
A Systematic Analysis of the Correlation between Flavor Active Differential Metabolites and Multiple Bean Ripening Stages of Coffea arabica L.
 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. Widely Targeted Metabolomics Analysis of Coffee Beans at Different Maturity Levels
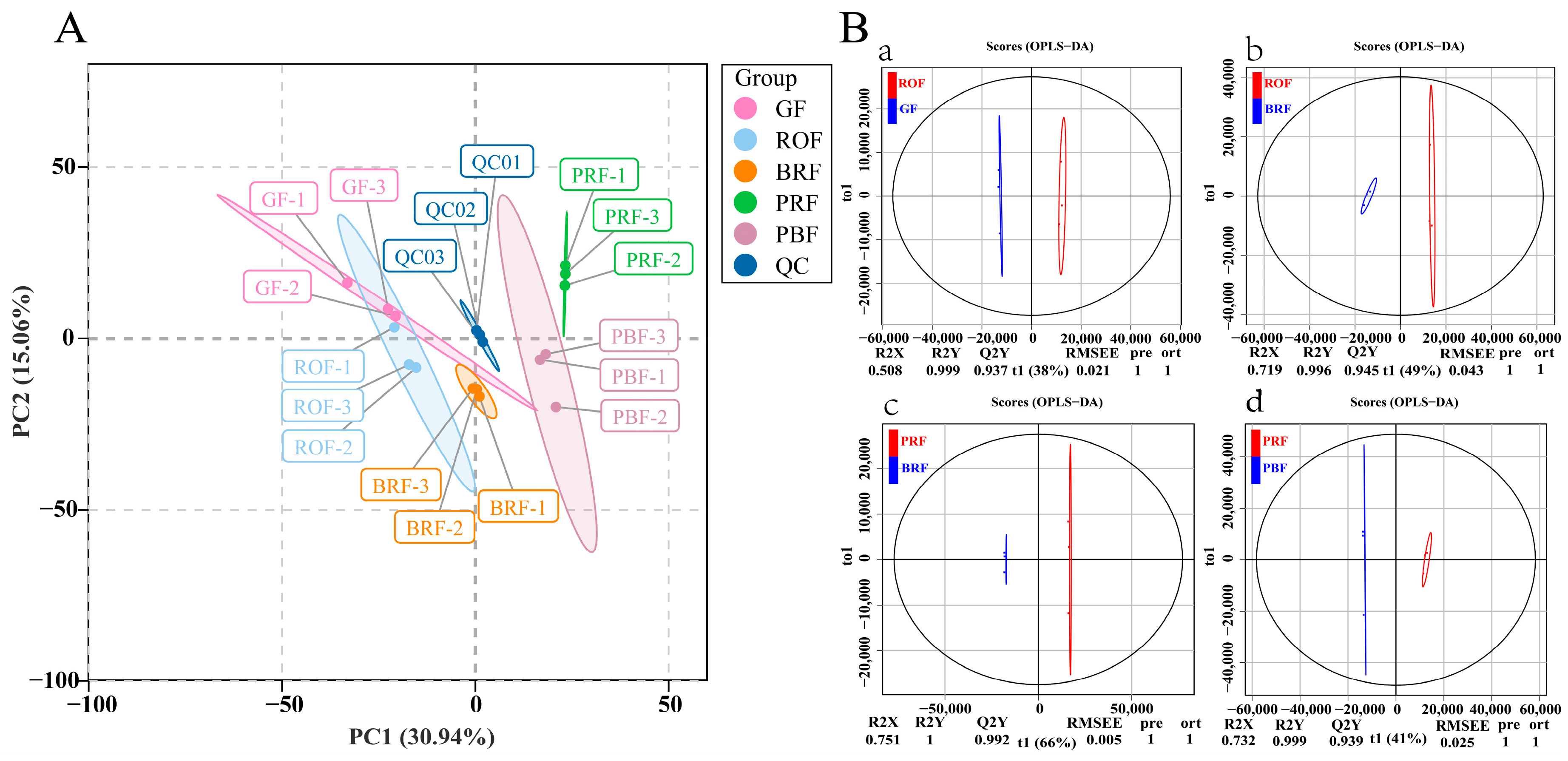
2.2. Multivariate Analysis of the Identified Metabolites
2.3. Identification and Classification of Differential Metabolites
2.4. Kyoto Encyclopedia of Genes and Genomes (KEGG) Enrichment Analysis of Differential Metabolites
2.5. Correlation Analysis of Characteristic Metabolites during Cherry Development and Ripening
3. Conclusions
4. Materials and Methods
4.1. Plant Materials and Test Condition
4.2. Sample Preparation and Extraction
4.3. UPLC Conditions
4.4. ESI Q-TRAP-MS/MS
4.5. Qualitative and Quantitative Analysis of Metabolites
4.6. Metabolites Analysis
4.7. KEGG Annotation and Enrichment Analysis
4.8. Quantification of Taste Indicators
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- DaMatta, F.M.; Avila, R.T.; Cardoso, A.A.; Martins, S.C.V.; Ramalho, J.C. Physiological and agronomic performance of the coffee crop in the context of climate change and global warming: A review. J. Agric. Food Chem. 2018, 66, 5264–5274. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.G.; Bi, X.F.; Liu, H.M.; Fu, X.F.; Li, Y.A.; Yang, Y.; Zhang, X.F.; Wu, R.R.; Li, G.P.; Lv, Y.L.; et al. Transcriptome and carotenoid profiling of different varieties of Coffea arabica provides insights into fruit color formation. Plant Divers. 2022, 44, 322–334. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.Y.; Li, J.P.; Jin, X.L.; Cesarino, I.; Zeng, W.; Li, Z. Characterization of sensory properties of Yunnan coffee. Curr. Res. Food Sci. 2022, 5, 1205–1215. [Google Scholar] [CrossRef] [PubMed]
- Cordoba, N.; Pataquiva, L.; Osorio, C.; Moreno, F.L.M.; Ruiz, R.Y. Effect of grinding, extraction time and type of coffee on the physicochemical and flavour characteristics of cold brew coffee. Sci. Rep. 2019, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Bressanello, D.; Liberto, E.; Cordero, C.; Rubiolo, P.; Pellegrino, G.; Ruosi, M.R.; Bicchi, C. Coffee aroma: Chemometric comparison of the chemical information provided by three different samplings combined with GC-MS to describe the sensory properties in cup. Food Chem. 2017, 214, 218–226. [Google Scholar] [CrossRef]
- Worku, M.; de Meulenaer, B.; Duchateau, L.; Boeckx, P. Effect of altitude on biochemical composition and quality of green arabica coffee beans can be affected by shade and postharvest processing method. Food Res. Int. 2018, 105, 278–285. [Google Scholar] [CrossRef]
