YTHDF1 in Tumor Cell Metabolism: An Updated Review


Abstract
:
1. Introduction
2. YTHDF1 and Tumorigenesis
2.1. Digestive System Tumors
2.1.1. Colorectal Cancer
2.1.2. Gastric Cancer
2.1.3. Esophageal Cancer
2.1.4. Liver Cancer
2.1.5. Pancreatic Cancer
2.2. Respiratory System Tumors
2.2.1. Non-Small Cell Lung Cancer
2.2.2. Nasopharyngeal Carcinoma
2.2.3. Laryngeal Cancer
2.3. Genitourinary System Tumors
2.3.1. Renal Cell Carcinoma
2.3.2. Bladder Cancer
2.3.3. Prostate Cancer
2.3.4. Breast Cancer
2.3.5. Cervical Cancer
2.4. Other Tumors
2.4.1. Thyroid Cancer
2.4.2. Osteosarcoma
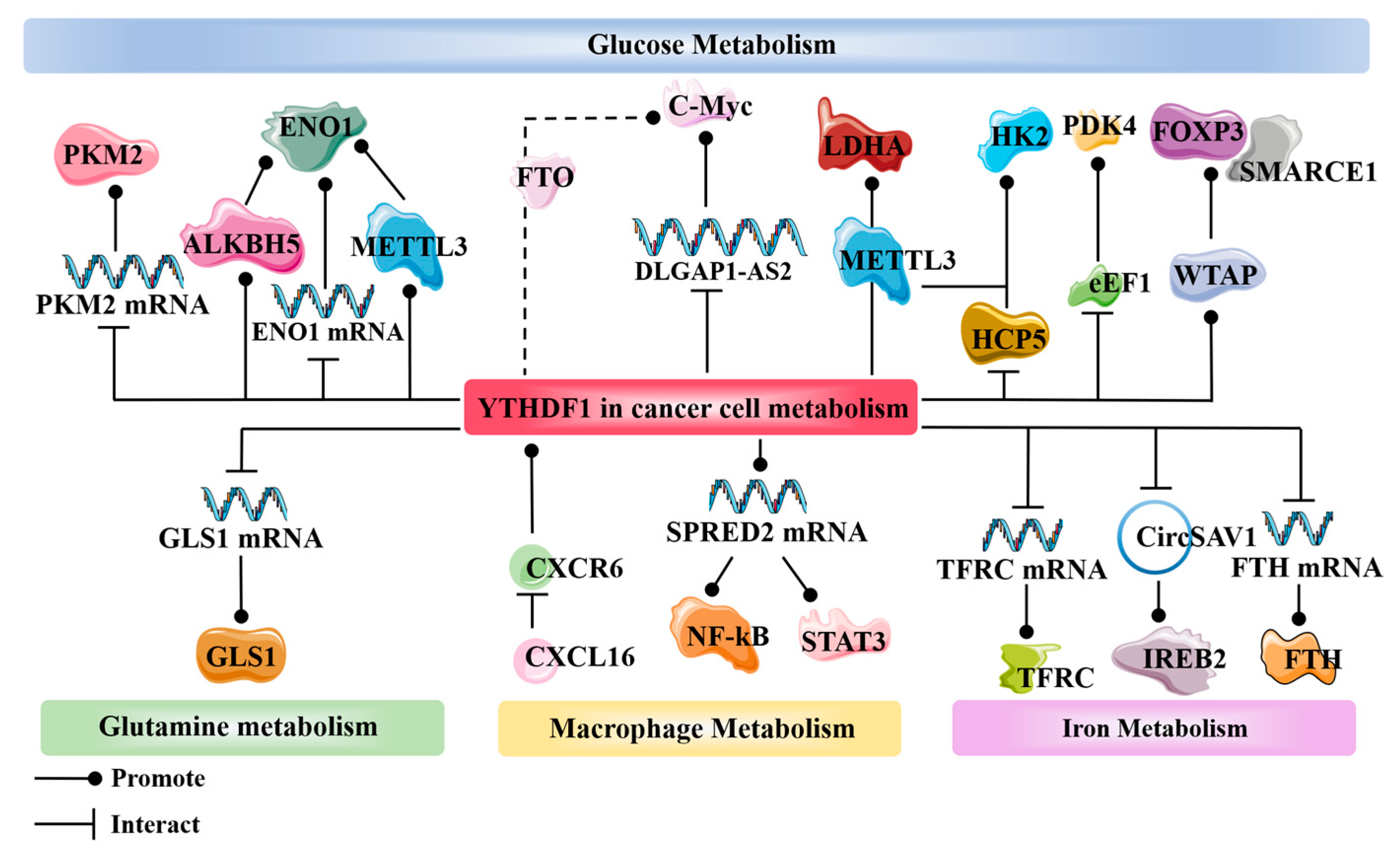
3. YTHDF1 and Cancer Metabolism
3.1. Glucose Metabolism
3.2. Glutamine Metabolism
3.3. Macrophage Metabolism
3.4. Iron Metabolism
3.5. Other Metabolisms
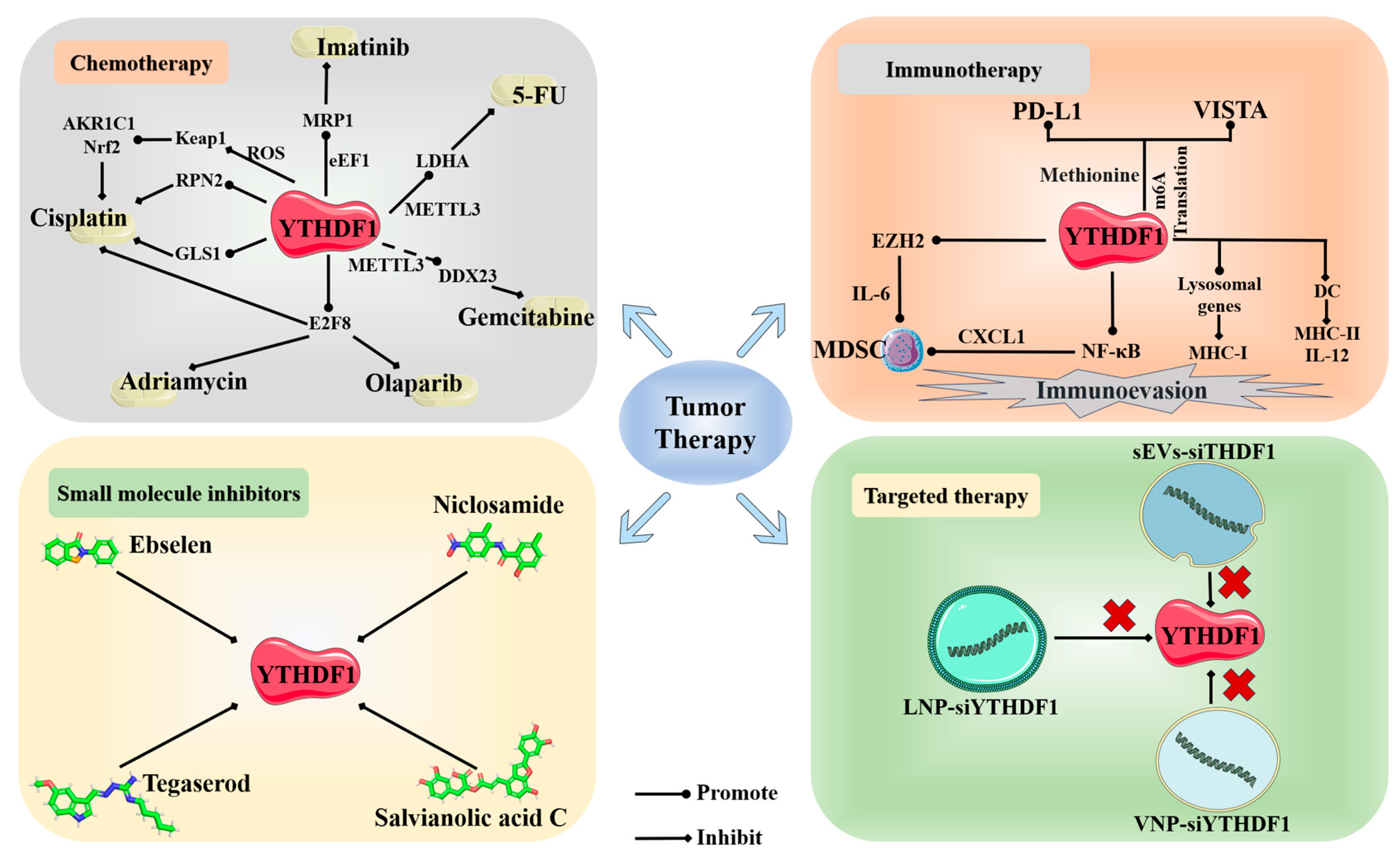
4. YTHDF1 and Cancer Therapy
4.1. Chemotherapy
4.2. Immunotherapy
4.3. Small Molecule Inhibitors
4.4. Other Therapies
5. Conclusions and Perspectives
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Desrosiers, R.; Friderici, K.; Rottman, F. Identification of methylated nucleosides in messenger RNA from Novikoff hepatoma cells. Proc. Natl. Acad. Sci. USA 1974, 71, 3971–3975. [Google Scholar] [CrossRef] [PubMed]
- Dominissini, D.; Moshitch-Moshkovitz, S.; Schwartz, S.; Salmon-Divon, M.; Ungar, L.; Osenberg, S.; Cesarkas, K.; Jacob-Hirsch, J.; Amariglio, N.; Kupiec, M.; et al. Topology of the human and mouse m6A RNA methylomes revealed by m6A-seq. Nature 2012, 485, 201–206. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.D.; Saletore, Y.; Zumbo, P.; Elemento, O.; Mason, C.E.; Jaffrey, S.R. Comprehensive analysis of mRNA methylation reveals enrichment in 3’ UTRs and near stop codons. Cell 2012, 149, 1635–1646. [Google Scholar] [CrossRef] [PubMed]
- Meyer, K.D.; Jaffrey, S.R. Rethinking m(6)A Readers, Writers, and Erasers. Annu. Rev. Cell Dev. Biol. 2017, 33, 319–342. [Google Scholar] [CrossRef] [PubMed]
- He, P.C.; He, C. m(6) A RNA methylation: From mechanisms to therapeutic potential. EMBO J. 2021, 40, e105977. [Google Scholar] [CrossRef]
- Bokar, J.A.; Shambaugh, M.E.; Polayes, D.; Matera, A.G.; Rottman, F.M. Purification and cDNA cloning of the AdoMet-binding subunit of the human mRNA (N6-adenosine)-methyltransferase. Rna 1997, 3, 1233–1247. [Google Scholar]
- Liu, J.; Yue, Y.; Han, D.; Wang, X.; Fu, Y.; Zhang, L.; Jia, G.; Yu, M.; Lu, Z.; Deng, X.; et al. A METTL3-METTL14 complex mediates mammalian nuclear RNA N6-adenosine methylation. Nat. Chem. Biol. 2014, 10, 93–95. [Google Scholar] [CrossRef]
- Agarwala, S.D.; Blitzblau, H.G.; Hochwagen, A.; Fink, G.R. RNA methylation by the MIS complex regulates a cell fate decision in yeast. PLoS Genet. 2012, 8, e1002732. [Google Scholar] [CrossRef]
- Jia, G.; Fu, Y.; Zhao, X.; Dai, Q.; Zheng, G.; Yang, Y.; Yi, C.; Lindahl, T.; Pan, T.; Yang, Y.G.; et al. N6-methyladenosine in nuclear RNA is a major substrate of the obesity-associated FTO. Nat. Chem. Biol. 2011, 7, 885–887. [Google Scholar] [CrossRef]
- Zheng, G.; Dahl, J.A.; Niu, Y.; Fedorcsak, P.; Huang, C.M.; Li, C.J.; Vågbø, C.B.; Shi, Y.; Wang, W.L.; Song, S.H.; et al. ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility. Mol. Cell 2013, 49, 18–29. [Google Scholar] [CrossRef]
- Huang, H.; Weng, H.; Sun, W.; Qin, X.; Shi, H.; Wu, H.; Zhao, B.S.; Mesquita, A.; Liu, C.; Yuan, C.L.; et al. Recognition of RNA N(6)-methyladenosine by IGF2BP proteins enhances mRNA stability and translation. Nat. Cell Biol. 2018, 20, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Patil, D.P.; Chen, C.K.; Pickering, B.F.; Chow, A.; Jackson, C.; Guttman, M.; Jaffrey, S.R. m(6)A RNA methylation promotes XIST-mediated transcriptional repression. Nature 2016, 537, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Alarcón, C.R.; Goodarzi, H.; Lee, H.; Liu, X.; Tavazoie, S.; Tavazoie, S.F. HNRNPA2B1 Is a Mediator of m(6)A-Dependent Nuclear RNA Processing Events. Cell 2015, 162, 1299–1308. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Hsu, P.J.; Chen, Y.S.; Yang, Y.G. Dynamic transcriptomic m(6)A decoration: Writers, erasers, readers and functions in RNA metabolism. Cell Res. 2018, 28, 616–624. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Lu, Z.; Gomez, A.; Hon, G.C.; Yue, Y.; Han, D.; Fu, Y.; Parisien, M.; Dai, Q.; Jia, G.; et al. N6-methyladenosine-dependent regulation of messenger RNA stability. Nature 2014, 505, 117–120. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, B.S.; Roundtree, I.A.; Lu, Z.; Han, D.; Ma, H.; Weng, X.; Chen, K.; Shi, H.; He, C. N6-methyladenosine Modulates Messenger RNA Translation Efficiency. Cell 2015, 161, 1388–1399. [Google Scholar] [CrossRef] [PubMed]
