; Chen, X.; He, W.; Xia, S.; Jiang, X.; Li, X.; Bai, J.; Li, N.; Chen, L.; Yang, B. Apigenin Enhanced Antitumor Effect of Cisplatin in Lung Cancer via Inhibition of Cancer Stem Cells. Nutr. Cancer 2021, 73, 1489–1497. [Google Scholar] [CrossRef]
- Yan, W.; Wu, T.H.Y.; Leung, S.S.Y.; To, K.K.W. Flavonoids Potentiated Anticancer Activity of Cisplatin in Non-Small Cell Lung Cancer Cells In Vitro by Inhibiting Histone Deacetylases. Life Sci. 2020, 258, 118211. [Google Scholar] [CrossRef]
- Zhan, Y.; Wang, Y.; Qi, M.; Liang, P.; Ma, Y.; Li, T.; Li, H.; Dai, C.; An, Z.; Qi, Y.; et al. BH3 Mimetic ABT-263 Enhances the Anticancer Effects of Apigenin in Tumor Cells with Activating EGFR Mutation. Cell Biosci. 2019, 9, 60. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Song, J.; Hwang, S.; Choi, J.; Song, G.; Lim, W. Apigenin Enhances Apoptosis Induction by 5-Fluorouracil through Regulation of Thymidylate Synthase in Colorectal Cancer Cells. Redox Biol. 2021, 47, 102144. [Google Scholar] [CrossRef] [PubMed]
- Papachristou, F.; Anninou, N.; Koukoulis, G.; Paraskakis, S.; Sertaridou, E.; Tsalikidis, C.; Pitiakoudis, M.; Simopoulos, C.; Tsaroucha, A. Differential Effects of Cisplatin Combined with the Flavonoid Apigenin on HepG2, Hep3B, and Huh7 Liver Cancer Cell Lines. Mutat. Res. Genet. Toxicol. Environ. Mutagen. 2021, 866, 503352. [Google Scholar] [CrossRef] [PubMed]
- Korga, A.; Ostrowska, M.; Jozefczyk, A.; Iwan, M.; Wojcik, R.; Zgorka, G.; Herbet, M.; Vilarrubla, G.G.; Dudka, J. Apigenin and Hesperidin Augment the Toxic Effect of Doxorubicin against HepG2 Cells. BMC Pharmacol. Toxicol. 2019, 20, 22. [Google Scholar] [CrossRef]
- Li, K.; Li, M.; Luo, Z.; Mao, Y.; Yu, Y.; He, Y.; Zhou, J.; Fei, Y.; Pei, Y.; Cai, K. Overcoming the Hypoxia-Induced Drug Resistance in Liver Tumor by the Concurrent Use of Apigenin and Paclitaxel. Biochem. Biophys. Res. Commun. 2020, 526, 321–327. [Google Scholar] [CrossRef]
- Aziz, N.A.; Froemming, G.R.A.; Kadir, S.H.S.A.; Ibahim, M.J. Apigenin Increases Cisplatin Inhibitory Effects on the Telomerase Activity of Triple Negative Breast Cancer Cells. Jurnal Teknologi 2018, 80, 123–132. [Google Scholar] [CrossRef]
- Korga-Plewko, A.; Michalczyk, M.; Adamczuk, G.; Humeniuk, E.; Ostrowska-Lesko, M.; Jozefczyk, A.; Iwan, M.; Wojcik, M.; Dudka, J. Apigenin and Hesperidin Downregulate DNA Repair Genes in MCF-7 Breast Cancer Cells and Augment Doxorubicin Toxicity. Molecules 2020, 25, 4421. [Google Scholar] [CrossRef]


| Cancer Type | Chemotherapy | Dosage | Assay Type | Molecular Effect | Study Conclusion | Ref. |
|---|---|---|---|---|---|---|
| Lung cancer | CIS | 41 µM CUR + 30 µM CIS for A549 cells; 33 µM CUR + 7 µM CIS for H2170 cells | A549 and H2170 cell lines | Suppression of the self-renewal capability of cancer stem cells. | Synergistic inhibition of NSCLC. | [83] |
| In vitro: 2–32 µM CUR + 0.5–8 µg/mL CIS. In vivo: 50 mg/kg CUR + 2.5 mg/kg CIS | A549, H1299, NCI-H460 cell lines and BALB/c mice | Upregulating the levels of CTR1 and Sp1 to increase more Pt2+ uptake. | Enhancing sensitivity and antitumor effects of CIS in NSCLC. | [85] | ||
| CIS + X-ray | 10 µmol/L CUR + 1 mg/L CIS | A549 cell line | Inhibition of EGFR-related signaling pathway. | Inhibition of cancer cell migration and invasiveness. Augmenting radio-sensitization effects against NSCLC. | [84] | |
| CZT | 30 µM CUR + 20 µM CZT | A549, H460, H1299 and H1066 cell lines | Increasing the levels of miR-142-5p through epigenetic and suppressing autophagy. | Enhancing NSCLC sensitivity to CZT treatment. | [86] | |