- Bastian, F.; Hutabarat, O.S.; Dirpan, A.; Nainu, F.; Harapan, H.; Emran, T.B.; Simal-Gandara, J. From Plantation to Cup: Changes in Bioactive Compounds during Coffee Processing. Foods 2021, 10, 2827. [Google Scholar] [CrossRef]
- Joët, T.; Laffargue, A.; Descroix, F.; Doulbeau, S.; Bertrand, B.; de Kochko, A.; Dussert, S. Influence of environmental factors, wet processing and their interactions on the biochemical composition of green Arabica coffee beans. Food Chem. 2010, 118, 693–701. [Google Scholar] [CrossRef]
- Velásquez, S.; Peña, N.; Bohórquez, J.C.; Gutierrez, N.; Sacks, G.L. Volatile and sensory characterization of roast coffees—Effects of cherry maturity. Food Chem. 2019, 274, 137–145. [Google Scholar] [CrossRef]
- Amorim, A.C.L.; Hovell, A.M.C.; Pinto, A.C.; Eberlin, M.N.; Arruda, N.P.; Pereira, E.J.; Bizzo, H.R.; Catharino, R.R.; Morais, Z.B.; Rezende, C.M. Green and Roasted Arabica Coffees Differentiated by Ripeness, Process and Cup Quality via Electrospray Ionization Mass Spectrometry Fingerprinting. J. Braz. Chem. Soc. 2009, 20, 313–321. [Google Scholar] [CrossRef]
- Craig, A.P.; Franca, A.S.; Oliveira, L.S. Evaluation of the potential of FTIR and chemometrics for separation between defective and non-defective coffees. Food Chem. 2012, 132, 1368–1374. [Google Scholar] [CrossRef]
- Smrke, S.; Kroslakova, I.; Gloess, A.N.; Yeretzian, C. Differentiation of degrees of ripeness of Catuai and Tipica green coffee by chromatographical and statistical techniques. Food Chem. 2015, 174, 637–642. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.L.; Gao, Y.; Peng, X.R.; Liu, J.H.; Su, H.G.; Huang, Y.J.; Qiu, M.H. Lactam ent-Kaurane Diterpene: A New Class of Diterpenoids Present in Roasted Beans of Coffea arabica. J. Agric. Food Chem. 2020, 68, 6112–6121. [Google Scholar] [CrossRef] [PubMed]
- Choi, M.Y.; Choi, W.; Park, J.H.; Lim, J.; Kwon, S.W. Determination of coffee origins by integrated metabolomic approach of combining multiple analytical data. Food Chem. 2010, 121, 1260–1268. [Google Scholar] [CrossRef]
- Chung, S.W.C.; Wu, A.H.T. Development and validation of an analytical method for the analysis of Sterigmatocystin in roasted coffee beans and black pepper using liquid chromatography-tandem mass spectrometry. Food Addit. Contam. Part A-Chem. Anal. Control Expo. Risk Assess. 2020, 37, 355–362. [Google Scholar] [CrossRef] [PubMed]
- Sunarharum, W.B.; Williams, D.J.; Smyth, H.E. Complexity of coffee flavor: A compositional and sensory perspective. Food Res. Int. 2014, 62, 315–325. [Google Scholar] [CrossRef]