- Xiao, W.; Adhikari, S.; Dahal, U.; Chen, Y.S.; Hao, Y.J.; Sun, B.F.; Sun, H.Y.; Li, A.; Ping, X.L.; Lai, W.Y.; et al. Nuclear m(6)A Reader YTHDC1 Regulates mRNA Splicing. Mol. Cell 2016, 61, 507–519. [Google Scholar] [CrossRef]
- Nachtergaele, S.; He, C. The emerging biology of RNA post-transcriptional modifications. RNA Biol. 2017, 14, 156–163. [Google Scholar] [CrossRef]
- Zhuang, M.; Li, X.; Zhu, J.; Zhang, J.; Niu, F.; Liang, F.; Chen, M.; Li, D.; Han, P.; Ji, S.J. The m6A reader YTHDF1 regulates axon guidance through translational control of Robo3.1 expression. Nucleic Acids Res. 2019, 47, 4765–4777. [Google Scholar] [CrossRef]
- Chen, Z.; Zhong, X.; Xia, M.; Zhong, J. The roles and mechanisms of the m6A reader protein YTHDF1 in tumor biology and human diseases. Mol. Ther. Nucleic Acids 2021, 26, 1270–1279. [Google Scholar] [CrossRef]
- Du, H.; Zhao, Y.; He, J.; Zhang, Y.; Xi, H.; Liu, M.; Ma, J.; Wu, L. YTHDF2 destabilizes m(6)A-containing RNA through direct recruitment of the CCR4-NOT deadenylase complex. Nat. Commun. 2016, 7, 12626. [Google Scholar] [CrossRef] [PubMed]
- Shi, H.; Wang, X.; Lu, Z.; Zhao, B.S.; Ma, H.; Hsu, P.J.; Liu, C.; He, C. YTHDF3 facilitates translation and decay of N(6)-methyladenosine-modified RNA. Cell Res. 2017, 27, 315–328. [Google Scholar] [CrossRef] [PubMed]
- Zaccara, S.; Jaffrey, S.R. A Unified Model for the Function of YTHDF Proteins in Regulating m(6)A-Modified mRNA. Cell 2020, 181, 1582–1595.e1518. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Sepich-Poore, C.; Zhou, X.; Wei, J.; He, C. The mechanism underlying redundant functions of the YTHDF proteins. Genome Biol. 2023, 24, 17. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Dong, D.; Xia, Y.; Hao, L.; Wang, W.; Zhao, C. YTHDF1 promotes breast cancer cell growth, DNA damage repair and chemoresistance. Cell Death Dis. 2022, 13, 230. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Yuan, Y.; Li, Y.; Mutti, L.; Peng, J.; Jiang, X. The function and clinical implication of YTHDF1 in the human system development and cancer. Biomark. Res. 2023, 11, 5. [Google Scholar] [CrossRef] [PubMed]
- Anita, R.; Paramasivam, A.; Priyadharsini, J.V.; Chitra, S. The m6A readers YTHDF1 and YTHDF3 aberrations associated with metastasis and predict poor prognosis in breast cancer patients. Am. J. Cancer Res. 2020, 10, 2546–2554. [Google Scholar]
- Bai, Y.; Yang, C.; Wu, R.; Huang, L.; Song, S.; Li, W.; Yan, P.; Lin, C.; Li, D.; Zhang, Y. YTHDF1 Regulates Tumorigenicity and Cancer Stem Cell-Like Activity in Human Colorectal Carcinoma. Front. Oncol. 2019, 9, 332. [Google Scholar] [CrossRef]
- Wang, S.; Gao, S.; Zeng, Y.; Zhu, L.; Mo, Y.; Wong, C.C.; Bao, Y.; Su, P.; Zhai, J.; Wang, L.; et al. N6-Methyladenosine Reader YTHDF1 Promotes ARHGEF2 Translation and RhoA Signaling in Colorectal Cancer. Gastroenterology 2022, 162, 1183–1196. [Google Scholar] [CrossRef]
- Li, J.; Ahmad, M.; Sang, L.; Zhan, Y.; Wang, Y.; Yan, Y.; Liu, Y.; Mi, W.; Lu, M.; Dai, Y.; et al. O-GlcNAcylation promotes the cytosolic localization of the m(6)A reader YTHDF1 and colorectal cancer tumorigenesis. J. Biol. Chem. 2023, 299, 104738. [Google Scholar] [CrossRef]
- Chen, X.Y.; Liang, R.; Yi, Y.C.; Fan, H.N.; Chen, M.; Zhang, J.; Zhu, J.S. The m(6)A Reader YTHDF1 Facilitates the Tumorigenesis and Metastasis of Gastric Cancer via USP14 Translation in an m(6)A-Dependent Manner. Front. Cell Dev. Biol. 2021, 9, 647702. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Nie, J.; Xu, K.; Liu, Y.; Tong, W.; Li, A.; Zuo, W.; Liu, Z.; Yang, F. YY1 is regulated by ALKBH5-mediated m6A modification and promotes autophagy and cancer progression through targeting ATG4B. Aging 2023, 15, 9590–9613. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; He, Q.; Liu, J.; Li, N.; Xiao, K.; Chen, H. PLAGL2 promotes Snail expression and gastric cancer progression via UCA1/miR-145-5p/YTHDF1 axis. Carcinogenesis 2023, 44, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.; Sun, M.; Chen, Y.; Zhuo, S.; Song, G.; Wang, T.; Zhang, Z. H19 recruited N6-methyladenosine (m6A) reader YTHDF1 to promote SCARB1 translation and facilitate angiogenesis in gastric cancer. Chin. Med. J. 2023, 136, 1719–1731. [Google Scholar] [CrossRef] [PubMed]