| GEF | In vitro: 5–10 µM CUR + 0–20 µM GEF. In vivo: 1 g/kg CUR + 100 mg/kg GEF | H157, H1299, PC-9 cell lines and BALBL/c mice | Inhibition of Sp1/EGFR activity to induce autophagy mediated apoptosis. | Reduction in tumor volume. Elevating the sensitivity to GEF in NSCLC patients with mutated EGFR. | [87] | |
| GEM | 3 µM CUR + 58.2 µM GEM for A549 cells; 3 µM CUR + 98.72 µM GEM for A549/GEM cells | A549 and A549/GEM drug-resistant cell lines | Downregulating expression of MMP9, vimentin, and N-cadherin and upregulating E-cadherin to slow EMT. | Elevating sensitivity of GEM-resistant NSCLC and decreasing migration and invasion. | [88] | |
| PTX | 75, 25, 50% (w/w) CUR + 25, 50, 75% (w/w) PTX | Calu-3 and A549 cell lines | Induction of apoptosis/necrotic cell death and G2/M cell cycle arrests. Increased intracellular ROS, mitochondrial depolarization, and reduced ATP content. | The combination exerts a more potent cytotoxic effect. CUR neutralizes cytotoxic effects of PTX. | [89] | |
| Colorectal cancer | CIS | 10, 20 µM CUR + 0.4, 8 µM CIS | HT-29/CIS drug-resistant cell line | Inhibition glutamine through miR-137-mediated. | CUR treatment overcame CIS resistance and suppressed proliferation of CRC. | [90] |
| 5-FU | 30 μM CUR + 20 mg/L 5-FU for HT-29 cells; 10 μM CUR + 10 mg/L 5-FU for SW480 cells | HT-29 and SW480 cell lines | G2/M Phase cell cycle arrest and downregulation of NNMT by p-STAT3 depression. | The combination inhibits CRC proliferation. | [91] | |
| In vitro and in vivo: 10 μM CUR + 5 μM 5-FU | SW620 cell line and female nude mice | Inhibited pERK signaling and downregulated L1 expression in SW620 cells. | Significantly increased apoptosis rate extended the survival of immunodeficient mice in the combination group as compared to that of the 5-FU group. | [92] | ||
| 5–40 μM CUR + 1.39 μg/mL 5-FU | rHCT-116/5-FU drug-resistant cell line | Regulating the TET1-NKD2-WNT signal pathway to inhibit the EMT progress. | CUR might exert an anti-resistant effect to 5-FU in HCT-116 cells. | [93] | ||
| 2–25 μg/mL CUR + 0.05–1000 μg/mL 5-FU | HCT-8 and HCT-8/5-FU drug-resistant cell lines | Downregulation of P-gp and HSP-27. | Inhibition of tumor growth. Reversal effects on MDR. | [94] | ||
| IRI | In vitro: 2–14 μg/mL CUR + 2–14 ng/mL IRI. In vivo: 5 mg/kg CUR every other day 3 times + IRI 25 mg/kg every other day 3 times | CT-26 cell line and C57 BL/6j mice | Upregulated ICD-related proteins including CALR and HMGB1a. | CUR may synergistically improve the antitumor effect of IRI by promoting the immunogenic cell death (ICD) effect. | [95] | |
| 100 mg/kg CUR by intragastric administration for 8 days + 75 mg/kg IRI for 4 days | IRI-treated BALB/c nude mice | Downregulation of NF-κB. | Protective effect against IRI-induced intestinal mucosal injury. | [96] | ||
| 2.5–20 µM CUR + 10–100 µM IRI | LoVo and LoVo/CPT-11 drug-resistant cell lines | Downregulation of CD44, EpCAM, CD24, Bcl-2 and upregulation of Bax. | Attenuated chemoresistance of CRC cells via targeting and inducing apoptosis in CRC. | [97] | ||
| 5–15 µg/mL CUR + 2.5–100 µg/mL IRI | LoVo/CPT-11R drug-resistant cell line | Increase of E-cadherin; downregulation of vimentin and N-cadherin. | Suppressed epithelial-mesenchymal transition (EMT). | [98] | ||