- Dixon, R.A.; Xie, D.Y.; Sharma, S.B. Proanthocyanidins—A final frontier in flavonoid research? New Phytol. 2005, 165, 9–28. [Google Scholar] [CrossRef]
- Eskelinen, M.H.; Kivipelto, M. Caffeine as a Protective Factor in Dementia and Alzheimer’s Disease. J. Alzheimers Dis. 2010, 20, 167–174. [Google Scholar] [CrossRef]
- Saravanakumar, K.; Park, S.; Sathiyaseelan, A.; Kim, K.N.; Cho, S.H.; Mariadoss, A.V.A.; Wang, M.H. Metabolite Profiling of Methanolic Extract of Gardenia jaminoides by LC-MS/MS and GC-MS and Its Anti-Diabetic, and Anti-Oxidant Activities. Pharmaceuticals 2021, 14, 102. [Google Scholar] [CrossRef]
- Roychoudhury, S.; Das, D.; Das, S.; Jha, N.K.; Pal, M.; Kolesarova, A.; Kesari, K.K.; Kalita, J.C.; Slama, P. Clinical Potential of Himalayan Herb Bergenia ligulata: An Evidence-Based Study. Molecules 2022, 27, 7039. [Google Scholar] [CrossRef]
- Liu, W.W.; Li, J.D.; Zhang, X.M.; Zu, Y.X.; Yang, Y.; Liu, W.J.; Xu, Z.H.; Gao, H.; Sun, X.; Jiang, X.W.; et al. Current Advances in Naturally Occurring Caffeoylquinic Acids: Structure, Bioactivity, and Synthesis. J. Agric. Food Chem. 2020, 68, 10489–10516. [Google Scholar] [CrossRef]
- Horman, I.; Viani, R. The nature and conformation of the caffeine-chloroginate complex of coffee. J. Food Sci 1972, 37, 925. [Google Scholar] [CrossRef]
- Hu, G.L.; Peng, X.R.; Wang, X.; Li, X.; Li, X.; Qiu, M.H. Excavation of coffee maturity markers and further research on their changes in coffee cherries of different maturity. Food Res. Int. 2020, 132, 7. [Google Scholar] [CrossRef] [PubMed]
- Ardo, Y. Flavour formation by amino acid catabolism. Biotechnol. Adv. 2006, 24, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Morris, W.L.; Ross, H.A.; Ducreux, L.J.M.; Bradshaw, J.E.; Bryan, G.J.; Taylor, M.A. Umami compounds are a determinant of the flavor of potato (Solanum tuberosum L.). J. Agric. Food Chem. 2007, 55, 9627–9633. [Google Scholar] [CrossRef] [PubMed]
- Wu, F.N.; Tang, J.; Pei, F.; Wang, S.; Chen, G.T.; Hu, Q.H.; Zhao, L.Y. The influence of four drying methods on nonvolatile taste components of White Hypsizygus marmoreus. Eur. Food Res. Technol. 2015, 240, 823–830. [Google Scholar] [CrossRef]
- Liu, C.J.; Yang, N.; Yang, Q.; Ayed, C.; Linforth, R.; Fisk, I.D. Enhancing Robusta coffee aroma by modifying flavour precursors in the green coffee bean. Food Chem. 2019, 281, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Rogers, W.J.; Michaux, S.; Bastin, M.; Bucheli, P. Changes to the content of sugars, sugar alcohols, myo-inositol, carboxylic acids and inorganic anions in developing grains from different varieties of Robusta (Coffea canephora) and Arabica (C. arabica) coffees. Plant Sci. 1999, 149, 115–123. [Google Scholar] [CrossRef]
