
Anti-Obesity Activities of the Compounds from Perilla frutescens var. acuta and Chemical Profiling of the Extract

 , , and
, , and 
Abstract
1. Introduction
2. Results
2.1. Inhibitory Activity against Adipocyte Differentiation
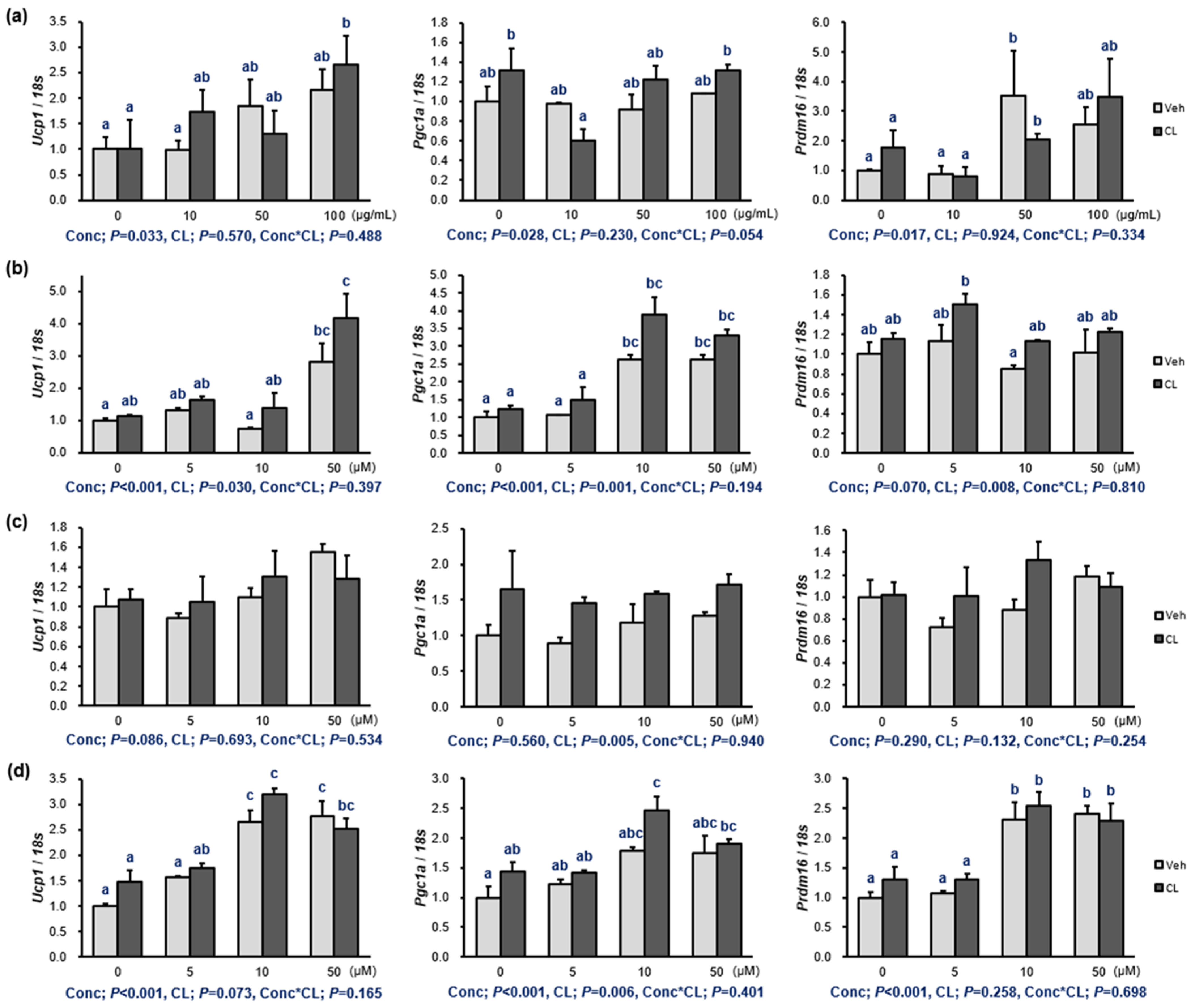
2.2. Thermogenesis of the Extract and 1–3
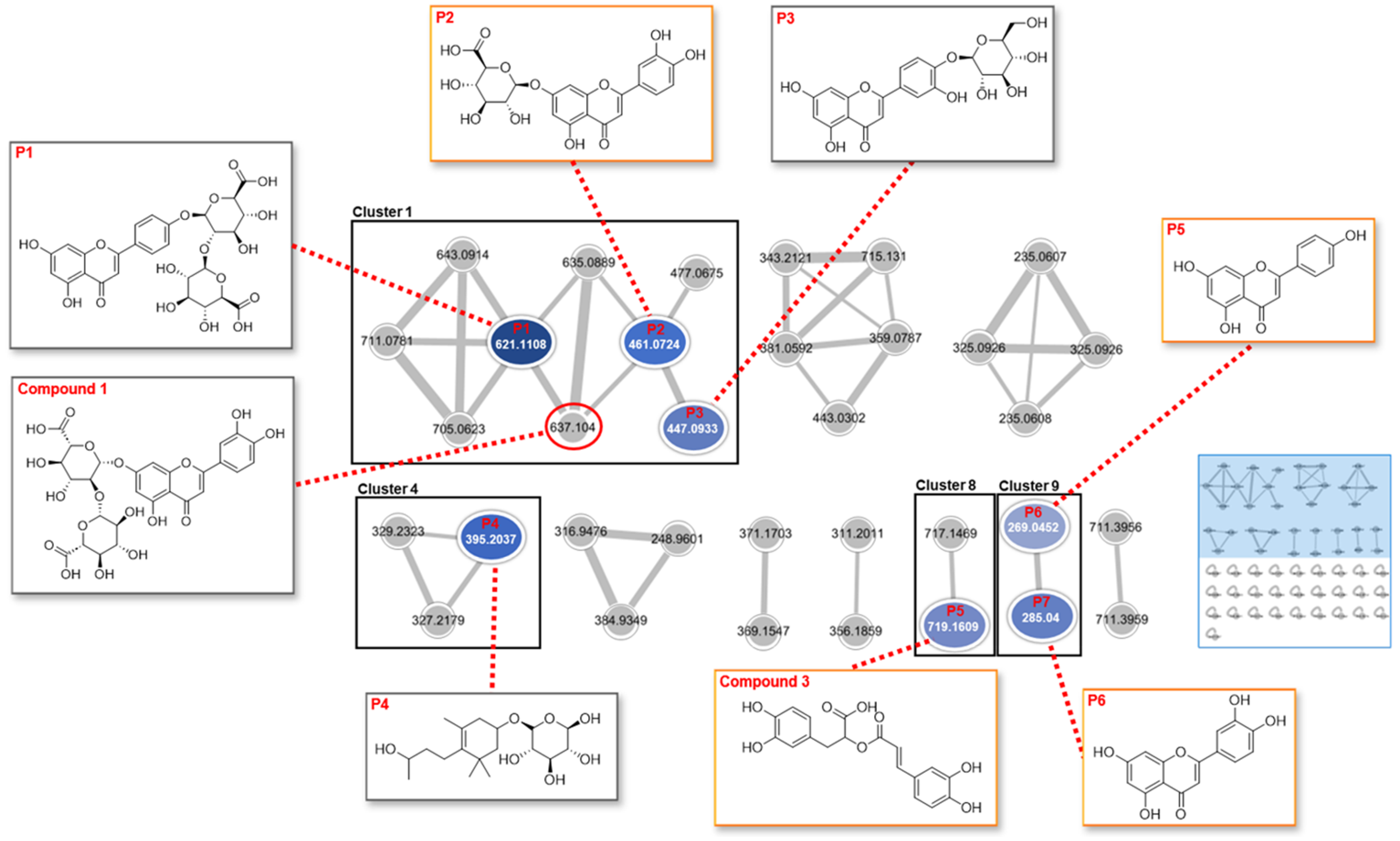
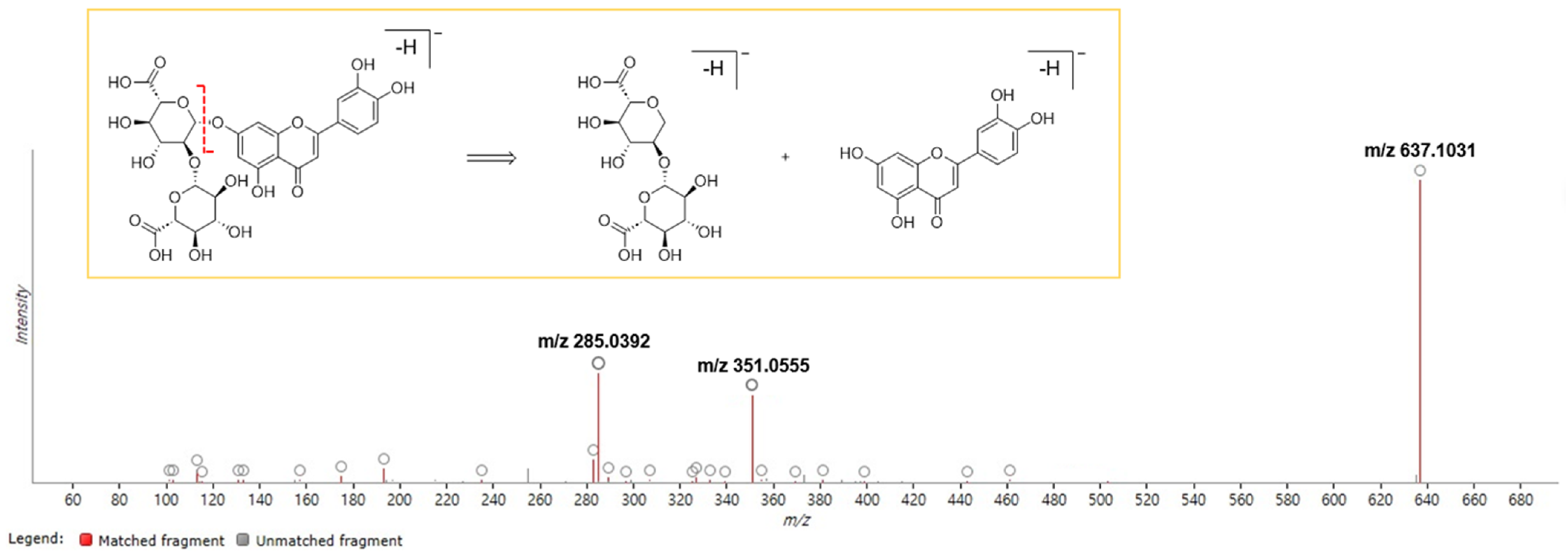
2.3. Feature-Based Molecular Networking of the Extract
3. Discussion and Conclusions
4. Materials and Methods
4.1. Plant Materials and Sample Preparation
4.2. Cell Culture
4.3. Cell Viability
4.4. Anti-Adipogenic and Thermogenic Gene Expression
4.5. Statistical Analysis
4.6. UPLC-MS/MS Analysis
4.6.1. UHPLC and MS/MS Conditions
4.6.2. Feature-Based Molecular Networking
4.6.3. Identification of Chemical Markers
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Blüher, M. Obesity: Global epidemiology and pathogenesis. Nat. Rev. Endocrinol. 2019, 15, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Müller, T.D.; Blüher, M.; Tschöp, M.H.; DiMarchi, R.D. Anti-obesity drug discovery: Advances and challenges. Nat. Rev. Drug Discov. 2022, 21, 201–223. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-N.; Kang, M.-C.; Kang, N.; Kim, S.-Y.; Hyun, C.-G.; Roh, S.W.; Ko, E.-Y.; Cho, K.; Jung, W.-K.; Ahn, G. 2,4,6-Trihydroxybenzaldehyde, a potential anti-obesity treatment, suppressed adipocyte differentiation in 3T3-L1 cells and fat accumulation induced by high-fat diet in C57BL/6 mice. Environ. Toxicol. Pharmacol. 2015, 39, 962–968. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Yang, M.; Han, Y.; Zhao, H.; Sun, L. PRDM16 regulating adipocyte transformation and thermogenesis: A promising therapeutic target for obesity and diabetes. Front. Pharmacol. 2022, 13, 870250. [Google Scholar] [CrossRef] [PubMed]