- Morgan, E.; Soerjomataram, I.; Rumgay, H.; Coleman, H.G.; Thrift, A.P.; Vignat, J.; Laversanne, M.; Ferlay, J.; Arnold, M. The Global Landscape of Esophageal Squamous Cell Carcinoma and Esophageal Adenocarcinoma Incidence and Mortality in 2020 and Projections to 2040: New Estimates from GLOBOCAN 2020. Gastroenterology 2022, 163, 649–658.e642. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.; Chen, F.; Li, L.; Dou, S.; Li, Q.; Huang, Y.; Li, Z.; Liu, W.; Zhang, G. Intracellular Fusobacterium nucleatum infection increases METTL3-mediated m6A methylation to promote the metastasis of esophageal squamous cell carcinoma. J. Adv. Res. 2023, in press. [Google Scholar] [CrossRef]
- Donne, R.; Lujambio, A. The liver cancer immune microenvironment: Therapeutic implications for hepatocellular carcinoma. Hepatology 2023, 77, 1773–1796. [Google Scholar] [CrossRef]
- Qu, N.; Zhang, X.; Wu, X.; Zhou, X.; Deng, Z.; Ma, L.; Liu, Y.; Ge, W.; Jiang, H.; Xu, L.; et al. Clinical implications of m6A-related regulators YTHDF1 and YTHDF2 in hepatocellular carcinoma. Precis. Med. Sci. 2022, 11, 174–185. [Google Scholar] [CrossRef]
- Zheng, H.; Cheng, Z.J.; Liang, B.; Wang, Z.G.; Tao, Y.P.; Huang, S.Y.; Ni, J.S.; Li, H.F.; Yang, L.; Yuan, S.X.; et al. N(6)-Methyladenosine Modification of ANLN Enhances Hepatocellular Carcinoma Bone Metastasis. Int. J. Biol. Sci. 2023, 19, 1009–1023. [Google Scholar] [CrossRef]
- Huang, X.; Zhu, L.; Wang, L.; Huang, W.; Tan, L.; Liu, H.; Huo, J.; Su, T.; Zhang, M.; Kuang, M.; et al. YTHDF1 promotes intrahepatic cholangiocarcinoma progression via regulating EGFR mRNA translation. J. Gastroenterol. Hepatol. 2022, 37, 1156–1168. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Gao, H.L.; Wang, W.Q.; Yu, X.J.; Liu, L. Molecular drivers and cells of origin in pancreatic ductal adenocarcinoma and pancreatic neuroendocrine carcinoma. Exp. Hematol. Oncol. 2020, 9, 28. [Google Scholar] [CrossRef]
- Huang, R.; Yang, L.; Zhang, Z.; Liu, X.; Fei, Y.; Tong, W.M.; Niu, Y.; Liang, Z. RNA m(6)A Demethylase ALKBH5 Protects Against Pancreatic Ductal Adenocarcinoma via Targeting Regulators of Iron Metabolism. Front. Cell Dev. Biol. 2021, 9, 724282. [Google Scholar] [CrossRef]
- Wu, S.; Ai, Y.; Huang, H.; Wu, G.; Zhou, S.; Hong, W.; Akuetteh, P.D.P.; Jin, G.; Zhao, X.; Zhang, Y.; et al. A synthesized olean-28,13β-lactam targets YTHDF1-GLS1 axis to induce ROS-dependent metabolic crisis and cell death in pancreatic adenocarcinoma. Cancer Cell Int. 2022, 22, 143. [Google Scholar] [CrossRef]
- Chen, Z.; Fillmore, C.M.; Hammerman, P.S.; Kim, C.F.; Wong, K.K. Non-small-cell lung cancers: A heterogeneous set of diseases. Nat. Rev. Cancer 2014, 14, 535–546. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, X.; Cai, J.; Ren, L.; Liu, B.; Wu, M.; Lu, W.; Li, R.; Zhang, C.; Huang, C.; et al. The pathological tissue expression pattern and clinical significance of m6A-regulatory genes in non-small cell lung cancer. J. Gene Med. 2022, 24, e3397. [Google Scholar] [CrossRef]
- Dou, X.; Wang, Z.; Lu, W.; Miao, L.; Zhao, Y. METTL3 promotes non-small cell lung cancer (NSCLC) cell proliferation and colony formation in a m6A-YTHDF1 dependent way. BMC Pulm. Med. 2022, 22, 324. [Google Scholar] [CrossRef]
- Sun, Z.; Su, Z.; Zhou, Z.; Wang, S.; Wang, Z.; Tong, X.; Li, C.; Wang, Y.; Chen, X.; Lei, Z.; et al. RNA demethylase ALKBH5 inhibits TGF-β-induced EMT by regulating TGF-β/SMAD signaling in non-small cell lung cancer. Faseb J. 2022, 36, e22283. [Google Scholar] [CrossRef]
- Brooks, L.; Yao, Q.Y.; Rickinson, A.B.; Young, L.S. Epstein-Barr virus latent gene transcription in nasopharyngeal carcinoma cells: Coexpression of EBNA1, LMP1, and LMP2 transcripts. J. Virol. 1992, 66, 2689–2697. [Google Scholar] [CrossRef]
- Countryman, J.; Miller, G. Activation of expression of latent Epstein-Barr herpesvirus after gene transfer with a small cloned subfragment of heterogeneous viral DNA. Proc. Natl. Acad. Sci. USA 1985, 82, 4085–4089. [Google Scholar] [CrossRef]
- Xia, T.L.; Li, X.; Wang, X.; Zhu, Y.J.; Zhang, H.; Cheng, W.; Chen, M.L.; Ye, Y.; Li, Y.; Zhang, A.; et al. N(6)-methyladenosine-binding protein YTHDF1 suppresses EBV replication and promotes EBV RNA decay. EMBO Rep. 2021, 22, e50128. [Google Scholar] [CrossRef]
- Li, T.; Tian, L.; Cao, J.; Liu, M. Cancer-associated fibroblasts secret extracellular vesicles to support cell proliferation and epithelial-mesenchymal transition in laryngeal squamous cell carcinoma. Mol. Cell. Probes 2023, 72, 101934. [Google Scholar] [CrossRef]
- Bahadoram, S.; Davoodi, M.; Hassanzadeh, S.; Bahadoram, M.; Barahman, M.; Mafakher, L. Renal cell carcinoma: An overview of the epidemiology, diagnosis, and treatment. G. Ital. Nefrol. 2022, 39, 1. [Google Scholar]
- Zhang, L.; Luo, X.; Qiao, S. METTL14-mediated N6-methyladenosine modification of Pten mRNA inhibits tumour progression in clear-cell renal cell carcinoma. Br. J. Cancer 2022, 127, 30–42. [Google Scholar] [CrossRef]