| OXA | In vitro: HCT116 and SW480 cells 0–8 µM CUR + 0.5–32 µM OXA; HCT116/OXA cells 4 µM CUR + 8 µM OXA. In vivo: 60 mg/kg CUR + 10 mg/kg OXA | HCT116, SW480, HCT116/OXA drug-resistant cell lines and BALB/c nude mice | Inhibition of TGF-β/Smad2/Smad3 signaling. | Inhibition of cell proliferation and reduced tumor weight and volume. | [99] | |
| RG | 15, 30 μM CUR + 0–1 μM RG | HCT-116 (KRAS mutant) and HT-29 (KRAS wild-type) cell lines | - | CUR behaved like MEK-specific inhibitor (U0126) to enhance RG-induced growth inhibition, apoptosis, and autophagy in HCT-116 cells. | [100] | |
| Liver cancer | CXB | 1.25–40 μM CUR + 3.125–100 μM CXB | HepG2 cell line | Inhibition of cell proliferation via the downregulation of NF-κβ, PGE2, MDA and Akt phosphorylation; suppression of cyclin D1 and VEGF expression. Increased apoptotic activities via the upregulation of caspase 3 activity. | Synergistic antiproliferative interaction. Possible use of lower and safer doses of CXB. | [38] |
| CIS | 15.6–500 μM CUR + 10, 25 μM CIS | HepG2 cell line | - | The combination of CIS with CUR inhibited cell viability and exhibited a significant agonist effect in selected cancer cells in a time and dose dependent manner. | [101] | |
| 5-FU | In vitro: 5, 10 μM CUR + 2.5, 5, 10 μM 5-FU. In vivo: 56.65 mg/kg CUR + 10 mg/kg 5-FU | SMMC-7721, Bel-7402, HepG-2, MHCC97H cell lines and BALB/c nude mice | Decreased expression of NF-κB protein in the nucleus. Increased expression of NF-κB protein in cytoplasm. Downregulation of COX-2 expression. | Synergistic effects and in vivo tumor growth inhibition. | [102] | |
| PTX | 5, 10, 20 μM CUR + 0.16–10.24 μM PTX | Hep3B and HepG2 cell lines | Downregulation of Lin28. | Synergistic effect. CUR increased the sensitivity of HCC cells to PTX. | [103] | |
| SOR | 60, 120 μM CUR + 0.25–10 μM SOR | Hep3b and HepG2 cell lines | S-phase and G2/M phase arrest of liver cancer cells, induced apoptosis, reduced the protein levels of cyclins A, B2 and D1, phosphorylated retinoblastoma and B-cell lymphoma (Bcl), increased the protein levels of Bcl-2-associated X protein, cleaved caspase-3 and cleaved caspase-9. | CUR augmented the apoptosis-inducing potential of SOR. | [104] | |
| Gastric cancer | DOX | 2.5–30 μg/mL CUR + 2.5–30 μg/mL DOX | AGS cell line | Activation of pro-apoptotic protein Bax, repression of anti-apoptotic protein Bcl-2, upregulation of caspase 9 activity. | Decrease in AGS cell viability. Dose-dependent inhibition of cell invasion and migration. | [81] |
| Breast cancer | APA | 25–100 μM CUR + 25–100 μM APA | MCF7 cell line | Induced apoptosis of breast cancer cells through increased expression of apoptosis-inducing BAX and SMAC genes as well as decreased expression of apoptosis inhibitor BCL2 and SURVIVIN genes. | Combination therapy exerts more profound anti-proliferation effects on breast cancer cells than APA or CUR monotherapy. | [105] |
| CARB | 5 µM CUR + 2 µM CARB | CAL-51, CAL-51-R and MDA-MB-231 cell lines | Increasing ROS production, which downregulated the DNA repair protein RAD51, leading to upregulation of γH2AX. | CUR sensitizes TNBC to the anticancer effect of CARB. | [106] | |
| DOX | 25 µM CUR + 5 µM DOX | MCF-7/DOX drug-resistant cell line | Reduced the Aurora-A expression. Triggered P53 stabilization. Growth arrest and apoptosis induction. | Reversed DOX insensitivity and increased sensitivity in DOX-resistant MCF-7 and MCF-7 cell lines. | [107] | |