- Textbook Compilation Committee of Korean Pharmacognosy. Pharmacognosy, 2nd ed.; Dong-Myeung Press: Seoul, Republic of Korea, 2001; pp. 522–523. [Google Scholar]
- Wu, X.; Dong, S.; Chen, H.; Guo, M.; Sun, Z.; Luo, H. Perilla frutescens: A traditional medicine and food homologous plant. Chin. Herb. Med. 2023, 15, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Hou, T.; Netala, V.R.; Zhang, H.; Xing, Y.; Li, H.; Zhang, Z. Perilla frutescens: A rich source of pharmacological active compounds. Molecules 2022, 27, 3578. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.S.; Kim, M.; Lee, S.J.; Cha, Y.S. Antiobesity effects of purple perilla (Perilla frutescens var. acuta) on adipocyte differentiation and mice fed a high-fat diet. J. Food Sci. 2018, 83, 2384–2393. [Google Scholar] [CrossRef]
- Choi, J.-Y.; Kim, J.-W.; Kwon, E.-Y. Effects of the Perilla frutescens var. acuta Kudo ethanol extract (PFE) on the improvement of metabolic syndrome and non-alcoholic fatty liver disease induced by high-fat diet. J. Korean Soc. Food. Sci. Nutr. 2020, 49, 1045–1053. [Google Scholar] [CrossRef]
- Youn, I.; Han, S.; Jung, H.J.; Noh, S.G.; Chung, H.Y.; Koo, Y.K.; Shin, S.; Seo, E.K. Anti-inflammatory activity of the constituents from the leaves of Perilla frutescens var. acuta. Pharmaceuticals 2023, 16, 1655. [Google Scholar] [CrossRef]