- Li, A.; Cao, C.; Gan, Y.; Wang, X.; Wu, T.; Zhang, Q.; Liu, Y.; Yao, L.; Zhang, Q. ZNF677 suppresses renal cell carcinoma progression through N6-methyladenosine and transcriptional repression of CDKN3. Clin. Transl. Med. 2022, 12, e906. [Google Scholar] [CrossRef]
- Ying, X.; Huang, Y.; Liu, B.; Hu, W.; Ji, D.; Chen, C.; Zhang, H.; Liang, Y.; Lv, Y.; Ji, W. Targeted m(6)A demethylation of ITGA6 mRNA by a multisite dCasRx-m(6)A editor inhibits bladder cancer development. J. Adv. Res. 2023, in press. [Google Scholar] [CrossRef]
- Hu, J.; Qiu, D.; Yu, A.; Hu, J.; Deng, H.; Li, H.; Yi, Z.; Chen, J.; Zu, X. YTHDF1 Is a Potential Pan-Cancer Biomarker for Prognosis and Immunotherapy. Front. Oncol. 2021, 11, 607224. [Google Scholar] [CrossRef]
- Zhu, J.; Tong, H.; Sun, Y.; Li, T.; Yang, G.; He, W. YTHDF1 Promotes Bladder Cancer Cell Proliferation via the METTL3/YTHDF1-RPN2-PI3K/AKT/mTOR Axis. Int. J. Mol. Sci. 2023, 24, 6905. [Google Scholar] [CrossRef]
- Han, C.; Chen, S.; Ma, H.; Wen, X.; Wang, Z.; Xu, Y.; Jin, X.; Yu, X.; Wang, M. RPN2 Predicts Poor Prognosis and Promotes Bladder Cancer Growth and Metastasis via the PI3K-Akt Pathway. Onco Targets Ther. 2021, 14, 1643–1657. [Google Scholar] [CrossRef]
- Shafi, A.A.; Yen, A.E.; Weigel, N.L. Androgen receptors in hormone-dependent and castration-resistant prostate cancer. Pharmacol. Ther. 2013, 140, 223–238. [Google Scholar] [CrossRef]
- Li, P.; Shi, Y.; Gao, D.; Xu, H.; Zou, Y.; Wang, Z.; Li, W. ELK1-mediated YTHDF1 drives prostate cancer progression by facilitating the translation of Polo-like kinase 1 in an m6A dependent manner. Int. J. Biol. Sci. 2022, 18, 6145–6162. [Google Scholar] [CrossRef] [PubMed]
- Kashyap, D.; Pal, D.; Sharma, R.; Garg, V.K.; Goel, N.; Koundal, D.; Zaguia, A.; Koundal, S.; Belay, A. Global Increase in Breast Cancer Incidence: Risk Factors and Preventive Measures. BioMed Res. Int. 2022, 2022, 9605439. [Google Scholar] [CrossRef] [PubMed]
- Tai, J.; Wang, L.; Guo, H.; Yan, Z.; Liu, J. Prognostic implications of N(6)-methyladenosine RNA regulators in breast cancer. Sci. Rep. 2022, 12, 1222. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Chen, L.; Zhang, H.; Qiu, X.; Huang, Q.; Wan, F.; Le, Z.; Geng, S.; Zhang, A.; Qiu, S.; et al. Tumor-intrinsic YTHDF1 drives immune evasion and resistance to immune checkpoint inhibitors via promoting MHC-I degradation. Nat. Commun. 2023, 14, 265. [Google Scholar] [CrossRef] [PubMed]
- Jing, L.; Zhou, K.; Wang, Z.; Li, Y.; Fan, Y.; Liu, T.; Shan, Z.; Lin, Y. YTHDF1 shapes “cold” tumor and inhibits CD8+ T cells infiltration and function in breast cancer. Exp. Cell Res. 2023, 432, 113778. [Google Scholar] [CrossRef] [PubMed]
- Ji, H.; Zhang, J.A.; Liu, H.; Li, K.; Wang, Z.W.; Zhu, X. Comprehensive characterization of tumor microenvironment and m6A RNA methylation regulators and its effects on PD-L1 and immune infiltrates in cervical cancer. Front. Immunol. 2022, 13, 976107. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.W.; Lang, B.H.H.; McLeod, D.S.A.; Newbold, K.; Haymart, M.R. Thyroid cancer. Lancet 2023, 401, 1531–1544. [Google Scholar] [CrossRef]
- Zhu, Y.; Peng, X.; Zhou, Q.; Tan, L.; Zhang, C.; Lin, S.; Long, M. METTL3-mediated m6A modification of STEAP2 mRNA inhibits papillary thyroid cancer progress by blocking the Hedgehog signaling pathway and epithelial-to-mesenchymal transition. Cell Death Dis. 2022, 13, 358. [Google Scholar] [CrossRef]
- Yuan, Y.; Yan, G.; He, M.; Lei, H.; Li, L.; Wang, Y.; He, X.; Li, G.; Wang, Q.; Gao, Y.; et al. ALKBH5 suppresses tumor progression via an m(6)A-dependent epigenetic silencing of pre-miR-181b-1/YAP signaling axis in osteosarcoma. Cell Death Dis. 2021, 12, 60. [Google Scholar] [CrossRef]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Lin, J.; Xia, L.; Liang, J.; Han, Y.; Wang, H.; Oyang, L.; Tan, S.; Tian, Y.; Rao, S.; Chen, X.; et al. The roles of glucose metabolic reprogramming in chemo- and radio-resistance. J. Exp. Clin. Cancer Res. CR 2019, 38, 218. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Shao, F.; Guo, D.; Wang, W.; Wang, J.; Zhu, R.; Gao, Y.; He, J.; Lu, Z. WNT/β-catenin-suppressed FTO expression increases m(6)A of c-Myc mRNA to promote tumor cell glycolysis and tumorigenesis. Cell Death Dis. 2021, 12, 462. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Zhang, Y.; Chen, H.; Sun, L.N.; Zhang, B.; Yue, D.S.; Wang, C.L.; Zhang, Z.F. METTL3-induced DLGAP1-AS2 promotes non-small cell lung cancer tumorigenesis through m(6)A/c-Myc-dependent aerobic glycolysis. Cell Cycle 2022, 21, 2602–2614. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Huang, H.; Shi, J.H.; Yu, X.; Ding, R.; Zhang, Y.; Han, Q.; Ni, Z.Y.; Li, X.; Zhao, R.; et al. USP47 inhibits m6A-dependent c-Myc translation to maintain regulatory T cell metabolic and functional homeostasis. J. Clin. Investig. 2023, 133, e169365. [Google Scholar] [CrossRef]