- Koshiro, Y.; Jackson, M.J.; Nagai, C.; Ashihara, H. Changes in the content of sugars and organic acids during ripening of coffea arabica and coffea canephora fruits. Eur. Chem. Bull. 2015, 4, 378–383. [Google Scholar]
- Chu, R.; Wan, L.S.; Peng, X.R.; Yu, M.Y.; Zhang, Z.R.; Zhou, L.; Li, Z.R.; Qiu, M.H. Characterization of New Ent-kaurane Diterpenoids of Yunnan Arabica Coffee Beans. Nat. Prod. Bioprospect. 2016, 6, 217–223. [Google Scholar] [CrossRef]
- Oh, S.H.; Hwang, Y.P.; Choi, J.H.; Jin, S.W.; Lee, G.H.; Han, E.H.; Chung, Y.H.; Chung, Y.C.; Jeong, H.G. Kahweol inhibits proliferation and induces apoptosis by suppressing fatty acid synthase in HER2-overexpressing cancer cells. Food Chem. Toxicol. 2018, 121, 326–335. [Google Scholar] [CrossRef] [PubMed]
- Iwamoto, H.; Izumi, K.; Natsagdorj, A.; Naito, R.; Makino, T.; Kadomoto, S.; Hiratsuka, K.; Shigehara, K.; Kadono, Y.; Narimoto, K.; et al. Coffee diterpenes kahweol acetate and cafestol synergistically inhibit the proliferation and migration of prostate cancer cells. Prostate 2019, 79, 12. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Wu, G.; Guo, L.; Hu, F.C.; Zhou, L.Y.; Xu, B.; Yin, Q.C.; Chen, Z. Metabolic Profiling and Potential Taste Biomarkers of Two Rambutans during Maturation. Molecules 2023, 28, 1390. [Google Scholar] [CrossRef] [PubMed]
- Setoyama, D.; Iwasa, K.; Seta, H.; Shimizu, H.; Fujimura, Y.; Miura, D.; Wariishi, H.; Nagai, C.; Nakahara, K. High-Throughput Metabolic Profiling of Diverse Green Coffea arabica Beans Identified Tryptophan as a Universal Discrimination Factor for Immature Beans. PLoS ONE 2013, 8, e70098. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bi, X.; Yu, H.; Hu, F.; Fu, X.; Li, Y.; Li, Y.; Yang, Y.; Liu, D.; Li, G.; Shi, R.; et al. A Systematic Analysis of the Correlation between Flavor Active Differential Metabolites and Multiple Bean Ripening Stages of Coffea arabica L. Molecules 2024, 29, 180. https://doi.org/10.3390/molecules29010180
Bi X, Yu H, Hu F, Fu X, Li Y, Li Y, Yang Y, Liu D, Li G, Shi R, et al. A Systematic Analysis of the Correlation between Flavor Active Differential Metabolites and Multiple Bean Ripening Stages of Coffea arabica L. Molecules. 2024; 29(1):180. https://doi.org/10.3390/molecules29010180
Chicago/Turabian StyleBi, Xiaofei, Haohao Yu, Faguang Hu, Xingfei Fu, Yanan Li, Yaqi Li, Yang Yang, Dexin Liu, Guiping Li, Rui Shi, and et al. 2024. "A Systematic Analysis of the Correlation between Flavor Active Differential Metabolites and Multiple Bean Ripening Stages of Coffea arabica L." Molecules 29, no. 1: 180. https://doi.org/10.3390/molecules29010180