- Otto, T.C.; Lane, M.D. Adipose development: From stem cell to adipocyte. Crit. Rev. Biochem. Mol. Biol. 2005, 40, 229–242. [Google Scholar] [CrossRef]
- Visioli, F.; Lastra, C.A.D.L.; Andres-Lacueva, C.; Aviram, M.; Calhau, C.; Cassano, A.; D’Archivio, M.; Faria, A.; Favé, G.; Fogliano, V. Polyphenols and human health: A prospectus. Crit. Rev. Food Sci. Nutr. 2011, 51, 524–546. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Pérez, C.; Segura-Carretero, A.; Contreras, M.d.M. Phenolic compounds as natural and multifunctional anti-obesity agents: A review. Crit. Rev. Food Sci. Nutr. 2019, 59, 1212–1229. [Google Scholar] [CrossRef]
- Ghaben, A.L.; Scherer, P.E. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Barak, Y.; Nelson, M.C.; Ong, E.S.; Jones, Y.Z.; Ruiz-Lozano, P.; Chien, K.R.; Koder, A.; Evans, R.M. PPAR-γ is required for placental, cardiac, and adipose tissue development. Mol. Cell 1999, 4, 585–595. [Google Scholar] [CrossRef]
- Rosen, E.D.; Sarraf, P.; Troy, A.E.; Bradwin, G.; Moore, K.; Milstone, D.S.; Spiegelman, B.M.; Mortensen, R.M. PPAR-γ is required for the differentiation of adipose tissue in vivo and in vitro. Mol. Cell 1999, 4, 611–617. [Google Scholar] [CrossRef] [PubMed]
- Lane, M.; Lin, F.; MacDougald, O.; Vasseur-Cognet, M. Control of adipocyte differentiation by CCAAT/enhancer binding protein alpha (C/EBP alpha). Int. J. Obes. Relat. Metab. Disord. 1996, 20, S91–S96. [Google Scholar] [PubMed]