| 10 µM CUR + 2.5–100 µM DOX | MCF-7/DOX and MDA-MB-231/DOX drug-resistant cell lines | Suppression of PI3K/Akt, GSK3β, β-catenin phosphorylation. Inhibition of efflux function of ABCB4 via the inhibition of ATPase activities of ABCB4. Inhibition of EMT via the upregulation of E-cadherin; downregulation of Smad2 phosphorylation. | Enhanced the sensitivity of breast cancer cells to DOX. Increased intracellular levels of DOX and reversed chemoresistance. | [108] | ||
| LAP | 1.5 μM CUR + 5, 20 nM LAP | AU-565 cell line | Inhibition of cell proliferation via the suppression of Akt phosphorylation. Reversal of HER2-induced chemoresistance via the downregulation of HER2. | Potentiated action of the metastasis treatment drug LAP. | [109] | |
| PTX | In vitro: 0.01, 0.1 µM CUR + 0.01–100 µM PAX. In vivo: 50 mg/kg CUR, p.o., 3 times/week + 10 mg/kg PTX, i.p., once/week | MCF-7EAC-tumor bearing mice | Inhibition of the ALDH-1 and PTX-induced Pgp-1 expression. Synergistic cytotoxic interaction via upregulation of Bax, caspase-7, -9 and downregulation of Bcl-2 expression. Inhibition of PTX-induced Pgp-1, and -ALDH-1 expression in the animal model. | Using CUR enhanced the tumor response to PTX. | [110] | |
| 30 µM CUR + 10 nM PTX | MCF-7 and MDA-MB-234 cell lines | Increased caspase 3 activation, PARP cleavage, loss of membrane integrity. Increased apoptotic effect of PTX. Reduced PTX-induced NF-κB. | Higher level of apoptosis compared with either substance alone. | [111] |
| Cancer Type | Chemotherapy | Dosage | Assay Type | Molecular Effect | Study Conclusion | Ref. |
|---|---|---|---|---|---|---|
| Lung cancer | GEM | In vitro: 10 µM RES + 1 µM GEM. In vivo: 25 mg/kg GEM i.p. 2×/week + 1 µmol/kg RES 5×/week | HCC827 cell lines and HCC827 xenografts in nude mice | Downregulation of mRNA and protein levels of ENG, activation of ERK signaling pathway. | RES promoted tumor microvessel growth, increased blood perfusion and drug delivery into tumor that resulted in enhanced anticancer effect of GEM. | [157] |
| Colorectal cancer | 5-FU | 10 mg/kg b.w. RES p.o./day + 12.5 mg/kg b.w. 5-FU i.p. injected on days 1, 3, and 5; repeated every 4 weeks for 4 months | Methyl nitrosourea-induced colon cancer in male albino rats | Decrease of NF-κB and reduction of COX-2, induced p53 gene expression. | RES biochemically modulated and enhanced the therapeutic effects of 5-FU. | [169] |
| 0–200 µM RES + 10 µM 5-FU | DLD1 and HCT116 cell lines | Abolished CD44 expression, inhibition of STAT3 and Akt signaling pathways, decreased binding of STAT3 to the hTERT promoter, subsequently reduced telomerase activity. | RES enhanced the antitelomeric and pro-apoptotic potential of 5-FU in CRC, and led to re-sensitization to chemotherapy. | [170] | ||
| 5 μM RES + 1 nM 5-FU | HCT116 and HCT116R/5-FU drug-resistant cell lines | Suppressed expression of NF-κB, MMP-9 and CXCR4, induced caspase-3 cleavage, suppressed vimentin, transcription factor slug and induction of E-cadherin. | RES chemosensitizes CRC cells to 5-FU in TNF-β-induced inflammatory tumor microenvironment. | [171] | ||
| Liver cancer | CIS | 12.5 μg/mL RES + 0.625 μg/mL CIS, 25 µg/mL RES + 1.25 µ/mL CIS | C3A and SMCC7721 cell lines | Reduced glutamine transporter ASCT2 expression and glutamine uptake, affected expression of cytochrome c, caspase-9 and activated caspase-3. | Synergistic effects and enhanced CIS toxicity in human hepatoma cell lines. | [158] |