- Zhang, K.; Zhang, T.; Yang, Y.; Tu, W.; Huang, H.; Wang, Y.; Chen, Y.; Pan, K.; Chen, Z. N(6)-methyladenosine-mediated LDHA induction potentiates chemoresistance of colorectal cancer cells through metabolic reprogramming. Theranostics 2022, 12, 4802–4817. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Guo, X.; Li, L.; Gao, Z.; Su, X.; Ji, M.; Liu, J. N(6)-methyladenosine METTL3 promotes cervical cancer tumorigenesis and Warburg effect through YTHDF1/HK2 modification. Cell Death Dis. 2020, 11, 911. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, Z.; Shi, W.; Shen, J.; Guan, Y.; Ni, F. HLA complex P5 upregulation is correlated with poor prognosis and tumor progression in esophageal squamous cell carcinoma. Bioengineered 2022, 13, 9301–9311. [Google Scholar] [CrossRef]
- Li, Z.; Peng, Y.; Li, J.; Chen, Z.; Chen, F.; Tu, J.; Lin, S.; Wang, H. N(6)-methyladenosine regulates glycolysis of cancer cells through PDK4. Nat. Commun. 2020, 11, 2578. [Google Scholar] [CrossRef]
- Ma, L.; Xue, X.; Zhang, X.; Yu, K.; Xu, X.; Tian, X.; Miao, Y.; Meng, F.; Liu, X.; Guo, S.; et al. The essential roles of m(6)A RNA modification to stimulate ENO1-dependent glycolysis and tumorigenesis in lung adenocarcinoma. J. Exp. Clin. Cancer Res. CR 2022, 41, 36. [Google Scholar] [CrossRef]
- Zhang, Y.; Tian, X.; Bai, Y.; Liu, X.; Zhu, J.; Zhang, L.; Wang, J. WTAP mediates FOXP3 mRNA stability to promote SMARCE1 expression and augment glycolysis in colon adenocarcinoma. Mamm. Genome 2022, 33, 654–671. [Google Scholar] [CrossRef]
- Yao, X.; Li, W.; Li, L.; Li, M.; Zhao, Y.; Fang, D.; Zeng, X.; Luo, Z. YTHDF1 upregulation mediates hypoxia-dependent breast cancer growth and metastasis through regulating PKM2 to affect glycolysis. Cell Death Dis. 2022, 13, 258. [Google Scholar] [CrossRef] [PubMed]
- Altman, B.J.; Stine, Z.E.; Dang, C.V. From Krebs to clinic: Glutamine metabolism to cancer therapy. Nat. Rev. Cancer 2016, 16, 749. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Zhang, Z.; Li, P.; Zhang, Z.; Zeng, M.; Liang, Z.; Li, D.; Wang, L.; Chen, Y.; Liang, Y.; et al. Reprogramming of glutamine metabolism and its impact on immune response in the tumor microenvironment. Cell Commun. Signal 2022, 20, 114. [Google Scholar] [CrossRef] [PubMed]
- Leone, R.D.; Zhao, L.; Englert, J.M.; Sun, I.M.; Oh, M.H.; Sun, I.H.; Arwood, M.L.; Bettencourt, I.A.; Patel, C.H.; Wen, J.; et al. Glutamine blockade induces divergent metabolic programs to overcome tumor immune evasion. Science 2019, 366, 1013–1021. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Byun, J.K.; Choi, Y.K.; Park, K.G. Targeting glutamine metabolism as a therapeutic strategy for cancer. Exp. Mol. Med. 2023, 55, 706–715. [Google Scholar] [CrossRef] [PubMed]
- Yelamanchi, S.D.; Jayaram, S.; Thomas, J.K.; Gundimeda, S.; Khan, A.A.; Singhal, A.; Keshava Prasad, T.S.; Pandey, A.; Somani, B.L.; Gowda, H. A pathway map of glutamate metabolism. J. Cell Commun. Signal 2016, 10, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Vanhove, K.; Derveaux, E.; Graulus, G.J.; Mesotten, L.; Thomeer, M.; Noben, J.P.; Guedens, W.; Adriaensens, P. Glutamine Addiction and Therapeutic Strategies in Lung Cancer. Int. J. Mol. Sci. 2019, 20, 252. [Google Scholar] [CrossRef]
- Chen, P.; Liu, X.Q.; Lin, X.; Gao, L.Y.; Zhang, S.; Huang, X. Targeting YTHDF1 effectively re-sensitizes cisplatin-resistant colon cancer cells by modulating GLS-mediated glutamine metabolism. Mol. Ther. Oncolytics 2021, 20, 228–239. [Google Scholar] [CrossRef]
- Mills, C.D.; Kincaid, K.; Alt, J.M.; Heilman, M.J.; Hill, A.M. M-1/M-2 macrophages and the Th1/Th2 paradigm. J. Immunol. 2000, 164, 6166–6173. [Google Scholar] [CrossRef]
- He, Z.; Zhang, S. Tumor-Associated Macrophages and Their Functional Transformation in the Hypoxic Tumor Microenvironment. Front. Immunol. 2021, 12, 741305. [Google Scholar] [CrossRef]
- Hong, L.; Wang, X.; Zheng, L.; Wang, S.; Zhu, G. Tumor-associated macrophages promote cisplatin resistance in ovarian cancer cells by enhancing WTAP-mediated N6-methyladenosine RNA methylation via the CXCL16/CXCR6 axis. Cancer Chemother. Pharmacol. 2023, 92, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Zhang, X.; Yang, P.; Zhang, X.; Peng, Y.; Li, D.; Yu, Y.; Wu, Y.; Wang, Y.; Zhang, J.; et al. RNA m6A methylation orchestrates cancer growth and metastasis via macrophage reprogramming. Nat. Commun. 2021, 12, 1394. [Google Scholar] [CrossRef]