- Rui, Y.; Tong, L.; Cheng, J.; Wang, G.; Qin, L.; Wan, Z. Rosmarinic acid suppresses adipogenesis, lipolysis in 3T3-L1 adipocytes, lipopolysaccharide-stimulated tumor necrosis factor-α secretion in macrophages, and inflammatory mediators in 3T3-L1 adipocytes. Food Nutr. Res. 2017, 61, 1330096. [Google Scholar] [CrossRef] [PubMed]
- Nothias, L.-F.; Petras, D.; Schmid, R.; Dührkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M. Feature-based molecular networking in the GNPS analysis environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef] [PubMed]
- Hildebrandt, X.; Ibrahim, M.; Peltzer, N. Cell death and inflammation during obesity: “Know my methods, WAT (son)”. Cell Death Differ. 2023, 30, 279–292. [Google Scholar] [CrossRef]
- Shin, S.; Ajuwon, K.M. Lipopolysaccharide alters thermogenic and inflammatory genes in white adipose tissue in mice fed diets with distinct 18-carbon fatty-acid composition. Lipids 2018, 53, 885–896. [Google Scholar] [CrossRef]
- Maffei, M.; Halaas, J.; Ravussin, E.; Pratley, R.; Lee, G.; Zhang, Y.; Fei, H.; Kim, S.; Lallone, R.; Ranganathan, S. Leptin levels in human and rodent: Measurement of plasma leptin and ob RNA in obese and weight-reduced subjects. Nat. Med. 1995, 1, 1155–1161. [Google Scholar] [CrossRef] [PubMed]
- Yamauchi, T.; Kamon, J.; Minokoshi, Y.; Ito, Y.; Waki, H.; Uchida, S.; Yamashita, S.; Noda, M.; Kita, S.; Ueki, K.; et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat. Med. 2002, 8, 1288–1295. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| tR (min) | m/z | Adduct | Cosine Score | Mass Error (ppm) | Compound Name | |
|---|---|---|---|---|---|---|
| 3 | 7.68 | 719.1609 | 2M-H | 0.86 | 1.53 | Rosmarinic acid |