| SOR | In vitro: 80 μM RES + 2.5, 5, 10 μM SOR. In vivo: RES (20 mg/kg, i.p.) + SOR (25 mg/kg, p.o.) 2×/week for 3 weeks. | HepG2, Huh7 HCC cell lines and BALB/c nude mice | Accumulation of cells in S phase and decrease of G0/G1 phase, decreased levels of CDK2 and CDC25A and increased level of cyclin A, increased levels of cleaved caspase-3, caspase-8, and caspase-9 proteins, decreased expression of PKA, p-AMPK, and eEF2K. | Synergistic effects in vitro and in vivo. | [159] | |
| 40, 80 μM RES + 0.25–10 μM SOR | Hep3b and HepG2 cell lines | - | RES potentiated the lethality of SOR. | [104] | ||
| Gastric cancer | CIS | 20 μM RES + 1 μg/mL CIS | AGS cell line | Upregulation of Bax and the cleaved form of PARP, downregulation of Bcl-2, increased PERK, p-eIF2α and CHOP protein levels. Activation of PERK/eIF2α/ATF4/CHOP signaling pathway, induction of G2/M cell cycle arrest. | Synergistically inhibited cell growth of cancer cell lines. | [151] |
| Breast Cancer | CIS | 12.5, 25, 50 μM RES + 4 μM CIS | MDA-MB-231cell lines and female BALB/c mice MDA-MB-231 xenografts | The expressions of P-AKT, P-PI3K, Smad2, Smad3, P-JNK, and P-ERK induced by TGF-β1 were reversed after RES and CIS co-treatment. | Synergistic effect on the inhibition of breast cancer cell viability, migration, and invasion in vitro; enhanced anti-tumor effect and reduced side effect of CIS in vivo. | [160] |
| 57.5, 72 μM RES + 18.5, 23 μM CIS | MDA-MB-231 cell line | Activation of the caspase-9 and caspase-3 enzymes, higher mitochondrial membrane depolarization. | Co-treatment induced a higher rate of apoptosis. | [161] | ||
| 0–250 μM RES + 2–50 μM CIS | MCF-7, MCF-7R, T47-D and MDA-MB-231 cell lines | Enhanced antiproliferative effect, reduction of the HR initiation complex mRNA components in MCF-7 and MCF-7R cells. | Co-treatment lowered the concentrations of CIS needed for the equivalent effect compared with CIS alone. | [162] | ||
| DOX | 30 µM RES + 100 nM DOX | MCF7 cell line | - | RES potentiated long-term toxicity of DOX, probably due to the long-term increase of apoptosis and senescence in MCF-7 cells. | [163] | |
| 50 μmol/L RES + 4 μg/mL DOX | MCF-7 and MCF-7/ADR drug-resistant cell lines | RES reversed DOX induced upregulation of vimentin and N-cadherin and β-catenin, upregulated SIRT1 expression, reversed EMT and inhibited cell migration in MCF7/ADR cells. | RES reversed DOX-resistance in MCF-7/ADR cells. | [164] | ||
| 100, 200, 300 μM RES + 2 mg/mL DOX | MCF-7/ADR drug resistant cell line | Activation of caspase-8 and caspase-9, inhibition of proliferation and decreased cell viability, miRNA miR-122-5p upregulation and miR-542-3p downregulation, the expression levels of targeted proteins of these miRNAs significantly reduced. | RES chemo-sensitizes drug resistant cancer cell lines. | [65] | ||
| In vitro: 10 mg/L RES + 1 mg/L DOX In vivo: 3 mg/kg DOX i.p. every week and 50 mg/kg RES p.o. for 4 weeks | MCF-7, MCF-7/DOX drug resistant cell lines and nude mice xenograft model | In vitro: PI3K and cleaved caspase-3 upregulation, reduced ratios p-Akt/Akt and p-mTOR/mTOR in MCF-7/DOX cells. In vivo: significant increase in the expression of PI3K and cleaved caspase-3, reduced p70 S6K and Ki67 expression. | In vitro, RES reversed DOX resistance, inhibited DOX-resistant breast cancer cell propagation and metastasis and facilitated cell apoptosis. In vivo, RES and DOX synergistically reduced the tumor volume. | [165] | ||