- Li, Y.; Li, M.; Liu, L.; Xue, C.; Fei, Y.; Wang, X.; Zhang, Y.; Cai, K.; Zhao, Y.; Luo, Z. Cell-Specific Metabolic Reprogramming of Tumors for Bioactivatable Ferroptosis Therapy. ACS Nano 2022, 16, 3965–3984. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.; Wang, Z.; Chen, X.; Jiang, X.; Dong, Z.; Hu, S.; Li, W.; Liu, Y.; Liao, B.; Han, W.; et al. YTHDF1-enhanced iron metabolism depends on TFRC m(6)A methylation. Theranostics 2020, 10, 12072–12089. [Google Scholar] [CrossRef] [PubMed]
- Xia, H.; Wu, Y.; Zhao, J.; Cheng, C.; Lin, J.; Yang, Y.; Lu, L.; Xiang, Q.; Bian, T.; Liu, Q. N6-Methyladenosine-modified circSAV1 triggers ferroptosis in COPD through recruiting YTHDF1 to facilitate the translation of IREB2. Cell Death Differ. 2023, 30, 1293–1304. [Google Scholar] [CrossRef] [PubMed]
- Diao, H.; Tan, H.; Hu, Y.; Wang, R.; Cai, P.; Huang, B.; Shao, X.; Yan, M.; Yin, C.; Zhang, Y. The m(6)A Reader YTHDF1 Promotes Lung Carcinoma Progression via Regulating Ferritin Mediate Ferroptosis in an m(6)A-Dependent Manner. Pharmaceuticals 2023, 16, 185. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Jin, P.; Wang, X. N(6)-methyladenosine regulator YTHDF1 represses the CD8 + T cell-mediated antitumor immunity and ferroptosis in prostate cancer via m(6)A/PD-L1 manner. Apoptosis 2023, 1–12. [Google Scholar] [CrossRef]
- Smith, J.A. The cell cycle and related concepts in cell proliferation. J. Pathol. 1982, 136, 149–166. [Google Scholar] [CrossRef]
- Shi, Y.; Fan, S.; Wu, M.; Zuo, Z.; Li, X.; Jiang, L.; Shen, Q.; Xu, P.; Zeng, L.; Zhou, Y.; et al. YTHDF1 links hypoxia adaptation and non-small cell lung cancer progression. Nat. Commun. 2019, 10, 4892. [Google Scholar] [CrossRef]
- Li, H.; Zhong, Y.; Cao, G.; Shi, H.; Liu, Y.; Li, L.; Yin, P.; Chen, J.; Xiao, Z.; Du, B. METTL3 promotes cell cycle progression via m(6)A/YTHDF1-dependent regulation of CDC25B translation. Int. J. Biol. Sci. 2022, 18, 3223–3236. [Google Scholar] [CrossRef]
- Lou, X.; Ning, J.; Liu, W.; Li, K.; Qian, B.; Xu, D.; Wu, Y.; Zhang, D.; Cui, W. YTHDF1 Promotes Cyclin B1 Translation through m(6)A Modulation and Contributes to the Poor Prognosis of Lung Adenocarcinoma with KRAS/TP53 Co-Mutation. Cells 2021, 10, 1669. [Google Scholar] [CrossRef] [PubMed]
- Duan, X.; Yang, L.; Wang, L.; Liu, Q.; Zhang, K.; Liu, S.; Liu, C.; Gao, Q.; Li, L.; Qin, G.; et al. m6A demethylase FTO promotes tumor progression via regulation of lipid metabolism in esophageal cancer. Cell Biosci. 2022, 12, 60. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Sun, L.; Guo, H.; Zhou, S.; Wang, C.; Ji, C.; Meng, F.; Liang, S.; Zhang, B.; Yuan, Y.; et al. Targeting SLP2-mediated lipid metabolism reprograming restricts proliferation and metastasis of hepatocellular carcinoma and promotes sensitivity to Lenvatinib. Oncogene 2023, 42, 374–388. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Zhang, Q.; Chen, M.; Li, B.; Wang, N.; Li, C.; Gao, Z.; Zhang, D.; Yang, L.; Xu, Z.; et al. N(6)-methyladenosine modification regulates imatinib resistance of gastrointestinal stromal tumor by enhancing the expression of multidrug transporter MRP1. Cancer Lett. 2022, 530, 85–99. [Google Scholar] [CrossRef] [PubMed]
- Lin, C.; Li, T.; Wang, Y.; Lai, S.; Huang, Y.; Guo, Z.; Zhang, X.; Weng, S. METTL3 enhances pancreatic ductal adenocarcinoma progression and gemcitabine resistance through modifying DDX23 mRNA N6 adenosine methylation. Cell Death Dis. 2023, 14, 221. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.; Huang, Y.; Huang, W.; Zhang, Y.; Zhang, K.; Li, Y.; Ouyang, H. Wnt3a/YTHDF1 Regulated Oxaliplatin-Induced Neuropathic Pain Via TNF-α/IL-18 Expression in the Spinal Cord. Cell Mol. Neurobiol. 2023, 43, 1583–1594. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Tan, Y.T.; Chen, Y.X.; Zheng, X.J.; Wang, W.; Liao, K.; Mo, H.Y.; Lin, J.; Yang, W.; Piao, H.L.; et al. Methionine deficiency facilitates antitumour immunity by altering m(6)A methylation of immune checkpoint transcripts. Gut 2023, 72, 501–511. [Google Scholar] [CrossRef]
- Wang, L.; Zhu, L.; Liang, C.; Huang, X.; Liu, Z.; Huo, J.; Zhang, Y.; Zhang, Y.; Chen, L.; Xu, H.; et al. Targeting N6-methyladenosine reader YTHDF1 with siRNA boosts antitumor immunity in NASH-HCC by inhibiting EZH2-IL-6 axis. J. Hepatol. 2023, 79, 1185–1200. [Google Scholar] [CrossRef]