| P1 | 6.55 | 621.1108 | M-H | 0.76 | 1.28 | Apigenin-4′-O-[(β-D-glucuronosyl-(1→2)-β-D-glucuronide] |
| P2 | 6.92 | 461.0724 | M-H | 0.85 | 1.32 | Luteolin-7-O-D-glucuronide |
| P3 | 6.46 | 447.0933 | M-H | 0.87 | 0.68 | Luteolin-4′-O-glucoside |
| P4 | 9.94 | 395.2037 | M+Na-2H | 0.71 | 9.34 | Tsangane L-3-glucoside |
| P5 | 9.39 | 269.0452 | M-H | 0.84 | 1.81 | Apigenin |
| P6 | 8.61 | 285.0400 | M-H | 0.88 | 0.00 | Luteolin |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Youn, I.; Piao, D.; Park, J.; Ock, S.A.; Han, S.; Han, A.-R.; Shin, S.; Seo, E.K. Anti-Obesity Activities of the Compounds from Perilla frutescens var. acuta and Chemical Profiling of the Extract. Molecules 2024, 29, 2465. https://doi.org/10.3390/molecules29112465
Youn I, Piao D, Park J, Ock SA, Han S, Han A-R, Shin S, Seo EK. Anti-Obesity Activities of the Compounds from Perilla frutescens var. acuta and Chemical Profiling of the Extract. Molecules. 2024; 29(11):2465. https://doi.org/10.3390/molecules29112465
Chicago/Turabian StyleYoun, Isoo, Donglan Piao, Jisu Park, Seung A Ock, Sujin Han, Ah-Reum Han, Sunhye Shin, and Eun Kyoung Seo. 2024. "Anti-Obesity Activities of the Compounds from Perilla frutescens var. acuta and Chemical Profiling of the Extract" Molecules 29, no. 11: 2465. https://doi.org/10.3390/molecules29112465
APA StyleYoun, I., Piao, D., Park, J., Ock, S. A., Han, S., Han, A.-R., Shin, S., & Seo, E. K. (2024). Anti-Obesity Activities of the Compounds from Perilla frutescens var. acuta and Chemical Profiling of the Extract. Molecules, 29(11), 2465. https://doi.org/10.3390/molecules29112465