| 20 mg/kg/day RES with 2.5 mg/kg DOX in six injections for 2 weeks | MCF-7 cell line and xenografts in mice | 99mTc-MIBI uptake in MCF-7 cells was significantly reduced due to higher apoptosis in tumor cells. | The combination of RES and DOX enhanced the antitumor effect and reduced DOX cardiotoxicity and hepatotoxicity. | [166] | ||
| PTX | 1 μM RES + 1, 10, 100 nM PTX | MCF-7, T47D (ERα+) and MDA-MB 231 (ERα−) cell lines | - | RES enhanced cell sensitivity to PTX and lowered the doses of PTX. | [167] |
| Cancer Type | Chemotherapy | Dosage | Assay Type | Molecular Effect | Study Conclusion | Ref. |
|---|---|---|---|---|---|---|
| Lung cancer | CIS | EGCG (1.5 mg/mouse/day IP) for 5 days and CIS (2 or 4 mg/kg IP) on day 5; EGCG (1.5 mg/mouse/day) and single-dose CIS (2 mg/mouse) on day 0 or 5 | A549 cell xenograft bearing BALB/c nude mice | Increased CIS concentration in tumor tissue and tumor growth delay due to EGCG induced vascular normalization. | EGCG synergistically potentiated CIS antitumor efficacy especially when CIS was applied during the vascular normalization window. | [175] |
| 0–25 µM EGCG + 2.5 µM CIS; 15 µM EGCG + IC90 CIS | H460 cell line | Inhibition of ERCC1/XPF activity and the repair of CIS induced interstrand crosslinks. | EGCG enhanced sensitivity to CIS in a lung cancer cell line. | [176] | ||
| DOX | 0.5 μM EGCG + 0–100 μM DOX | Nonresponsive A549 cell line | Decreased drug efflux, MDR signaling and invasiveness. Increased drug internalization, cell cycle arrest, stress induced damage and cell death. | EGCG reversed the compromised functionality of DOX in a nonresponsive A549 cell line and improved its oxidative damage-mediated antitumor effect by modulating redox signaling. | [177] | |
| DOX or 5-FU | 0.3 μM EGCG + 10, 20 µM 5-FU for HCT15 cells; 36 μM EGCG + 0.25, 0.5 µM DOX for A549 cells | HCT15 colon and A549 lung cancer cell lines | Increased growth inhibitory effect of 5-FU and DOX, as well as their effect on apoptosis, but not on cell cycle. EGCG sensitized 5-FU and DOX to further suppress ERK phosphorylation. | EGCG combination with DOX or 5-FU reduced cancer cell growth in different cancer cell lines. | [181] | |
| ETO | 0.05–500 µM EGCG + 0–100 μM ETO | Nonresponsive A549 cell line | Downregulation of MRP-1 and increased intracellular uptake and retention of ETO. Suppression of MMP-2 and MMP-9 activity. EGCG helped maintain an optimum level of Nrf2 which contributed to overcoming ETO resistance. | EGCG sensitized lung adenocarcinoma cells towards ETO chemotherapy by inducing G2/M arrest and suppressing the multidrug resistance. | [178] | |
| GEF or OSM | 10–150 µM EGCG + 1, 2.5, 5 µM GEF for PC9-GR1, PC9-GR3 and PC9-GR4 cells; 10–150 µM EGCG + 0.5, 1, 2 µM OSM for PC9-GR3 cells | PC9/GEF drug resistant cell lines: PC9-GR1, PC9-GR3 and PC9-GR4 | The results shed light on the possible involvement of FASN/EGFR/STAT3 pathways. | EGCG combination with either GEF or OSM resulted in mostly additive effects. | [179] | |
| GEF | In vitro: 34 μM EGCG + 1.87 μM GEF; 40 µM EGCG + 10 µM GEF. In vivo: 200 mg/kg/day EGCG and 10 mg/kg/day GEF, p.o. | A549/GEF drug resistant cell line and A549 cell xenograft bearing BALB/c nude mice | Inhibition of GEF induced autophagy and ERK phosphorylation, as well as LC3-II/I and ATG5 expression, while the expression of p62 increased. | Synergistic inhibition of GEF resistant NSCLC cell proliferation and tumor growth suppression in a xenograft mouse model. | [180] | |