- Bao, Y.; Zhai, J.; Chen, H.; Wong, C.C.; Liang, C.; Ding, Y.; Huang, D.; Gou, H.; Chen, D.; Pan, Y.; et al. Targeting m(6)A reader YTHDF1 augments antitumour immunity and boosts anti-PD-1 efficacy in colorectal cancer. Gut 2023, 72, 1497–1509. [Google Scholar] [CrossRef]
- Bai, X.; Wong, C.C.; Pan, Y.; Chen, H.; Liu, W.; Zhai, J.; Kang, W.; Shi, Y.; Yamamoto, M.; Tsukamoto, T.; et al. Loss of YTHDF1 in gastric tumors restores sensitivity to antitumor immunity by recruiting mature dendritic cells. J. Immunother. Cancer 2022, 10, e003663. [Google Scholar] [CrossRef]
- Sekulovski, N.; MacLean, J.A.; Bheemireddy, S.R.; Yu, Z.; Okuda, H.; Pru, C.; Plunkett, K.N.; Matzuk, M.; Hayashi, K. Niclosamide’s potential direct targets in ovarian cancer†. Biol. Reprod. 2021, 105, 403–412. [Google Scholar] [CrossRef] [PubMed]
- Micaelli, M.; Dalle Vedove, A.; Cerofolini, L.; Vigna, J.; Sighel, D.; Zaccara, S.; Bonomo, I.; Poulentzas, G.; Rosatti, E.F.; Cazzanelli, G.; et al. Small-Molecule Ebselen Binds to YTHDF Proteins Interfering with the Recognition of N (6)-Methyladenosine-Modified RNAs. ACS Pharmacol. Transl. Sci. 2022, 5, 872–891. [Google Scholar] [CrossRef] [PubMed]
- Zou, Z.; Wei, J.; Chen, Y.; Kang, Y.; Shi, H.; Yang, F.; Shi, Z.; Chen, S.; Zhou, Y.; Sepich-Poore, C.; et al. FMRP phosphorylation modulates neuronal translation through YTHDF1. Mol. Cell 2023, 83, 4304–4317.e8. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Lin, J.; Huang, D.; Ye, C.; Chen, D. Salvianolic Acid C Inhibits the Epithelial-Mesenchymal Transition and Ameliorates Renal Tubulointerstitial Fibrosis. Front Biosci-Landmark 2023, 28, 121. [Google Scholar] [CrossRef] [PubMed]
- Hong, Y.G.; Yang, Z.; Chen, Y.; Liu, T.; Zheng, Y.; Zhou, C.; Wu, G.C.; Chen, Y.; Xia, J.; Wen, R.; et al. The RNA m6A Reader YTHDF1 Is Required for Acute Myeloid Leukemia Progression. Cancer Res. 2023, 83, 845–860. [Google Scholar] [CrossRef] [PubMed]
- Shah, E.D.; Lacy, B.E.; Chey, W.D.; Chang, L.; Brenner, D.M. Tegaserod for Irritable Bowel Syndrome with Constipation in Women Younger Than 65 Years Without Cardiovascular Disease: Pooled Analyses of 4 Controlled Trials. Am. J. Gastroenterol. 2021, 116, 1601–1611. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, Y.; Mao, Q.; Jiang, X.; Jiang, W.; Chen, J.; Xu, W.; Zhong, L.; Sun, X. Overexpression of YTHDF1 is associated with poor prognosis in patients with hepatocellular carcinoma. Cancer Biomark. 2018, 21, 859–868. [Google Scholar] [CrossRef]
- You, Q.; Wang, F.; Du, R.; Pi, J.; Wang, H.; Huo, Y.; Liu, J.; Wang, C.; Yu, J.; Yang, Y.; et al. m(6) A Reader YTHDF1-Targeting Engineered Small Extracellular Vesicles for Gastric Cancer Therapy via Epigenetic and Immune Regulation. Adv. Mater. 2023, 35, e2204910. [Google Scholar] [CrossRef]
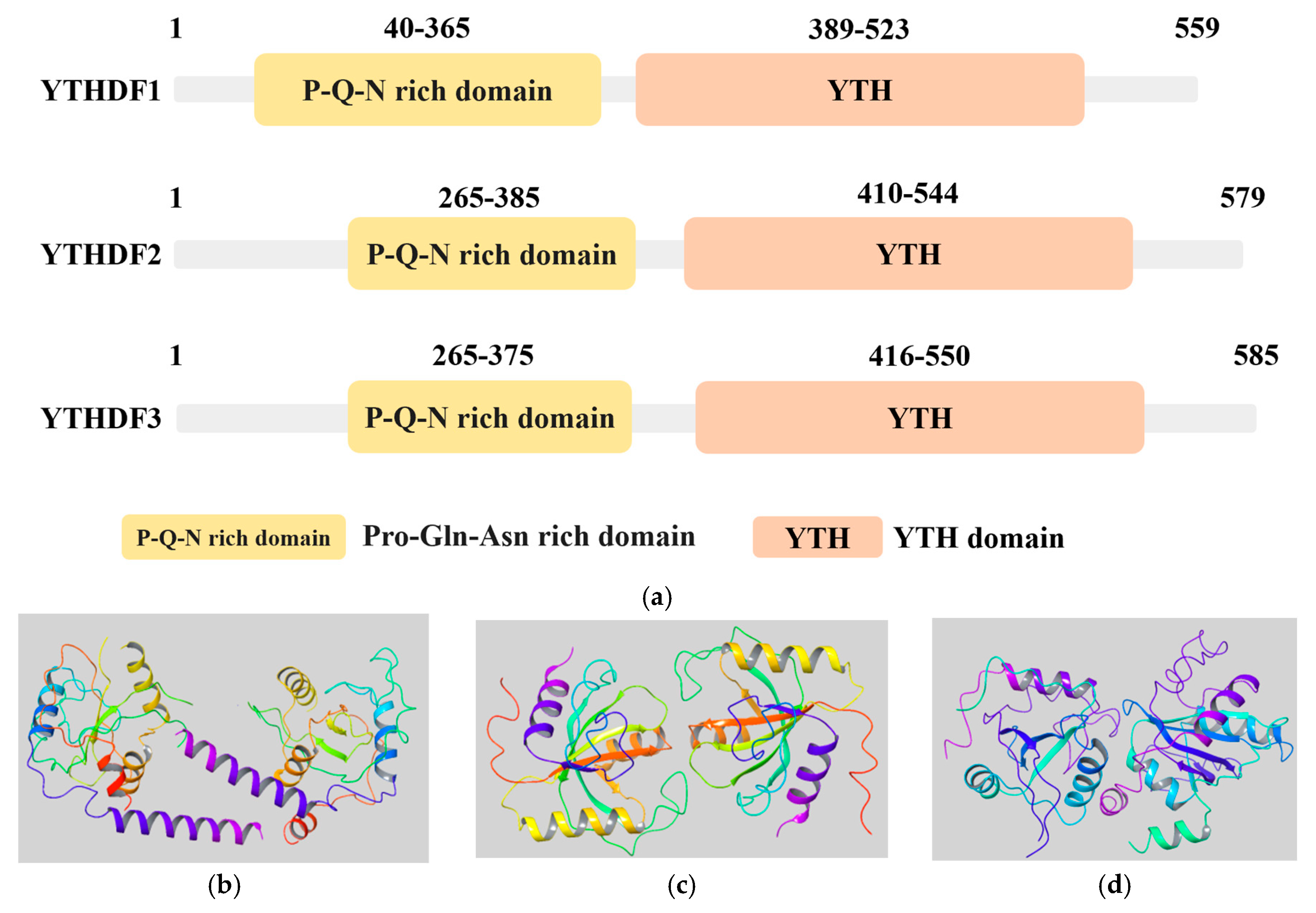

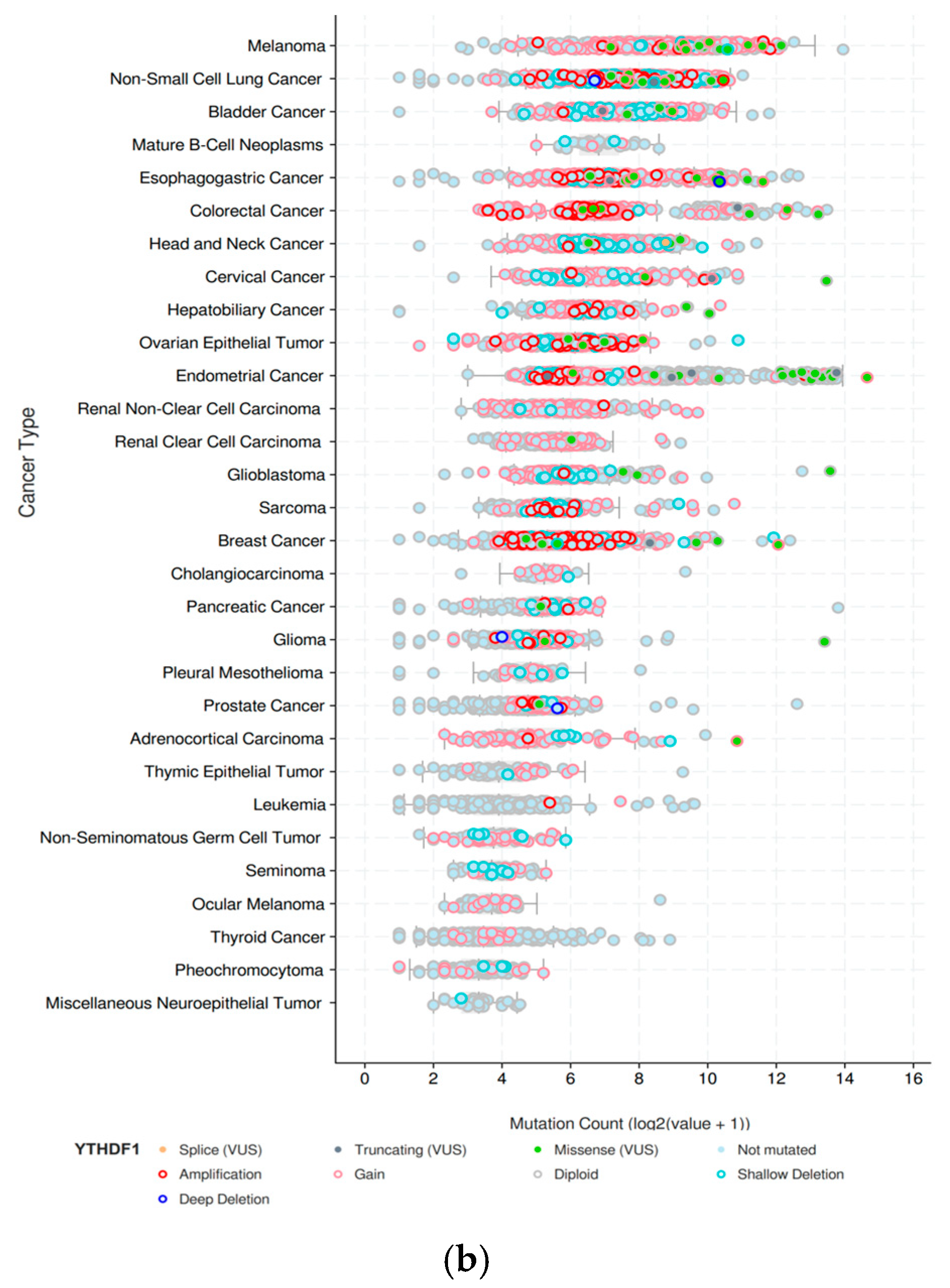

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Mechanism | Function | Type of Cancer | Reference |
| c-Myc | The OGT facilitates the cytoplasmic localization of YTHDF1, thereby upregulating the expression of c-Myc | Inducing tumor initiation | CRC | [30] |
| YY1 | YTHDF1 enhances YY1 expression, and YY1 upregulates ATG4B by binding to its promoter | Dysregulating autophagy | GC | [32] |
| Snail | YTHDF1 interacts with eEF-2, recognizing and facilitating the expression of Snail | Inducing EMT and tumor migration | GC | [33] |
| SCARB1 | H19 binds to YTHDF1, recognizing and promoting the translation of SCARB1 mRNA | Promoting tumor proliferation, migration, and angiogenesis | GC | [34] |
| c-Myc | Fn upregulates the expression of METTL3 and enhances the translation of c-Myc mRNA through YTHDF1 | Promoting tumor proliferation and migration | ESCC | [36] |
| ANLN | YTHDF1 collaborates with METTL3 to facilitate the translation of ANLN through an m6A-dependent mechanism | Promoting bone metastasis | HCC | [39] |
| EGFR | YTHDF1 binds to the 3’-UTR site of EGFR mRNA and promotes translation | Promoting tumor proliferation and migration | HCC | [40] |
| KGA, GAC | The downregulation of YTHDF1 by B28 subsequently decreases the expression of GLS1 isoform KGA/GAC | Increasing ROS, emerging bioenergetic crisis and apoptosis | PDAC | [44] |
| FRAS1 | METTL3-FRAS1 and YTHDF1 collaboratively regulate CDON | Facilitating tumor proliferation and colony formation | NSCLC | [47] |
| TGFβR2, SMAD3 | YTHDF1 collaborates with YTHDF3 in the selective recognition of TGFβR2 and SMAD mRNA | Inducing EMT | NSCLC | [48] |
| BZLF1, BRLF1 | YTHDF1, together with ZAP, DX17, and DCP2, collaboratively decaps and decreases the stability of BZLF1 mRNA and BRLF1 mRNA | Inhibiting the activation and re-infection of EBV | NPC | [51] |