| Colorectal cancer | DOX or 5-FU | 0.3 μM EGCG + 10, 20 µM 5-FU for HCT15 cells; 36 μM EGCG + 0.25, 0.5 µM DOX for A549 cells | HCT15 colon and A549 lung cancer cell lines | Increased growth inhibitory effect of 5-FU and DOX, as well as their effect on apoptosis, but not on cell cycle. EGCG sensitized 5-FU and DOX to further suppress ERK phosphorylation. | EGCG combination with DOX or 5-FU reduced cancer cell growth in different cancer cell lines. | [181] |
| DOX | 100, 109, 117 µM EGCG + 3.63, 3.08, 3.07 µM DOX | Caco-2 cell line | EGCG decreased the dose of DOX needed to reach cytotoxicity by mediating P-gp activity. | Non-toxic EGCG concentrations combined with DOX resulted in antagonism or slight additivity in Caco-2 cell line. | [182] | |
| 5-FU | 100 µM EGCG + 0.1–100 µM 5-FU | HT-29 cell line | Significant decrease in cell proliferation. | Cotreatment enhanced the sensitivity of HT-29 cells to 5-FU by 12-fold. | [183] | |
| 5-FU or IRI | 0.2, 1, 6, 12 µM EGCG + 1, 10 µM 5-FU for KM12 cells; 0.2, 1, 6, 12 µM EGCG + 1, 10 µM 5-FU for WiDr cells; 2, 20 µM EGCG + 2, 40 µM IRI for SW837 cells | KM12, WiDr and SW837 cell lines | Synergy can only be observed in some cell lines and the underlying mechanism is possibly a combination of several mechanisms, not just a simple induction of MET. | The combination of EGCG and 5-FU resulted in synergy for WiDr cell line, while no synergy was observed for KM12 cell line. The combination of EGCG and IRI did not result in synergy for SW837 cell line. | [184] | |
| 5-FU | In vitro: 50 µM EGCG + 1, 5, 10, 15, 20, 30 µM 5-FU. In vivo: intratumoral injection of 25 mg/kg EGCG + 20 mg/kg 5-FU for 14 successive days | HCT-116, DLD1 cell lines and DLD1 tumor bearing BALB/c nude mice | Significant enhancement of cancer cell apoptosis and DNA damage. Inhibition of GRP78 expression and increased NF-κB and miR-155-5p levels, followed by a decrease in MDR1 expression and promotion of 5-FU accumulation in cancer cells. Activation of caspase-3 and PARP, inhibition of Bcl-2 expression and increased level of Bad. | EGCG enhanced the sensitivity of colon cancer cells to 5-FU. | [185] | |
| IRI | In vitro: 20, 50 μM EGCG + 0.5 μM IRI. In vivo: 5 mg/kg EGCG 1 time per day, i.p. + 4 mg/kg IRI 2 times per week, i.p. | RKO, HCT116 cell lines and HCT116 tumor bearing BALB/c nude mice | Increased intracellular GRP78 expression, decreased mitochondrial membrane potential, as well as intracellular ROS and induced cell apoptosis. | EGCG increased the sensitivity of colorectal cancer cells to IRI. | [186] | |
| 2, 5, 10, 20, 50, 100, 200 μM EGCG + 0.5 μM IRI for RKO cells. 5, 10, 20, 50 μM EGCG + 0.5 μM IRI for HCT116 cells | RKO and HCT116 cell lines | Enhanced inhibitory effect on tumor cells, induced cell apoptosis and prevention of tumor cell migration and invasion. Inhibition of topoisomerase I caused cell cycle arrest in S or G2 phase. | EGCG and IRI combination resulted in enhanced DNA damage in human colorectal cancer cells and synergistic antitumor effects. | [187] | ||
| Liver cancer | SOR | 100 mg/kg EGCG + 10 mg/kg SOR | Diethyl nitrosamine induced hepatocellular carcinoma in Wistar albino rats | Histopathological observations revealed a satisfying decline in tissue degeneration and hyperchromatism. Significantly lower alpha-fetoprotein and liver enzyme levels were detected, as well as a greater antioxidant capacity. | EGCG and SOR combination had a comparable effect with standard dose SOR. The combination resulted in enhanced chemoprotection and is considered effective against hepatocellular carcinoma. | [188] |