| TUC338 | YTHDF1 potentially regulates the translation stability of the lncRNA TUC338 together with METTL3 | Enhancing the invasiveness of tumor cells | LC | [52] |
| Pten | METTL14 overexpression enhances the stability of Pten mRNA by an m6A–YTHDF1-dependent mechanism | Inhibiting tumor proliferation, migration and AKT signaling activation | RCC | [54] |
| ZNF677 | YTHDF1 enhances the expression level of ZNF677, which significantly inhibits CDKN3 at both mRNA and protein levels | Inhibiting tumor proliferation and inducing apoptosis | RCC | [55] |
| RPN2 | A high expression of METTL3/YTHDF1 increases the level of RPN2, thereby activating the PI3K/AKT pathway | Accelerating tumor proliferation and progression | BLCA | [58] |
| PLK1 | The expression of YTHDF1 is activated by ELK1, thereby promoting the translation of m6A-dependent Polo-like kinase 1 | Promoting tumorigenesis and metastasis | PRAD | [61] |
| STEAP2 | Low expression of METTL3 leads to a decrease in the YTHDF1-mediated translation of STEAP2 mRNA | Enhancing tumor invasion and poor prognosis | PTC | [68] |
| YAP | YTHDF1 recognizes methylated YAP transcripts to promote its translation | Enhancing tumor proliferation, invasion and metastasis | OS | [69] |
| Downstream Effectors | Mechanism | Metabolic Types | Types of Tumors | References |
|---|---|---|---|---|
| c-Myc | YTHDF1 mediates the enhancement of c-Myc mRNA stability by DLGAP1-AS2. | Glucose Metabolism | NSCLC | [73] |
| The downregulation of FTO promotes the recruitment of YTHDF1 and induces the translation of c-Myc mRNA. | LUAD | [72] | ||
| LDHA | YTHDF1 is recruited by METTL3-methylated LDHA mRNA and facilitates translation. | Glucose Metabolism | CRC | [75] |
| PKM2 | YTHDF1 enhances the translation and signaling of PKM2. | Glucose Metabolism | BLCA | [81] |
| ENO1 | Methylated ENO1 mRNA is recognized by YTHDF1, facilitating the translation of ENO1 mRNA. | Glucose Metabolism | LUAD | [79] |
| HK2 | METTL3 interacts with HK2 mRNA through the recognition function of YTHDF1. | Glucose Metabolism | CC | [76] |
| PDK4 | The interaction between YTHDF1 and eEF1 enhances the translation of PDK4 mRNA. | Glucose Metabolism | CC | [76] |
| HK2, HCP5 | The direct interaction between YTHDF1 and HCP5 amplifies the translation of HK2 mRNA. | Glucose Metabolism | ESCC | [77] |
| FOXP3 | The collaborative action of YTHDF1 and WTAP stabilizes FOXP3 mRNA, enabling its interaction with SMARCE1 and activation. | Glucose Metabolism | CRC | [80] |
| GLS1 | YTHDF1 selectively binds to the 3’ UTR of GLS1 mRNA and enhances translation. | Glutamine metabolism | CRC | [88] |
| SPRED2 | YTHDF1 mediates the translation of SPRED2, thereby activating NF-kB and STAT3. | Macrophage Metabolism | LUNG | [92] |
| TFRC | YTHDF1 selectively binds to both the 3’UTR and 5’UTR of TRFC mRNA, thereby enhancing translation. | Iron Metabolism | HPSCC | [94] |
| IREB2 | YTHDF1 interacts with CircSAV1 to form a ternary complex with IREB2 mRNA, thereby promoting the translation of IREB2. | Iron Metabolism | COPD | [95] |
| FTH | YTHDF1 facilitates the translation of FTH, thereby accelerating the transport of free iron. | Iron Metabolism | LUAD | [96] |
| CDC25B | YTHDF1 is modulated by METTL3 to regulate the translation of CDC25B. | Cellular Cycle Metabolism | CC | [100] |
| Cyclin B1 | YTHDF1 promotes translation of Cyclin B1. | Cellular Cycle Metabolism | LUAD | [101] |
| HSD17B11 | YTHDF1 decreases the translation efficiency of HSD17B11. | Lipid metabolism | ESCC | [102] |
| SLP2 | YTHDF1 facilitates the translation of SLP2 in an m6A-dependent manner, which promotes the interaction between SLP2 and the C-terminus of JNK2, thereby enhancing the activity of SREBP1. | Lipid metabolism | HCC | [103] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rong, H.; Wang, D.; Wang, Y.; Dong, C.; Wang, G. YTHDF1 in Tumor Cell Metabolism: An Updated Review. Molecules 2024, 29, 140. https://doi.org/10.3390/molecules29010140
Rong H, Wang D, Wang Y, Dong C, Wang G. YTHDF1 in Tumor Cell Metabolism: An Updated Review. Molecules. 2024; 29(1):140. https://doi.org/10.3390/molecules29010140
Chicago/Turabian StyleRong, Haichuan, Danyang Wang, Yiran Wang, Chenshuang Dong, and Guiling Wang. 2024. "YTHDF1 in Tumor Cell Metabolism: An Updated Review" Molecules 29, no. 1: 140. https://doi.org/10.3390/molecules29010140
APA StyleRong, H., Wang, D., Wang, Y., Dong, C., & Wang, G. (2024). YTHDF1 in Tumor Cell Metabolism: An Updated Review. Molecules, 29(1), 140. https://doi.org/10.3390/molecules29010140