| Gastric cancer | CIS | 25 μg/mL EGCG + 5 μg/mL CIS | BGC-823 cell lines | Significant nuclear shrinkage and reduction in proliferation rate, cloning efficiency and cell migration. Cycle arrest in G1 phase, increased apoptosis and up-regulation of p19Arf, p53 and p21Cip1 gene and protein expression. | EGCG enhanced CIS antitumor effect against gastric cancer cells. | [189] |
| Breast cancer | ATO and/or irradiation | 10–100 µM EGCG + 2 Gy radiation; 10–100 µM EGCG and 4 µM ATO. 10–100 µM EGCG, 4 µM ATO and 2 Gy radiation. | MCF-7 cell lines | Bax upregulation and Bcl-2 downregulation. | Combination of EGCG and ATO with or without radiation showed synergistic effects in breast cancer treatment visible in the rise of cell death. | [190] |
| CLF | 10 µM EGCG + 640 nM CLF for MCF-7 cells; 10 µM EGCG + 50 nM CLF for MDA-MB-231 cells | MCF-7 and MDA-MB-231 cell lines | Enhanced inhibitory effect of CLF on RARB promoter methylation and consequential induction of RARB expression. An increase in PTEN and CDKN1A transcript levels was also observed. | EGCG and CLF combination synergistically inhibited cell growth and induced apoptosis. The combination exerted a promising anticancer effect. | [191] | |
| SAHA | 5 μM EGCG + 3 μM SAHA | ERα (+) MCF-7, ERα (−) MDA-MB-157, MDA-MB-231 and HCC1806 cell lines | Decreased expression of cIAP2 and increased expression of caspase 7. Changes in histone modifications indicate an involvement of epigenetic mechanisms in cIAP2 expression modulation. Increased apoptosis and a reduction of TNBC cell migration. | EGCG and SAHA combination successfully induced breast cancer cell apoptosis and reduced their migratory ability. | [192] | |
| 5 μM EGCG + 3 μM SAHA | ERα (+) MCF-7, ERα (−) MDA-MB-157, MDA-MB-231 and HCC1806 cell lines | Decreased density of cancer cells and changes in expression of ERα, oncogenic miR-221/222, p27 and PTEN. Changes in histone acetylation indicate an involvement of epigenetic mechanisms in tumor suppressor expression modulation. Increased E-cadherin and decreased N-cadherin expression levels. DNMT activity and the migratory capacity of TNBC cells were reduced. | EGCG and SAHA combination successfully limited the growth, proliferation and migration of breast cancer cells. | [193] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jakobušić Brala, C.; Karković Marković, A.; Kugić, A.; Torić, J.; Barbarić, M. Combination Chemotherapy with Selected Polyphenols in Preclinical and Clinical Studies—An Update Overview. Molecules 2023, 28, 3746. https://doi.org/10.3390/molecules28093746
Jakobušić Brala C, Karković Marković A, Kugić A, Torić J, Barbarić M. Combination Chemotherapy with Selected Polyphenols in Preclinical and Clinical Studies—An Update Overview. Molecules. 2023; 28(9):3746. https://doi.org/10.3390/molecules28093746
Chicago/Turabian StyleJakobušić Brala, Cvijeta, Ana Karković Marković, Azra Kugić, Jelena Torić, and Monika Barbarić. 2023. "Combination Chemotherapy with Selected Polyphenols in Preclinical and Clinical Studies—An Update Overview" Molecules 28, no. 9: 3746. https://doi.org/10.3390/molecules28093746
APA StyleJakobušić Brala, C., Karković Marković, A., Kugić, A., Torić, J., & Barbarić, M. (2023). Combination Chemotherapy with Selected Polyphenols in Preclinical and Clinical Studies—An Update Overview. Molecules, 28(9), 3746. https://doi.org/10.3390/molecules28093746